FOXO1 - FOXO1 - Wikipedia






Forkhead box protein O1 (FOXO1) také známý jako vidlice v rhabdomyosarkomu (FKHR) je a protein že u lidí je kódován FOXO1 gen.[5] FOXO1 je transkripční faktor který hraje důležitou roli při regulaci glukoneogeneze a glykogenolýza podle inzulínová signalizace, a je také ústředním bodem rozhodnutí předběžnéadipocyt zavázat se k adipogeneze.[6] Je primárně regulováno prostřednictvím fosforylace na více zbytcích; své transkripční aktivita závisí na stavu fosforylace.[7][8]
Funkce
Adipogeneze

FOXO1 negativně reguluje adipogeneze.[9] Přesný mechanismus, kterým je toho dosaženo, není v současné době zcela objasněn. V aktuálně přijímaném modelu FOXO1 negativně reguluje adipogenezi vazbou na promotorová místa PPARG a zabránění jeho transkripci. K zahájení adipogeneze je zapotřebí stoupající hladiny PPARG; prevencí jeho transkripce FOXO1 brání nástupu adipogeneze. Během stimulace inzulinem je FOXO1 vyloučen z jádra a následně není schopen zabránit transkripci PPARG a inhibovat adipogenezi.[10] Existují však podstatné důkazy, které naznačují, že existují další faktory, které zprostředkovávají interakci mezi FOX01 a promotorem PPARG, a že inhibice adipogeneze není zcela závislá na FOXO1 zabraňujícím transkripci PPARG.[11] Neschopnost zavázat se k adipogenezi je primárně způsobena aktivním FOXO1 zastavujícím buňku v G0 / G1 aktivací dosud neznámých následných cílů, přičemž předpokládaný cíl je SOD2.[12]
FOXO1 patří do vidlice rodina transkripční faktory které se vyznačují výrazným doména vidlicové hlavy. Specifická funkce tohoto genu dosud nebyla stanovena; Může však hrát roli v myogenní růst a diferenciace.[13] FOXO1 je nezbytný pro udržení lidské pluripotence ESC. Tato funkce je pravděpodobně zprostředkována přímou kontrolou exprese genu OCT4 a SOX2 prostřednictvím FOX01 prostřednictvím obsazení a aktivace příslušných promotorů.[14] V jaterních buňkách se zdá, že tento transkripční faktor zvyšuje expresi PEPCK a glykogen-6-fosfatáza (stejné enzymy, které jsou blokovány prostřednictvím metformin /AMPK /SHP cesta). Blokování tohoto transkripčního faktoru nabízí příležitost pro nové terapie pro diabetes mellitus.[15] V pankreatických alfa-buňkách je FOXO1 důležitý při regulaci prepro-glukagon výraz.[16] V pankreatických beta buňkách zprostředkovává FOXO1 glukagonu podobný peptid-1 účinky na hmotnost beta-buněk pankreatu.[17]
Glukoneogeneze a glykogenolýza
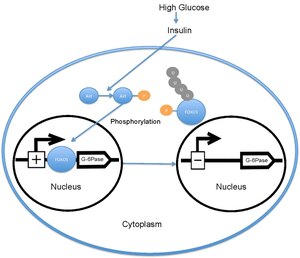
Když je hladina glukózy v krvi vysoká, pankreas se uvolní inzulín do krevního řečiště. Inzulin pak způsobí aktivaci PI3K, který následně fosforyluje Akt. Akt poté fosforyluje FOXO1, což způsobí jaderné vyloučení.[18][19] Tento fosforylovaný FOXO1 je pak ubikvitinován a degradován proteosomem.[20] Fosforylace FOX01 je nevratná; to prodlužuje inhibiční účinek inzulínu na metabolismus glukózy a produkci glukózy v játrech. Přepis glukóza 6-fosfatáza následně klesá, což následně snižuje sazby glukoneogeneze a glykogenolýza.[21] FOXO1 také aktivuje transkripci fosfoenolpyruvátkarboxykináza, který je nutný pro glukoneogenezi.[22] Činnost FOXO1 je také regulována prostřednictvím CBP indukovaná acetylace[23] na Lys -242, Lys-245 a Lys-262. Tyto lysinové zbytky jsou umístěny uvnitř Doména vázající DNA; acetylace inhibuje schopnost FOXO1 interagovat s promotorem glukóza-6 fosfatázy snížením stability komplexu FOXO1-DNA. Navíc tato acetylace zvyšuje rychlost fosforylace na Ser-253 o Akt. Mutace Ser-253 na Ala-253 činí FOXO1 konstitutivně aktivní. SIRT1 obrátí tento proces acetylace; přesný mechanismus, kterým SIRT1 deacetyluje FOXO1, je však stále předmětem šetření; v současné době se předpokládá, že acetylace zmírňuje transkripční aktivitu FOXO1 a poskytuje tak další úroveň metabolické regulace, která je nezávislá na cestě inzulín / PI3K.[24]
Apoptóza
FOXO1 může hrát důležitou roli v apoptóza protože je fosforylován a inhibován AKT.[25] Když je FOXO1 nadměrně vyjádřen v člověku LNCaP prostata rakovinné buňky, to způsobuje apoptóza.[25] FOXO1 také reguluje ligand indukující apoptózu související s TNF (STEZKA ), který způsobil FOXO1 apoptóza v člověku rakovina prostaty buněčná linie LAPC4 když byla použita nadměrná exprese zprostředkovaná FOXO1 adenovirem.[25] FOXO1 zvyšuje hladinu ligandu Fas (FasL ) transkripčně vedoucí k apoptotická buněčná smrt.[25] FOXO1 navíc transaktivuje Bim protein, který je členem Rodina Bcl-2 který podporuje apoptóza a hraje roli ve vnitřní mitochondriální apoptotické dráze.[25] Dále se ukázalo, že DNA poškození buněk vyvolané poškozením p53 - nedostatečné a p53 -proficientních buněk bylo sníženo, když byl lidský FOXO1 umlčen siRNA.[25] v Diabetes typu 2 the beta buňky slinivky břišní, které se normálně produkují inzulín podstoupit apoptózu, což výrazně snižuje produkci inzulínu. Mastné kyseliny v beta buňkách aktivují FOXO1, což vede k apoptóze beta buněk.[26]
Regulace buněčného cyklu
Aktivace FOXO1 hraje roli v progrese buněčného cyklu nařízení.[25] Přepis a poločas rozpadu inhibitor cyklin-dependentní kinázy p27KIP1 stoupá, když je aktivní FOXO1.[25] Studie zjistila, že FOXO1 reguluje nukleární lokalizaci p27KIP1 v prasečích granulózních buňkách a dopady buněčný cyklus postup.[25] Kromě toho je spojena zástava buněčného cyklu zprostředkovaná FOXO1 cyklin D1 a cyklin D2 potlačení u savců.[25] Bylo zjištěno, že lidský FOXO1 je spojen s cyklin D1 promotor využívající imunoprecipitační testy na chromatinu (ChIP testy ).[25] H215R je lidský mutant FOXO1, který se nemůže vázat na kanonický FRE k vyvolání exprese p27KIP1, potlačovat cyklin D1 a cyklin D2 propagační činnost a povzbuzuje buněčný cyklus zástava při cyklinu G1 (CCNG1 ).[25] V důsledku toho aktivace FOXO1 brání cyklus buněčného dělení při cyklinu G1 (CCNG1 ) jedním z dvou způsobů stimulace nebo potlačení genové transkripce.[25]
Mechanismus účinku
Ve svém nefosforylovaném stavu je FOXO1 lokalizován do jádra, kde se váže na sekvenci inzulínové odpovědi umístěnou v promotér pro glukóza 6-fosfatáza a zvyšuje jeho rychlost transkripce. FOXO1 prostřednictvím zvýšení transkripce glukóza-6-fosfatázy nepřímo zvyšuje rychlost produkce glukózy v játrech.[22] Pokud je však FOXO1 fosforylován Akt na Thr-24, Ser-256 a Ser-319 je vyloučen z jádra, kde je ubikvitinovaný a degradován. The fosforylace FOXO1 podle Akt následně snižuje produkci glukózy v játrech snížením transkripce 6-fosfatázy glukózy.
Nařízení
Existují tři procesy, a to acetylace, fosforylace, a ubikvitinace které jsou odpovědné za regulaci činnosti vidlicového boxu O1 (FOXO1).[27]
Fosforylace
Fosforylace proteinu FOXO1 je výsledkem aktivace Dráha PI3K / AKT.[27] Sérum a glukokortikoidy indukovatelná kináza SGK může také fosforylovat a inaktivovat transkripční faktor FOXO1.[25] FOXO1 přemístit z jádro na cytoplazma a deaktivovat fosforylace na dobře definovaných místech proteinovými kinázami AKT / SGK1.[27] Transkripční faktor FOXO1 může přímo fosforylovat AKT /SGK1 na třech místech T24, S256 a S319.[28] FOXO1 navíc ztrácí své interakce s DNA když je fosforylován AKT /SGK1 protože S256, který je jedním ze tří serverů AKT / SGK, mění Doména vázající DNA náboj z kladného náboje do záporného náboje.[27]
Inzulinové signální substráty 1 a 2 inzulínové signální kaskády také regulují FOXO1 prostřednictvím fosforylace AKT.[27] AKT, který se označuje jako protein kináza B, fosforyluje FOXO1 a hromadí se v cytosol.[27]
Kasein kináza 1, proteinkináza aktivovaná růstovým faktorem, také fosforyluje a potencuje FOXO1 a translokuje FOXO1 do cytoplazma.[27]
Výzkum
Protože FOXO1 poskytuje spojení mezi transkripcí a metabolickou kontrolou pomocí inzulínu, je také potenciálním cílem genetické kontroly Diabetes typu 2. V myším modelu rezistentním na inzulín je zvýšená produkce glukózy v játrech v důsledku ztráty citlivosti na inzulín; rychlost jaterní glukoneogeneze a glykogenolýzy se zvyšuje ve srovnání s normálními myšmi; je to pravděpodobně způsobeno neregulovaným FOXO1. Když se stejný experiment opakoval s haploinsufficient FOXO1, citlivost na inzulín se částečně obnovila a produkce jaterní glukózy se následně snížila.[29] Podobně u myší krmených stravou s vysokým obsahem tuků (HFD) dochází ke zvýšené inzulínové rezistenci v kostních a jaterních buňkách. Když však byly haploinsufficient FOXO1 myši léčeny stejným HFD, došlo k výraznému snížení inzulínové rezistence jak v kosterních, tak v jaterních buňkách. Tento účinek byl významně zvýšen současným podáváním rosiglitazon, což je běžně předepisovaný antidiabetikum.[30] Tyto výsledky vytvářejí příležitost pro nový přístup založený na genové terapii ke zmírnění desenzibilizace inzulínu u diabetu 2. typu.
U cukrovky (typu 1 i typu 2) přispívá glukoneogeneze v ledvinách ke glukóze v krvi více než u normálních subjektů.[31] Zesílení potlačení FOXO1 inzulinem může snížit glukoneogenezi v játrech i ledvinách.[31]
U myší krmených HFD byla kombinace FOXO1 a haploinsufficiency Notch-1 účinnější při obnovení citlivosti na inzulín než samotná FOXO1 haploinsufficiency.[32]
Buňky produkující inzulín by mohly být generovány inhibicí FOXO1 ve střevě organoidy generované ze střevních kmenových buněk izolovaných z dospělé tkáně.[33]
Klinický význam
- Translokace tohoto genu pomocí PAX3 byl spojen s alveolární rhabdomyosarkom.[5][34]
- v Glukoneogeneze Gen FOXO1 reguluje glukóza hladiny kvůli nízkému výkonu jater glukóza.[27] U myší řeže krev nalačno glukóza hladiny inhibicí formulace glukoneogenní geny.[27]
- FOXO1 hraje roli v ochraně buněk před oxidační stres.[27] Zdá se, že podporuje smrt buněk, když oxidační stres je vysoký v tkáních, které se účastní diabetických komplikací.[27] V takových situacích má místo ochranné role destruktivní roli.[27]
- FOXO1 pomáhá hojení ran u myší prostřednictvím koordinace reakce keratinocyty a funkce v keratinocyty svrhnout oxidační stres.[27] Hojení ran je velmi komplikovaný biologický proces a studie ukázaly, že transkripční faktor FOXO1 pomáhá při řízení událostí, které zlepšují proces hojení keratinocyty.[35] Lokalizace jaderné energie FOXO1 se v roce 2004 zvýšila čtyřikrát hojení ran keratinocyty.[35] Podporuje migraci EU keratinocyty prostřednictvím regulace regulace růstového faktoru.[35]
- V Vrozený imunitní systém Bylo prokázáno, že FOXO1 zvyšuje zánět prostřednictvím zvýšené formulace několika prozánětlivých genů.[27] Zprostředkovává formulaci prozánětlivé cytokiny v reakci na vysokou glukóza úrovně, TNF a stimulace LPS.[27]
- v Adaptivní imunitní systém FOXO1 reguluje návrat periferních zařízení B buňky upregulací L-sekce a kontroluje rekombinaci periferního přepínače třídy B buňky a v T buňky zvyšuje přežití CD8 Paměť.[27]
- v Karcinogeneze FOXO1 hraje roli nádorového supresoru a jeho inaktivace byla dokumentována u mnoha druhů lidí rakovina.[27] Potlačuje přežití nádorové buňky indukcí apoptóza v buňkách rakoviny prostaty a buňkách gliomu zvýšením regulace proapoptotických faktorů.[27] Zvýšená aktivace FOXO1 může inhibovat metastáza buněk rakoviny prostaty do jiných orgánů potlačením migrace a invaze nebo potlačením Běhová doména obsahující Runx2 transkripční aktivita.[27]
Interakce
Bylo prokázáno, že FOXO1 komunikovat s:
- androgenní receptor,[36]
- estrogenový receptor alfa,[37]
- CREB-vazebný protein,[38] a
- protein tuberózní sklerózy 2.[39]
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000150907 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000044167 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b Galili N, Davis RJ, Fredericks WJ, Mukhopadhyay S, Rauscher FJ, Emanuel BS, Rovera G, Barr FG (listopad 1993). "Fúze genu domény vidlicové hlavy k PAX3 v alveolárním rhabdomyosarkomu solidního nádoru". Genetika přírody. 5 (3): 230–5. doi:10.1038 / ng1193-230. PMID 8275086.
- ^ Nakae J, Kitamura T, Kitamura Y, Biggs WH, Arden KC, Accili D (leden 2003). "Frakční transkripční faktor Foxo1 reguluje diferenciaci adipocytů". Vývojová buňka. 4 (1): 119–29. doi:10.1016 / S1534-5807 (02) 00401-X. PMID 12530968.
- ^ Rena G, Guo S, Cichy SC, Unterman TG, Cohen P (červen 1999). „Fosforylace člena rodiny FKHR transkripčního faktoru pomocí proteinkinázy B“. The Journal of Biological Chemistry. 274 (24): 17179–83. doi:10.1074 / jbc.274.24.17179. PMID 10358075.
- ^ Guo S, Rena G, Cichy S, He X, Cohen P, Unterman T (červen 1999). „Fosforylace serinu 256 proteinovou kinázou B narušuje transaktivaci pomocí FKHR a zprostředkovává účinky inzulínu na aktivitu promotoru proteinu-1 vázajícího růstový faktor prostřednictvím konzervované sekvence inzulínové odpovědi“. The Journal of Biological Chemistry. 274 (24): 17184–92. doi:10.1074 / jbc.274.24.17184. PMID 10358076.
- ^ Farmář SR (leden 2003). „Frakční transkripční faktor Foxo1: možná souvislost mezi obezitou a inzulínovou rezistencí“. Molekulární buňka. 11 (1): 6–8. doi:10.1016 / S1097-2765 (03) 00003-0. PMID 12535515.
- ^ Armoni M, Harel C, Karni S, Chen H, Bar-Yoseph F, Ver MR, Quon MJ, Karnieli E (červenec 2006). „FOXO1 potlačuje promotory genu receptoru aktivovaného peroxisomovým proliferátorem - gama1 a -gama2 v primárních adipocytech. Nové paradigma pro zvýšení citlivosti na inzulín“. The Journal of Biological Chemistry. 281 (29): 19881–91. doi:10,1074 / jbc.M600320200. PMID 16670091.
- ^ Puigserver P, Rhee J, Donovan J, Walkey CJ, Yoon JC, Oriente F, Kitamura Y, Altomonte J, Dong H, Accili D, Spiegelman BM (květen 2003). „Inzulínem regulovaná jaterní glukoneogeneze prostřednictvím interakce FOXO1-PGC-1alfa“. Příroda. 423 (6939): 550–5. doi:10.1038 / nature01667. PMID 12754525.
- ^ Adachi M, Osawa Y, Uchinami H, Kitamura T, Accili D, Brenner DA (duben 2007). „Frakční transkripční faktor FoxO1 reguluje proliferaci a transdiferenciaci jaterních stelátových buněk“ (PDF). Gastroenterologie. 132 (4): 1434–46. doi:10.1053 / j.gastro.2007.01.033. PMID 17408630.
- ^ „Entrez Gene: FOXO1 forkhead box O1 (rhabdomyosarcoma)“.
- ^ Zhang X, Yalcin S, Lee DF, Yeh TY, Lee SM, Su J, Mungamuri SK, Rimmelé P, Kennedy M, Sellers R, Landthaler M, Tuschl T, Chi NW, Lemischka I, Keller G, Ghaffari S (červenec 2011 ). „FOXO1 je základním regulátorem pluripotence v lidských embryonálních kmenových buňkách“. Přírodní buněčná biologie. 13 (9): 1092–9. doi:10.1038 / ncb2293. PMC 4053529. PMID 21804543.
- ^ Nagashima T, Shigematsu N, Maruki R, Urano Y, Tanaka H, Shimaya A, Shimokawa T, Shibasaki M (listopad 2010). „Objev nových inhibitorů O1 forkhead boxu pro léčbu cukrovky typu 2: zlepšení glykémie nalačno u diabetických myší db / db“. Molekulární farmakologie. 78 (5): 961–70. doi:10,1124 / mol. 110,065714. PMID 20736318.
- ^ McKinnon CM, Ravier MA, Rutter GA (prosinec 2006). „FoxO1 je vyžadován pro regulaci exprese genu preproglukagonu inzulinem v pankreatických buňkách alfaTC1-9“. The Journal of Biological Chemistry. 281 (51): 39358–69. doi:10,1074 / jbc.M605022200. PMID 17062568.
- ^ Buteau J, Spatz ML, Accili D (květen 2006). „Transkripční faktor FoxO1 zprostředkovává účinky peptidu-1 podobného glukagonu na hmotnost beta-buněk pankreatu“. Cukrovka. 55 (5): 1190–6. doi:10.2337 / db05-0825. PMID 16644672.
- ^ Rena G, Woods YL, Prescott AR, Peggie M, Unterman TG, Williams MR, Cohen P (květen 2002). „Dvě nová fosforylační místa na FKHR, která jsou zásadní pro jeho jaderné vyloučení“. Časopis EMBO. 21 (9): 2263–71. doi:10.1093 / emboj / 21.9.2263. PMC 125977. PMID 11980723.
- ^ Rena G, Prescott AR, Guo S, Cohen P, Unterman TG (březen 2001). „Role vidlice v místech fosforylace rhabdomyosarkomu (FKHR) při regulaci 14-3-3 vazby, transaktivace a jaderného cílení“. The Biochemical Journal. 354 (Pt 3): 605–12. doi:10.1042/0264-6021:3540605. PMC 1221692. PMID 11237865.
- ^ Matsuzaki H, Daitoku H, Hatta M, Tanaka K, Fukamizu A (září 2003). „Inzulínem indukovaná fosforylace cílů FKHR (Foxo1) na proteazomální degradaci“. Sborník Národní akademie věd Spojených států amerických. 100 (20): 11285–90. doi:10.1073 / pnas.1934283100. PMC 208749. PMID 13679577.
- ^ Daitoku H, Fukamizu A (červen 2007). "FOXO transkripční faktory v regulačních sítích dlouhověkosti". Journal of Biochemistry. 141 (6): 769–74. doi:10.1093 / jb / mvm104. PMID 17569704.
- ^ A b Nakae J, Kitamura T, Silver DL, Accili D (listopad 2001). „Frakční transkripční faktor Foxo1 (Fkhr) uděluje citlivost na inzulín expresi glukóza-6-fosfatázy“. The Journal of Clinical Investigation. 108 (9): 1359–67. doi:10,1172 / JCI12876. PMC 209440. PMID 11696581.
- ^ Matsuzaki H, Daitoku H, Hatta M, Aoyama H, Yoshimochi K, Fukamizu A (srpen 2005). „Acetylace Foxo1 mění jeho schopnost vázat DNA a citlivost na fosforylaci“. Sborník Národní akademie věd Spojených států amerických. 102 (32): 11278–83. doi:10.1073 / pnas.0502738102. PMC 1183558. PMID 16076959.
- ^ Jing E, Gesta S, Kahn CR (srpen 2007). „SIRT2 reguluje diferenciaci adipocytů prostřednictvím acetylace / deacetylace FoxO1“. Buněčný metabolismus. 6 (2): 105–14. doi:10.1016 / j.cmet.2007.07.003. PMC 2083635. PMID 17681146.
- ^ A b C d E F G h i j k l m n Lu H, Huang H (srpen 2011). „FOXO1: potenciální cíl pro lidské nemoci“. Aktuální drogové cíle. 12 (9): 1235–44. doi:10.2174/138945011796150280. PMC 4591039. PMID 21443466.
- ^ Sun T, Han X (2019). „Smrt versus dediferenciace: Molekulární základy snížení hmotnosti beta buněk u diabetu 2. typu“. Semináře z buněčné a vývojové biologie. doi:10.1016 / j.semcdb.2019.12.002. PMID 31831356.
- ^ A b C d E F G h i j k l m n Ó p q r s Wang Y, Zhou Y, Graves DT (duben 2014). „Faktory transkripce FOXO: jejich klinický význam a regulace“. BioMed Research International. 2014: 925350. doi:10.1155/2014/925350. PMC 4016844. PMID 24864265.
- ^ Hedrick SM, Hess Michelini R, Doedens AL, Goldrath AW, Stone EL (září 2012). „Transkripční faktory FOXO v biologii T buněk“. Recenze přírody. Imunologie. 12 (9): 649–61. doi:10.1038 / nri3278. PMC 3875397. PMID 22918467.
- ^ Nakae J, Biggs WH, Kitamura T, Cavenee WK, Wright CV, Arden KC, Accili D (říjen 2002). „Regulace působení inzulínu a funkce beta-buněk pankreatu mutovanými alelami genu kódujícího transkripční faktor vidlice Foxo1“. Genetika přírody. 32 (2): 245–53. doi:10.1038 / ng890. PMID 12219087.
- ^ Kim JJ, Li P, Huntley J, Chang JP, Arden KC, Olefsky JM (červen 2009). „Haploinsufficiency FoxO1 chrání před stravou s vysokým obsahem tuků vyvolanou inzulínovou rezistencí se zvýšenou aktivací gamma receptoru aktivovaného proliferátorem peroxisomu v tukové tkáni. Cukrovka. 58 (6): 1275–82. doi:10 2337 / db08-1001. PMC 2682681. PMID 19289458.
- ^ A b Swe MT, Pongchaidecha A, Chatsudthipong V, Chattipakorn N, Lungkaphin A (2019). "Molekulární signální mechanismy renální glukoneogeneze u nediabetických a diabetických stavů". Journal of Cellular Physiology. 234 (6): 8134–8151. doi:10,1002 / jcp.27598. PMID 30370538.
- ^ Pajvani UB, Shawber CJ, Samuel VT, Birkenfeld AL, Shulman GI, Kitajewski J, Accili D (červenec 2011). „Inhibice signalizace Notch zlepšuje inzulínovou rezistenci způsobem závislým na FoxO1“. Přírodní medicína. 17 (8): 961–7. doi:10,1038 / nm.2378. PMC 3387563. PMID 21804540.
- ^ Bouchi R, Foo KS, Hua H, Tsuchiya K, Ohmura Y, Sandoval PR, Ratner LE, Egli D, Leibel RL, Accili D (červen 2014). „Inhibice FOXO1 poskytuje funkční buňky produkující inzulín v lidských střevních organoidních kulturách“. Příroda komunikace. 5: 4242. doi:10.1038 / ncomms5242. PMC 4083475. PMID 24979718.
- ^ Linardic CM (říjen 2008). „Fúzní gen PAX3-FOXO1 v rhabdomyosarkomu“. Dopisy o rakovině. 270 (1): 10–8. doi:10.1016 / j.canlet.2008.03.035. PMC 2575376. PMID 18457914.
- ^ A b C Xiao E, Graves DT (srpen 2015). „Dopad diabetu na ochrannou roli FOXO1 při hojení ran“. Journal of Dental Research. 94 (8): 1025–6. doi:10.1177/0022034515586353. PMC 530387. PMID 25978971.
- ^ Li P, Lee H, Guo S, Unterman TG, Jenster G, Bai W (leden 2003). „AKT-nezávislá ochrana buněk rakoviny prostaty před apoptózou zprostředkovanou tvorbou komplexů mezi androgenovým receptorem a FKHR“. Molekulární a buněčná biologie. 23 (1): 104–18. doi:10.1128 / MCB.23.1.104-118.2003. PMC 140652. PMID 12482965.
- ^ Schuur ER, Loktev AV, Sharma M, Sun Z, Roth RA, Weigel RJ (září 2001). „Ligandově závislá interakce estrogenového receptoru-alfa s členy rodiny transkripčních faktorů vidlice“. The Journal of Biological Chemistry. 276 (36): 33554–60. doi:10,1074 / jbc.M105555200. PMID 11435445.
- ^ Nasrin N, Ogg S, Cahill CM, Biggs W, Nui S, Dore J, Calvo D, Shi Y, Ruvkun G, Alexander-Bridges MC (září 2000). „DAF-16 rekrutuje komplex koaktivátoru vázajícího protein CREB na promotor proteinu vázajícího růstový faktor podobný inzulínu 1 v buňkách HepG2“. Sborník Národní akademie věd Spojených států amerických. 97 (19): 10412–7. doi:10.1073 / pnas.190326997. PMC 27038. PMID 10973497.
- ^ Cao Y, Kamioka Y, Yokoi N, Kobayashi T, Hino O, Onodera M, Mochizuki N, Nakae J (prosinec 2006). „Interakce FoxO1 a TSC2 indukuje inzulínovou rezistenci aktivací savčího cíle dráhy rapamycinu / p70 S6K“. The Journal of Biological Chemistry. 281 (52): 40242–51. doi:10,1074 / jbc.M608116200. PMID 17077083.
externí odkazy
- FOXO1A + protein, + člověk v americké národní lékařské knihovně Lékařské předměty (Pletivo)
Tento článek včlení text z United States National Library of Medicine, který je v veřejná doména.