Biosyntéza - Biosynthesis
Biosyntéza je vícestupňový, enzym -katalyzovaný proces kde substráty jsou převedeny na složitější produkty v živých organismech. V biosyntéze jednoduché sloučeniny jsou modifikovány, převedeny na jiné sloučeniny nebo spojeny dohromady za vzniku makromolekuly. Tento proces často sestává z metabolické cesty. Některé z těchto biosyntetických drah jsou umístěny v jedné buňce organela, zatímco jiné zahrnují enzymy, které se nacházejí v několika buněčných organelách. Příklady těchto biosyntetických drah zahrnují produkci lipidová membrána komponenty a nukleotidy. Biosyntéza je obvykle synonymní s anabolismus.
Nezbytné prvky pro biosyntézu zahrnují: předchůdce sloučeniny, chemická energie (např. ATP ) a katalytické enzymy, které mohou vyžadovat koenzymy (např.NADH, NADPH ). Tyto prvky vytvářejí monomery, stavební kameny pro makromolekuly. Některé důležité biologické makromolekuly zahrnují: bílkoviny, které se skládají z aminokyselina monomery připojené prostřednictvím peptidové vazby, a DNA molekuly, které jsou složeny z nukleotidů spojených prostřednictvím fosfodiesterové vazby.
Vlastnosti chemických reakcí
K biosyntéze dochází v důsledku řady chemických reakcí. K uskutečnění těchto reakcí jsou nutné následující prvky:[1]
- Prekurzorové sloučeniny: tyto sloučeniny jsou výchozí molekuly nebo substráty v reakci. Ty lze také považovat za reaktanty v daném chemickém procesu.
- Chemická energie: chemickou energii lze nalézt ve formě vysokoenergetických molekul. Tyto molekuly jsou potřebné pro energeticky nepříznivé reakce. Kromě toho hydrolýza těchto sloučenin pohání reakci vpřed. Molekuly s vysokou energií, jako např ATP, mít tři fosfáty. Koncový fosfát se často během hydrolýzy odštěpí a přenese do jiné molekuly.
- Katalytické enzymy: tyto molekuly jsou zvláštní bílkoviny které katalyzují reakci zvýšením rychlost reakce a spouštění aktivační energie.
- Koenzymy nebo kofaktory: kofaktory jsou molekuly, které pomáhají při chemických reakcích. Mohou být ionty kovů, vitaminové deriváty jako např NADH a acetyl CoA nebo nevitaminové deriváty, jako je ATP. V případě NADH molekula přenáší vodík, zatímco acetyl CoA přenáší vodík acetylová skupina a ATP přenáší fosfát.
V nejjednodušším smyslu mají reakce, které se vyskytují v biosyntéze, následující formát:[2]
![{ displaystyle { ce {Reaktant -> [] [enzym] Produkt}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/d97fbef58f64fb0b7d05d90fa0a73b05f5919fc8)
Některé varianty této základní rovnice, které budou podrobněji popsány dále, jsou:[3]
- Jednoduché sloučeniny, které se převádějí na jiné sloučeniny, obvykle jako součást vícestupňové reakční cesty. Dva příklady tohoto typu reakce se vyskytují během tvorby nukleové kyseliny a nabíjení z tRNA před překlad. U některých z těchto kroků je vyžadována chemická energie:
- Jednoduché sloučeniny, které se pomocí kofaktorů převádějí na jiné sloučeniny. Například syntéza fosfolipidy vyžaduje acetyl CoA, zatímco syntéza jiné membránové složky, sfingolipidy, vyžaduje NADH a FADH pro formování sfingosin páteř. Obecná rovnice pro tyto příklady je:
- Jednoduché sloučeniny, které spojují dohromady a vytvářejí makromolekulu. Například, mastné kyseliny spojte se a vytvořte fosfolipidy. Fosfolipidy a cholesterol komunikovat nekovalentně za účelem vytvoření lipidová dvojvrstva. Tuto reakci lze popsat následovně:

![{ displaystyle { ce {{Prekurzor ~ molekula} + kofaktor -> [] [enzym] makromolekula}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b192f25eec9ceebed179bfeb43ae27795f3a91be)

Lipid
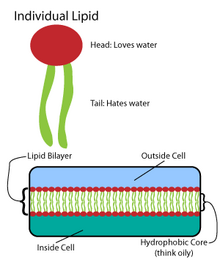
Mnoho složitých makromolekul je syntetizováno ve vzoru jednoduchých opakovaných struktur.[4] Například nejjednodušší struktury lipidů jsou mastné kyseliny. Mastné kyseliny jsou uhlovodík deriváty; obsahují a karboxylová skupina „hlava“ a „ocas“ uhlovodíkového řetězce.[4] Tyto mastné kyseliny vytvářejí větší složky, které zase zahrnují nekovalentní interakce za vzniku lipidové dvojvrstvy.[4]Řetězce mastných kyselin se nacházejí ve dvou hlavních složkách membránových lipidů: fosfolipidy a sfingolipidy. Třetí hlavní membránová složka, cholesterol, neobsahuje tyto jednotky mastných kyselin.[5]
Fosfolipidy
Základ všech biomembrán tvoří a dvouvrstvá struktura fosfolipidů.[6] Fosfolipidová molekula je amfipatický; obsahuje a hydrofilní polární hlava a hydrofobní nepolární ocas.[4] Fosfolipidové hlavy interagují navzájem a s vodným médiem, zatímco uhlovodíkové zbytky se orientují ve středu, daleko od vody.[7] Tyto poslední interakce řídí dvojvrstvou strukturu, která působí jako bariéra pro ionty a molekuly.[8]
Existují různé typy fosfolipidů; v důsledku toho se jejich cesty syntézy liší. První krok syntézy fosfolipidů však zahrnuje tvorbu fosfatidát nebo diacylglycerol 3-fosfát na endoplazmatické retikulum a vnější mitochondriální membrána.[7] Cesta syntézy je uvedena níže:

Cesta začíná glycerol 3-fosfátem, který se převádí na lysofosfatidát přidáním řetězce mastných kyselin poskytovaného acyl koenzym A.[9] Poté se lysofosfatidát převede na fosfatidát přidáním dalšího řetězce mastných kyselin, k němuž přispívá druhý acyl CoA; všechny tyto kroky jsou katalyzovány glycerolfosfátem acyltransferáza enzym.[9] Syntéza fosfolipidů pokračuje v endoplazmatickém retikulu a cesta biosyntézy se liší v závislosti na složkách konkrétního fosfolipidu.[9]
Sfingolipidy
Stejně jako fosfolipidy mají tyto deriváty mastných kyselin polární hlavu a nepolární ocasy.[5] Na rozdíl od fosfolipidů mají sfingolipidy a sfingosin páteř.[10] Sfingolipidy existují v eukaryotický buněk a jsou obzvláště hojné v centrální nervový systém.[7] Například sfingomyelin je součástí myelinová vrstva nervových vláken.[11]
Sfingolipidy jsou tvořeny z ceramidy které sestávají z řetězce mastných kyselin připojeného k aminoskupině páteře sfingosinu. Tyto ceramidy jsou syntetizovány z acylace sfingosinu.[11] Biosyntetická cesta pro sfingosin je uvedena níže:

Jak ukazuje obrázek, během syntézy sfingosinu prochází palmitoyl CoA a serin a kondenzační reakce což vede k tvorbě dehydrosphingosinu.[7] Tento produkt se poté redukuje za vzniku dihydrospingosinu, který se prostřednictvím. Převádí na sfingosin oxidační reakce podle FAD.[7]
Cholesterol
Tento lipid patří do třídy molekul zvaných steroly.[5] Steroly mají čtyři kondenzované kruhy a hydroxylová skupina.[5] Cholesterol je obzvláště důležitá molekula. Nejen, že slouží jako součást lipidových membrán, je také předchůdcem několika steroid hormony, včetně kortizol, testosteron, a estrogen.[12]
Cholesterol je syntetizován z acetyl CoA.[12] Cesta je zobrazena níže:

Obecněji řečeno, tato syntéza probíhá ve třech stupních, přičemž první stupeň probíhá v cytoplazma a druhá a třetí fáze vyskytující se v endoplazmatickém retikulu.[9] Fáze jsou následující:[12]
- 1. Syntéza isopentenylpyrofosfát, „stavební kámen“ cholesterolu
- 2. Vznik skvalen kondenzací šesti molekul isopentenylfosfátu
- 3. Konverze skvalenu na cholesterol několika enzymatickými reakcemi
Nukleotidy
Biosyntéza nukleotidy zahrnuje enzymkatalyzovaný reakce, které přeměňují substráty na složitější produkty.[1] Nukleotidy jsou stavebními kameny DNA a RNA. Nukleotidy se skládají z pětičlenného kruhu vytvořeného z ribóza cukr v RNA a deoxyribóza cukr v DNA; tyto cukry jsou spojeny s a purin nebo pyrimidin základna s a glykosidová vazba a a fosfát skupina u 5 'umístění cukru.[13]
Purinové nukleotidy

DNA nukleotidy adenosin a guanosin sestávají z purinové báze připojené k ribózovému cukru s glykosidovou vazbou. V případě RNA nukleotidů deoxyadenosin a deoxyguanosin, purinové báze jsou připojeny k deoxyribózovému cukru s glykosidovou vazbou. Purinové báze na DNA a RNA nukleotidech jsou syntetizovány ve dvanáctikrokovém reakčním mechanismu přítomném ve většině jednobuněčných organismů. Vyšší eukaryoty zaměstnat podobné mechanismus reakce v deseti reakčních krocích. Purinové báze se syntetizují konvertováním fosforibosylpyrofosfát (PRPP) do inosin monofosfát (IMP), což je první klíčový meziprodukt v biosyntéze purinové báze.[14] Další enzymatická modifikace IMP produkuje adenosinové a guanosinové báze nukleotidů.
- Prvním krokem v biosyntéze purinů je a kondenzační reakce, provádí glutamin-PRPP amidotransferáza. Tento enzym přenáší aminoskupina z glutamin k PRPP, formování 5-fosforibosylamin. Následující krok vyžaduje aktivaci glycin přidáním a fosfát skupina z ATP.
- GAR syntetáza[15] provádí kondenzaci aktivovaného glycinu na PRPP a tvoří se glycineamid ribonukleotid (GAR).
- GAR transformyláza dodává formyl skupina na aminoskupinu GAR za vzniku formylglycinamid ribonukleotidu (FGAR).
- FGAR amidotransferáza[16] katalyzuje přidání dusíkové skupiny k FGAR za vzniku formylglycinamidin ribonukleotidu (FGAM).
- FGAM cykláza katalyzuje uzavření kruhu, které zahrnuje odstranění molekuly vody za vzniku 5členného imidazol prsten 5-aminoimidazol ribonukleotid (VZDUCH).
- N5-CAIR syntetáza přenáší a karboxyl skupina, tvořící meziprodukt N5-karboxyaminoimidazol ribonukleotid (N5-CAIR).[17]
- N5-CAIR mutáza přeskupí karboxylovou funkční skupinu a přenese ji na imidazolový kruh a vytvoří se karboxyaminoimidazol ribonukleotid (CAIR). Dvoustupňový mechanismus tvorby CAIR z prostředí AIR se většinou nachází v jednobuněčných organismech. Vyšší eukaryota obsahují enzym AIR karboxyláza,[18] který přenáší karboxylovou skupinu přímo na imidazolový kruh AIR a vytváří CAIR.
- SAICAR syntetáza tvoří a peptidová vazba mezi aspartát a přidaná karboxylová skupina imidazolového kruhu, která se tvoří N-sukcinyl-5-aminoimidazol-4-karboxamid ribonukleotid (SAICAR).
- SAICAR lyáza odstraní uhlíkovou kostru přidaného aspartátu, ponechá aminoskupinu a vytvoří se 5-aminoimidazol-4-karboxamid ribonukleotid (AICAR).
- AICAR transformyláza přenáší karbonylovou skupinu na AICAR a tvoří se N-formylaminoimidazol-4-karboxamid ribonukleotid (FAICAR).
- Poslední krok zahrnuje enzym IMP syntáza, který provádí uzavření purinového kruhu a tvoří meziprodukt inosinmonofosfátu.[5]
Pyrimidinové nukleotidy

Další DNA a RNA nukleotidové báze, které jsou spojeny s ribózovým cukrem přes glykosidovou vazbu, jsou tymin, cytosin a uracil (který se nachází pouze v RNA).Uridinmonofosfát biosyntéza zahrnuje enzym, který se nachází v mitochondriální vnitřní membrána a multifunkční enzymy, které se nacházejí v cytosol.[19]
- První krok zahrnuje enzym karbamoylfosfát syntáza kombinování glutamin s CO2 v reakci závislé na ATP karbamoyl fosfát.
- Aspartát karbamoyltransferáza kondenzuje karbamoylfosfát s aspartátem za vzniku uridosukcinátu.
- Dihydroorotáza provádí uzavření kroužku reakce, která ztrácí vodu dihydroorotát.
- Dihydroorotát dehydrogenáza, umístěný uvnitř mitochondriální vnitřní membrány,[19] oxiduje dihydroorotát na orotovat.
- Orotát fosforibosylhydroláza (OMP pyrofosforyláza) kondenzuje orotát s PRPP tvořit orotidin-5'-fosfát.
- OMP dekarboxyláza katalyzuje konverzi orotidin-5'-fosfátu na UMP.[20]
Po syntéze uridinové nukleotidové báze se syntetizují další báze, cytosin a thymin. Biosyntéza cytosinu je dvoustupňová reakce, která zahrnuje přeměnu UMP na UTP. Fosfát přídavek k UMP je katalyzován a kináza enzym. Enzym CTP syntáza katalyzuje další reakční krok: konverzi UTP na CTP převodem aminoskupina od glutaminu po uridin; toto tvoří cytosinovou bázi CTP.[21] Mechanismus, který zobrazuje reakci UTP + ATP + glutamin ⇔ CTP + ADP + glutamát, je níže:

Cytosin je nukleotid, který je přítomen v DNA i RNA. Uracil se však nachází pouze v RNA. Proto poté, co je syntetizován UTP, musí být převeden na deoxy forma, která má být začleněna do DNA. Tato přeměna zahrnuje enzym ribonukleosid trifosfát reduktáza. Tato reakce, která odstraní 2'-OH ribózového cukru za vzniku deoxyribózy, není ovlivněna bázemi připojenými k cukru. Tato nespecifičnost umožňuje ribonukleosid trifosfát reduktáze převést vše nukleotid trifosfáty na deoxyribonukleotid podobným mechanismem.[21]
Na rozdíl od uracilu se tyminové báze nacházejí většinou v DNA, nikoli v RNA. Buňky normálně neobsahují tyminové báze, které jsou spojeny s ribózovými cukry v RNA, což naznačuje, že buňky syntetizují pouze tymin vázaný na deoxyribózu. Enzym thymidylát syntetáza je zodpovědný za syntézu thyminových zbytků z skládka na dTMP. Tato reakce přenáší a methyl seskupte na uracilovou bázi dUMP a vytvořte dTMP.[21] Vpravo je uvedena reakce thymidylátsyntázy, dUMP + 5,10-methylenetetrahydrofolát ⇔ dTMP + dihydrofolát.
DNA
I když existují rozdíly mezi eukaryotický a prokaryotický Syntéza DNA, následující část označuje klíčové vlastnosti replikace DNA sdílené oběma organismy.
DNA se skládá z nukleotidy ke kterým se připojil fosfodiesterové vazby.[4] Syntéza DNA, který se odehrává v jádro, je polokonzervativní proces, což znamená, že výsledná molekula DNA obsahuje původní řetězec z rodičovské struktury a nový řetězec.[22] Syntéza DNA je katalyzována rodinou DNA polymerázy které vyžadují čtyři deoxynukleosid trifosfáty, a pramen šablony a primer s volným 3'OH, do kterého se začleňují nukleotidy.[23]
Aby došlo k replikaci DNA, a replikační vidlice je tvořen enzymy zvanými helikázy které odvíjejí šroubovice DNA.[23] Topoizomerázy na replikační vidlici odeberte supercoily způsobené odvíjením DNA a jednovláknové proteiny vázající DNA udržovat dva jednovláknové šablony DNA stabilizované před replikací.[13]
Syntéza DNA je iniciována RNA polymeráza primase, který vytváří RNA primer s volným 3'OH.[23] Tento primer je připojen k jednořetězcovému templátu DNA a DNA polymeráza prodlužuje řetězec začleněním nukleotidů; DNA polymeráza také koriguje nově syntetizovaný řetězec DNA.[23]
Během polymerační reakce katalyzované DNA polymerázou, a nukleofilní útok nastává 3'OH rostoucího řetězce na nejvnitřnějším atomu fosforu deoxynukleosid trifosfátu; tím se získá vznik a fosfodiesterový most který připojí nový nukleotid a uvolní se pyrofosfát.[9]
Během replikace se současně vytvářejí dva typy vláken: přední vlákno, který je syntetizován nepřetržitě a roste směrem k replikační vidličce, a zaostávající vlákno, který je vyráběn diskontinuálně v Okazaki fragmenty a vyrůstá z replikační vidlice.[22] Fragmenty Okazaki jsou kovalentně připojil se DNA ligáza za vzniku spojitého vlákna.[22]Poté, aby se dokončila replikace DNA, se odstraní RNA primery a výsledné mezery se nahradí DNA a spojí se pomocí DNA ligázy.[22]
Aminokyseliny
Protein je polymer, který je složen z aminokyseliny které jsou propojeny pomocí peptidové vazby. Je tam více než 300 aminokyselin našel v přírodě, z nichž pouze dvacet, známý jako standardní aminokyseliny, jsou stavebními kameny pro bílkoviny.[24] Pouze zelené rostliny a většina mikroby jsou schopni syntetizovat všech 20 standardních aminokyselin, které jsou potřebné pro všechny živé druhy. Savci může syntetizovat pouze deset z dvaceti standardních aminokyselin. Ostatní aminokyseliny, valin, methionin, leucin, isoleucin, fenylalanin, lysin, threonin a tryptofan pro dospělé a histidin, a arginin pro kojence se získávají dietou.[25]
Základní struktura aminokyselin
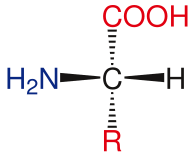
Obecná struktura standardních aminokyselin zahrnuje a primární aminoskupina, a karboxylová skupina a funkční skupina připojený k α-uhlík. Různé aminokyseliny jsou identifikovány funkční skupinou. Výsledkem tří různých skupin připojených k a-uhlíku jsou aminokyseliny asymetrické molekuly. Pro všechny standardní aminokyseliny, kromě glycin, α-uhlík je a chirální centrum. V případě glycinu má α-uhlík dva atomy vodíku, což této molekule dodává symetrii. S výjimkou prolin, všechny aminokyseliny nalezené v životě mají L-izoforma konformace. Prolin má funkční skupinu na a-uhlíku, která tvoří kruh s aminoskupinou.[24]

Zdroj dusíku
Jeden hlavní krok v biosyntéze aminokyselin zahrnuje začlenění dusíkové skupiny na a-uhlík. V buňkách existují dvě hlavní cesty začlenění dusíkových skupin. Jedna cesta zahrnuje enzym glutamin oxoglutarátaminotransferáza (GOGAT), který odstraní amide aminoskupina glutamin a přenese to na 2-oxoglutarát, produkovat dva glutamát molekuly. V této katalytické reakci slouží glutamin jako zdroj dusíku. Vpravo je obrázek ilustrující tuto reakci.
Další cesta pro zabudování dusíku na a-uhlík aminokyselin zahrnuje enzym glutamát dehydrogenáza (GDH). GDH je schopen převádět amoniak na 2-oxoglutarát a tvoří glutamát. Dále enzym glutamin syntetáza (GS) je schopen přenášet amoniak na glutamát a syntetizovat glutamin a doplňovat glutamin.[26]
Glutamátová rodina aminokyselin
The glutamát rodina aminokyselin zahrnuje aminokyseliny, které jsou odvozeny od aminokyseliny glutamátu. Tato rodina zahrnuje: glutamát, glutamin, prolin, a arginin. Tato rodina také zahrnuje aminokyselinu lysin, který je odvozen od α-ketoglutarát.[27]
Biosyntéza glutamátu a glutaminu je klíčovým krokem výše zmíněné asimilace dusíku. Enzymy GOGAT a GDH katalyzovat asimilace dusíku reakce.
V bakteriích je to enzym glutamát 5-kináza iniciuje biosyntézu prolinu přenosem fosfátové skupiny z ATP na glutamát. Další reakce je katalyzována enzymem pyrrolin-5-karboxylát syntáza (P5CS), který katalyzuje redukci ϒ-karboxyl skupina L-glutamát 5-fosfátu. To má za následek tvorbu glutamát semialdehydu, který spontánně cyklizuje na pyrrolin-5-karboxylát. Pyrrolin-5-karboxylát je dále redukován enzymem pyrrolin-5-karboxylát reduktázou (P5CR) za vzniku aminokyseliny prolin.[28]
V prvním kroku biosyntézy argininu v bakteriích je glutamát acetylovaný přenosem acetylové skupiny z acetyl-CoA v poloze N-a; tím se zabrání spontánní cyklizaci. Enzym N-acetylglutamát syntáza (glutamát N-acetyltransferáza) je zodpovědný za katalyzování kroku acetylace. Následující kroky jsou katalyzovány enzymy N-acetylglutamátkináza, N-acetyl-gama-glutamyl-fosfátreduktáza, a acetylornithin / sukcinyldiaminopimelát aminotransferáza a získá se N-acetyl-L-ornithin. Acetylová skupina acetylornithinu je odstraněna enzymem acetylornithináza (AO) nebo ornitin acetyltransferáza (OAT), a to přináší ornitin. Poté enzymy citrulin a argininosukcinát převést ornitin na arginin.[29]
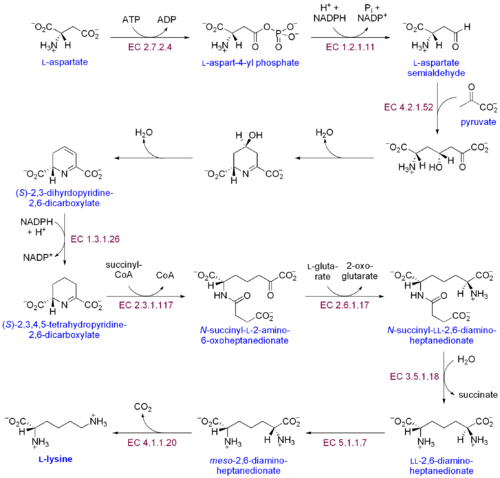
Existují dvě odlišné cesty biosyntézy lysinu: cesta kyseliny diaminopimelické a cesta dráha α-aminoadipátu. Nejběžnější ze dvou syntetických drah je cesta kyseliny diaminopimelické; sestává z několika enzymatických reakcí, které přidávají uhlíkové skupiny k aspartátu za vzniku lysinu:[30]
- Aspartát kináza iniciuje cestu kyseliny diaminopimelické fosforylací aspartátu a produkcí aspartylfosfátu.
- Aspartát semialdehyd dehydrogenáza katalyzuje NADPH -závislá redukce aspartylfosfátu za vzniku aspartát semialdehydu.
- 4-hydroxy-tetrahydrodipikolinát syntáza dodává pyruvát Skupina k p-aspartyl-4-semialdehydu a molekula vody je odstraněna. To způsobuje cyklizace a získá se (2S, 4S) -4-hydroxy-2,3,4,5-tetrahydrodipikolinát.
- 4-hydroxy-tetrahydrodipikolinátreduktáza katalyzuje redukci (2S, 4S) -4-hydroxy-2,3,4,5-tetrahydrodipikolinátu pomocí NADPH za vzniku A'-piperidein-2,6-dikarboxylátu (2,3,4,5-tetrahydrodipikolinátu) a H2Ó.
- Tetrahydrodipikolinát acyltransferáza katalyzuje acetylační reakci, která vede k otevření kruhu a poskytuje N-acetyl-a-amino-ε-ketopimelát.
- N-sukcinyl-a-amino-ε-ketopimelát-glutamátaminotransamináza katalyzuje transaminační reakci, která odstraňuje keto skupinu N-acetyl-a-amino-ε-ketopimelátu a nahrazuje ji aminoskupinou, čímž se získá N-sukcinyl-L-diaminopimelát.[31]
- N-acyldiaminopimelát deacyláza katalyzuje deacylaci N-sukcinyl-L-diaminopimelátu, čímž se získá L, L-diaminopimelát.[32]
- DAP epimeráza katalyzuje konverzi L, L-diaminopimelátu na mezo forma L, L-diaminopimelátu.[33]
- DAP dekarboxyláza katalyzuje odstranění karboxylové skupiny za vzniku L-lysinu.
Rodina serinových aminokyselin
The serin rodina aminokyselin zahrnuje: serin, cystein, a glycin. Většina mikroorganismů a rostlin získává síru pro syntézu methionin z aminokyseliny cystein. Kromě toho přeměna serinu na glycin poskytuje uhlíky potřebné pro biosyntézu methioninu a histidin.[27]
Během biosyntézy serinu[34] enzym fosfoglycerátdehydrogenáza katalyzuje počáteční reakci oxiduje 3-fosfo-D-glycerát poddat se 3-fosfonooxypyruvát.[35] Následující reakce je katalyzována enzymem fosfoserinaminotransferáza, který přenáší aminoskupinu z glutamátu na 3-fosfonooxypyruvát za vzniku L-fosfoserin.[36] Poslední krok je katalyzován enzymem fosfoserin fosfatáza, který defosforyláty L-fosfoserin za vzniku L-serin.[37]
Existují dvě známé cesty pro biosyntézu glycinu. Organismy, které používají ethanol a acetát jako hlavní zdroj uhlíku využívají glykonogenní cesta k syntéze glycin. Další cesta biosyntézy glycinu je známá jako glykolytický cesta. Tato cesta převádí serin syntetizovaný z meziproduktů glykolýza na glycin. V glykolytické dráze enzym serin hydroxymethyltransferáza katalyzuje štěpení serinu za vzniku glycinu a přenáší štěpenou uhlíkovou skupinu serinu na tetrahydrofolát, formování 5,10-methylen-tetrahydrofolát.[38]
Cysteinová biosyntéza je dvoustupňová reakce, která zahrnuje zabudování anorganické látky síra. V mikroorganismech a rostlinách enzym serinová acetyltransferáza katalyzuje přenos acetylové skupiny z acetyl-CoA na L-serin, čímž se získá O-acetyl-L-serin.[39] Následující reakční krok, katalyzovaný enzymem O-acetylserin (thiol) lyáza, nahrazuje acetylovou skupinu O-acetyl-L-serinu sulfidem za vzniku cysteinu.[40]
Aspartátová rodina aminokyselin
The aspartát rodina aminokyselin zahrnuje: threonin, lysin, methionin, isoleucin a aspartovat. Lysin a isoleucin jsou považovány za součást aspartátové rodiny, přestože je součástí jejich uhlíkové kostry pyruvát. V případě methioninu je methylový uhlík odvozen od serinu a skupiny síry, ale u většiny organismů je odvozen od cysteinu.[27]
Biosyntéza aspartátu je jednokroková reakce, která je katalyzována jediným enzymem. Enzym aspartátaminotransferáza katalyzuje přenos aminoskupiny z aspartátu na α-ketoglutarát za vzniku glutamátu a oxaloacetát.[41] Asparagin je syntetizován přídavkem aminoskupiny závislé na ATP na aspartát; asparagin syntetáza katalyzuje přidání dusíku z glutaminu nebo rozpustného amoniaku na aspartát za vzniku asparaginu.[42]

Biosyntetická cesta lysinu kyseliny diaminopimelické patří do rodiny aminokyselin aspartátu. Tato cesta zahrnuje devět enzymem katalyzovaných reakcí, které přeměňují aspartát na lysin.[43]
- Aspartát kináza katalyzuje počáteční krok v dráze kyseliny diaminopimelové přenosem a fosforyl z ATP na karboxylátovou skupinu aspartátu, čímž se získá aspartyl-p-fosfát.[44]
- Aspartát-semialdehyd dehydrogenáza katalyzuje redukční reakci o defosforylace aspartyl-p-fosfátu za vzniku aspartát-p-semialdehydu.[45]
- Dihydrodipikolinát syntáza katalyzuje kondenzace reakce aspartát-p-semialdehydu s pyruvátem za vzniku kyseliny dihydrodipikolinové.[46]
- 4-hydroxy-tetrahydrodipikolinátreduktáza katalyzuje redukci kyseliny dihydrodipikolinové za vzniku kyseliny tetrahydrodipikolinové.[47]
- Tetrahydrodipikolinát N-sukcinyltransferáza katalyzuje přenos sukcinylové skupiny ze sukcinyl-CoA na kyselinu tetrahydrodipikolinovou za vzniku N-sukcinyl-L-2,6-diaminoheptandioátu.[48]
- N-sukcinyldiaminopimelát aminotransferáza katalyzuje přenos aminoskupiny z glutamátu na N-sukcinyl-L-2,6-diaminoheptandioát za vzniku kyseliny N-sukcinyl-L, L-diaminopimelové.[49]
- Sukcinyl-diaminopimelát-desukcinyláza katalyzuje odstranění acylové skupiny z N-sukcinyl-L, L-diaminopimelové kyseliny, čímž se získá kyselina L, L-diaminopimelová.[50]
- Diaminopimelát epimeráza katalyzuje inverzi a-uhlíku kyseliny L, L-diaminopimelové za vzniku kyselina mesodiaminopimelová.[51]
- Siaminopimelát dekarboxyláza katalyzuje poslední krok v biosyntéze lysinu, který odstraňuje skupinu oxidu uhličitého z kyseliny mezodiaminopimelové za vzniku L-lysinu.[52]
Proteiny

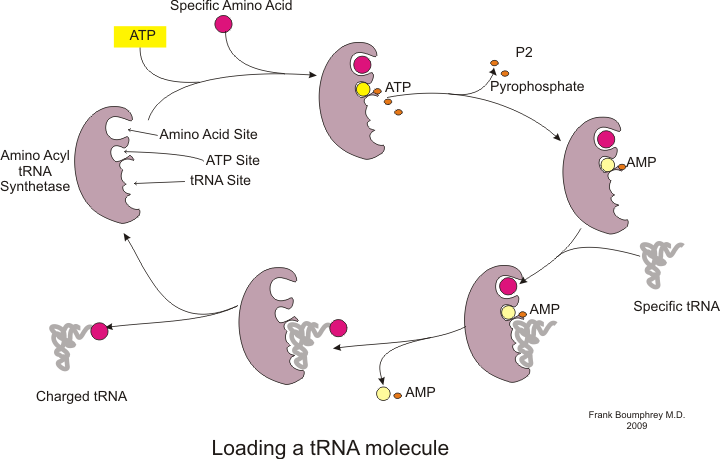
K syntéze bílkovin dochází prostřednictvím procesu zvaného překlad.[53] Během překladu volal genetický materiál mRNA čte ribozomy generovat protein polypeptid řetěz.[53] Tento proces vyžaduje přenos RNA (tRNA), která slouží jako adaptér vázáním aminokyseliny na jednom konci a interakce s mRNA na druhém konci; druhé párování mezi tRNA a mRNA zajišťuje, že je do řetězce přidána správná aminokyselina.[53] Syntéza proteinů probíhá ve třech fázích: zahájení, prodloužení a ukončení.[13] Prokaryotický (archaeal a bakteriální ) překlad se liší od eukaryotický překlad; tato část se však většinou zaměří na společné rysy obou organismů.
Další pozadí
Před zahájením translace musí dojít k procesu navázání specifické aminokyseliny na odpovídající tRNA. Tato reakce, nazývaná nabíjení tRNA, je katalyzována aminoacyl tRNA syntetáza.[54] Specifická tRNA syntetáza je zodpovědná za rozpoznání a nabití konkrétní aminokyseliny.[54] Kromě toho má tento enzym speciální diskriminační oblasti, které zajišťují správnou vazbu mezi tRNA a její příbuznou aminokyselinou.[54] Prvním krokem pro připojení aminokyseliny k její odpovídající tRNA je tvorba aminoacyl-AMP:[54]

Poté následuje přenos aminoacylové skupiny z aminoacyl-AMP do molekuly tRNA. Výsledná molekula je aminoacyl-tRNA:[54]

Kombinace těchto dvou kroků, z nichž oba jsou katalyzovány aminoacyl tRNA syntetázou, produkuje nabitou tRNA, která je připravena přidávat aminokyseliny do rostoucího polypeptidového řetězce.
Kromě vazby na aminokyselinu má tRNA tří nukleotidovou jednotku nazvanou antikodon že základní páry se specifickými nukleotidovými triplety na mRNA kodony; kodony kódují specifickou aminokyselinu.[55] Tato interakce je možná díky ribozomu, který slouží jako místo pro syntézu bílkovin. Ribozom má tři vazebná místa pro tRNA: aminoacylové místo (A místo), peptidylové místo (P místo) a výstupní místo (E místo).[56]
V přepisu mRNA je mnoho kodonů a je velmi běžné, že aminokyselina je specifikována více než jedním kodonem; tento jev se nazývá zvrhlost.[57] Celkově existuje 64 kodonů, 61 z každého kódu pro jednu z 20 aminokyselin, zatímco zbývající kodony specifikují ukončení řetězce.[57]
Překlad v krocích
Jak již bylo zmíněno dříve, překlad probíhá ve třech fázích: zahájení, prodloužení a ukončení.
Krok 1: Zahájení
Dokončení iniciační fáze závisí na následujících třech událostech:[13]
1. Nábor ribozomu do mRNA
2. Vazba nabitého iniciátoru tRNA na P místo ribozomu
3. Správné zarovnání ribozomu s počátečním kodonem mRNA
Krok 2: Prodloužení
Po zahájení je polypeptidový řetězec prodloužen prostřednictvím interakcí antikodon: kodon, přičemž ribozom přidává aminokyseliny do polypeptidového řetězce po jedné. K zajištění správného přidání aminokyselin musí dojít k následujícím krokům:[58]
1. Vazba správné tRNA na místo A ribozomu
2. Vznik a peptidová vazba mezi tRNA v místě A a polypeptidovým řetězcem připojeným k tRNA v místě P
3. Přemístění nebo postup komplexu tRNA-mRNA o tři nukleotidy
Translokace „nastartuje“ tRNA v místě E a posune tRNA z místa A do místa P a ponechá místo A volné pro příchozí tRNA pro přidání další aminokyseliny.
Krok 3: Ukončení
Poslední fáze překladu nastane, když a stop kodon vstupuje na web A.[1] Poté nastanou následující kroky:
1. Uznání kodonů uvolňovací faktory, který způsobí hydrolýza polypeptidového řetězce z tRNA umístěné v místě P[1]
2. Uvolnění polypeptidového řetězce[57]
3. Disociace a „recyklace“ ribozomu pro budoucí překladové procesy[57]
Souhrnná tabulka klíčových hráčů v překladu je uvedena níže:
| Klíčoví hráči v překladu | Fáze překladu | Účel |
|---|---|---|
| tRNA syntetáza | před zahájením | Odpovídá za nabíjení tRNA |
| mRNA | zahájení, prodloužení, ukončení | Šablona pro syntézu proteinů; obsahuje oblasti pojmenované kodony, které kódují aminokyseliny |
| tRNA | zahájení, prodloužení, ukončení | Spojuje místa ribozomů A, P, E; páry antikodonových bází s kodonem mRNA, aby bylo zajištěno, že do rostoucího polypeptidového řetězce je začleněna správná aminokyselina |
| ribozom | zahájení, prodloužení, ukončení | Řídí syntézu proteinů a katalyzuje tvorbu peptidové vazby |
Nemoci spojené s nedostatkem makromolekul

Chyby v biosyntetických drahách mohou mít škodlivé následky, včetně malformace makromolekul nebo nedostatečné produkce funkčních molekul. Níže jsou uvedeny příklady, které ilustrují narušení, ke kterým dochází v důsledku těchto neúčinností.
- Familiární hypercholesterolemie: tato porucha je charakterizována absencí funkčnosti receptory pro LDL.[59] Nedostatky ve tvorbě LDL receptorů mohou způsobit vadné receptory, které narušují endocytární cesta, inhibující vstup LDL do jater a dalších buněk.[59] To způsobuje hromadění LDL v krevní plazmě, což má za následek aterosklerotické plaky které zužují tepny a zvyšují riziko infarktu.[59]
- Lesch-Nyhanův syndrom: toto genetické onemocnění je charakterizováno sebepoškozování duševní nedostatek a dna.[60] Je to způsobeno absencí hypoxanthin-guanin fosforibosyltransferáza, což je nezbytný enzym pro tvorbu purinových nukleotidů.[60] Nedostatek enzymu snižuje hladinu nezbytných nukleotidů a způsobuje akumulaci biosyntézy meziprodukty, což má za následek výše uvedené neobvyklé chování.[60]
- Těžká kombinovaná imunodeficience (SCID): SCID se vyznačuje ztrátou T buňky.[61] Nedostatek těchto složek imunitního systému zvyšuje náchylnost k infekčním agens, protože postižené osoby se nemohou vyvinout imunologická paměť.[61] Tato imunologická porucha je výsledkem nedostatku adenosin deanimáza aktivita, která způsobí nárůst DATP. Tyto molekuly dATP pak inhibují ribonukleotidreduktázu, která brání syntéze DNA.[61]
- Huntingtonova choroba: tento neurologický onemocnění je způsobeno chybami, ke kterým dochází během syntézy DNA.[62] Tyto chyby nebo mutace vedou k expresi mutanta huntingtin protein, který obsahuje opakující se glutamin zbytky, které jsou kódovány rozbalením Opakuje se trinukleotid CAG v gen.[62] Huntingtonova choroba je charakterizována ztrátou neuronů a glióza. Mezi příznaky onemocnění patří: porucha hybnosti, poznávací pokles a poruchy chování.[63]
Viz také
- Lipidy
- Fosfolipidová dvojvrstva
- Nukleotidy
- DNA
- replikace DNA
- Proteinogenní aminokyselina
- Codonův stůl
- Prostaglandin
- Porfyriny
- Chlorofyly a bakteriochlorofyly
- Vitamin B12
Reference
- ^ A b C d Alberts, Bruce (2007). Molekulární biologie buňky. New York: Garland Science. ISBN 978-0815341055.
- ^ Zumdahl, Steven S. Zumdahl, Susan A. (2008). Chemie (8. vydání). CA: Cengage Learning. ISBN 978-0547125329.
- ^ Pratt, Donald Voet, Judith G. Voet, Charlotte W. (2013). Základy biochemie: život na molekulární úrovni (4. vydání). Hoboken, NJ: Wiley. ISBN 978-0470547847.
- ^ A b C d E Lodish, Harvey; et al. (2007). Molekulární buněčná biologie (6. vydání). New York: W.H. Freemane. ISBN 978-0716743668.
- ^ A b C d E Cox, David L. Nelson, Michael M. (2008). Lehningerovy principy biochemie (5. vydání). New York: W.H. Freemane. ISBN 9780716771081.
- ^ Hanin, Izrael (2013). Fosfolipidy: biochemické, farmaceutické a analytické úvahy. Springer. ISBN 978-1475713664.
- ^ A b C d E Vance, Dennis E .; Vance, Jean E. (2008). Biochemie lipidů, lipoproteinů a membrán (5. vydání). Amsterdam: Elsevier. ISBN 978-0444532190.
- ^ Katsaras, J .; et al. (2001). Lipidové dvojvrstvy: struktura a interakce; se 6 stoly. Berlin [u.a.]: Springer. ISBN 978-3540675556.
- ^ A b C d E Stryer, Jeremy M. Berg; John L. Tymoczko; Lubert (2007). Biochemie (6. vydání, 3. vydání, vydání). New York: Freeman. ISBN 978-0716787242.
- ^ Gault, ČR; LM Obeid; YA Hannun (2010). Přehled metabolismu sfingolipidů: od syntézy po rozpad. Adv Exp Med Biol. Pokroky v experimentální medicíně a biologii. 688. s. 1–23. doi:10.1007/978-1-4419-6741-1_1. ISBN 978-1-4419-6740-4. PMC 3069696. PMID 20919643.
- ^ A b Siegel, George J. (1999). Základní neurochemie: molekulární, buněčné a lékařské aspekty (6. vyd.). Philadelphia, PA [USA]: Lippincott Williams & Wilkins. ISBN 978-0397518203.
- ^ A b C Harris, J. Robin (2010). Vazba cholesterolu a transportní proteiny cholesterolu: struktura a funkce ve zdraví a nemoci. Dordrecht: Springer. ISBN 978-9048186211.
- ^ A b C d Watson, James D .; et al. (2007). Molekulární biologie genu (6. vydání). San Francisco, Kalifornie: Benjamin Cummings. ISBN 978-0805395921.
- ^ Kappock, TJ; Ealick, SE; Stubbe, J (říjen 2000). "Modulární vývoj purinové biosyntetické dráhy". Aktuální názor na chemickou biologii. 4 (5): 567–72. doi:10.1016 / s1367-5931 (00) 00133-2. PMID 11006546.
- ^ Sampei, G; Baba, S; Kanagawa, M; Yanai, H; Ishii, T; Kawai, H; Fukai, Y; Ebihara, A; Nakagawa, N; Kawai, G (říjen 2010). "Krystalové struktury glycinamid ribonukleotid syntetázy, PurD, z termofilních eubakterií". Journal of Biochemistry. 148 (4): 429–38. doi:10.1093 / jb / mvq088. PMID 20716513.
- ^ Hoskins, AA; Anand, R; Ealick, SE; Stubbe, J (17. srpna 2004). „Komplex formylglycinamid ribonukleotid amidotransferázy z Bacillus subtilis: tvorba komplexu zprostředkovaného metabolity“. Biochemie. 43 (32): 10314–27. doi:10.1021 / bi049127h. PMID 15301530.
- ^ Mueller, EJ; Meyer, E; Rudolph, J; Davisson, VJ; Stubbe, J (1. března 1994). „N5-karboxyaminoimidazol ribonukleotid: důkaz nového meziproduktu a dvou nových enzymatických aktivit v de novo purinové biosyntetické dráze Escherichia coli“. Biochemie. 33 (8): 2269–78. doi:10.1021 / bi00174a038. PMID 8117684.
- ^ Firestine, SM; Poon, SW; Mueller, EJ; Stubbe, J; Davisson, VJ (4. října 1994). „Reakce katalyzované 5-aminoimidazolovými ribonukleotidovými karboxylázami z Escherichia coli a Gallus gallus: případ odlišných katalytických mechanismů“. Biochemie. 33 (39): 11927–34. doi:10.1021 / bi00205a031. PMID 7918411.
- ^ A b Srere, PA (1987). "Komplexy sekvenčních metabolických enzymů". Roční přehled biochemie. 56 (1): 89–124. doi:10.1146 / annurev.bi.56.070187.000513. PMID 2441660.
- ^ Broach, editoval Jeffrey N. Strathern, Elizabeth W. Jones, James R. (1981). Molekulární biologie kvasinek Saccharomyces. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. ISBN 978-0879691394.CS1 maint: další text: seznam autorů (odkaz)
- ^ A b C O'Donovan, GA; Neuhard, J (září 1970). "Metabolismus pyrimidinu v mikroorganismech". Bakteriologické recenze. 34 (3): 278–343. doi:10.1128 / MMBR.34.3.278-343.1970. PMC 378357. PMID 4919542.
- ^ A b C d Geer, Gerald Karp; odpovědný za revizi kapitoly 15 Peter van der (2004). Buněčná a molekulární biologie: koncepty a experimenty (4. vydání, Wiley International ed.). New York: J. Wiley & Sons. ISBN 978-0471656654.
- ^ A b C d Griffiths, Anthony J. F. (1999). Moderní genetická analýza (2. tisk. Vyd.). New York: Freeman. ISBN 978-0716731184.
- ^ A b Wu, G (květen 2009). "Aminokyseliny: metabolismus, funkce a výživa". Aminokyseliny. 37 (1): 1–17. doi:10.1007 / s00726-009-0269-0. PMID 19301095. S2CID 1870305.
- ^ Mousdale, D. M .; Coggins, J. R. (1991). Syntéza aminokyselin. Cílové stránky pro herbicidní opatření. str. 29–56. doi:10.1007/978-1-4899-2433-9_2. ISBN 978-1-4899-2435-3.
- ^ Miflin, B. J .; Lea, P. J. (1977). "Metabolismus aminokyselin". Roční přehled fyziologie rostlin. 28: 299–329. doi:10.1146 / annurev.pp.28.060177.001503.
- ^ A b C Umbarger, HE (1978). "Biosyntéza aminokyselin a její regulace". Roční přehled biochemie. 47 (1): 532–606. doi:10.1146 / annurev.bi.47.070178.002533. PMID 354503.
- ^ Pérez-Arellano, I; Carmona-Alvarez, F; Martínez, AI; Rodríguez-Díaz, J; Cervera, J (březen 2010). „Pyrrolin-5-karboxylát syntáza a prolinová biosyntéza: od osmotolerance ke vzácnému metabolickému onemocnění“. Věda o bílkovinách. 19 (3): 372–82. doi:10,1002 / pro.340. PMC 2866264. PMID 20091669.
- ^ Xu, Y; Labedan, B; Glansdorff, N (březen 2007). „Překvapivá biosyntéza argininu: přehodnocení enzymologie a vývoje dráhy v mikroorganismech“. Recenze mikrobiologie a molekulární biologie. 71 (1): 36–47. doi:10.1128 / MMBR.00032-06. PMC 1847373. PMID 17347518.
- ^ "MetaCyc: biosyntéza L-lysinu I".
- ^ PETERKOFSKY, B; GILVARG, C (květen 1961). "N-sukcinyl-L-diaminopimelic-glutamová transamináza". The Journal of Biological Chemistry. 236: 1432–8. PMID 13734750.
- ^ KINDLER, SH; GILVARG, C (prosinec 1960). „Deacyláza N-sukcinyl-L-2,6-diaminopimelové kyseliny“. The Journal of Biological Chemistry. 235: 3532–5. PMID 13756049.
- ^ Born, TL; Blanchard, JS (říjen 1999). "Structure/function studies on enzymes in the diaminopimelate pathway of bacterial cell wall biosynthesis". Aktuální názor na chemickou biologii. 3 (5): 607–13. doi:10.1016/s1367-5931(99)00016-2. PMID 10508663.
- ^ "Escherichia coli K-12 substr. MG1655". serine biosynthesis. SRI International. Citováno 12. prosince 2013.
- ^ Bell, JK; Grant, GA; Banaszak, LJ (Mar 30, 2004). "Multiconformational states in phosphoglycerate dehydrogenase". Biochemie. 43 (12): 3450–8. doi:10.1021/bi035462e. PMID 15035616.
- ^ Dubnovitsky, AP; Kapetaniou, EG; Papageorgiou, AC (January 2005). "Enzyme adaptation to alkaline pH: atomic resolution (1.08 A) structure of phosphoserine aminotransferase from Bacillus alcalophilus". Věda o bílkovinách. 14 (1): 97–110. doi:10.1110/ps.041029805. PMC 2253317. PMID 15608117.
- ^ Wang, W; Kim, R; Jancarik, J; Yokota, H; Kim, SH (Jan 10, 2001). "Crystal structure of phosphoserine phosphatase from Methanococcus jannaschii, a hyperthermophile, at 1.8 A resolution". Struktura. 9 (1): 65–71. doi:10.1016/s0969-2126(00)00558-x. PMID 11342136.
- ^ Monschau, N; Stahmann, KP; Sahm, H; McNeil, JB; Bognar, AL (May 1, 1997). "Identification of Saccharomyces cerevisiae GLY1 as a threonine aldolase: a key enzyme in glycine biosynthesis". Mikrobiologické dopisy FEMS. 150 (1): 55–60. doi:10.1111/j.1574-6968.1997.tb10349.x. PMID 9163906.
- ^ Pye, VE; Tingey, AP; Robson, RL; Moody, PC (Sep 24, 2004). "The structure and mechanism of serine acetyltransferase from Escherichia coli". The Journal of Biological Chemistry. 279 (39): 40729–36. doi:10.1074/jbc.M403751200. PMID 15231846.
- ^ Huang, B; Vetting, MW; Roderick, SL (May 2005). "The active site of O-acetylserine sulfhydrylase is the anchor point for bienzyme complex formation with serine acetyltransferase". Journal of Bacteriology. 187 (9): 3201–5. doi:10.1128/JB.187.9.3201-3205.2005. PMC 1082839. PMID 15838047.
- ^ McPhalen, CA; Vincent, MG; Picot, D; Jansonius, JN; Lesk, AM; Chothia, C (Sep 5, 1992). "Uzavření domény v mitochondriální aspartátaminotransferáze". Journal of Molecular Biology. 227 (1): 197–213. doi:10.1016 / 0022-2836 (92) 90691-C. PMID 1522585.
- ^ Larsen, TM; Boehlein, SK; Schuster, SM; Richards, NG; Thoden, JB; Holden, HM; Rayment, I (Dec 7, 1999). "Three-dimensional structure of Escherichia coli asparagine synthetase B: a short journey from substrate to product". Biochemie. 38 (49): 16146–57. CiteSeerX 10.1.1.453.5998. doi:10.1021/bi9915768. PMID 10587437.
- ^ Velasco, AM; Leguina, JI; Lazcano, A (October 2002). "Molekulární vývoj biosyntetických drah lysinu". Journal of Molecular Evolution. 55 (4): 445–59. doi:10.1007 / s00239-002-2340-2. PMID 12355264. S2CID 19460256.
- ^ Kotaka, M; Ren, J; Lockyer, M; Hawkins, AR; Stammers, DK (Oct 20, 2006). "Structures of R- and T-state Escherichia coli aspartokinase III. Mechanisms of the allosteric transition and inhibition by lysine". The Journal of Biological Chemistry. 281 (42): 31544–52. doi:10,1074 / jbc.M605886200. PMID 16905770.
- ^ Hadfield, A; Kryger, G; Ouyang, J; Petsko, GA; Ringe, D; Viola, R (Jun 18, 1999). "Structure of aspartate-beta-semialdehyde dehydrogenase from Escherichia coli, a key enzyme in the aspartate family of amino acid biosynthesis". Journal of Molecular Biology. 289 (4): 991–1002. doi:10.1006/jmbi.1999.2828. PMID 10369777.
- ^ Mirwaldt, C; Korndörfer, I; Huber, R (Feb 10, 1995). "The crystal structure of dihydrodipicolinate synthase from Escherichia coli at 2.5 A resolution". Journal of Molecular Biology. 246 (1): 227–39. doi:10.1006/jmbi.1994.0078. PMID 7853400.
- ^ Cirilli, M; Zheng, R; Scapin, G; Blanchard, JS (Sep 16, 2003). "The three-dimensional structures of the Mycobacterium tuberculosis dihydrodipicolinate reductase-NADH-2,6-PDC and -NADPH-2,6-PDC complexes. Structural and mutagenic analysis of relaxed nucleotide specificity". Biochemie. 42 (36): 10644–50. doi:10.1021/bi030044v. PMID 12962488.
- ^ Beaman, TW; Binder, DA; Blanchard, JS; Roderick, SL (Jan 21, 1997). "Three-dimensional structure of tetrahydrodipicolinate N-succinyltransferase". Biochemie. 36 (3): 489–94. doi:10.1021/bi962522q. PMID 9012664.
- ^ Weyand, S; Kefala, G; Weiss, MS (Mar 30, 2007). "The three-dimensional structure of N-succinyldiaminopimelate aminotransferase from Mycobacterium tuberculosis". Journal of Molecular Biology. 367 (3): 825–38. doi:10.1016/j.jmb.2007.01.023. PMID 17292400.
- ^ Nocek, BP; Gillner, DM; Fan, Y; Holz, RC; Joachimiak, A (Apr 2, 2010). "Structural basis for catalysis by the mono- and dimetalated forms of the dapE-encoded N-succinyl-L,L-diaminopimelic acid desuccinylase". Journal of Molecular Biology. 397 (3): 617–26. doi:10.1016/j.jmb.2010.01.062. PMC 2885003. PMID 20138056.
- ^ Pillai, B; Cherney, M; Diaper, CM; Sutherland, A; Blanchard, JS; Vederas, JC; James, MN (Nov 23, 2007). "Dynamics of catalysis revealed from the crystal structures of mutants of diaminopimelate epimerase". Sdělení o biochemickém a biofyzikálním výzkumu. 363 (3): 547–53. doi:10.1016/j.bbrc.2007.09.012. PMID 17889830.
- ^ Gokulan, K; Rupp, B; Pavelka MS, Jr; Jacobs WR, Jr; Sacchettini, JC (May 16, 2003). "Crystal structure of Mycobacterium tuberculosis diaminopimelate decarboxylase, an essential enzyme in bacterial lysine biosynthesis". The Journal of Biological Chemistry. 278 (20): 18588–96. doi:10.1074/jbc.M301549200. PMID 12637582.
- ^ A b C Weaver, Robert F. (2005). Molekulární biologie (3. vyd.). Boston: McGraw-Hill Higher Education. ISBN 978-0-07-284611-9.
- ^ A b C d E Cooper, Geoffrey M. (2000). Buňka: molekulární přístup (2. vyd.). Washington (DC): ASM Press. ISBN 978-0878931064.
- ^ Jackson, R.J.; et al. (Únor 2010). "The mechanism of eukaryotic translation initiation and principles of its regulation". Molekulární buněčná biologie. 10: 113–127. PMC 4461372. PMID 20094052.
- ^ Zelená, Rachel; Harry F. Noller; et al. (1997). "Ribosomes and Translation". Annu. Biochem. 66: 679–716. doi:10.1146/annurev.biochem.66.1.679. PMID 9242921.
- ^ A b C d Pestka (editors), Herbert Weissbach, Sidney (1977). Molecular Mechanisms of protein biosynthesis. New York: Academic Press. ISBN 978-0127442501.CS1 maint: další text: seznam autorů (odkaz)
- ^ Frank, J; Haixiao Gao; et al. (Září 2007). "The process of mRNA–tRNA translocation". PNAS. 104 (50): 19671–19678. doi:10.1073/pnas.0708517104. PMC 2148355. PMID 18003906.
- ^ A b C Bandeali, Salman J.; Daye, Jad; Virani, Salim S. (30 November 2013). "Novel Therapies for Treating Familial Hypercholesterolemia". Current Atherosclerosis Reports. 16 (1): 382. doi:10.1007/s11883-013-0382-0. PMID 24293346. S2CID 8903481.
- ^ A b C Kang, Tae Hyuk; Park, Yongjin; Bader, Joel S.; Friedmann, Theodore; Cooney, Austin John (9 October 2013). "The Housekeeping Gene Hypoxanthine Guanine Phosphoribosyltransferase (HPRT) Regulates Multiple Developmental and Metabolic Pathways of Murine Embryonic Stem Cell Neuronal Differentiation". PLOS ONE. 8 (10): e74967. doi:10.1371/journal.pone.0074967. PMC 3794013. PMID 24130677.
- ^ A b C Walport, Ken Murphy, Paul Travers, Mark (2011). Janewayova imunobiologie (8. vyd.). Oxford: Taylor & Francis. ISBN 978-0815342434.
- ^ A b Hughes, edited by Donald C. Lo, Robert E. (2010). Neurobiology of Huntington's disease : applications to drug discovery (2. vyd.). Boca Raton: CRC Press/Taylor & Francis Group. ISBN 978-0849390005.CS1 maint: další text: seznam autorů (odkaz)
- ^ Biglan, Kevin M.; Ross, Christopher A.; Langbehn, Douglas R.; Aylward, Elizabeth H.; Stout, Julie C.; Queller, Sarah; Carlozzi, Noelle E.; Duff, Kevin; Beglinger, Leigh J.; Paulsen, Jane S. (26 June 2009). "Motor abnormalities in premanifest persons with Huntington's disease: The PREDICT-HD study". Poruchy pohybu. 24 (12): 1763–1772. doi:10.1002/mds.22601. PMC 3048804. PMID 19562761.
Pobočky chemie | |
|---|---|
| Fyzický | |
| Organické | |
| Anorganické | |
| Analytické | |
| Ostatní | |
| Viz také | |
| |