Octopamin - Octopamine
 | |
 | |
| Klinické údaje | |
|---|---|
| Ostatní jména | OCT, norsympathol, norsynefrin, odst-Oktamin, beta-hydroxytyramin, para-hydroxyfenyl-ethanolamin, a- (aminomethyl) -4-hydroxybenzenmethanol, l- (p-hydroxyfenyl) -2-aminoethanol |
| Trasy z správa | Ústní |
| ATC kód | |
| Fyziologický data | |
| Zdroj papírové kapesníky | nervový systém bezobratlých; stopový amin u obratlovců |
| Cílové tkáně | v celém systému u bezobratlých |
| Receptory | TAAR1 (savci) OctαR, OctβR, TyrR (bezobratlí), Oct-TyrR |
| Agonisté | Formamidiny (amitraz (AMZ) a chlordimeform (CDM)) |
| Antagonisté | epinastin (3-amino-9, 13b-dihydro-lH-dibenz (c, f) imidazo (1,5a) azepin hydrochlorid) |
| Předchůdce | tyramin |
| Biosyntéza | tyramin-p-hydroxyláza; dopamin-p-hydroxyláza |
| Metabolismus | str-hydroxymandlová kyselina;[1][2] N-acetyltransferázy; fenylethanolamin N-methyltransferáza |
| Právní status | |
| Právní status |
|
| Farmakokinetické data | |
| Biologická dostupnost | 99.42 % |
| Metabolismus | str-hydroxymandlová kyselina;[1][2] N-acetyltransferázy; fenylethanolamin N-methyltransferáza |
| Odstranění poločas rozpadu | 15 minut v hmyzu. U lidí mezi 76 a 175 minutami |
| Vylučování | Až 93% požitého oktopaminu je vyloučeno močí do 24 hodin[1] |
| Identifikátory | |
| |
| Číslo CAS | |
| PubChem CID | |
| IUPHAR / BPS | |
| ChemSpider | |
| UNII | |
| ChEBI | |
| ChEMBL | |
| Řídicí panel CompTox (EPA) | |
| Informační karta ECHA | 100.002.890 |
| Chemické a fyzikální údaje | |
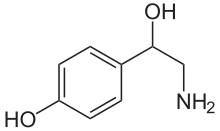
| Vzorec | C8H11NÓ2 |
| Molární hmotnost | 153.181 g · mol−1 |
| 3D model (JSmol ) | |
| |
| |
| | |
Octopamin (molekulární vzorec C8H11NE2; také známý jako odst-oktopaminy a další) je organická chemická látka úzce souvisí s norepinefrin a syntetizovány biologicky homologní cestou. Jeho název pochází ze skutečnosti, že byl poprvé identifikován ve slinných žlázách chobotnice.
U mnoha druhů bezobratlých je oktopamin důležitý neurotransmiter a hormon. v protostomy - členovci, měkkýši a několik druhů červů - nahrazuje norepinefrin a vykonává funkce zjevně podobné funkcím norepinefrinu u savců, funkce, které byly popsány jako mobilizace těla a nervového systému k činnosti. U savců se octopamin nachází pouze ve stopových množstvích a nebyla pro něj pevně stanovena žádná biologická funkce. Vyskytuje se také přirozeně v mnoha rostlinách, včetně hořký pomeranč.[3][4] Octopamin byl prodáván pod obchodními názvy, jako je Epirenor, Norden, a Norfen pro použití jako sympatomimetický lék, k dispozici na lékařský předpis.
Funkce
Buněčné efekty
Octopamin uplatňuje své účinky vazbou na a aktivuje receptory umístěné na povrchu buněk. Tyto receptory byly studovány hlavně u hmyzu, kde je lze rozdělit do tří typů: alfa-adrenergní (OctαR), které jsou strukturně a funkčně podobné noradrenergním alfa-1 receptorům u savců; beta-adrenergní látky (OctβR), které jsou strukturálně a funkčně podobné noradrenergním beta receptorům u savců; a smíšené receptory octopaminu / tyraminu (TyrR), které jsou strukturálně a funkčně podobné noradrenergním alfa-2 receptorům u savců.[5] Receptory ve třídě TyrR jsou však obecně silněji aktivovány tyramin než octopaminem.[5]
U obratlovců nebyly identifikovány žádné receptory specifické pro oktopamin. Octopamin se slabě váže na receptory pro norepinefrin a epinefrin, ale není jasné, zda to má nějaký funkční význam. Silněji se váže na receptory spojené se stopovými aminy (TAARs), zejména TAAR1.[5]
Bezobratlí
Octopamin byl poprvé objeven italským vědcem Vittorio Erspamer v roce 1948[6] ve slinných žlázách chobotnice a od té doby bylo zjištěno, že působí jako neurotransmiter, neurohormon a neuromodulátor v bezobratlých. Ačkoli Erspamer objevil jeho přirozený výskyt a pojmenoval jej, octopamin ve skutečnosti existoval po mnoho let jako farmaceutický produkt.[7] Je široce používán v energeticky náročném chování všem hmyzem, korýši (kraby, humři, raky) a pavouky. Mezi takové chování patří létání,[8] kladení vajec,[9] a skákání.[10][11]
Octopamin působí jako hmyz ekvivalent norepinefrin a podílí se na regulaci agrese u bezobratlých s různými účinky na různé druhy. Studie prokázaly, že snížení neurotransmiteru oktopaminu a prevence kódování tyramin beta hydroxyláza (enzym, který se převádí tyramin na oktopamin) snižuje agresi v Drosophila aniž by to ovlivnilo jiné chování.[12]
U hmyzu je octopamin uvolňován vybraným počtem neuronů, ale působí široce v celém centrálním mozku, na všechny smyslové orgány a na několik jiných než neuronálních tkání.[13][14] V hrudních gangliích je octopamin primárně uvolňován neurony DUM (dorální nepárový medián) a VUM (ventrální nepárový medián), které uvolňují oktopamin na nervové, svalové a periferní cíle.[15][16] Tyto neurony jsou důležité pro zprostředkování energeticky náročného motorického chování, jako je skákání vyvolané únikem a let. Například neuron kobylky DUMeti uvolňuje oktopamin do svalu tibie extensor, aby zvýšil svalové napětí a rychlost relaxace. Tyto akce podporují účinnou kontrakci svalů nohou při skákání.[13] Během letu jsou neurony DUM také aktivní a uvolňují oktopamin v celém těle, aby synchronizovaly energetický metabolismus, dýchání, svalovou aktivitu a letovou interneuronovou aktivitu.[8] Octopamin v kobylky je čtyřikrát koncentrovanější v axonu než v soma a snižuje kobylku myogenní rytmus.[17]
V včelí med a ovocný let „Octopamin hraje hlavní roli v učení a paměti. V světluška Uvolňování oktopaminu vede k produkci světla v lucerně.[18][19]
U humrů se zdá, že oktopamin řídí a koordinuje neurohormony do určité míry v centrálním nervovém systému a bylo pozorováno, že injekce octopaminu do humra a raků vedla k prodloužení končetiny a břicha.[20]
Heberlein a kol.[21] provedli studie tolerance alkoholu u ovocných mušek; zjistili, že mutace, která způsobila nedostatek oktopaminu, také způsobila nižší toleranci alkoholu.[22][23][24][25]
The smaragdový šváb vosa bodne hostitele za jeho larvy (šváb) v ganglionu hlavy (mozku). Jed blokuje receptory oktopaminu[26] a šváb nevykazuje normální únikové reakce a nadměrně se upravuje. Stává se poslušným a vosa jej táhne za anténu jako vodítko a vede ho do doupěte vosy.[27]
V hlístice „Octopamin se nachází ve vysokých koncentracích u dospělých, což snižuje chování při snášení vajec a čerpání hltanu s antagonistickým účinkem na serotonin.[28]
Octopaminergní nervy v měkkýš mohou být přítomny v srdci s vysokou koncentrací v nervovém systému.[29]
U larev v orientální armyworm „Octopamin je imunologicky prospěšný a zvyšuje míru přežití u populací s vysokou hustotou.[30]
Obratlovci
v obratlovců, nahrazuje octopamin norepinefrin v soucitný neurony s chronickým užíváním inhibitory monoaminooxidázy. Může být odpovědný za společné vedlejší účinek z ortostatická hypotenze s těmito agenty, i když existují také důkazy, že je to ve skutečnosti zprostředkováno zvýšenými hladinami N-acetylserotonin.
Jedna studie poznamenala, že oktopamin může být důležitým aminem, který ovlivňuje terapeutické účinky inhibitorů, jako jsou inhibitory monoaminooxidázy, zejména proto, že při léčbě tímto inhibitorem bylo pozorováno velké zvýšení hladin oktopaminu. Octopamin byl pozitivně identifikován ve vzorcích moči savců, jako jsou lidé, krysy a králíci léčeni inhibitory monoaminooxidázy. Velmi malé množství oktopaminu bylo také nalezeno v určitých zvířecích tkáních. Bylo pozorováno, že v těle králíka srdce a ledviny drží nejvyšší koncentrace oktopaminu. Bylo zjištěno, že oktopamin byl 93% eluován močí do 24 hodin poté, co byl v těle produkován jako vedlejší produkt iproniazidu u králíků.[7]
Farmakologie
Octopamin byl prodáván pod obchodními názvy, jako je Epirenor, Norden, a Norfen pro použití v medicíně jako a sympatomimetický lék, k dispozici na lékařský předpis. O jeho klinické užitečnosti nebo bezpečnosti však existuje jen velmi málo informací.[31]
v savci, Octopamin může mobilizovat uvolňování Tlustý z adipocyty (tukové buňky), což vedlo k jeho propagaci na internetu Internet jako pomoc při hubnutí. Uvolněný tuk je však pravděpodobně okamžitě absorbován do jiných buněk a neexistují žádné důkazy o tom, že oktopamin usnadňuje hubnutí. Octopamin se také může zvýšit krevní tlak v kombinaci s jinými stimulanty, jako v některých doplňky na hubnutí.[32]
The Světová antidopingová agentura uvádí oktopamin jako zakázanou látku pro soutěžní použití jako „specifický stimulant“[33] na Seznamu zakázaných 2019.
Insekticidy
Octopaminový receptor je terčem insekticidů, protože jeho zablokování vede ke snížení hladin cAMP. Esenciální oleje mohou mít takový neuro-insekticidní účinek,[34] a tento mechanismus oktopamin-receptor je přirozeně využíván rostlinami s aktivními insekticidními fytochemikáliemi.[35]
Biochemické mechanismy
Savci
Octopamin je jedním ze čtyř primárních endogenní agonisté člověka receptor související se stopovými aminy 1.[36]
Bezobratlí
Octopamin se váže na příslušné receptory spojené s G-proteinem (GPCR) a iniciuje tak cestu přenosu signálu buňky. Byly definovány nejméně tři skupiny octopaminového GPCR. OctαR (receptory OCTOPAMINE1) jsou více příbuzné s α-adrenergními receptory, zatímco OctβR (receptory OCTOPAMINE2) jsou více příbuzné s β-adrenergními receptory. Octopaminové / tyraminové receptory (včetně Oct-TyrR) mohou vázat oba ligandy a vykazovat specifické spojení s agonisty. Oct-TyrR je uveden v obou genových skupinách OCTOPAMINE a TYRAMINE RECEPTORS.[37]
Biosyntéza
U lidí

Viz také
Reference
- ^ A b Hengstmann, J. H .; Konen, W; Konen, C; Eichelbaum, M; Dengler, H. J. (1974). "Fyziologická dispozice p-oktopaminu u člověka". Archiv farmakologie Naunyn-Schmiedeberg. 283 (1): 93–106. doi:10.1007 / bf00500148. PMID 4277715. S2CID 35523412.
- ^ d’Andrea, Giovanni; Nordera, Gianpietro; Pizzolato, Gilberto; Bolner, Andrea; Colavito, Davide; Flaibani, Raffaella; Leon, Alberta (2010). „Stopový metabolismus aminů u Parkinsonovy choroby: Nízké cirkulující hladiny oktopaminu v raných stadiích onemocnění“. Neurovědy Dopisy. 469 (3): 348–51. doi:10.1016 / j.neulet.2009.12.025. PMID 20026245. S2CID 12797090.
- ^ Tang F, Tao L, Luo X, Ding L, Guo M, Nie L, Yao S (září 2006). „Stanovení oktopaminu, synefrinu a tyraminu v bylinách Citrus pomocí iontové kapaliny vylepšené„ zelené “chromatografie“. Journal of Chromatography A. 1125 (2): 182–8. doi:10.1016 / j.chroma.2006.05.049. PMID 16781718.
- ^ Jagiełło-Wójtowicz E (1979). "Mechanismus centrálního působení oktopaminu". Polský žurnál farmakologie a farmacie. 31 (5): 509–16. PMID 121158.
- ^ A b C Pflüger HJ, Stevensonb PA (2005). „Evoluční aspekty octopaminergních systémů s důrazem na členovce“. Struktura a vývoj členovců. 34 (3): 379–396. doi:10.1016 / j.asd.2005.04.004.CS1 maint: používá parametr autoři (odkaz)
- ^ Erspamer, V. (2009). „Účinné látky v zadních slinných žlázách chobotnice. II. Tyramin a oktopamin (oxyoktopamin)“. Acta Pharmacologica et Toxicologica. 4 (3–4): 224–47. doi:10.1111 / j.1600-0773.1948.tb03345.x.
- ^ A b Kakimoto Y, Armstrong MD (únor 1962). „O identifikaci oktopaminu u savců“. The Journal of Biological Chemistry. 237: 422–7. PMID 14453200.
- ^ A b Orchard, I; Ramirez, J. M.; Lange, AB (leden 1993). "Multifunkční role octopaminu v letu Locust". Každoroční přezkum entomologie. 38 (1): 227–249. doi:10.1146 / annurev.en.38.010193.001303. ISSN 0066-4170.
- ^ Lee, Hyun-Gwan; Seong, Chang-Soo; Kim, Young-Cho; Davis, Ronald L; Han, Kyung-An (1. prosince 2003). „Octopaminový receptor OAMB je nutný pro ovulaci u Drosophila melanogaster“. Vývojová biologie. 264 (1): 179–190. doi:10.1016 / j.ydbio.2003.07.018. ISSN 0012-1606. PMID 14623240.
- ^ Pollack, Alan J .; Ritzmann, Roy E .; Westin, Joanne (1988). "Aktivace DUM buněčných interneuronů ventrálními obřími interneurony u švába, periplaneta americana". Journal of Neurobiology. 19 (6): 489–497. doi:10,1002 / neu.480190602. ISSN 1097-4695. PMID 3171574.
- ^ Orchard, Ian (1. dubna 1982). „Octopamin v hmyzu: neurotransmiter, neurohormon a neuromodulátor“. Kanadský žurnál zoologie. 60 (4): 659–669. doi:10.1139 / z82-095. ISSN 0008-4301.
- ^ Zhou C, Rao Y, Rao Y (září 2008). „Podskupina oktopaminergních neuronů je důležitá pro agresi Drosophila“. Přírodní neurovědy. 11 (9): 1059–67. doi:10.1038 / nn.2164. PMID 19160504. S2CID 1134848.
- ^ A b Atwood, H.L .; Klose, M. K. (1. ledna 2009), „Modulace neuromuskulárního přenosu na neuromuskulárních spojích bezobratlých“, v Squire, Larry R. (ed.), Encyclopedia of Neuroscience, Oxford: Academic Press, s. 671–690, ISBN 978-0-08-045046-9, vyvoláno 10. července 2020
- ^ Roeder, T. (prosinec 1999). "Octopamin u bezobratlých". Pokrok v neurobiologii. 59 (5): 533–561. doi:10.1016 / s0301-0082 (99) 00016-7. ISSN 0301-0082. PMID 10515667. S2CID 25654298.
- ^ Eckert, Manfred; Rapus, Jürgen; Nürnberger, Asja; Penzlin, Heinz (1992). „Nová specifická protilátka odhaluje imunoreaktivitu podobnou octopaminu ve ventrálním nervovém kabelu švába“. Journal of Comparative Neurology (francouzsky). 322 (1): 1–15. doi:10,1002 / k.903220102. ISSN 1096-9861. PMID 1430305. S2CID 41099770.
- ^ Sinakevitch, Irina G .; Geffard, Michel; Pelhate, Marcel; Lapied, Bruno (duben 1994). „Octopaminová imunoreaktivita v dorzálních nepárových středních neuronech (DUM) inervujících pomocnou žlázu mužského švába Periplaneta americana“. Výzkum buněk a tkání. 276 (1): 15–21. doi:10.1007 / bf00354779. ISSN 0302-766X. S2CID 23485136.
- ^ Evans, P. D .; O'Shea, M. (duben 1978). „Identifikace oktopaminergního neuronu a modulace myogenního rytmu v kobylce“. The Journal of Experimental Biology. 73: 235–260. ISSN 0022-0949. PMID 25941.
- ^ Greenfield MD (listopad 2001). „Chybějící odkaz v bioluminiscenci světlušek odhalen: ŽÁDNÁ regulace dýchání fotocitů“. BioEssays. 23 (11): 992–5. doi:10,1002 / bies.1144. PMID 11746215.
- ^ Trimmer BA, Aprille JR, Dudzinski DM, Lagace CJ, Lewis SM, Michel T a kol. (Červen 2001). "Oxid dusnatý a ovládání blikání světlušek". Věda. 292 (5526): 2486–8. doi:10.1126 / science.1059833. PMID 11431567. S2CID 1095642.
- ^ Livingstone MS, Harris-Warrick RM, Kravitz EA (duben 1980). "Serotonin a octopamin produkují opačné polohy u humrů". Věda. 208 (4439): 76–9. Bibcode:1980Sci ... 208 ... 76L. doi:10.1126 / science.208.4439.76. PMID 17731572. S2CID 32141532.
- ^ Heberlein U, Wolf FW, Rothenfluh A, Guarnieri DJ (srpen 2004). "Molekulární genetická analýza intoxikace ethanolem v Drosophila melanogaster". Integrativní a srovnávací biologie. 44 (4): 269–74. CiteSeerX 10.1.1.536.262. doi:10.1093 / icb / 44.4.269. PMID 21676709. S2CID 14762870.
- ^ Moore MS, DeZazzo J, Luk AY, Tully T, Singh CM, Heberlein U (červen 1998). „Intoxikace ethanolem v Drosophile: Genetické a farmakologické důkazy pro regulaci signální cestou cAMP“. Buňka. 93 (6): 997–1007. doi:10.1016 / S0092-8674 (00) 81205-2. PMID 9635429. S2CID 15312752.
- ^ Tecott LH, Heberlein U (prosinec 1998). „Pijeme?“. Buňka. 95 (6): 733–5. doi:10.1016 / S0092-8674 (00) 81695-5. PMID 9865690.
- ^ Williams, Ruth (22. června 2005). „Bar Flies: Co nás naši příbuzní hmyzu mohou naučit o toleranci k alkoholu“. Nahý vědec.
- ^ Vince, Gaia (22. srpna 2005). "'Gen kocoviny je klíčem k toleranci vůči alkoholu “. Nový vědec.
- ^ Hopkin, Michael (2007). "Jak vyrobit zombie šváb". Příroda. doi:10.1038 / novinky.2007.312.
- ^ Gal R, Rosenberg LA, Libersat F (prosinec 2005). „Vosa parazitoidní používá koktejl jedu vstřikovaný do mozku k manipulaci s chováním a metabolizmem své kořisti švábů“. Archiv biochemie a fyziologie hmyzu. 60 (4): 198–208. doi:10.1002 / arch.20092. PMID 16304619.
- ^ Horvitz, H. R .; Chalfie, M .; Trent, C .; Sulston, J. E .; Evans, P. D. (28. května 1982). "Serotonin a oktopamin u hlístice Caenorhabditis elegans". Věda. 216 (4549): 1012–1014. doi:10.1126 / science.6805073. ISSN 0036-8075. PMID 6805073.
- ^ Dougan, D. F. H .; Duffield, P. H .; Wade, D. N .; Duffield, A. M. (1. ledna 1981). „Výskyt a syntéza oktopaminu v srdci a gangliích měkkýšů Tapes watlingi“. Srovnávací biochemie a fyziologie Část C: Srovnávací farmakologie. 70 (2): 277–280. doi:10.1016/0306-4492(81)90064-2. ISSN 0306-4492.
- ^ Kong, Hailong; Yuan, Lin; Dong, Chuanlei; Zheng, Minyuan; Jing, Wanghui; Tian, Zhen; Hou, Qiuli; Cheng, Yunxia; Zhang, Lei; Jiang, Xingfu; Luo, Lizhi (prosinec 2020). „Imunologická regulace genem receptoru octopaminu podobného β-adrenergním látkám v přeplněných larvách orientálního Armyworm, Mythmina separata“. Vývojová a srovnávací imunologie. 113: 103802. doi:10.1016 / j.dci.2020.103802. ISSN 1879-0089. PMID 32712170.
- ^ Stohs SJ (leden 2015). "Fyziologické funkce a farmakologické a toxikologické účinky p-oktopaminu". Drogová a chemická toxikologie. 38 (1): 106–12. doi:10.3109/01480545.2014.900069. PMID 24654910. S2CID 21901553.
- ^ Haller CA, Benowitz NL, Jacob P (září 2005). "Hemodynamické účinky doplňků na hubnutí bez ephedry u lidí". American Journal of Medicine. 118 (9): 998–1003. doi:10.1016 / j.amjmed.2005.02.034. PMID 16164886.
- ^ „Zakázáno v soutěži - stimulanty“. WADA. Citováno 6. května 2019.
- ^ Enan, Essam (1. listopadu 2001). „Insekticidní aktivita éterických olejů: octopaminergní místa působení“. Srovnávací biochemie a fyziologie, část C: Toxikologie a farmakologie. 130 (3): 325–337. doi:10.1016 / S1532-0456 (01) 00255-1. ISSN 1532-0456.
- ^ Ratan, Rameshwar Singh (1. září 2010). „Mechanismus účinku sekundárních insekticidních metabolitů rostlinného původu“. Ochrana plodin. 29 (9): 913–920. doi:10.1016 / j.cropro.2010.05.008. ISSN 0261-2194.
- ^ Maguire JJ, Davenport AP (20. února 2018). "Stopový aminový receptor: TA1 receptor ". Průvodce IUPHAR / BPS FARMAKOLOGIE. Mezinárodní unie základní a klinické farmakologie. Citováno 16. července 2018.
- ^ „Gene Group: OCTOPAMINE RECEPTORS“, FlyBase, 16. října 2018.
- ^ Broadley KJ (březen 2010). "Cévní účinky stopových aminů a amfetaminů". Farmakologie a terapeutika. 125 (3): 363–375. doi:10.1016 / j.pharmthera.2009.11.005. PMID 19948186.
- ^ Lindemann L, Hoener MC (květen 2005). „Renesance stopových aminů inspirovaná novou rodinou GPCR“. Trendy ve farmakologických vědách. 26 (5): 274–281. doi:10.1016 / j.tips.2005.03.007. PMID 15860375.
- ^ Wang X, Li J, Dong G, Yue J (únor 2014). "Endogenní substráty mozkového CYP2D". European Journal of Pharmacology. 724: 211–218. doi:10.1016 / j.ejphar.2013.12.025. PMID 24374199.