Faktor určující varlata - Testis-determining factor





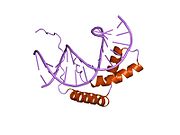
Faktor určující varlata (TDF), také známý jako oblast určující pohlaví Y (SRY) protein, je Protein vázající DNA (také známý jako genově regulační protein /transkripční faktor ) zakódováno SRY gen který je zodpovědný za zasvěcení muže určení pohlaví v Therian savci (placentární savci a vačnatci ).[5] SRY je intronless sex - určující gen na Y chromozom.[6] Mutace v tomto genu vedou k řadě poruchy pohlavního vývoje (DSD) s různými účinky na fenotyp a genotyp jednotlivce.
TDF je členem Gen SOX (SRY-like box) rodina DNA vázající proteiny. V komplexu s Protein SF1, TDF funguje jako transkripční faktor který způsobí upregulaci ostatních transkripční faktory nejdůležitější SOX9.[7] Své výraz způsobuje rozvoj primárního sexuální šňůry, které se později vyvinuly do semenonosné tubuly. Tyto šňůry se tvoří ve střední části dosud nediferencovaného pohlavní žláza, přeměnit to na varle. Nyní vyvolané Leydigovy buňky varlete pak začněte vylučovat testosteron, zatímco Sertoliho buňky vyrobit anti-Müllerian hormon.[8] Účinky genu SRY se obvykle vyskytují 6–8 týdnů po tvorbě plodu, což inhibuje anatomický strukturální růst žen u mužů. Pracuje také na rozvoji dominantních mužských vlastností.
Genový vývoj a regulace
Vývoj
SRY může vzniknout z a genová duplikace genu vázaného na chromozom X SOX3, člen Soxova rodina.[9][10] K této duplikaci došlo po rozdělení mezi monotremes a Therians. Monotrémy postrádají SRY a některé jejich pohlavní chromozomy sdílejí homologii s ptačími chromozomy.[11] SRY je rychle se vyvíjející gen a jeho regulace byla obtížně studovatelná, protože určování pohlaví není v živočišné říši vysoce konzervovaným jevem.[12] Dokonce i uvnitř vačnatci a placenty, které používají SRY v jejich procesu určování pohlaví, akce SRY liší se mezi druhy.[10] Genová sekvence se také mění; zatímco jádro genu, Skupina s vysokou mobilitou (HMG) pole, je mezi druhy konzervativní, jiné oblasti genu nejsou.[10] SRY je jedním z pouhých čtyř genů na lidském chromozomu Y, u nichž bylo prokázáno, že vznikly z původního chromozomu Y.[13] Ostatní geny na lidském chromozomu Y vznikly z autosomu, který se spojil s původním chromozomem Y.[13]
Nařízení
SRY Gen má málo společného s geny určujícími pohlaví jiných modelových organismů, proto jsou myši hlavním modelovým výzkumným organismem, který lze pro jeho studium využít. Pochopení jeho regulace je dále komplikováno, protože i mezi druhy savců existuje jen málo zachování proteinové sekvence. Jedinou konzervovanou skupinou mezi myší a jinými savci je Skupina s vysokou mobilitou (HMG) pole oblast, která je zodpovědná za vazbu DNA. Výsledkem mutací v této oblasti je obrácení pohlaví, kde se vyrábí opačné pohlaví.[14] Protože tam je malá ochrana, SRY promotor, regulační prvky a regulace nejsou dobře známy. V rámci příbuzných savčích skupin existují homologie v rámci prvních 400-600 párů bází proti proudu od počátečního místa translace. In vitro studie na člověku SRY promotor ukázal, že pro oblast je vyžadována oblast alespoň 310 bp proti směru od počátečního místa translace SRY funkce promotoru. Bylo prokázáno, že vazba tří transkripčních faktorů, steroidogenního faktoru 1 (SF1 ), Specificita Protein 1 (Transkripční faktor Sp1 ) a Wilmsův nádorový protein 1 (WT1 ), na sekvenci lidského promotoru, ovlivňují expresi SRY.[14]
Oblast promotor má dvě Sp1 vazebná místa, na -150 a -13, která fungují jako regulační místa. Sp1 je transkripční faktor, který váže konsensuální sekvence bohaté na GC a mutaci SRY vazebná místa vede k 90% snížení transkripce genu. Studie z SF1 vedly k méně jednoznačným výsledkům. Mutace SF1 může vést k obrácení pohlaví a vypuštění vede k neúplnému vývoji pohlavních žláz. Není však jasné, jak SF1 interaguje s SR1 přímo promotér.[15] Oblast promotor má také dvě WT1 vazebná místa na -78 a -87 bp z kodonu ATG. WT1 je transkripční faktor, který má čtyři C-konce Zinkové prsty a N-koncová oblast bohatá na Pro / Glu a primárně funguje jako aktivátor. Mutace Zinkové prsty nebo deaktivace WT1 má za následek zmenšení velikosti mužské pohlavní žlázy. Odstranění genu vedlo k dokončení obrácení pohlaví. Není jasné, jak WT1 funkce pro regulaci nahoru SRYNěkteré výzkumy však naznačují, že pomáhá stabilizovat zpracování zpráv.[15] Tato hypotéza však má komplikace, protože WT1 je také zodpovědný za expresi antagonisty mužského vývoje, DAX1, což je zkratka pro obrácení pohlaví citlivé na dávkování, kritická oblast hypoplázie nadledvin, na chromozomu X, gen 1. Další kopie DAX1 u myší vede k obrácení pohlaví. Není jasné, jak DAX1 funkcí a bylo navrženo mnoho různých cest, včetně SRY transkripční destabilizace a vazba RNA. Z práce na potlačení mužského vývoje existují důkazy DAX1 může interferovat s funkcí SF1 a následně přepis SRY náborem korepresorů.[14]
Existují také důkazy, že GATA vázající protein 4 (GATA4) a FOG2 přispívají k aktivaci SRY sdružením s jeho promotérem. Jak tyto proteiny regulují SRY transkripce není jasná, ale mutanty FOG2 a GATA4 mají významně nižší hladiny SRY transkripce.[16] FOG mají motivy zinkových prstů, které mohou vázat DNA, ale neexistují žádné důkazy o interakci FOG2 s SRY. Studie naznačují, že FOG2 a GATA4 se asociují s proteiny remodelace nukleosomů, což by mohlo vést k jeho aktivaci.[17]
Funkce
Během těhotenství jsou buňky prvotní pohlavní žlázy, které leží podél urogenitálního hřebene, v bipotenciálním stavu, což znamená, že mají schopnost stát se buď mužskými buňkami (Sertoli a Leydig buňky) nebo ženské buňky (folikul buňky a theca buňky). TDF iniciuje diferenciaci varlat aktivací transkripčních faktorů specifických pro muže, které těmto bipotenciálním buňkám umožňují diferenciaci a proliferaci. TDF toho dosahuje zvýšenou regulací SOX9, transkripční faktor s vazebným místem pro DNA velmi podobný TDF. SOX9 vede k upregulaci růstového faktoru 9 fibroblastů (Fgf9 ), což zase vede k další upregulaci SOX9. Jakmile je dosaženo správné hladiny SOX9, bipotenciální buňky pohlavní žlázy se začnou diferencovat na buňky Sertoli. Kromě toho budou buňky exprimující TDF pokračovat v proliferaci za vzniku prvotního varlete. I když se jedná o základní řadu událostí, je třeba tento krátký přehled brát opatrně, protože existuje mnoho dalších faktorů, které ovlivňují diferenciaci pohlaví.
Akce v jádru
Protein TDF se skládá ze tří hlavních oblastí. Centrální oblast zahrnuje doménu HMG (skupina s vysokou mobilitou), která obsahuje sekvence nukleární lokalizace a působí jako doména vázající DNA. The C-terminál doména nemá žádnou konzervovanou strukturu a N-terminál doménu lze fosforylovat, aby se zvýšila vazba na DNA.[15] Proces začíná jaderná lokalizace TDF do acetylace signálních oblastí jaderné lokalizace, což umožňuje vazbu importin β a klimodulin k TDF, což usnadňuje jeho import do jádra. Jakmile jsou v jádru, TDF a SF1 (steroidogenní faktor 1, další regulátor transkripce) komplexují a váží se na TESCO (varlat specifický enhancer jádra Sox9), varlat specifický enhancerový prvek genu Sox9 v prekurzorech buněk Sertoli, umístěných proti proudu od místa začátku transkripce genu Sox9.[7] Konkrétně se jedná o HMG oblast TDF, která se váže na vedlejší drážku cílové sekvence DNA, což způsobuje ohnutí a uvolnění DNA. Zavedení této konkrétní „architektury“ DNA usnadňuje transkripci genu Sox9.[15] V jádru Sertoliho buněk SOX9 přímo cílí na Amh gen i prostaglandin D syntáza (Ptgds) gen. Vazba SOX9 na zesilovač poblíž Amh promotor umožňuje syntézu Amh zatímco vazba SOX9 na Ptgds gen umožňuje produkci prostaglandinu D2 (PGD2). Zpětný vstup SOX9 do jádra je usnadněn autokrinní nebo parakrinní signalizací prováděnou PGD2.[18] Protein SOX9 poté zahájí a Pozitivní zpětná vazba smyčka, zahrnující SOX9, který působí jako svůj vlastní transkripční faktor a vede k syntéze velkého množství SOX9.[15]
SOX9 a diferenciace varlat
The Protein SF1 sám o sobě vede k minimální transkripci SOX9 gen v obou bipotenciálních gonadálních buňkách XX a XY podél urogenitálního hřebene. Vazba komplexu TDF-SF1 na testespecifický enhancer (TESCO) na SOX9 však vede k významné up-regulaci genu pouze v gonádě XY, zatímco transkripce v gonádě XX zůstává zanedbatelná. Část této up-regulace se dosahuje samotným SOX9 prostřednictvím smyčky pozitivní zpětné vazby; jako TDF, SOX9 komplexuje s SF1 a váže se na TESCO enhancer, což vede k další expresi SOX9 v XY gonádě. Dva další proteiny, FGF9 (fibroblastový růstový faktor 9) a PDG2 (prostaglandin D2) také udržují tuto regulaci nahoru. Ačkoli jejich přesné dráhy nejsou plně pochopeny, ukázalo se, že jsou nezbytné pro pokračující expresi SOX9 na úrovních nezbytných pro vývoj varlat.[7]
SOX9 Předpokládá se, že TDF a TDF jsou odpovědné za buněčnou autonomní diferenciaci podpůrných buněčných prekurzorů v pohlavních žlázách do Sertoliho buněk, což je začátek vývoje varlat. U těchto počátečních Sertoliho buněk ve středu gonády se předpokládá, že jsou výchozím bodem pro vlnu FGF9, která se šíří po vyvíjející se gonádě XY, což vede k další diferenciaci Sertoliho buněk prostřednictvím upregulace SOX9.[19] SOX9 a TDF jsou také považovány za odpovědné za mnoho pozdějších procesů vývoje varlat (jako je diferenciace Leydigových buněk, tvorba pohlavní šňůry a tvorba varlat specifické pro varlata), ačkoli přesné mechanismy zůstávají nejasné.[20] Ukázalo se však, že SOX9 v přítomnosti PDG2 působí přímo na Amh (kódující anti-Müllerianův hormon) a je schopen vyvolat tvorbu varlat v gonádách myší XX, což naznačuje jeho zásadní význam pro vývoj varlat.[19]
Vliv poruch SRY na sexuální výraz
Embrya jsou gonadálně identická, bez ohledu na genetické pohlaví, až do určitého bodu vývoje, kdy faktor určující varlata způsobí vývoj mužských pohlavních orgánů. Typickým mužským karyotypem je XY, zatímco ženským je XX. Existují však výjimky, ve kterých hraje SRY hlavní roli. Jednotlivci s Klinefelterův syndrom zdědí normální chromozom Y a více chromozomů X, což jim dává karyotyp XXY. Tyto osoby jsou považovány za muže.[21] Atypická genetická rekombinace během křížení, kdy se vyvíjí spermie, může vést k karyotypům, které neodpovídají jejich fenotypové expresi.
Většinu času, když vyvíjející se spermie prochází crossoverem během meiózy, zůstává gen SRY na chromozomu Y. Pokud se gen SRY přenese do chromozomu X místo toho, aby zůstal na chromozomu Y, k vývoji varlat již nedojde. Toto je známé jako Swyerův syndrom, charakterizovaný karyotypem XY a ženským fenotypem. Jedinci, kteří mají tento syndrom, mají normálně vytvořené dělohy a vejcovody, ale pohlavní žlázy nejsou funkční. Jednotlivci Swyerova syndromu jsou obecně vychováváni jako ženy a mají ženskou genderovou identitu.[22] Na druhém spektru se XX mužský syndrom vyskytuje, když má tělo ženské chromozomy a SRY se k jednomu z nich připojí translokací. Lidé s mužským syndromem XX mají ženský genotyp, ale mužské fyzické rysy.[23] Jedinci s jedním z těchto syndromů mohou mít opožděnou pubertu, neplodnost a růstové rysy opačného pohlaví, se kterým se ztotožňují. U expresorů XX mužského syndromu se může vyvinout prsa a u pacientů se Swyerovým syndromem se mohou vyskytnout vousy.[22][24]
| Klinefelterův syndrom |
|
| Swyerův syndrom |
|
| XX mužský syndrom |
|
I když přítomnost nebo nepřítomnost SRY obecně určovala, zda dojde nebo nedojde k vývoji varlat, bylo navrženo, že existují další faktory, které ovlivňují funkčnost SRY.[25] Proto existují jedinci, kteří mají gen SRY, ale stále se vyvíjejí jako ženy, buď proto, že samotný gen je defektní nebo mutovaný, nebo proto, že je defektní jeden z přispívajících faktorů.[26] K tomu může dojít u jedinců vykazujících karyotyp pozitivní na XY, XXY nebo XX SRY.
Další systémy určující pohlaví, které se spoléhají na SRY / TDF nad XY, jsou procesy, které přicházejí poté, co je SRY přítomen nebo chybí ve vývoji embrya. V normálním systému, pokud je SRY přítomen pro XY, TDF aktivuje dřeň a vyvine pohlavní žlázy do varlat. Poté bude produkován testosteron a zahájen vývoj dalších mužských pohlavních znaků. Srovnatelně, pokud SRY není přítomen pro XX, bude nedostatek TDF založený na žádném chromozomu Y. Nedostatek TDF umožní kůře embryonálních pohlavních žláz vyvinout se do vaječníků, které pak budou produkovat estrogen, a povedou k rozvoji dalších ženských sexuálních charakteristik.[27]
Role v jiných nemocech
SRY bylo prokázáno komunikovat s androgenní receptor a jedinci s karyotypem XY a funkčním genem SRY mohou mít navenek ženský fenotyp kvůli podkladové syndrom androgenní necitlivosti (AIS).[28] Jedinci s AIS nejsou schopni správně reagovat na androgeny kvůli defektu genu pro androgenní receptor a postižení jedinci mohou mít úplnou nebo částečnou AIS.[29] SRY souvisí také se skutečností, že u mužů je pravděpodobnější vývoj než u žen dopamin související choroby, jako je schizofrenie a Parkinsonova choroba. SRY kóduje protein, který řídí koncentraci dopaminu, neurotransmiteru, který přenáší signály z mozku, které řídí pohyb a koordinaci.[30] Výzkum na myších ukázal, že mutace v SOX10, transkripční faktor kódovaný SRY, je spojen se stavem dominantního megakolonu u myší.[31] Tento model myši se používá ke zkoumání vazby mezi SRY a Hirschsprungova choroba nebo vrozený megakolon u lidí.[31] Existuje také souvislost mezi SRY kódovaným transkripčním faktorem SOX9 a kampomelická dysplázie (CD).[32] Tato mutace missense je vadná chondrogeneze nebo proces tvorby chrupavky a projevuje se jako kosterní CD.[33] Dvě třetiny z 46 XY jedinců s diagnostikovanou CD mají kolísavé množství reverze pohlaví mezi ženami.[32]
Použití při olympijském screeningu
Jedno z nejkontroverznějších použití tohoto objevu bylo prostředkem ověření pohlaví na olympijské hry, v rámci systému implementovaného Mezinárodní olympijský výbor v roce 1992. Sportovcům s genem SRY nebylo povoleno účastnit se jako žen, i když všichni sportovci, u kterých to bylo „zjištěno“ na Letní olympijské hry 1996 byli ovládáni falešně pozitivní výsledky a nebyli diskvalifikováni. Konkrétně bylo zjištěno, že osm ženských účastnic (z celkového počtu 3387) v těchto hrách mělo gen SRY. Po dalším zkoumání jejich genetických podmínek však byli všichni tito sportovci ověřeni jako ženy a bylo jim umožněno soutěžit. Bylo zjištěno, že tito sportovci mají buď částečné, nebo úplné necitlivost na androgeny, navzdory tomu, že mají gen SRY, což z nich dělá fenotypicky samice a nedává jim žádnou výhodu oproti jiným ženským konkurentům.[34] Na konci 90. let vyzvala řada příslušných profesních společností ve Spojených státech k odstranění ověření pohlaví, včetně Americká lékařská asociace s uvedením, že použitá metoda byla nejistá a neúčinná.[35] Chromozomální screening byl k Letní olympijské hry 2000,[35][36][37] ale poté následovaly další formy testování založené na hladinách hormonů.[38]
Probíhající výzkum
Navzdory pokroku, kterého bylo v posledních několika desetiletích dosaženo ve studiu určování pohlaví, genu SRY a proteinu TDF, stále probíhá práce na dalším porozumění v těchto oblastech. V molekulární síti určující pohlaví stále existují faktory, které je třeba identifikovat, a chromozomální změny v mnoha dalších případech zvrácení lidského pohlaví nejsou dosud známy. Vědci pokračují v hledání dalších genů určujících pohlaví pomocí technik, jako je microarray screening genitálních hřebenových genů v různých vývojových stádiích, screening mutageneze u myší na fenotypy reverze pohlaví a identifikace genů, na které transkripční faktory působí imunoprecipitace chromatinu.[15]
Viz také
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000184895 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000069036 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Berta P, Hawkins JR, Sinclair AH, Taylor A, Griffiths BL, Goodfellow PN, Fellous M (listopad 1990). "Genetický důkaz, který se rovná SRY a faktoru určujícím varlata". Příroda. 348 (6300): 448–50. Bibcode:1990 Natur.348..448B. doi:10.1038 / 348448A0. PMID 2247149. S2CID 3336314.
- ^ Wallis MC, Waters PD, Graves JA (říjen 2008). "Určení pohlaví u savců - před a po vývoji SRY". Buněčné a molekulární biologické vědy. 65 (20): 3182–95. doi:10.1007 / s00018-008-8109-z. PMID 18581056. S2CID 31675679.
- ^ A b C Kashimada K, Koopman P (prosinec 2010). „Sry: hlavní přepínač v určování pohlaví savců“. Rozvoj. 137 (23): 3921–30. doi:10,1242 / dev.048983. PMID 21062860.
- ^ Mittwoch U (říjen 1988). "Rasa být mužem". Nový vědec. 120 (1635): 38–42.
- ^ Katoh K, Miyata T (prosinec 1999). „Heuristický přístup metody maximální věrohodnosti pro odvození fylogenetického stromu a aplikace savčího původu SOX-3 genu SRY určujícího varlata“. FEBS Dopisy. 463 (1–2): 129–32. doi:10.1016 / S0014-5793 (99) 01621-X. PMID 10601652. S2CID 24519808.
- ^ A b C Bakloushinskaya, I Y (2009). „Vývoj determinace pohlaví u savců“. Bulletin biologie. 36 (2): 167–174. doi:10.1134 / S1062359009020095. S2CID 36988324.
- ^ Veyrunes F, Waters PD, Miethke P, Rens W, McMillan D, Alsop AE, Grützner F, Deakin JE, Whittington CM, Schatzkamer K, Kremitzki CL, Graves T, Ferguson-Smith MA, Warren W, Marshall Graves JA (červen 2008 ). „Sexuální chromozomy ptakopysků podobné ptákům naznačují nedávný původ sexuálních chromozomů savců“. Výzkum genomu. 18 (6): 965–73. doi:10,1101 / gr. 7101908. PMC 2413164. PMID 18463302.
- ^ Bowles J, Schepers G, Koopman P (listopad 2000). "Fylogeneze rodiny SOX vývojových transkripčních faktorů na základě sekvenčních a strukturálních indikátorů". Vývojová biologie. 227 (2): 239–55. doi:10,1006 / dbio.2000.9883. PMID 11071752.
- ^ A b Graves JA (prosinec 2015). „Divní savci poskytují poznatky o vývoji pohlavních chromozomů savců a kompenzaci dávky“. Journal of Genetics. 94 (4): 567–74. doi:10.1007 / s12041-015-0572-3. PMID 26690510. S2CID 186238659.
- ^ A b C Ely D, Underwood A, Dunphy G, Boehme S, Turner M, Milsted A (listopad 2010). "Recenze chromozomu Y, Sry a hypertenze". Steroidy. 75 (11): 747–53. doi:10.1016 / j.steroids.2009.10.015. PMC 2891862. PMID 19914267.
- ^ A b C d E F Harley VR, Clarkson MJ, Argentaro A (srpen 2003). „Molekulární působení a regulace faktorů určujících varlata, SRY (oblast určující pohlaví na chromozomu Y) a SOX9 [SRY-související skupina s vysokou mobilitou (HMG), rámeček 9]“. Endokrinní hodnocení. 24 (4): 466–87. doi:10.1210 / er.2002-0025. PMID 12920151.
- ^ Knower KC, Kelly S, Harley VR (2003). „Zapnutí muže - SRY, SOX9 a určení pohlaví u savců“ (PDF). Cytogenetický a genomový výzkum. 101 (3–4): 185–98. doi:10.1159/000074336. PMID 14684982. S2CID 20940513. Archivovány od originál dne 9. srpna 2017.
- ^ Friedman, Theodore (2011). Advances in Genetics Vol 76. 108: Elsevier Inc. ISBN 9780123864826.CS1 maint: umístění (odkaz)
- ^ Sekido, Ryohei; Lovell-Badge, Robin (2009). „Určení pohlaví a SRY: až k mrknutí a šťouchnutí?“. Trendy v genetice. 25 (1): 19–29. doi:10.1016 / j.tig.2008.10.008. PMID 19027189.
- ^ A b McClelland K, Bowles J, Koopman P (leden 2012). „Určení mužského pohlaví: pohledy na molekulární mechanismy“. Asijský deník andrologie. 14 (1): 164–71. doi:10.1038 / aja.2011.169. PMC 3735148. PMID 22179516.
- ^ Sekido R, Lovell-Badge R (2013). "Genetická kontrola vývoje varlat". Sexuální vývoj. 7 (1–3): 21–32. doi:10.1159/000342221. PMID 22964823.
- ^ Reference, Genetics Home. "Klinefelterův syndrom". Genetická domácí reference. Citováno 3. března 2020.
- ^ A b Reference, Genetics Home. "Swyerův syndrom". Genetická domácí reference. Citováno 3. března 2020.
- ^ „Mužský syndrom XX | Encyclopedia.com“. www.encyclopedia.com. Citováno 3. března 2020.
- ^ Reference, Genetics Home. „46, XX testikulární porucha pohlavního vývoje“. Genetická domácí reference. Citováno 3. března 2020.
- ^ Polanco JC, Koopman P (únor 2007). „Sry a váhavé počátky mužského vývoje“. Vývojová biologie. 302 (1): 13–24. doi:10.1016 / j.ydbio.2006.08.049. PMID 16996051.
- ^ Biason-Lauber A, Konrad D, Meyer M, DeBeaufort C, Schoenle EJ (květen 2009). „Ovarie a ženský fenotyp u dívky s 46, karyotypem XY a mutacemi v genu CBX2“. American Journal of Human Genetics. 84 (5): 658–63. doi:10.1016 / j.ajhg.2009.03.016. PMC 2680992. PMID 19361780.
- ^ Marieb, Elaine Nicpon, 1936- (2018). Lidská anatomie a fyziologie. Hoehn, Katja (jedenácté vydání). [Hoboken, New Jersey]. ISBN 978-0-13-458099-9. OCLC 1004376412.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Yuan X, Lu ML, Li T, Balk SP (prosinec 2001). „SRY interaguje s a negativně reguluje transkripční aktivitu androgenového receptoru“. The Journal of Biological Chemistry. 276 (49): 46647–54. doi:10,1074 / jbc.M108404200. PMID 11585838.
- ^ Lister Hill National Center for Biomedical Communications (2008). "Syndrom necitlivosti na androgen". Genetická domácí reference. Americká národní lékařská knihovna.
- ^ Dewing P, Chiang CW, Sinchak K, Sim H, Fernagut PO, Kelly S, Chesselet MF, Micevych PE, Albrecht KH, Harley VR, Vilain E (únor 2006). „Přímá regulace funkce mozku dospělých pomocí faktoru SRY specifického pro muže“. Aktuální biologie. 16 (4): 415–20. doi:10.1016 / j.cub.2006.01.017. PMID 16488877. S2CID 5939578.
- ^ A b Herbarth B, Pingault V, Bondurand N, Kuhlbrodt K, Hermans-Borgmeyer I, Puliti A, Wegner M (1998). „Mutace genu Sox10 souvisejícího se Sry v dominujícím megakolonu, myší model pro lidskou Hirschsprungovu chorobu“. Sborník Národní akademie věd. 95 (9): 5161–5165. Bibcode:1998PNAS ... 95.5161H. doi:10.1073 / pnas.95.9.5161. PMC 20231. PMID 9560246.
- ^ A b Pritchett J, Athwal V, Roberts N, Hanley NA, Hanley KP (2011). „Pochopení role SOX9 v získaných chorobách: poučení z vývoje“. Trendy v molekulární medicíně. 17 (3): 166–174. doi:10.1016 / j.molmed.2010.12.001. PMID 21237710.
- ^ „OMIM Entry - # 114290 - CAMPOMELIC DYSPLASIA“. omim.org. Citováno 29. února 2020.
- ^ „Olympic Gender Testing“.
- ^ A b Facius GM (1. srpna 2004). „Hlavní lékařská chyba 20. století“. Testování pohlaví. facius-homepage.dk. Archivovány od originál dne 26. ledna 2010. Citováno 12. června 2011.
- ^ Elsas LJ, Ljungqvist A, Ferguson-Smith MA, Simpson JL, Genel M, Carlson AS, Ferris E, de la Chapelle A, Ehrhardt AA (2000). „Ověření pohlaví atletek“. Genetika v medicíně. 2 (4): 249–54. doi:10.1097/00125817-200007000-00008. PMID 11252710.
- ^ Dickinson BD, Genel M, Robinowitz CB, Turner PL, Woods GL (říjen 2002). "Ověření pohlaví olympijských atletek". Medicína a věda ve sportu a cvičení. 34 (10): 1539–42, diskuse 1543. doi:10.1097/00005768-200210000-00001. PMID 12370551.
- ^ „Předpisy MOV o ženském hyperandrogenismu“ (PDF). Mezinárodní olympijský výbor. 22. června 2012. Archivováno (PDF) z původního dne 13. srpna 2012. Citováno 9. srpna 2012.
Další čtení
- Haqq CM, King CY, Ukiyama E, Falsafi S, Haqq TN, Donahoe PK, Weiss MA (prosinec 1994). „Molekulární podstata sexuálního stanovení savců: aktivace genové exprese látky inhibující Müllerian pomocí SRY“. Věda. 266 (5190): 1494–500. Bibcode:1994Sci ... 266.1494H. doi:10.1126 / science.7985018. PMID 7985018.
- Goodfellow PN, Lovell-Badge R (1993). "SRY a určení pohlaví u savců". Výroční přehled genetiky. 27: 71–92. doi:10.1146 / annurev.ge.27.120193.000443. PMID 8122913.
- Hawkins JR (1993). "Mutační analýza SRY u XY žen". Lidská mutace. 2 (5): 347–50. doi:10,1002 / humu.1380020504. PMID 8257986.
- Harley VR (2002). "Molekulární působení faktorů určujících varlata SRY a SOX9". Genetika a biologie stanovení pohlaví. Nalezen Novartis. Symp. Symposia Novartis Foundation. 244. str. 57–66, diskuse 66–7, 79–85, 253–7. doi:10.1002 / 0470868732.ch6. ISBN 978-0-470-86873-7. PMID 11990798.
- Jordan BK, Vilain E (2003). „Sry a genetika determinace pohlaví“. Přiřazení pediatrického pohlaví. Adv. Exp. Med. Biol. Pokroky v experimentální medicíně a biologii. 511. s. 1–13, diskuse 13–4. doi:10.1007/978-1-4615-0621-8_1. ISBN 978-1-4613-5162-7. PMID 12575752.
- Oh HJ, Lau YF (březen 2006). "KRAB: partner pro SRY působení na chromatin". Molekulární a buněčná endokrinologie. 247 (1–2): 47–52. doi:10.1016 / j.mce.2005.12.011. PMID 16414182. S2CID 19870331.
- Polanco JC, Koopman P (únor 2007). „Sry a váhavé počátky mužského vývoje“. Vývojová biologie. 302 (1): 13–24. doi:10.1016 / j.ydbio.2006.08.049. PMID 16996051.
- Hawkins JR, Taylor A, Berta P, Levilliers J, Van der Auwera B, Goodfellow PN (únor 1992). "Mutační analýza SRY: nesmysly a missense mutace v obrácení pohlaví XY". Genetika člověka. 88 (4): 471–4. doi:10.1007 / BF00215684. PMID 1339396. S2CID 9332496.
- Hawkins JR, Taylor A, Goodfellow PN, Migeon CJ, Smith KD, Berkovitz GD (listopad 1992). "Důkazy o zvýšené prevalenci mutací SRY u žen XY s úplnou spíše než částečnou gonadální dysgenezí". American Journal of Human Genetics. 51 (5): 979–84. PMC 1682856. PMID 1415266.
- Ferrari S, Harley VR, Pontiggia A, Goodfellow PN, Lovell-Badge R, Bianchi ME (prosinec 1992). „SRY, jako HMG1, rozeznává ostré úhly v DNA“. Časopis EMBO. 11 (12): 4497–506. doi:10.1002 / j.1460-2075.1992.tb05551.x. PMC 557025. PMID 1425584.
- Jäger RJ, Harley VR, Pfeiffer RA, Goodfellow PN, Scherer G (prosinec 1992). "Rodinná mutace v genu určujícím varlata SRY sdílená oběma pohlavími". Genetika člověka. 90 (4): 350–5. doi:10.1007 / BF00220457. PMID 1483689. S2CID 19470332.
- Vilain E, McElreavey K, Jaubert F, Raymond JP, Richaud F, Fellous M (květen 1992). "Rodinný případ se sekvenční variantou v oblasti určující varlat související se dvěma pohlavními fenotypy". American Journal of Human Genetics. 50 (5): 1008–11. PMC 1682588. PMID 1570829.
- Müller J, Schwartz M, Skakkebaek NE (červenec 1992). „Analýza oblasti určující pohlaví chromozomu Y (SRY) u pacientů s reverzním pohlavím: bodová mutace v SRY způsobující reverzi pohlaví u 46, XY žen“. The Journal of Clinical Endocrinology and Metabolism. 75 (1): 331–3. doi:10.1210 / jc.75.1.331. PMID 1619028.
- McElreavey KD, Vilain E, Boucekkine C, Vidaud M, Jaubert F, Richaud F, Fellous M (červenec 1992). "XY obrácení pohlaví spojené s nesmyslnou mutací v SRY". Genomika. 13 (3): 838–40. doi:10.1016 / 0888-7543 (92) 90164-N. PMID 1639410.
- Sinclair AH, Berta P, Palmer MS, Hawkins JR, Griffiths BL, Smith MJ, Foster JW, Frischauf AM, Lovell-Badge R, Goodfellow PN (červenec 1990). „Gen z oblasti určující lidské pohlaví kóduje protein s homologií se zachovaným motivem vázajícím DNA“. Příroda. 346 (6281): 240–4. Bibcode:1990 Natur.346..240S. doi:10.1038 / 346240a0. PMID 1695712. S2CID 4364032.
- Berkovitz GD, Fechner PY, Zacur HW, Rock JA, Snyder HM, Migeon CJ, Perlman EJ (listopad 1991). „Klinické a patologické spektrum 46, XY gonadální dysgeneze: její význam pro pochopení diferenciace pohlaví“. Lék. 70 (6): 375–83. doi:10.1097/00005792-199111000-00003. PMID 1956279. S2CID 37972412.
- Berta P, Hawkins JR, Sinclair AH, Taylor A, Griffiths BL, Goodfellow PN, Fellous M (listopad 1990). "Genetický důkaz, který se rovná SRY a faktoru určujícím varlata". Příroda. 348 (6300): 448–50. Bibcode:1990 Natur.348..448B. doi:10.1038 / 348448A0. PMID 2247149. S2CID 3336314.
- Jäger RJ, Anvret M, hala K, Scherer G (listopad 1990). „Lidská žena XY s mutací posunu rámce v kandidátním genu určujícím varlata SRY“. Příroda. 348 (6300): 452–4. Bibcode:1990 Natur.348..452J. doi:10.1038 / 348452a0. PMID 2247151. S2CID 4326539.
- Ellis NA, Goodfellow PJ, Pym B, Smith M, Palmer M, Frischauf AM, Goodfellow PN (leden 1989). „Hranice pseudoautosomů u člověka je definována Alu opakovací sekvencí vloženou do chromozomu Y“. Příroda. 337 (6202): 81–4. Bibcode:1989 Natur.337 ... 81E. doi:10.1038 / 337081a0. PMID 2909893. S2CID 2890077.
- Whitfield LS, Hawkins TL, Goodfellow PN, Sulston J (květen 1995). „41 kilobází analyzované sekvence z pseudoautosomálních a pohlaví určujících oblastí krátkého ramene lidského chromozomu Y“. Genomika. 27 (2): 306–11. doi:10.1006 / geno.1995.1047. PMID 7557997.
externí odkazy
- GeneReviews / NCBI / NIH / UW vstup na 46, XX Testikulární porucha sexuálního vývoje
- OMIM entries on 46, XX Testicular Disorder of Sex Development
- Geny, + sry v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- Určení pohlaví + Region + Y + Protein v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- PDBe-KB poskytuje přehled všech strukturních informací dostupných v PDB pro protein Y oblasti určující lidský sex
Galerie PDB | |
|---|---|
|