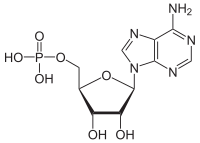
Adenosinmonofosfát - Adenosine monophosphate
Tento článek obsahuje seznam obecných Reference, ale zůstává z velké části neověřený, protože postrádá dostatečné odpovídající vložené citace. (únor 2013) (Zjistěte, jak a kdy odstranit tuto zprávu šablony) |
 | |
 | |
| Jména | |
|---|---|
| Název IUPAC [(2R, 3S, 4R, 5R) -5- (6-aminopurin-9-yl) -3,4-dihydroxyoxolan-2-yl] methyl-dihydrogenfosforečnan | |
| Ostatní jména Adenosin 5'-monofosfát, kyselina 5'-adenylová | |
| Identifikátory | |
3D model (JSmol ) | |
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| DrugBank | |
| Informační karta ECHA | 100.000.455 |
| KEGG | |
| Pletivo | Adenosin + monofosfát |
PubChem CID | |
| UNII | |
Řídicí panel CompTox (EPA) | |
| |
| |
| Vlastnosti | |
| C10H14N5Ó7P | |
| Molární hmotnost | 347,22 g / mol |
| Vzhled | bílý krystalický prášek |
| Hustota | 2,32 g / ml |
| Bod tání | 178 až 185 ° C (352 až 365 ° F; 451 až 458 K) |
| Bod varu | 798,5 ° C (1469,3 ° F; 1071,7 K) |
| Kyselost (strK.A) | 0.9[Citace je zapotřebí ], 3.8, 6.1 |
Pokud není uvedeno jinak, jsou uvedeny údaje o materiálech v nich standardní stav (při 25 ° C [77 ° F], 100 kPa). | |
| Reference Infoboxu | |
Adenosinmonofosfát (AMP), také známý jako 5'-adenylová kyselina, je nukleotid. AMP se skládá z a fosfát skupina, cukr ribóza a nukleobáze adenin; to je ester z kyselina fosforečná a nukleosid adenosin.[1] Jako substituent má formu předpony adenylyl-.[2]
AMP hraje důležitou roli v mnoha buněčných metabolických procesech, na které se přeměňuje ADP a / nebo ATP. AMP je také součástí syntézy RNA.[3] AMP je přítomen ve všech známých formách života.[4]
Výroba a degradace
AMP nemá vysokou energii fosfoanhydrid vazba spojená s ADP a ATP. AMP lze vyrobit z ADP:
- 2 ADP → ATP + AMP
Nebo AMP může být produkován hydrolýza jednoho fosfát s vysokou energií svazek ADP:
- ADP + H2O → AMP + Pi
AMP lze také vytvořit hydrolýzou ATP do AMP a pyrofosfát:
- ATP + H2O → AMP + PPi
Když se RNA rozloží živými systémy, vzniknou nukleosidmonofosfáty, včetně adenosinmonofosfátu.
AMP lze regenerovat na ATP následujícím způsobem:
- AMP + ATP → 2 ADP (adenylát kináza v opačném směru)
- ADP + Pi → ATP (tento krok se nejčastěji provádí v aerobech pomocí ATP syntáza v době oxidační fosforylace )
AMP lze převést na IMP podle enzym myoadenylát deamináza, uvolnění amoniak skupina.
V katabolický cestou lze adenosinmonofosfát převést na kyselina močová, který se vylučuje z těla u savců.[5]
Fyziologická role v regulaci
Regulace kinázy aktivované AMP
Enzym eukaryotických buněk 5 'adenosinmonofosfátem aktivovaná protein kináza nebo AMPK využívá AMP pro homeostatický energetické procesy v době vysokého výdeje buněčné energie, jako je cvičení.[6] Od štěpení ATP a odpovídající fosforylace reakce, jsou využívány v různých procesech v celém těle jako zdroj energie, produkce ATP je nezbytná k dalšímu vytváření energie pro tyto savčí buňky. AMPK jako snímač buněčné energie je aktivován snižováním hladin ATP, což je přirozeně doprovázeno zvyšováním hladin ADP a AMP.[7]
Ačkoli se zdá, že fosforylace je hlavní aktivátor pro AMPK některé studie naznačují, že AMP je alosterický regulátor stejně jako a přímý agonista pro AMPK.[8] Další studie dále naznačují, že vysoký poměr hladin AMP: ATP v buňkách aktivuje AMPK spíše než jen AMP.[9] Například druh Caenorhabditis elegans a Drosophila melanogaster a bylo zjištěno, že jejich AMP-aktivované kinázy byly aktivovány AMP, zatímco druhy droždí a rostlinné kinázy nebyly alostericky aktivovány AMP.[9]
AMP se váže na y-podjednotka AMPK, vedoucí k aktivaci kinázy, a nakonec a kaskáda dalších procesů, jako je aktivace katabolický cesty a inhibice z anabolický cesty k regeneraci ATP. Katabolické mechanismy, které generují ATP uvolňováním energie z rozpadu molekul, jsou aktivovány enzymem AMPK, zatímco anabolické mechanismy, které využívají energii z ATP k tvorbě produktů, jsou inhibovány.[10] Ačkoli γ-podjednotka může vázat AMP / ADP / ATP, pouze vazba AMP / ADP vede ke konformačnímu posunu enzymového proteinu. Tato odchylka ve vazbě AMP / ADP versus ATP vede k posunu v defosforylace stav pro enzym.[11] Defosforylace AMPK prostřednictvím různých proteinů fosfatázy zcela deaktivuje katalytickou funkci. AMP / ADP chrání AMPK před inaktivací vazbou na y-podjednotka a udržování stavu defosforylace.[12]
tábor
AMP může také existovat jako cyklická struktura známá jako cyklický AMP (nebo cAMP). V určitých buňkách enzym adenylátcykláza vyrábí cAMP z ATP a tato reakce je obvykle regulována hormony, jako je adrenalin nebo glukagon. cAMP hraje důležitou roli v intracelulární signalizaci.[13]
Viz také
Reference
- ^ "Adenosinmonofosfát (sloučenina)". PubChem. NCBI. Citováno 30. dubna 2020.
- ^ „Nomenklatura sacharidů: (Doporučení 1996)“. Journal of Carbohydrate Chemistry. 16 (8): 1191–1280. 1997. doi:10.1080/07328309708005748.
- ^ Jauker M, Griesser H, Richert C (listopad 2015). „Spontánní tvorba pramenů RNA, peptidylové RNA a kofaktorů“. Angewandte Chemie. 54 (48): 14564–9. doi:10,1002 / anie.201506593. PMC 4678511. PMID 26435376.
- ^ "Adenosinmonofosfát". Databáze lidských metabolomů. Citováno 3. července 2020.
- ^ Maiuolo J, Oppedisano F, Gratteri S, Muscoli C, Mollace V (červen 2016). „Regulace metabolismu a vylučování kyseliny močové“. International Journal of Cardiology. 213: 8–14. doi:10.1016 / j.ijcard.2015.08.109. PMID 26316329.
- ^ Richter EA, Ruderman NB (březen 2009). „AMPK a biochemie cvičení: důsledky pro lidské zdraví a nemoci“. The Biochemical Journal. 418 (2): 261–75. doi:10.1042 / BJ20082055. PMC 2779044. PMID 19196246.
- ^ Carling D, Mayer FV, Sanders MJ, Gamblin SJ (červenec 2011). „Protein kináza aktivovaná AMP: energetický senzor přírody“. Přírodní chemická biologie. 7 (8): 512–8. doi:10.1038 / nchembio.610. PMID 21769098.
- ^ Faubert B, Vincent EE, Poffenberger MC, Jones RG (leden 2015). „AMP-aktivovaná protein kináza (AMPK) a rakovina: mnoho tváří metabolického regulátoru“. Dopisy o rakovině. 356 (2 Pt A): 165–70. doi:10.1016 / j.canlet.2014.01.018. PMID 24486219.
- ^ A b Hardie DG (15. září 2011). „Proteinkináza aktivovaná AMP - energetický senzor, který reguluje všechny aspekty funkce buněk“. Geny a vývoj. 25 (18): 1895–1908. doi:10.1101 / gad.17420111. ISSN 0890-9369. PMC 3185962. PMID 21937710.
- ^ Hardie DG (únor 2011). „Snímání energie pomocí AMP-aktivované proteinkinázy a její účinky na metabolismus svalů“. Sborník společnosti pro výživu. 70 (1): 92–9. doi:10.1017 / S0029665110003915. PMID 21067629.
- ^ Krishan S, Richardson DR, Sahni S (březen 2015). „Adenozinmonofosfátem aktivovaná kináza a její klíčová role v katabolismu: struktura, regulace, biologická aktivita a farmakologická aktivace“. Molekulární farmakologie. 87 (3): 363–77. doi:10,1124 / mol. 114,095810. PMID 25422142.
- ^ Xiao B, Sanders MJ, Underwood E, Heath R, Mayer FV, Carmena D, Jing C, Walker PA, Eccleston JF, Haire LF, Saiu P, Howell SA, Aasland R, Martin SR, Carling D, Gamblin SJ (duben 2011 ). „Struktura AMPK savců a její regulace pomocí ADP“. Příroda. 472 (7342): 230–3. doi:10.1038 / nature09932. PMC 3078618. PMID 21399626.
- ^ Ravnskjaer K, Madiraju A, Montminy M (2015). Metabolická kontrola. Příručka experimentální farmakologie. 233. Springer, Cham. str. 29–49. doi:10.1007/164_2015_32. ISBN 9783319298047. PMID 26721678.
Další čtení
- Ming D, Ninomiya Y, Margolskee RF (srpen 1999). „Blokování aktivace chuťových receptorů gustducinu inhibuje chuťové reakce na hořké sloučeniny“. Sborník Národní akademie věd Spojených států amerických. 96 (17): 9903–8. doi:10.1073 / pnas.96.17.9903. PMC 22308. PMID 10449792.