Protein vázající TATA - TATA-binding protein




| TBP | |||||||||
|---|---|---|---|---|---|---|---|---|---|
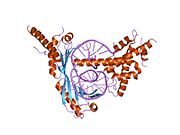
 krystalová struktura kvasinkového ternárního komplexu brf1-tbp-dna | |||||||||
| Identifikátory | |||||||||
| Symbol | TBP | ||||||||
| Pfam | PF00352 | ||||||||
| Pfam klan | CL0407 | ||||||||
| InterPro | IPR000814 | ||||||||
| STRÁNKA | PDOC00303 | ||||||||
| SCOP2 | 1 TB / Rozsah / SUPFAM | ||||||||
| |||||||||
The Protein vázající TATA (TBP) je obecný transkripční faktor který se váže konkrétně na a DNA sekvence zvaná TATA box. Tato sekvence DNA je asi 30 základní páry proti proudu od místo zahájení transkripce v některých eukaryotický gen promotéři.[4]
TBP genová rodina
TBP je členem malé genové rodiny faktorů souvisejících s TBP.[5] U ovocné mušky byl identifikován první faktor související s TBP (TRF / TRF1) Drosophila, ale zdá se, že je specifický pro mouchy nebo hmyz. Následně TBPL1 / TRF2 byl nalezen v genomech mnoha lidí metazoans, zatímco obratlovců genomy kódují třetího člena rodiny obratlovců, TBPL2 / TRF3. Ve specifických typech buněk nebo na specifických promotorech může být TBP nahrazen jedním z těchto faktorů souvisejících s TBP, z nichž některé interagují s TATA boxem podobně jako TBP.
Role jako transkripční faktor
TBP je podjednotka eukaryotické obecný transkripční faktor TFIID. TFIID je první protein, který se váže na DNA během tvorby transkripční preiniciační komplex z RNA polymeráza II (RNA Pol II).[6] Jako jeden z mála proteinů v preiniciačním komplexu, který váže DNA sekvenčně specifickým způsobem, pomáhá umístit RNA polymerázu II nad místo zahájení transkripce genu. Odhaduje se však, že pouze 10–20% lidských promotorů má TATA boxy. TBP proto pravděpodobně není jediným proteinem podílejícím se na umístění RNA polymerázy II. Většina lidských promotorů je TATA -méně úklidový gen promotéři. Vazba TBP na tyto promotory je usnadněna regulátory genů pro domácnost.[7][8] Zajímavé je, že transkripce začíná v úzké oblasti kolem 30 bp za TATA boxem na promotorech obsahujících TATA,[9] zatímco počáteční místa transkripce promotorů bez TATA jsou dispergována v oblasti 200 bp.[10][8]
Vazba TFIID na TATA box v promotér oblast genu iniciuje nábor dalších faktorů potřebných pro RNA Pol II k zahájení transkripce. Mezi další přijaté transkripční faktory patří TFIIA, TFIIB, a TFIIF. Každý z těchto transkripčních faktorů obsahuje několik proteinových podjednotek.
TBP je také důležité pro přepis do RNA polymeráza I a RNA polymeráza III, a proto se účastní iniciace transkripce všemi třemi RNA polymerázami.[11]
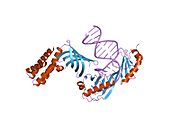
TBP je zapojena do Tání DNA (dvouvláknová separace) ohnutím DNA o 80 ° (sekvence bohatá na AT, na kterou se váže, usnadňuje snadné tavení). TBP je neobvyklý protein v tom, že váže vedlejší drážku pomocí β listu.
Dalším charakteristickým rysem TBP je dlouhý řetězec glutaminů na N-konci proteinu. Tato oblast moduluje aktivitu vazby DNA na C-konci a modulace vazby DNA ovlivňuje rychlost tvorby transkripčního komplexu a zahájení transkripce. Mutace, které rozšiřují počet opakování CAG kódujících toto polyglutaminový trakt, a tím zvyšují délku polyglutaminového řetězce, jsou spojeny s spinocerebelární ataxie 17, a neurodegenerativní porucha klasifikováno jako a polyglutaminová nemoc.[12]
Interakce DNA-protein
Když se TBP váže na a TATA box v rámci DNA, narušuje DNA vložením postranních řetězců aminokyselin mezi páry bází, částečným odvíjením šroubovice a dvojnásobným zauzlováním. Zkreslení je dosaženo velkým množstvím povrchového kontaktu mezi proteinem a DNA. TBP se váže na záporně nabité fosfáty v páteři DNA prostřednictvím kladně nabitých lysin a arginin aminokyselinové zbytky. Ostrý ohyb v DNA je vytvořen projekcí čtyř objemných fenylalanin zbytky do vedlejší drážky. Jak se DNA ohýbá, zvyšuje se její kontakt s TBP, což zvyšuje interakci DNA-protein.
Kmen uložený na DNA touto interakcí iniciuje tání nebo separaci řetězců. Protože tato oblast DNA je bohatá na adenin a tymin zbytky, které párují bazy pouze přes dva Vodíkové vazby, řetězce DNA se snáze oddělují. Oddělení dvou vláken odhaluje základy a umožňuje RNA polymeráza II zahájit přepis souboru gen.
C-konec TBP se skládá ze spirálovitého tvaru, který (neúplně) doplňuje T-A-T-A oblast DNA. Tato neúplnost umožňuje DNA pasivně ohýbat vazbu.
Informace o použití TBP v buňkách viz: RNA polymeráza I, RNA polymeráza II, a RNA polymeráza III.
Interakce protein-protein
Ukázalo se, že protein vázající TATA komunikovat s:
- BRF1,[13][14]
- BTAF1,[15][16]
- C-Fos,[17]
- C-jun,[18]
- EDF1,[19][20][21]
- GTF2B (TFIIB),[22][23]
- GTF2A1 (TFIIA podjednotka 1),[22][24][25][26]
- GTF2F1 (TFIIF podjednotka 1)[15][27][28]
- GTF2H4 (TFIIH podjednotka 4),[15]
- Mdm2,[29][30]
- MSX1,[31][32][33]
- NFYB,[34]
- P53,[35][36]
- PAX6,[36]
- POLR2A,[15]
- POU2F1,[37]
- RELA,[38][39]
- NR2B1,[40]
- TAF1,[34][41][42][43]
- TAF4,[44]
- TAF5,[34][42][45]
- TAF6,[34][42][44]
- TAF7,[34][44]
- TAF9.[34][46]
- TAF10,[34][42]
- TAF11,[34][47][48]
- TAF13,[47] a
- TAF15.[49]
Složitá montáž
TATA box vazba protein (TBP) je vyžadován pro zahájení transkripce RNA polymerázy I, II a III, od promotéři s nebo bez TATA boxu.[50][51] V přítomnosti promotoru bez TATA se TBP váže pomocí faktorů spojených s TBP (TAF).[52][53] TBP se sdružuje s řadou faktorů, včetně obecných transkripční faktory TFIIA, -B, -D, -E a -H k vytvoření obrovských předinicializačních komplexů s více podjednotkami na jádru promotér. Prostřednictvím jeho spojení s různými transkripce faktory, může TBP iniciovat transkripce z různých RNA polymerázy. Existuje několik souvisejících TBP, včetně TBP-like (TBPL) bílkoviny.[54]
Struktura
C-terminální jádro TBP (~ 180 zbytků) je vysoce konzervativní a obsahuje dvě 88-aminokyseliny opakuje které vytvářejí sedlový tvar struktura který se rozkročí nad DNA; v tomto regionu váže do pole TATA a interaguje s transkripční faktory a regulační bílkoviny .[55] Naproti tomu N-koncová oblast se liší jak v délce, tak v sekvence.
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000112592 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Kornberg RD (2007). „Molekulární základ eukaryotické transkripce“. Proc. Natl. Acad. Sci. USA. 104 (32): 12955–61. Bibcode:2007PNAS..10412955K. doi:10.1073 / pnas.0704138104. PMC 1941834. PMID 17670940.
- ^ Akhtar W, Veenstra GJ (1. ledna 2011). „Faktory související s TBP: paradigma rozmanitosti v iniciaci transkripce“. Cell & Bioscience. 1 (1): 23. doi:10.1186/2045-3701-1-23. PMC 3142196. PMID 21711503.
- ^ Lee TI, Young RA (2000). "Transkripce genů kódujících eukaryotický protein". Výroční přehled genetiky. 34: 77–137. doi:10.1146 / annurev.genet.34.1.77. PMID 11092823.
- ^ Lam KC, Mühlpfordt F, Vaquerizas JM, Raja SJ, Holz H, Luscombe NM, Manke T, Akhtar A (2012). „Komplex NSL reguluje úklidové geny v Drosophile“. Genetika PLOS. 8 (6): e1002736. doi:10.1371 / journal.pgen.1002736. PMC 3375229. PMID 22723752.
- ^ A b Lam KC, Chung HR, Semplicio G, Iyer SS, Gaub A, Bhardwaj V, Holz H, Georgiev P, Akhtar A (únor 2019). „Pro zachování věrnosti transkripce a potlačení transkripčního šumu je nutná krajina nukleosomů zprostředkovaná komplexem NSL“. Geny a vývoj. 33 (7–8): 452–465. doi:10,1101 / gad.321489.118. PMC 6446542. PMID 30819819.
- ^ Carninci P, Sandelin A, Lenhard B, Katayama S, Shimokawa K, Ponjavic J a kol. (Červen 2006). „Analýza genomu a vývoje evoluce savčích promotorů v celém genomu“. Genetika přírody. 38 (6): 626–35. doi:10.1038 / ng1789. PMID 16645617. S2CID 22205897.
- ^ Ni T, Corcoran DL, Rach EA, Song S, Spana EP, Gao Y, Ohler U, Zhu J (červenec 2010). „Strategie párování na konci mapující komplexní prostředí iniciace transkripce“. Přírodní metody. 7 (7): 521–7. doi:10.1038 / nmeth.1464. PMC 3197272. PMID 20495556.
- ^ Vannini A, Cramer P (únor 2012). „Zachování mezi zařízeními pro iniciaci transkripce RNA polymerázy I, II a III“. Molekulární buňka. 45 (4): 439–46. doi:10.1016 / j.molcel.2012.01.023. PMID 22365827.
- ^ „Entrez Gene: TBP TATA box binding protein“.
- ^ McCulloch V, Hardin P, Peng W, Ruppert JM, Lobo-Ruppert SM (srpen 2000). „Alternativně sestříhané varianty hBRF fungují na různých promotorech RNA polymerázy III“. EMBO J.. 19 (15): 4134–43. doi:10.1093 / emboj / 19.15.4134. PMC 306597. PMID 10921893.
- ^ Wang Z, Roeder RG (červenec 1995). „Struktura a funkce podjednotky lidského transkripčního faktoru TFIIIB, která je evolučně konzervovaná a obsahuje jak domény související s proteinem TFIIB, tak s proteinem s vysokou mobilitou 2“. Proc. Natl. Acad. Sci. USA. 92 (15): 7026–30. Bibcode:1995PNAS ... 92,7026W. doi:10.1073 / pnas.92.15.7026. PMC 41464. PMID 7624363.
- ^ A b C d Scully R, Anderson SF, Chao DM, Wei W, Ye L, Young RA, Livingston DM, Parvin JD (květen 1997). „BRCA1 je složkou holoenzymu RNA polymerázy II“. Proc. Natl. Acad. Sci. USA. 94 (11): 5605–10. Bibcode:1997PNAS ... 94.5605S. doi:10.1073 / pnas.94.11.5605. PMC 20825. PMID 9159119.
- ^ Chicca JJ, Auble DT, Pugh BF (březen 1998). „Klonování a biochemická charakterizace TAF-172, lidského homologu kvasinek Mot1“. Mol. Buňka. Biol. 18 (3): 1701–10. doi:10.1128 / MCB.18.3.1701. PMC 108885. PMID 9488487.
- ^ Metz R, Bannister AJ, Sutherland JA, Hagemeier C, O'Rourke EC, Cook A, Bravo R, Kouzarides T (září 1994). „c-Fos indukovaná aktivace promotoru obsahujícího TATA box zahrnuje přímý kontakt s proteinem vázajícím TATA“. Mol. Buňka. Biol. 14 (9): 6021–9. doi:10.1128 / MCB.14.9.6021. PMC 359128. PMID 8065335.
- ^ Franklin CC, McCulloch AV, Kraft AS (únor 1995). „In vitro asociace mezi rodinou proteinů Jun a obecnými transkripčními faktory, TBP a TFIIB“. Biochem. J. 305 (3): 967–74. doi:10.1042 / bj3050967. PMC 1136352. PMID 7848298.
- ^ Brendel C, Gelman L, Auwerx J (červen 2002). „Multiproteinový můstkový faktor-1 (MBF-1) je kofaktorem pro jaderné receptory, které regulují metabolismus lipidů“. Mol. Endokrinol. 16 (6): 1367–77. doi:10.1210 / oprava.16.6.0843. PMID 12040021.
- ^ Mariotti M, De Benedictis L, Avon E, Maier JA (srpen 2000). „Interakce mezi faktorem 1 souvisejícím s endoteliální diferenciací a kalmodulinem in vitro a in vivo“. J. Biol. Chem. 275 (31): 24047–51. doi:10,1074 / jbc.M001928200. PMID 10816571.
- ^ Kabe Y, Goto M, Shima D, Imai T, Wada T, Morohashi Ki, Shirakawa M, Hirose S, Handa H (listopad 1999). „Role lidského MBF1 jako transkripčního koaktivátoru“. J. Biol. Chem. 274 (48): 34196–202. doi:10.1074 / jbc.274.48.34196. PMID 10567391.
- ^ A b Tang H, Sun X, Reinberg D, Ebright RH (Únor 1996). „Interakce protein-protein při iniciaci eukaryotické transkripce: struktura preiniciačního komplexu“. Proc. Natl. Acad. Sci. USA. 93 (3): 1119–24. Bibcode:1996PNAS ... 93.1119T. doi:10.1073 / pnas.93.3.1119. PMC 40041. PMID 8577725.CS1 maint: používá parametr autoři (odkaz)
- ^ Bushnell DA, Westover KD, Davis RE, Kornberg RD (únor 2004). "Strukturální základ transkripce: RNA polymeráza II-TFIIB kokrystal při 4,5 Angstromu". Věda. 303 (5660): 983–8. Bibcode:2004Sci ... 303..983B. doi:10.1126 / science.1090838. PMID 14963322. S2CID 36598301.
- ^ DeJong J, Bernstein R, Roeder RG (duben 1995). "Lidský obecný transkripční faktor TFIIA: charakterizace cDNA kódující malou podjednotku a požadavek na bazální a aktivovanou transkripci". Proc. Natl. Acad. Sci. USA. 92 (8): 3313–7. Bibcode:1995PNAS ... 92,3313D. doi:10.1073 / pnas.92.8.3313. PMC 42156. PMID 7724559.
- ^ Ozer J, Mitsouras K, Zerby D, Carey M, Lieberman PM (červen 1998). „Transkripční faktor IIA dereprimuje inhibici faktoru souvisejícího s proteinem vázajícím se na TATA (TBP) vazby TBP-DNA“. J. Biol. Chem. 273 (23): 14293–300. doi:10.1074 / jbc.273.23.14293. PMID 9603936.
- ^ Sun X, Ma D, Sheldon M, Yeung K, Reinberg D (říjen 1994). „Rekonstituce lidské aktivity TFIIA z rekombinantních polypeptidů: role v transkripci zprostředkované TFIID“. Genes Dev. 8 (19): 2336–48. doi:10,1101 / gad.8.19.2336. PMID 7958900.
- ^ Ruppert S, Tjian R (listopad 1995). „Lidský TAFII250 interaguje s RAP74: důsledky pro iniciaci RNA polymerázy II“. Genes Dev. 9 (22): 2747–55. doi:10,1101 / gad. 9.22.2747. PMID 7590250.
- ^ Malik S, Guermah M, Roeder RG (březen 1998). „Dynamický model pro funkci koaktivátoru PC4 v transkripci RNA polymerázy II“. Proc. Natl. Acad. Sci. USA. 95 (5): 2192–7. Bibcode:1998PNAS ... 95,2192M. doi:10.1073 / pnas.95.5.2192. PMC 19292. PMID 9482861.
- ^ Thut CJ, Goodrich JA, Tjian R (srpen 1997). „Represe transkripce zprostředkované p53 pomocí MDM2: duální mechanismus“. Genes Dev. 11 (15): 1974–86. doi:10.1101 / gad.11.15.1974. PMC 316412. PMID 9271120.
- ^ Léveillard T, Wasylyk B (prosinec 1997). „C-koncová oblast MDM2 se váže na TAFII250 a je nutná pro regulaci promotoru cyklinu A pomocí MDM2“. J. Biol. Chem. 272 (49): 30651–61. doi:10.1074 / jbc.272.49.30651. PMID 9388200.
- ^ Shetty S, Takahashi T, Matsui H, Ayengar R, Raghow R (květen 1999). „Transkripční autoreprese genu Msx1 je zprostředkována interakcemi proteinu Msx1 s multiproteinovým transkripčním komplexem obsahujícím protein vázající TATA, Sp1 a protein vázající protein cAMP-response-element-binding protein (CBP / p300)“. Biochem. J. 339 (3): 751–8. doi:10.1042/0264-6021:3390751. PMC 1220213. PMID 10215616.
- ^ Zhang H, Hu G, Wang H, Sciavolino P, Iler N, Shen MM, Abate-Shen C (květen 1997). „Heterodimerizace homeoproteinů Msx a Dlx vede k funkčnímu antagonismu“. Mol. Buňka. Biol. 17 (5): 2920–32. doi:10,1128 / mcb.17.5.2920. PMC 232144. PMID 9111364.
- ^ Zhang H, Catron KM, Abate-Shen C (březen 1996). „Role pro homeodoménu Msx-1 v regulaci transkripce: zbytky v N-koncovém rameni zprostředkovávají interakci vazebného proteinu TATA a transkripční represi.“. Proc. Natl. Acad. Sci. USA. 93 (5): 1764–9. Bibcode:1996PNAS ... 93.1764Z. doi:10.1073 / pnas.93.5.1764. PMC 39855. PMID 8700832.
- ^ A b C d E F G h Bellorini M, Lee DK, Dantonel JC, Zemzoumi K, Roeder RG, Tora L, Mantovani R (červen 1997). „Interakce NF-Y-TBP vázající se na CCAAT: NF-YB a NF-YC vyžadují krátké domény sousedící s jejich histonovými složenými motivy pro spojení s bazickými zbytky TBP“. Nucleic Acids Res. 25 (11): 2174–81. doi:10.1093 / nar / 25.11.2174. PMC 146709. PMID 9153318.
- ^ Seto E, Usheva A, Zambetti GP, Momand J, Horikoshi N, Weinmann R, Levine AJ, Shenk T (prosinec 1992). „Divoký typ p53 se váže na protein vázající TATA a potlačuje transkripci“. Proc. Natl. Acad. Sci. USA. 89 (24): 12028–32. Bibcode:1992PNAS ... 8912028S. doi:10.1073 / pnas.89.24.12028. PMC 50691. PMID 1465435.
- ^ A b Cvekl A, Kashanchi F, Brady JN, Piatigorsky J (červen 1999). „Interakce Pax-6 s proteinem vázajícím TATA a protein retinoblastomu“. Investovat. Oftalmol. Vis. Sci. 40 (7): 1343–50. PMID 10359315.
- ^ Zwilling S, Annweiler A, Wirth T (květen 1994). „Domény POU transkripčních faktorů Oct1 a Oct2 zprostředkovávají specifickou interakci s TBP“. Nucleic Acids Res. 22 (9): 1655–62. doi:10.1093 / nar / 22.9.1655. PMC 308045. PMID 8202368.
- ^ Guermah M, Malik S, Roeder RG (červen 1998). „Zapojení TFIID a USA komponent do transkripční aktivace promotoru viru lidské imunodeficience pomocí NF-kappaB a Sp1“. Mol. Buňka. Biol. 18 (6): 3234–44. doi:10,1128 / mcb.18.6.3234. PMC 108905. PMID 9584164.
- ^ Schmitz ML, Stelzer G, Altmann H, Meisterernst M, Baeuerle PA (březen 1995). „Interakce COOH-terminální transaktivační domény p65 NF-kappa B s proteinem vázajícím TATA, transkripčním faktorem IIB a koaktivátory“. J. Biol. Chem. 270 (13): 7219–26. doi:10.1074 / jbc.270.13.7219. PMID 7706261.
- ^ Schulman IG, Chakravarti D, Juguilon H, Romo A, Evans RM (srpen 1995). „Interakce mezi receptorem retinoidu X a konzervovanou oblastí proteinu vázajícího TATA zprostředkovávají transaktivaci závislou na hormonech“. Proc. Natl. Acad. Sci. USA. 92 (18): 8288–92. Bibcode:1995PNAS ... 92.8288S. doi:10.1073 / pnas.92.18.8288. PMC 41142. PMID 7667283.
- ^ Siegert JL, Robbins PD (leden 1999). „Rb inhibuje aktivitu vnitřní kinázy faktoru TAFII250 spojeného s proteinem spojujícím TATA“. Mol. Buňka. Biol. 19 (1): 846–54. doi:10.1128 / MCB.19.1.846. PMC 83941. PMID 9858607.
- ^ A b C d Ruppert S, Wang EH, Tjian R (březen 1993). „Klonování a exprese lidského TAFII250: faktor spojený s TBP podílející se na regulaci buněčného cyklu“. Příroda. 362 (6416): 175–9. Bibcode:1993 Natur.362..175R. doi:10.1038 / 362175a0. PMID 7680771. S2CID 4364676.
- ^ O'Brien T, Tjian R (květen 1998). "Funkční analýza lidské TAFII250 N-terminální kinázové domény". Mol. Buňka. 1 (6): 905–11. doi:10.1016 / S1097-2765 (00) 80089-1. PMID 9660973.
- ^ A b C Pointud JC, Mengus G, Brancorsini S, Monako L, Parvinen M, Sassone-Corsi P, Davidson I (květen 2003). „Intracelulární lokalizace TAF7L, paralog transkripčního faktoru TFIID podjednotky TAF7, je vývojově regulována během diferenciace mužských zárodečných buněk“. J. Cell Sci. 116 (Pt 9): 1847–1858. doi:10.1242 / jcs.00391. PMID 12665565.
- ^ Tao Y, Guermah M, Martinez E, Oelgeschläger T, Hasegawa S, Takada R, Yamamoto T, Horikoshi M, Roeder RG (březen 1997). „Specifické interakce a potenciální funkce lidského TAFII100“. J. Biol. Chem. 272 (10): 6714–21. doi:10.1074 / jbc.272.10.6714. PMID 9045704.
- ^ Martinez E, Palhan VB, Tjernberg A, Lymar ES, Gamper AM, Kundu TK, Chait BT, Roeder RG (říjen 2001). „Lidský komplex STAGA je transkripční koaktivátor acetylující chromatin, který interaguje s sestřihem pre-mRNA a faktory vázajícími poškození DNA in vivo“. Mol. Buňka. Biol. 21 (20): 6782–95. doi:10.1128 / MCB.21.20.6782-6795.2001. PMC 99856. PMID 11564863.
- ^ A b Mengus G, květen M, Jacq X, Staub A, Tora L, Chambon P, Davidson I (duben 1995). "Klonování a charakterizace hTAFII18, hTAFII20 a hTAFII28: tři podjednotky lidského transkripčního faktoru TFIID". EMBO J.. 14 (7): 1520–31. doi:10.1002 / j.1460-2075.1995.tb07138.x. PMC 398239. PMID 7729427.
- ^ May M, Mengus G, Lavigne AC, Chambon P, Davidson I (červen 1996). „Lidský TAF (II28) podporuje transkripční stimulaci aktivační funkcí 2 retinoidních X receptorů“. EMBO J.. 15 (12): 3093–104. doi:10.1002 / j.1460-2075.1996.tb00672.x. PMC 450252. PMID 8670810.
- ^ Hoffmann A, Roeder RG (červenec 1996). „Klonování a charakterizace lidského TAF20 / 15. Několik interakcí naznačuje ústřední roli při tvorbě komplexu TFIID“. J. Biol. Chem. 271 (30): 18194–202. doi:10.1074 / jbc.271.30.18194. PMID 8663456.
- ^ Hochheimer A, Tjian R (červen 2003). „Diverzifikované komplexy iniciace transkripce rozšiřují selektivitu promotoru a tkáňově specifickou genovou expresi“. Geny a vývoj. 17 (11): 1309–20. doi:10.1101 / gad.1099903. PMID 12782648.
- ^ Pugh BF (září 2000). "Řízení genové exprese regulací proteinu vázajícího TATA". Gen. 255 (1): 1–14. doi:10.1016 / s0378-1119 (00) 00288-2. PMID 10974559.
- ^ 1942-, Weaver, Robert Franklin (1. ledna 2012). Molekulární biologie. McGraw-Hill. ISBN 9780073525327. OCLC 789601172.CS1 maint: číselné názvy: seznam autorů (odkaz)
- ^ Louder RK, He Y, López-Blanco JR, Fang J, Chacón P, Nogales E (březen 2016). "Struktura promotorem vázaného TFIID a model sestavení lidského preinicializačního komplexu". Příroda. 531 (7596): 604–9. Bibcode:2016Natur.531..604L. doi:10.1038 / příroda17394. PMC 4856295. PMID 27007846.
- ^ Davidson I (červenec 2003). „Genetika TBP a faktory související s TBP“. Trendy v biochemických vědách. 28 (7): 391–8. doi:10.1016 / S0968-0004 (03) 00117-8. PMID 12878007.
- ^ Nikolov DB, Hu SH, Lin J, Gasch A, Hoffmann A, Horikoshi M, Chua NH, Roeder RG, Burley SK (listopad 1992). "Krystalová struktura proteinu vázajícího TFIID TATA-box". Příroda. 360 (6399): 40–6. Bibcode:1992 Natur.360 ... 40N. doi:10.1038 / 360040a0. PMID 1436073. S2CID 4307043.
externí odkazy
- GeneReviews / NCBI / NIH / UW vstup na spinocerebelární ataxii typu 17
- Interaktivní struktura TBP na massey.ac.nz
- PDB Molekula měsíce Protein vázající TATA
- TATA-Box + Binding + Protein v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- FactorBook TBP
- Přehled všech strukturálních informací dostupných v PDB pro UniProt: P20226 (Protein vázající lidský TATA-box) na PDBe-KB.
Galerie PDB | |
|---|---|
|