Blokování kanálů - Channel blocker

A blokátor kanálů je biologický mechanismus, ve kterém se konkrétní molekula používá k zabránění otevření iontových kanálů za účelem produkce a fyziologický reakce v buňce. Blokování kanálů je prováděno různými typy molekul, jako jsou kationty, anionty, aminokyseliny a další chemikálie. Tyto blokátory fungují jako iontový kanál antagonisté, zabraňující odezvě, kterou obvykle poskytuje otevření kanálu.
Iontové kanály umožňují selektivní průchod iontů přes buněčné membrány využitím proteinů, které fungují jako póry, které umožňují průchod elektrického náboje dovnitř a ven z buňky.[1] Tyto iontové kanály jsou nejčastěji brány, což znamená, že vyžadují zvláštní stimul, který způsobí otevření a uzavření kanálu. Tyto typy iontových kanálů regulují tok nabitých iontů přes membránu, a proto zprostředkovávají membránový potenciál buňky.
Molekuly, které působí jako blokátory kanálů, jsou důležité v oblasti farmakologie, protože velkou částí designu léčiv je použití antagonistů iontových kanálů při regulaci fyziologické odpovědi. Specifičnost molekul blokujících kanál na určitých kanálech z něj činí cenný nástroj při léčbě mnoha poruch.[2][3]
Pozadí
Iontové kanály
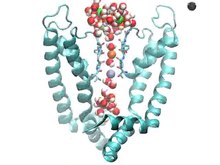
Pro pochopení mechanismu blokátorů kanálů je důležité pochopit složení iontových kanálů. Jejich hlavní funkcí je přispívat k klidový membránový potenciál buňky tokem ionty přes buněčnou membránu. K dosažení tohoto úkolu musí být ionty schopné překročit hydrofobní oblast a lipidová dvojvrstva nepříznivý proces. Na pomoc při transportu iontů tvoří iontové kanály hydrofilní póry přes membránu, což umožňuje obvykle nepříznivý přenos hydrofilních molekul.[4] Různé iontové kanály mají různé mechanismy funkce. Obsahují:
- napěťově řízené iontové kanály
- Iontové kanály, které jsou aktivovány změnami membránového potenciálu
- iontový kanál řízený ligandem
- Iontové kanály zprostředkované vazbou malých molekul na protein kanálu
- mechanosenzitivní iontové kanály
- Iontové kanály, které reagují na protažení, vibrace nebo změny teploty
- světelné iontové kanály
- Iontové kanály, které se otevírají nebo zavírají v reakci na světlo
Molekuly, které působí jako blokátory iontových kanálů, lze použít ve vztahu k některému z těchto různých kanálů. Například sodíkové kanály, které jsou nezbytné pro výrobu akční potenciály, jsou ovlivněni mnoha různými toxiny. Tetrodotoxin (TTX), toxin nacházející se v pufferfish, zcela blokuje transport iontů sodíku blokováním oblasti selektivního filtru kanálu.[5] Velká část struktury pórů iontových kanálů byla objasněna ze studií, které používají toxiny k inhibici funkce kanálu.[6][7][8]
Identita
Nástroje jako Rentgenová krystalografie a elektrofyziologie byly nezbytné při lokalizaci vazebných míst molekul bloků s otevřeným kanálem. Studiem biologického a chemického složení iontových kanálů mohou vědci určit složení molekul, které se vážou na určité oblasti. Rentgenová krystalografie poskytuje strukturální obraz dotyčného kanálu a molekuly.[9] Stanovení hydrofobicity domén kanálu prostřednictvím hydrofobní grafy také poskytuje informace o chemickém složení molekuly a o tom, proč se váže na určitou oblast. Například pokud se protein váže na hydrofobní oblast kanálu (a proto má transmembránovou oblast), může být dotyčná molekula složena z aminokyselin alanin, leucin nebo fenylalanin, protože jsou všichni sami hydrofobní.[10] Elektrofyziologie je také důležitým nástrojem při identifikaci struktury kanálu, protože analýza iontových faktorů, které vedou k aktivaci kanálu, může být rozhodující pro pochopení inhibičních účinků molekul bloku otevřeného kanálu.[3][9]
Fyziologie

Antagonista receptoru
Blokátory kanálů jsou antagonisté kanálů potřebných k produkci normální fyziologické funkce v buňkách. Mnoho kanálů má vazebná místa pro regulační prvky, které mohou podporovat nebo potlačovat normální funkci v závislosti na požadavcích v buňce a organismu. Normální funkcí vazby agonistů je generování buněčných změn vedoucích k různým následným účinkům; tyto účinky se pohybují od změny membránového potenciálu po zahájení signalizační kaskády.[11] Naopak, když se blokátory otevřeného kanálu vážou na buňku, brání normální funkci vazby agonisty. Například, napěťově řízené kanály otevírají a zavírají se na základě membránového potenciálu a jsou rozhodující při vytváření akčních potenciálů tím, že umožňují iontům stékat po stanovených gradientech. Blokátory otevřených kanálů se však mohou na tyto kanály vázat, aby zabránily proudění iontů, čímž zabrání iniciaci akčního potenciálu.[12]
Specifičnost molekul
Mnoho různých organické sloučeniny mohou působit jako blokátory kanálů navzdory specifičnosti kanálu. Kanály si vyvinuly struktury, které díky svým oblastem překlenujícím membránu mohou rozlišovat mezi různými ionty nebo sloučeninami. Například některé objekty jsou příliš velké na to, aby se vešly do kanálů, které jsou strukturně specifikovány pro přepravu menších objektů, jako je například iont draslíku, který se pokouší vejít do sodíkového kanálu. Naopak, některé objekty jsou příliš malé na to, aby byly řádně stabilizovány určitými póry kanálu, například sodíkový iont, který se pokouší projít draslíkovým kanálem.[11][13] V obou případech není tok kanálu povolen. Pokud však určitá sloučenina vykazuje adekvátní chemickou afinitu ke kanálu, může být tato sloučenina schopna vázat a blokovat póry kanálu. Například TTX může vázat a inaktivovat napěťově řízené sodíkové kanály, a to navzdory skutečnosti, že TTX je mnohem větší a chemicky odlišný od sodíkových iontů. Vzhledem k rozdílům ve velikosti a chemických vlastnostech mezi TTX a sodíkovým iontem se jedná o příklad struktury používané k blokování obvykle specifických kanálů. [14]
Kinetika
Blok kanálu může být indukován mnoha různými typy organických sloučenin, pokud se mohou vázat na určitou část pórů cílového kanálu. Kinetika blokátorů kanálů je primárně chápána, i když je používána jako anestetika. Lokální anestetika fungují indukcí stavu fázového bloku v cílených neuronech.[13] Zpočátku blokátory otevřeného kanálu účinně nezabraňují akčním potenciálům, protože je blokováno jen málo kanálů a samotný blokátor lze z kanálu uvolnit buď rychle, nebo pomalu v závislosti na jeho vlastnostech. Fázické bloky se však vyskytují, protože opakovaná depolarizace zvyšuje afinitu blokátorů ke kanálům v neuronu. Za tuto akci je zodpovědná kombinace zvýšení dostupných kanálů a změny konformace kanálu ke zvýšení vazebné afinity blokátoru.[13][15][16]
Klinický význam
Terapeutické použití
Rozličný neurodegenerativní onemocnění byly spojeny s nadměrným NMDA receptor aktivace znamená zprostředkování neurotoxicity závislé na vápníku. Vědci zkoumali mnoho různých antagonistů NMDA a jejich terapeutickou účinnost, přičemž žádný z nich neuzavřel bezpečnost ani účinnost.[17] Vědci roky zkoumali účinky bloku otevřeného kanálu, memantin, jako možnost léčby neurotoxicity. Předpokládali, že rychlejší rychlosti blokování a odblokování a celková kinetika memantinu mohou být základním důvodem klinické tolerance.[17][3] Jako nekonkurenční antagonista by memantin měl navzdory vysoké hladině přiblížit hladinu NMDA normálním hodnotám glutamát koncentrace. Na základě těchto informací vědci spekulovali, že jednoho dne může být memantin použit jako blok otevřeného kanálu, aby se zabránilo zvýšení hladiny glutamátu spojené s neurotoxicitou s malými nebo žádnými vedlejšími účinky ve srovnání s jinými možnostmi léčby.[17]
Alzheimerova choroba
Alzheimerova choroba, je spojena se specifickou neurodegenerativní poruchou glutaminergní neurotransmise přerušení, o nichž se předpokládá, že vedou k základním kognitivním příznakům Alzheimerovy choroby.[18][2][3] Vědci naznačují, že nekompetitivní agonisty NMDA receptorů lze použít jako pomoc při léčbě těchto příznaků, aniž by docházelo k závažným vedlejším účinkům.[18] Jako jeden z mála léčivých přípravků schválených pro léčbu Alzheimerovy choroby bylo prokázáno, že memantin umožňuje, aby excitační postsynaptické proudy zůstaly nedotčeny, a zároveň snižuje výskyt a amplitudu inhibičních postsynaptických proudů.[19] Důkazy podporují hypotézu, že jak silná závislost na napětí, tak rychlá kinetika memantinu mohou být zodpovědné za snížené vedlejší účinky a kognitivní pokrok.[20]
Cystická fibróza
Cystická fibróza je progresivní genetické onemocnění spojené s transmembránovým regulátorem CF (CFTR ) dysfunkce.[21] Blokování tohoto kanálu určitými cytoplazmatickými, záporně nabitými látkami má za následek snížený transport chloridových iontů a hydrogenuhličitanových aniontů, jakož i sníženou sekreci tekutin a solí. To má za následek nahromadění hustého hlenu, který je charakteristický pro cystickou fibrózu.[21]
Farmakologie
Anestetika
Blokátory kanálů jsou v oblasti anestetik zásadní. Inhibitory sodíkového kanálu se používají jako oba antiepileptika a antiarytmika, protože mohou inhibovat hyperexcitovatelné tkáně u pacienta.[22] Zavedení specifických blokátorů sodíkového kanálu do tkáně umožňuje preferenční vazbu blokátoru na sodíkové kanály, což vede k konečné inhibici toku sodíku do tkáně. V průběhu času tento mechanismus vede k celkovému snížení excitace tkáně. Prodloužená hyperpolarizace přerušuje normální zotavení kanálu a umožňuje neustálou inhibici a poskytuje dynamickou kontrolu anestetik v daném prostředí.[22]
Alzheimerova choroba
Nadměrná expozice glutamátu vede k neurotoxicitě u pacientů s Alzheimerovou chorobou. Konkrétně byla nadměrná aktivace glutamátových receptorů typu NMDA spojena s excitotoxicitou nervových buněk a buněčnou smrtí.[18][2] Potenciálním řešením je snížení aktivity NMDA receptoru, aniž by došlo k tak drastickým interferencím, které by způsobily klinické vedlejší účinky.[23]
Ve snaze zabránit další neurodegeneraci vědci použili memantin, blok otevřeného kanálu, jako formu léčby. Užívání memantinu dosud u pacientů s Alzheimerovou chorobou rychle vede ke klinickému pokroku u mnoha různých příznaků. Předpokládá se, že memantin pracuje efektivně díky své schopnosti rychle upravit jeho kinetiku, což brání nahromadění v kanálu a umožňuje normální synaptický přenos. Bylo zjištěno, že jiné blokátory kanálů blokují veškerou aktivitu NMDA receptoru, což vede k nežádoucím klinickým vedlejším účinkům.[3]
Dysfunkce kanálu CFTR
Transmembránové regulátory cystické fibrózy (CFTR) fungují při transportu chloridových iontů, hydrogenuhličitanů a tekutin.[24] Jsou vyjádřeny především v apikálních membránách epiteliální buňky v respiračních, pankreatických, gastrointestinálních a reprodukčních tkáních.[21][24] Abnormálně zvýšená funkce CFTR má za následek nadměrné vylučování tekutin. Vysoce afinitní inhibitory CFTR, jako je CFTRobyv-172 a GlyH-101 se ukázaly jako účinné při léčbě sekrečních průjmů.[25][26] Teoreticky mohou být blokátory kanálů CFTR užitečné také jako mužská antikoncepce. Kanály CFTR zprostředkovávají vstup aniontů hydrogenuhličitanu, který je nezbytný pro spermie kapacitní.[27]
Je známo, že různé typy látek blokují chloridové iontové kanály CFTR. Mezi nejznámější a nejvíce studované látky patří sulfonylmočoviny, arylaminobenzenoáty a disulfonové stilbeny.[28][29][30] Tyto blokátory jsou závislé na straně, protože vstupují do pórů výhradně z cytoplazmatické strany, jsou závislé na napětí, protože hyperpolarizované membránové potenciály upřednostňují záporně nabitý vstup látky do pórů z cytoplazmatické strany a na koncentraci chloridových iontů závisí jako elektrostaticky vysoké ionty chloridu odpuzujte záporně nabité blokátory zpět do cytoplazmy.[31]
Typy
Existuje několik různých hlavních tříd blokátorů kanálů, včetně:
- Vápník (Ca2+) blokátory kanálů
- Chlorid (Cl−) blokátory kanálů
- Draslík (K.+) blokátory kanálů
- Sodík (Na+) blokátory kanálů
Následující typy, na které působí ligandem řízené iontové kanály (LGIC) prostřednictvím vazby na jejich póry také existují:
- 5-HT3 antagonisté receptoru
- GABAA antagonisté receptoru
- Antagonisté receptoru nACh
- Antagonisté NMDA receptoru
Je také známo, že působí blokátory kanálů AMPA receptory, Glycinové receptory, Kainátové receptory, P2X receptory a Zinek (Zn2+) -aktivované kanály. Typ inhibice zprostředkovaný blokátory kanálů lze označit jako nesoutěžní nebo nekonkurenceschopný.
Viz také
Reference
- ^ „Lékařská definice iontového kanálu“. MedicineNet. Citováno 2017-03-20.
- ^ A b C Kocahan S, Doğan Z (únor 2017). „Mechanismy patogeneze a prevence Alzheimerovy choroby: mozek, nervová patologie, receptory N-methyl-D-aspartátu, proteiny Tau a další rizikové faktory“. Klinická psychofarmakologie a neurovědy. 15 (1): 1–8. doi:10.9758 / cpn.2017.15.1.1. PMC 5290713. PMID 28138104.
- ^ A b C d E Lipton SA (leden 2004). „Selhání a úspěchy antagonistů NMDA receptorů: molekulární základ pro použití blokátorů s otevřeným kanálem, jako je memantin, při léčbě akutních a chronických neurologických urážek“. NeuroRx. Neuroprotekce. 1 (1): 101–10. doi:10.1602 / neurorx.1.1.101. PMC 534915. PMID 15717010.
- ^ Ahern CA, Payandeh J, Bosmans F, Chanda B (leden 2016). „Stopařův průvodce galaxií sodíkovým kanálem s napětím“. The Journal of General Physiology. 147 (1): 1–24. doi:10.1085 / jgp.201511492. PMC 4692491. PMID 26712848.
- ^ Moore JW, poslanec Blaustein, Anderson NC, Narahashi T (květen 1967). „Základ selektivity tetrodotoxinu při blokování chobotnicových axonů“. The Journal of General Physiology. 50 (5): 1401–11. doi:10,1085 / jgp.50.5.1401. PMC 2225715. PMID 6033592.
- ^ Stevens M, Peigneur S, Tytgat J (2011-11-09). „Neurotoxiny a jejich vazebné oblasti na napěťově řízených sodíkových kanálech“. Hranice ve farmakologii. 2: 71. doi:10.3389 / fphar.2011.00071. PMC 3210964. PMID 22084632.
- ^ Miller C (prosinec 1988). "Soutěž o blok Ca2 (+) aktivovaného kanálu K + charybdotoxinem a tetraethylamoniem". Neuron. 1 (10): 1003–6. doi:10.1016/0896-6273(88)90157-2. PMID 2483092.
- ^ Aiyar J, Withka JM, Rizzi JP, Singleton DH, Andrews GC, Lin W, Boyd J, Hanson DC, Simon M, Dethlefs B (listopad 1995). „Topologie oblasti pórů kanálu K + odhalená strukturami toxinů štíra odvozenými z NMR“. Neuron. 15 (5): 1169–81. doi:10.1016/0896-6273(95)90104-3. PMID 7576659.
- ^ A b Findeisen F, Campiglio M, Jo H, Abderemane-Ali F, Rumpf CH, Pope L, Rossen ND, Flucher BE, DeGrado WF, Minor DL (březen 2017). „Sešívané napěťově řízené vápníkové kanály (CaV) Peptidy α-interakční domény (AID) fungují jako selektivní inhibitory CaV funkce interakce protein-protein“. ACS Chemical Neuroscience. 8: 1313–1326. doi:10.1021 / acschemneuro.6b00454. PMC 5481814. PMID 28278376.
- ^ Phoenix DA, Harris F (01.01.2002). „Hydrofobní moment a jeho použití při klasifikaci amfifilních struktur (přehled)“. Molekulární membránová biologie. 19 (1): 1–10. doi:10.1080/09687680110103631. PMID 11989818.
- ^ A b Jackson MB (únor 2010). „Otevřít blok kanálu a dále“. The Journal of Physiology. 588 (Pt 4): 553–4. doi:10.1113 / jphysiol.2009.183210. PMC 2828128. PMID 20173077.
- ^ Ahern CA, Payandeh J, Bosmans F, Chanda B (leden 2016). „Stopařův průvodce galaxií sodíkovým kanálem s napětím“. The Journal of General Physiology. 147 (1): 1–24. doi:10.1085 / jgp.201511492. PMC 4692491. PMID 26712848.
- ^ A b C Butterworth JF, Strichartz GR (duben 1990). "Molekulární mechanismy lokální anestézie: přehled". Anesteziologie. 72 (4): 711–34. doi:10.1097/00000542-199004000-00022. PMID 2157353.
- ^ Evans MH (září 1969). "Mechanismus otravy saxitoxinem a tetrodotoxinem". Britský lékařský bulletin. 25 (3): 263–7. doi:10.1093 / oxfordjournals.bmb.a070715. PMID 5812102.
- ^ Mert T, Gunes Y, Guven M, Gunay I, Ozcengiz D (březen 2002). "Porovnání bloků nervového vedení opioidem a lokálním anestetikem". European Journal of Pharmacology. 439 (1–3): 77–81. doi:10.1016 / S0014-2999 (02) 01368-7. PMID 11937095.
- ^ Mitolo-Chieppa D, Carratu MR (květen 1983). "Anestetika: elektrofyziologické základy jejich účinku blokujícího vedení". Komunikace farmakologického výzkumu. 15 (5): 439–50. doi:10.1016 / s0031-6989 (83) 80064-2. PMID 6351107.
- ^ A b C Chen HS, Pellegrini JW, Aggarwal SK, Lei SZ, Warach S, Jensen FE, Lipton SA (listopad 1992). „Blok otevřených kanálů odpovědí N-methyl-D-aspartátu (NMDA) memantinem: terapeutická výhoda proti neurotoxicitě zprostředkované NMDA receptory“. The Journal of Neuroscience. 12 (11): 4427–36. PMID 1432103.
- ^ A b C Müller WE, Mutschler E, Riederer P (červenec 1995). „Nekonkurenční antagonisté NMDA receptorů s rychlou kinetikou blokování otevřeného kanálu a silnou závislostí na napětí jako potenciální terapeutické látky pro Alzheimerovu demenci“. Farmakopsychiatrie. 28 (4): 113–24. doi:10.1055 / s-2007-979603. PMID 7491365.
- ^ Povysheva NV, Johnson JW (prosinec 2016). „Účinky memantinu na rovnováhu excitace a inhibice v prefrontální kůře“. Neurobiologie nemocí. 96: 75–83. doi:10.1016 / j.nbd.2016.08.006. PMC 5102806. PMID 27546057.
- ^ Dominguez, Evangelyn; Chin, Ting-Yu; Chen, Chih-Ping; Wu, Tzong-Yuan (01.12.2011). „Léčba středně těžké až těžké Alzheimerovy choroby: zaměření na memantin“. Taiwanský časopis porodnictví a gynekologie. 50 (4): 415–423. doi:10.1016 / j.tjog.2011.10.004. ISSN 1028-4559. PMID 22212311.
- ^ A b C Lubamba B, Dhooghe B, Noel S, Leal T (říjen 2012). „Cystická fibróza: pohled na patofyziologii a farmakoterapii CFTR“. Klinická biochemie. 45 (15): 1132–44. doi:10.1016 / j.clinbiochem.2012.05.034. PMID 22698459.
- ^ A b Ramos E, O'leary ME (říjen 2004). „Státně závislé zachycování flekainidu v srdečním sodíkovém kanálu“. The Journal of Physiology. 560 (Pt 1): 37–49. doi:10.1113 / jphysiol.2004.065003. PMC 1665201. PMID 15272045.
- ^ Lipton SA (květen 2007). „Patologicky aktivovaná terapeutika pro neuroprotekci: mechanismus blokování NMDA receptorů memantinem a S-nitrosylací“. Aktuální drogové cíle. 8 (5): 621–32. doi:10.2174/138945007780618472. PMID 17504105.
- ^ A b Frizzell RA, Hanrahan JW (červen 2012). "Fyziologie epiteliálního chloridu a sekrece tekutin". Perspektivy Cold Spring Harbor v medicíně. 2 (6): a009563. doi:10.1101 / cshperspect.a009563. PMC 3367533. PMID 22675668.
- ^ Muanprasat C, Sonawane ND, Salinas D, Taddei A, Galietta LJ, Verkman AS (srpen 2004). „Objev inhibitorů CFTR glycinhydrazidových pórů uzavírajících: mechanismus, analýza struktury a aktivity a účinnost in vivo“. The Journal of General Physiology. 124 (2): 125–37. doi:10.1085 / jgp.200409059. PMC 2229623. PMID 15277574.
- ^ Ma T, Thiagarajah JR, Yang H, Sonawane ND, Folli C, Galietta LJ, Verkman AS (prosinec 2002). „Inhibitor thiazolidinonu CFTR identifikovaný vysoce výkonným screeningem blokuje sekreci střevní tekutiny vyvolanou toxinem cholery“. The Journal of Clinical Investigation. 110 (11): 1651–8. doi:10.1172 / JCI16112. PMC 151633. PMID 12464670.
- ^ Chen H, Ruan YC, Xu WM, Chen J, Chan HC (01.11.2012). „Regulace mužské plodnosti pomocí CFTR a důsledky pro mužskou neplodnost“. Aktualizace lidské reprodukce. 18 (6): 703–13. doi:10.1093 / humupd / dms027. PMID 22709980.
- ^ Schultz BD, DeRoos AD, Venglarik CJ, Singh AK, Frizzell RA, Bridges RJ (srpen 1996). "Blokování glibenklamidu u chloridových kanálů CFTR". Americký žurnál fyziologie. 271 (2 Pt 1): L192-200. doi:10.1152 / ajplung.1996.271.2.L192. PMID 8770056.
- ^ Zhang ZR, Zeltwanger S, McCarty NA (květen 2000). "Přímé srovnání NPPB a DPC jako sondy CFTR exprimované v oocytech Xenopus". The Journal of Membrane Biology. 175 (1): 35–52. doi:10,1007 / s002320001053. PMID 10811966.
- ^ Linsdell P, Hanrahan JW (listopad 1996). „Disulfonický stilbenový blok cystických fibrózových transmembránových regulátorů vodivosti Cl-kanálů exprimovaných v buněčné linii savců a jeho regulace kritickým zbytkem pórů“. The Journal of Physiology. 496 (Pt 3): 687–93. doi:10.1113 / jphysiol.1996.sp021719. PMC 1160856. PMID 8930836.
- ^ Linsdell P (únor 2014). „Transmembránový regulátor vodivosti cystické fibrózy blokátory chloridových kanálů: Farmakologický, biofyzikální a fyziologický význam“. World Journal of Biological Chemistry. 5 (1): 26–39. doi:10,4331 / wjbc.v5.i1.26. PMC 3942540. PMID 24600512.