Chuťový receptor - Taste receptor
| Chuťový receptor | |
|---|---|
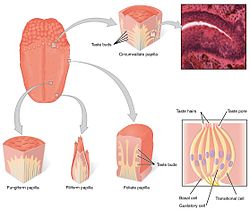 Chuťové receptory jazyka jsou přítomny v chuťových pohárcích papil. | |
| Identifikátory | |
| FMA | 84662 |
| Anatomická terminologie | |
A chuťový receptor je typ receptor což usnadňuje vnímání chuť. Když potraviny nebo jiné látky vstupují do úst, molekuly interagují se slinami a jsou vázány na receptory chuti v ústní dutině a na dalších místech. Molekuly, které dávají pocit chuti, jsou považovány za „sapidové“.[1]
Chuťové receptory jsou rozděleny do dvou rodin:[Citace je zapotřebí ]
- Typ 1, bonbón, poprvé charakterizováno v roce 2001:[2] TAS1R2 – TAS1R3
- Typ 2, hořký, poprvé charakterizováno v roce 2000:[3] U lidí je 25 známých různých hořkých receptorů, u koček 12, u kuřat tři a u myší 35 známých různých hořkých receptorů.[4]
Vizuální, olfaktivní, „sapiktivní“ (vnímání chutí), trigeminální (horké, chladné), mechanické, všechny přispívají k vnímání chuť. Z nich přechodný receptorový potenciální kationtový kanál podčeleď V člen 1 (TRPV1 ) vaniloidní receptory jsou odpovědné za vnímání tepla z některých molekul, jako je kapsaicin, a CMR1 receptor je zodpovědný za vnímání chladu z molekul jako např mentol, eukalyptol, a icilin.[1]
Distribuce tkání
The chuťový systém sestává z buněk chuťových receptorů v chuťové pohárky. Chuťové pohárky jsou zase obsaženy ve strukturách nazývaných papily. Chuť zahrnuje tři druhy papil: fungiformní papily, listové papily, a obklopit papily. (Čtvrtý typ - vláknité papily neobsahují chuťové pohárky). Kromě papil jsou receptory chuti také v patro a rané části zažívací ústrojí jako hrtan a horní jícen. Tam jsou tři lebeční nervy které inervují jazyk; the bludný nerv, glossofaryngeální nerv a lícní nerv. The glossofaryngeální nerv a chorda tympani pobočka lícní nerv inervujte chuťové receptory TAS1R a TAS2R. Vedle chuťových receptorů v jazyku je střevní epitel také vybaven jemným chemosenzorickým systémem, který sděluje senzorické informace několika efektorovým systémům zapojeným do regulace chuti k jídlu, imunitních odpovědí a gastrointestinální motility. [5]
V roce 2010 našli vědci hořké receptory v plicní tkáni, které způsobují uvolnění dýchacích cest, když dojde k hořké látce. Věří, že tento mechanismus je evolučně adaptivní, protože pomáhá léčit plicní infekce, ale lze jej také využít k léčbě astma a chronická obstrukční plicní nemoc.[6]
Funkce
Chuť pomáhá identifikovat toxiny udržovat výživa a regulují chuť k jídlu, imunitní odpovědi a gastrointestinální motilitu.[5] Dnes je známo pět základních chutí: slaná, sladká, hořká, kyselá a umami. Slané i kyselé chuťové vjemy jsou detekovány prostřednictvím iontové kanály. Sladké, hořké a umami chutě jsou však detekovány způsobem Vazba na G protein chuťové receptory.[7]
Někteří agenti mohou navíc fungovat jako modifikátory chuti, tak jako miraculin nebo kurculin pro sladké nebo sterubin na maska hořká.
Mechanismus účinku
Standardní hořký, sladký nebo umami chuťový receptor je a Receptor spojený s G proteinem se sedmi transmembránové domény. Aktivuje se vazba ligandu na receptory chuti druhý posel kaskády k depolarizaci chuťové buňky. Gustducin je nejběžnější podjednotkou chuti Gα, která má hlavní roli v recepci hořké chuti TAS2R. Gustducin je homolog pro transducin, G-protein podílející se na transdukci zraku.[8] Kromě toho chuťové receptory sdílejí použití iontového kanálu TRPM5, stejně jako fosfolipázy PLCβ2.[9]
Pikantní nebo glutamáty (Umami)
The TAS1R1 +TAS1R3 heterodimerový receptor funguje jako receptor umami, reaguje na L-aminokyselina vázání, zejména L-glutamát.[2] The umami chuť je nejčastěji spojována s potravinářskou přídatnou látkou glutaman sodný (MSG) a lze je vylepšit vazbou inosin monofosfát (IMP) a guanosinmonofosfát (GMP) molekuly.[10][11] Buňky exprimující TAS1R1 + 3 se nacházejí většinou v fungiformní papily na špičce a okrajích jazyka a chuťových buněk patra ve střeše úst.[2] Ukázalo se, že tyto buňky se synchronizují s chorda tympani nervy posílat své signály do mozku, i když určitá aktivace glossofaryngeální nerv bylo nalezeno.[10][12]
Alternativní kandidátské receptory chuti umami zahrnují sestřihové varianty metabotropních glutamátových receptorů, mGluR4 a mGluR1 a N-methyl-D-aspartát receptor glutamátového iontového kanálu typu.[7][13][14][15]
Bonbón
The TAS1R2 +TAS1R3 heterodimerový receptor funguje jako bonbón receptor vazbou na širokou škálu cukry a náhražky cukru.[2][16] Buňky exprimující TAS1R2 + 3 se nacházejí v obklopit papily a listové papily v zadní části jazyk a patro buňky chuťových receptorů ve střeše úst.[2] Tyto buňky jsou zobrazeny synapse na chorda tympani a glossofaryngeální nervy vysílat jejich signály do mozku.[7][12] The TAS1R3 homodimer také funguje jako sladký receptor podobně jako TAS1R2 + 3, ale má sníženou citlivost na sladké látky. Přírodní cukry snáze detekuje TAS1R3 receptor než náhražky cukru. To může pomoci vysvětlit, proč mají cukr a umělá sladidla různé chutě.[17] Genetické polymorfismy v TAS1R3 částečně vysvětlují rozdíl ve vnímání sladké chuti a spotřebě cukru mezi lidmi afroamerických předků a lidmi evropských a asijských předků.[18][19]
Hořký
Proteiny TAS2R (InterPro: IPR007960 ) fungují jako hořký chuťové receptory.[20] Existuje 43 lidí TAS2R geny, z nichž každý (kromě pěti pseudogeny ) chybí introny a kódy pro a GPCR protein.[7] Tyto proteiny, na rozdíl od proteinů TAS1R, mají krátké extracelulární domény a jsou umístěny v obklopit papily, patro, listové papily, a epiglottis chuťové pohárky, se sníženým projevem v fungiformní papily.[3][7] Ačkoli je jisté, že více TAS2R je exprimováno v jedné buňce chuťového receptoru, stále se diskutuje o tom, zda savci mohou rozlišovat mezi chutěmi různých hořkých ligandy.[3][7] Určité překrývání se však musí objevit, protože existuje mnohem více hořkých sloučenin než genů TAS2R. Mezi běžné hořké ligandy patří cykloheximid, denatonium, PROP (6-n-propyl-2-thiouracil ), PTC (fenylthiokarbamid ) a β-glukopyranosidy.[7]
Transdukce signálu hořkých podnětů je dosaženo pomocí α-podjednotky gustducin. Tato podjednotka G proteinu aktivuje chuť fosfodiesteráza a klesá cyklický nukleotid úrovně. Další kroky v transdukční dráze nejsou dosud známy. Βγ-podjednotka gustducinu také zprostředkovává chuť aktivací IP3 (inositol trifosfát ) a DAG (diglycerid ). Tito druzí poslové mohou otevírat bránové iontové kanály nebo mohou způsobit uvolnění vnitřních vápník.[21] Ačkoli jsou všechny TAS2R umístěny v buňkách obsahujících gustducin, vyřazení gustducinu úplně nezruší citlivost na hořké sloučeniny, což naznačuje nadbytečný mechanismus hořké ochutnávky[9] (nepřekvapivé vzhledem k tomu, že hořká chuť obecně signalizuje přítomnost a toxin ).[9] Jeden navrhovaný mechanismus hořké ochutnávky nezávislé na gustducinu je prostřednictvím interakce iontových kanálů specifickými hořkými ligandy, podobně jako interakce iontových kanálů, ke které dochází při ochutnávání kyselých a slaných podnětů.[7]
Jeden z nejlépe prozkoumaných proteinů TAS2R je TAS2R38, což přispívá k ochutnávání PROP i PTC. Je to první chuťový receptor, jehož polymorfismy je prokázáno, že jsou odpovědné za rozdíly ve vnímání chuti. Současné studie se zaměřují na stanovení dalších takových polymorfismů určujících chuťový fenotyp.[7] Novější studie ukazují, že genetické polymorfismy v jiných genech receptorů hořké chuti ovlivňují vnímání hořké chuti kofeinu, chininu a denatoniumbenzoátu.[22]
Kyselý
Historicky se předpokládalo, že kyselá chuť se vyrábí pouze tehdy, když je zdarma vodíkové ionty (H+) přímo depolarizované chuťové receptory. Nyní se však navrhují specifické receptory pro kyselou chuť s jinými způsoby působení. HCN1 a HCN4 (Kanály HCN ) byly dva takové návrhy; oba tyto receptory jsou kanály řízené cyklickými nukleotidy. Dva iontové kanály, které mají přispívat ke kyselé chuti, jsou ACCN1 a ÚKOL-1.
Sůl
Byly také navrženy různé receptory pro slanou chuť spolu s možnou detekcí chuti lipidů, komplexních sacharidů a vody. Důkazy pro tyto receptory jsou však přinejlepším nejisté a ve studiích na savcích často nepřesvědčivé. Například navrhovaný ENaC lze prokázat, že receptor pro detekci sodíku přispívá pouze k chuti sodíku v Drosophilia.[7]
Karbonatace
Enzym připojený k kyselému receptoru přenáší informace o sycené vodě.[23]
Tlustý
Možný chuťový receptor pro tuk, CD36, byl identifikován.[24] CD36 byl lokalizován do cirkumvalátu a folátu papily, které jsou přítomny v chuťové pohárky[25] a kde jazyková lipáza je produkován a výzkum ukázal, že receptor CD36 váže dlouhý řetězec mastné kyseliny.[26] Rozdíly v množství CD36 výraz u lidských subjektů byla spojena s jejich schopností ochutnat tuky,[27] vytvoření argumentu pro vztah receptoru k ochutnávání tuků. Další výzkum receptoru CD36 by mohl být užitečný při určování existence skutečného receptoru chutnajícího tuky.
GPR120 a GPR40 byly zapojeny do reakce na orální tuk,[28] a jejich absence vede ke snížené preferenci tuků a snížené neuronální odpovědi na orálně podávané mastné kyseliny.[29]
TRPM5 Bylo prokázáno, že se účastní orální tukové odpovědi a je identifikován jako možný orální tukový receptor, ale nedávné důkazy ji uvádějí jako primárně následného hráče.[30][31]
Typy
Geny pro receptory hořké chuti se nazývají TAS2R1 až TAS2R64, s mnoha mezerami kvůli neexistujícím genům, pseudogenům nebo navrhovaným genům, které nebyly anotovány k nejnovějšímu shromáždění lidského genomu. Mnoho genů receptorů hořké chuti má také matoucí názvy synonym s několika různými názvy genů odkazujících na stejný gen. V následující tabulce najdete úplný seznam genů receptorů lidské hořké chuti:
| Třída | Gen | Synonyma | Aliasy | Místo | Popis |
|---|---|---|---|---|---|
| typ 1 (bonbón) | TAS1R1 | GPR70 | 1p36.23 | ||
| TAS1R2 | GPR71 | 1p36.23 | |||
| TAS1R3 | 1p36 | ||||
| typ 2 (hořký) | TAS2R1 | 5p15 | |||
| TAS2R2 | 7p21.3 | pseudogen | |||
| TAS2R3 | 7q31.3-q32 | ||||
| TAS2R4 | 7q31.3-q32 | ||||
| TAS2R5 | 7q31.3-q32 | ||||
| TAS2R6 | 7 | není anotován v sestavě lidského genomu | |||
| TAS2R7 | 12p13 | ||||
| TAS2R8 | 12p13 | ||||
| TAS2R9 | 12p13 | ||||
| TAS2R10 | 12p13 | ||||
| TAS2R11 | chybí u lidí | ||||
| TAS2R12 | TAS2R26 | 12p13.2 | pseudogen | ||
| TAS2R13 | 12p13 | ||||
| TAS2R14 | 12p13 | ||||
| TAS2R15 | 12p13.2 | pseudogen | |||
| TAS2R16 | 7q31.1-q31.3 | ||||
| TAS2R17 | chybí u lidí | ||||
| TAS2R18 | 12p13.2 | pseudogen | |||
| TAS2R19 | TAS2R23, TAS2R48 | 12p13.2 | |||
| TAS2R20 | TAS2R49 | 12p13.2 | |||
| TAS2R21 | chybí u lidí | ||||
| TAS2R22 | 12 | není anotován v sestavě lidského genomu | |||
| TAS2R24 | chybí u lidí | ||||
| TAS2R25 | chybí u lidí | ||||
| TAS2R27 | chybí u lidí | ||||
| TAS2R28 | chybí u lidí | ||||
| TAS2R29 | chybí u lidí | ||||
| TAS2R30 | TAS2R47 | 12p13.2 | |||
| TAS2R31 | TAS2R44 | 12p13.2 | |||
| TAS2R32 | chybí u lidí | ||||
| TAS2R33 | 12 | není anotován v sestavě lidského genomu | |||
| TAS2R34 | chybí u lidí | ||||
| TAS2R35 | chybí u lidí | ||||
| TAS2R36 | 12 | není anotován v sestavě lidského genomu | |||
| TAS2R37 | 12 | není anotován v sestavě lidského genomu | |||
| TAS2R38 | 7q34 | ||||
| TAS2R39 | 7q34 | ||||
| TAS2R40 | GPR60 | 7q34 | |||
| TAS2R41 | 7q34 | ||||
| TAS2R42 | 12p13 | ||||
| TAS2R43 | 12p13.2 | ||||
| TAS2R45 | GPR59 | 12 | |||
| TAS2R46 | 12p13.2 | ||||
| TAS2R50 | TAS2R51 | 12p13.2 | |||
| TAS2R52 | chybí u lidí | ||||
| TAS2R53 | chybí u lidí | ||||
| TAS2R54 | chybí u lidí | ||||
| TAS2R55 | chybí u lidí | ||||
| TAS2R56 | chybí u lidí | ||||
| TAS2R57 | chybí u lidí | ||||
| TAS2R58 | chybí u lidí | ||||
| TAS2R59 | chybí u lidí | ||||
| TAS2R60 | 7 | ||||
| TAS2R62P | 7q34 | pseudogen | |||
| TAS2R63P | 12p13.2 | pseudogen | |||
| TAS2R64P | 12p13.2 | pseudogen |
Ztráta funkce
U mnoha druhů vykazovaly chuťové receptory ztrátu funkcí. Evoluční proces, jehož chuťové receptory ztratily svoji funkci, se považuje za adaptivní evoluci, která je spojena s ekologií krmení, aby se podpořila specializace a rozdvojení chuťových receptorů.[32] Ze všech chuťových receptorů je prokázáno, že hořké, sladké a umami mají korelaci mezi inaktivací chuťových receptorů a chováním při krmení.[32] Neexistují však žádné silné důkazy, které by podporovaly stavovce, chybí geny receptorů hořké chuti.[32]
Receptor sladké chuti je jedním z chuťových receptorů, kde došlo ke ztrátě funkce. U savců je převládajícím receptorem sladké chuti receptor chuti typu 1 Tas1r2 / Tas1r3.[33] Některé druhy savců, jako jsou kočky a upíři, prokázaly neschopnost sladce chutnat.[33] U těchto druhů je příčinou ztráty funkce sladkého receptoru pseudogenizace Tas1r2.[33] Pseudogenizace Tas1r2 je také pozorována u druhů, které nejsou savci, jako jsou kuřata a západní bezocasá žába, a tyto druhy také ukazují neschopnost chutnat sladce.[33] Pseudogenizace Tas1r2 je rozšířená a nezávislá v pořadí Carnivora.[33] Mnoho studií ukázalo, že pseudogenizace chuťových receptorů je způsobena škodlivou mutací v otevřených čtecích rámcích (ORF).[34] Ve studii bylo zjištěno, že u nefelšinových masožravých druhů tyto druhy vykazovaly mutace Tas1r2 narušující ORF a vyskytovaly se mezi druhy nezávisle.[33] Rovněž vykazovaly vysokou variabilitu v jejich liniích.[33] Předpokládá se, že k pseudogenizaci Tas1r2 došlo konvergentní evolucí, kdy masožravé druhy ztratily schopnost chutnat sladce kvůli dietnímu chování.[33]
Umami je také chuťovým receptorem, kde se funkce u mnoha druhů ztratila. Převládající receptory chuti umami jsou Tas1r1 / Tas1r3.[33] Ve dvou liniích vodních savců, včetně delfínů a lachtanů, bylo zjištěno, že Tas1r1 je pseudogenizovaný.[33] Pseudogenizace Tas1r1 byla také nalezena u suchozemských masožravých druhů.[33] Zatímco panda patří do řádu Carnivora, je býložravá, kde 99% její stravy tvoří bambus, a nemůže ochutnat umami.[35] Sekvence genomu pandy ukazuje, že její gen Tas1r1 je pseudogenizovaný.[35] Ve studii bylo zjištěno, že u všech druhů v pořadí Carnivora kromě pandy byl zachován otevřený čtecí rámec.[35] V pandě bylo zjištěno, že poměr nesynonymních a synonymních substitucí je mnohem vyšší než u jiných druhů, aby byl Carnivora.[35] Tato data korelují s daty fosilních záznamů pandy, která ukazují, kde panda přešla ze stravy masožravců na býložravce.[33] Proto se předpokládá, že ztráta funkce umami u pandy je způsobena změnou stravy, kde se panda stala méně závislou na mase.[33] Tyto studie však nevysvětlují býložravce, jako jsou koně a krávy, kteří si uchovali receptor Tas1r1.[35]
Celkově je ztráta funkce chuťového receptoru evolučním procesem, ke kterému došlo v důsledku dietní změny druhů.[34]
Reference
- ^ A b To, Hervé (2012). „Věda o troubě - výňatek z kapitoly 1“. Citováno 30.dubna 2014.
- ^ A b C d E Nelson G, Hoon MA, Chandrashekar J, Zhang Y, Ryba NJ, Zuker CS (srpen 2001). "Savčí receptory sladké chuti". Buňka. 106 (3): 381–90. doi:10.1016 / S0092-8674 (01) 00451-2. PMID 11509186.
- ^ A b C Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba NJ, Zuker CS (březen 2000). "Nová rodina savčích chuťových receptorů". Buňka. 100 (6): 693–702. doi:10.1016 / S0092-8674 (00) 80705-9. PMID 10761934.
- ^ http://bitterdb.agri.huji.ac.il/dbbitter.php#receptorBrowse
- ^ A b Steensels S, Depoortere I (2018). "Chemoreceptory ve střevě". Roční přehled fyziologie. 80: 117–141. doi:10,1146 / annurev-physiol-021317-121332. PMID 29029594.
- ^ Deshpande DA, Wang WC, McIlmoyle EL, Robinett KS, Schillinger RM, An SS, Sham JS, Liggett SB (listopad 2010). „Receptory hořké chuti na bronchodilatu hladkého svalstva dýchacích cest lokalizovanou signalizací vápníku a reverzní obstrukcí“. Přírodní medicína. 16 (11): 1299–304. doi:10,1038 / nm.2237. PMC 3066567. PMID 20972434.
- ^ A b C d E F G h i j Bachmanov AA, Beauchamp GK (2007). „Ochutnejte geny receptoru“. Každoroční přehled výživy. 27: 389–414. doi:10.1146 / annurev.nutr.26.061505.111329. PMC 2721271. PMID 17444812.
- ^ Sainz E, Cavenagh MM, LopezJimenez ND, Gutierrez JC, Battey JF, Northup JK, Sullivan SL (červen 2007). "G-proteinové vazebné vlastnosti lidských sladkých a aminokyselinových chuťových receptorů". Vývojová neurobiologie. 67 (7): 948–59. doi:10.1002 / dneu.20403. PMID 17506496.
- ^ A b C Zhang Y, Hoon MA, Chandrashekar J, Mueller KL, Cook B, Wu D, Zuker CS, Ryba NJ (únor 2003). "Kódování sladkého, hořkého a umami vkusu: různé receptorové buňky sdílející podobné signální dráhy". Buňka. 112 (3): 293–301. doi:10.1016 / S0092-8674 (03) 00071-0. PMID 12581520.
- ^ A b Nelson G, Chandrashekar J, Hoon MA, Feng L, Zhao G, Ryba NJ, Zuker CS (březen 2002). "Aminokyselinový chuťový receptor". Příroda. 416 (6877): 199–202. Bibcode:2002 Natur.416..199N. doi:10.1038 / příroda726. PMID 11894099.
- ^ Delay ER, Beaver AJ, Wagner KA, Stapleton JR, Harbaugh JO, Catron KD, Roper SD (říjen 2000). „Synergie preferencí chuti mezi agonisty receptoru glutamátu a inosinmonofosfátem u potkanů“. Chemické smysly. 25 (5): 507–15. doi:10.1093 / chemse / 25.5.507. PMID 11015322.
- ^ A b Danilova V, Hellekant G (březen 2003). „Srovnání odpovědí chorda tympani a glossofaryngeálních nervů na chuťové podněty u myší C57BL / 6J“. BMC Neuroscience. 4: 5. doi:10.1186/1471-2202-4-5. PMC 153500. PMID 12617752.
- ^ Značka JG (duben 2000). „Receptorové a transdukční procesy pro chuť umami“. The Journal of Nutrition. 130 (4S Suppl): 942S – 5S. doi:10.1093 / jn / 130.4.942S. PMID 10736357.
- ^ Chaudhari N, Landin AM, Roper SD (únor 2000). "Varianta metabotropního glutamátového receptoru funguje jako chuťový receptor". Přírodní neurovědy. 3 (2): 113–9. doi:10.1038/72053. PMID 10649565.
- ^ Toyono T, Seta Y, Kataoka S, Kawano S, Shigemoto R, Toyoshima K (červenec 2003). "Exprese skupiny metabotropního glutamátového receptoru I v chuťových papilách potkanů". Výzkum buněk a tkání. 313 (1): 29–35. doi:10.1007 / s00441-003-0740-2. PMID 12898387.
- ^ Li X, Staszewski L, Xu H, Durick K, Zoller M, Adler E (duben 2002). "Lidské receptory pro sladkou a umami chuť". Sborník Národní akademie věd Spojených států amerických. 99 (7): 4692–6. Bibcode:2002PNAS ... 99.4692L. doi:10.1073 / pnas.072090199. PMC 123709. PMID 11917125.
- ^ Zhao GQ, Zhang Y, Hoon MA, Chandrashekar J, Erlenbach I, Ryba NJ, Zuker CS (říjen 2003). "Receptory pro savčí sladkou a umami chuť". Buňka. 115 (3): 255–66. doi:10.1016 / S0092-8674 (03) 00844-4. PMID 14636554.
- ^ Hwang LD, Lin C, Gharahkhani P, Cuellar-Partida G, Ong JS, An J, Gordon SD, Zhu G, MacGregor S, Lawlor DA, Breslin PA, Wright MJ, Martin NG, Reed DR (duben 2019). „Nový pohled na lidskou sladkou chuť: celogenomová asociační studie vnímání a příjmu sladkých látek“. American Journal of Clinical Nutrition. 109: 1724–1737. doi:10.1093 / ajcn / nqz043. PMC 6537940. PMID 31005972.
- ^ Yousif, Ragheed (březen 2020). „Zkoumání molekulárních interakcí mezi neokulinem a receptory lidské chuti na sladké pomocí výpočetních přístupů“ (PDF). Sains Malaysiana. 49: 517. doi:10.17576 / jsm-2020-4903-06.
- ^ Chandrashekar J, Mueller KL, Hoon MA, Adler E, Feng L, Guo W, Zuker CS, Ryba NJ (březen 2000). Msgstr "T2R fungují jako receptory hořké chuti". Buňka. 100 (6): 703–11. doi:10.1016 / S0092-8674 (00) 80706-0. PMID 10761935.
- ^ Margolskee RF (leden 2002). "Molekulární mechanismy přenosu hořké a sladké chuti". The Journal of Biological Chemistry. 277 (1): 1–4. doi:10,1074 / jbc.R100054200. PMID 11696554.
- ^ Hwang LD, Gharahkhani P, Breslin PA, Gordon SD, Zhu G, Martin NG, Reed DR, Wright MJ (září 2018). „Bivariační genomová asociační analýza posiluje roli shluků hořkých receptorů na chromozomech 7 a 12 v lidské hořké chuti“. BMC Genomics. 19 (1): 678. doi:10.1186 / s12864-018-5058-2. PMC 6142396. PMID 30223776.
- ^ „Archivovaná kopie“. Archivovány od originál dne 3. 7. 2015. Citováno 2014-10-06.CS1 maint: archivovaná kopie jako titul (odkaz)
- ^ Laugerette F, Passilly-Degrace P, Patris B, Niot I, Febbraio M, Montmayeur JP, Besnard P (listopad 2005). „Zapojení CD36 do orosenzorické detekce dietních lipidů, preference spontánního tuku a sekrece trávicího traktu“. The Journal of Clinical Investigation. 115 (11): 3177–84. doi:10,1172 / JCI25299. PMC 1265871. PMID 16276419.
- ^ Simons PJ, Kummer JA, Luiken JJ, Boon L (prosinec 2011). "Apikální CD36 imunolokalizace v lidských a prasečích chuťových pohárcích z cirkumvalátových a listnatých papil". Acta Histochemica. 113 (8): 839–43. doi:10.1016 / j.acthis.2010.08.006. PMID 20950842.
- ^ Baillie AG, Coburn CT, Abumrad NA (září 1996). "Reverzibilní vazba mastných kyselin s dlouhým řetězcem na čištěný FAT, homolog tukového CD36". The Journal of Membrane Biology. 153 (1): 75–81. doi:10.1007 / s002329900111. PMID 8694909.
- ^ Pepino MY, Love-Gregory L, Klein S, Abumrad NA (březen 2012). „Translokázový gen mastných kyselin CD36 a lingvální lipáza ovlivňují orální citlivost na tuk u obézních jedinců“. Journal of Lipid Research. 53 (3): 561–6. doi:10.1194 / ml. M021873. PMC 3276480. PMID 22210925.
- ^ DiPatrizio NV (září 2014). „Je tuková chuť připravena na hlavní čas?“. Fyziologie a chování. 136: 145–54. doi:10.1016 / j.physbeh.2014.03.002. PMC 4162865. PMID 24631296.
- ^ Cartoni C, Yasumatsu K, Ohkuri T, Shigemura N, Yoshida R, Godinot N, le Coutre J, Ninomiya Y, Damak S (červen 2010). „Preference chuti mastných kyselin je zprostředkována GPR40 a GPR120“ (PDF). The Journal of Neuroscience. 30 (25): 8376–82. doi:10.1523 / JNEUROSCI.0496-10.2010. PMC 6634626. PMID 20573884.
- ^ Mattes RD (září 2011). „Hromadící se důkazy podporují chuťovou složku volných mastných kyselin u lidí“. Fyziologie a chování. 104 (4): 624–31. doi:10.1016 / j.physbeh.2011.05.002. PMC 3139746. PMID 21557960.
- ^ Liu P, Shah BP, Croasdell S, Gilbertson TA (červen 2011). „Přechodný receptorový potenciální kanál typu M5 je nezbytný pro chuť tuku“. The Journal of Neuroscience. 31 (23): 8634–42. doi:10.1523 / JNEUROSCI.6273-10.2011. PMC 3125678. PMID 21653867.
- ^ A b C Feng P, Zhao H (červen 2013). „Komplexní evoluční historie genů receptoru pro chuť sladké / umami obratlovců“. Bulletin čínské vědy. 58 (18): 2198–2204. Bibcode:2013ChSBu..58.2198F. doi:10.1007 / s11434-013-5811-5.
- ^ A b C d E F G h i j k l m Jiang P, Josue J, Li X, Glaser D, Li W, Brand JG, Margolskee RF, Reed DR, Beauchamp GK (březen 2012). „Velká ztráta chuti u masožravých savců“. Sborník Národní akademie věd Spojených států amerických. 109 (13): 4956–61. doi:10.1073 / pnas.1118360109. PMC 3324019. PMID 22411809.
- ^ A b Antinucci M, Risso D (2017-11-28). „Záležitost chuti: Ztráta funkce genů receptoru chuti u obratlovců specifická pro daný rod“. Frontiers in Molecular Biosciences. 4: 81. doi:10.3389 / fmolb.2017.00081. PMC 5712339. PMID 29234667.
- ^ A b C d E Zhao H, Yang JR, Xu H, Zhang J (prosinec 2010). „Pseudogenizace genu pro chuťový receptor umami Tas1r1 u pandy obrovské se shodovala s jejím přechodem ze stravy na bambus“. Molekulární biologie a evoluce. 27 (12): 2669–73. doi:10.1093 / molbev / msq153. PMC 3108379. PMID 20573776.
externí odkazy
- Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba JP, Zucker CS, Patton A (2000). „Nová rodina receptorů na chuť savců - vyšetřovací přezkum“. Oddělení biologie na Davidson College. Citováno 2008-08-11.
- chuť + receptory, + typ + 1 v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- chuť + receptory, + typ + 2 v americké národní lékařské knihovně Lékařské předměty (Pletivo)