Kalciový kanál typu L. - L-type calcium channel
| Vápníkový kanál, závislý na napětí | |
|---|---|
 Krystalografická struktura | |
| Identifikátory | |
| Symbol | Vápníkový kanál, závislý na napětí |


The Kalciový kanál typu L. (také známý jako dihydropyridinový kanál, nebo Kanál DHP ) je součástí rodiny aktivované vysokým napětím napěťově závislý vápníkový kanál.[2]„L“ znamená dlouhodobý odkaz na délku aktivace. Tento kanál má čtyři podjednotky (Cav1.1, Cav1.2, 1.3, 1.4 ).
Vápenaté kanály typu L jsou odpovědné za excitaci -kontrakce spojení kosterní, hladký, srdeční sval, a pro aldosteron sekrece v endokrinní buňky z kůra nadledvin.[1] Vyskytují se také v neuronech a pomocí vápníkových kanálů typu L v endokrinních buňkách regulují neurohormony a neurotransmitery. Bylo také zjištěno, že hrají roli v genové expresi, stabilitě mRNA, přežití neuronů, ischemicky indukovaném axonálním poškození, synaptické účinnosti a aktivaci i deaktivaci dalších iontových kanálů.[3]
V srdečních myocytech prochází vápníkový kanál typu L dovnitř Ca2+ proud a spouští uvolňování vápníku ze sarkoplazmatického retikula aktivací ryanodinový receptor 2 (RyR2) (uvolňování vápníku indukované vápníkem).[4] Fosforylace těchto kanálů zvyšuje jejich propustnost pro vápník a zvyšuje kontraktilitu jejich příslušných srdečních myocytů.
Typ L. blokátor kalciového kanálu léky se používají jako srdeční antiarytmika nebo antihypertenziva, v závislosti na tom, zda mají léky vyšší afinitu k srdce (dále jen fenylalkylaminy, jako verapamil ) nebo pro krevní cévy ( dihydropyridiny, jako nifedipin ).[5]
V kosterním svalu je velmi vysoká koncentrace vápníkových kanálů typu L, která se nachází v T-tubuly. Výsledkem depolarizace svalů jsou velké hradlové proudy, ale neobvykle nízký tok vápníku, což se nyní vysvětluje velmi pomalou aktivací iontových proudů. Z tohoto důvodu malý nebo žádný Ca2+ prochází přes membránu T-tubulu během jediného akčního potenciálu.
Dějiny
V roce 1953 objevili Paul Fatt a Bernard Katz napěťově řízené vápníkové kanály ve svalu korýšů. Kanály vykazovaly různá aktivační napětí a vlastnosti vedení vápníku a byly tak rozděleny na vysokonapěťové aktivační kanály (HVA) a nízkonapěťové aktivační kanály (LVA). Po dalším experimentování bylo zjištěno, že by se kanály HVA otevřely 1,4-dihydropyridin (DHP ).[6] Pomocí DHP zjistili, že kanály HVA byly specifické pro určité tkáně a reagovaly odlišně, což vedlo k další kategorizaci kanálů HVA do typu L, Typ P., a N-typ.[3] Vápníkové kanály typu L byly sekvenovány peptidem a bylo zjištěno, že existují 4 druhy vápníkových kanálů typu L: a1S (kosterní sval), α1C (srdeční), α1 D (nachází se v mozku) a α1F (nachází se v sítnici).[6] V roce 2000, poté, co byl proveden další výzkum na α1 podjednotek v napěťově řízených vápníkových kanálech byla použita nová nomenklatura, která se nazývá vápníkové kanály typu L CaV1, přičemž jeho podjednotky se nazývají CaV1.1, CaV1.2, CaV1.3 a CaV1.4.[3] Výzkum podjednotek CaV1 nadále odhaluje více o jejich struktuře, funkci a farmaceutických aplikacích.[7]
Struktura
Vápníkové kanály typu L obsahují 5 různých podjednotek, a1 (170–240 kDa), α2 (150 kDa), δ (17-25 kDa), β (50-78 kDa) a γ (32 kDa) podjednotek.[8] Podjednotky α2, δ a β jsou nekovalentně navázány na podjednotku α1 a modulují přenos iontů a biofyzikální vlastnosti podjednotky α1. Podjednotky α2 a δ jsou v extracelulárním prostoru, zatímco podjednotky β a γ jsou umístěny v cytosolickém prostoru.[8]
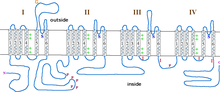
Podjednotka α1 je heterotetramer, který má čtyři transmembránové oblasti, známé jako domény I-IV, které procházejí plazmou šestkrát α-šroubovice, nazývané S0-S6 (S0 a S1 společně procházejí membránou jednou).[3] Podjednotka α1 jako celek obsahuje doménu snímání napětí, vodivé póry a hradlovací zařízení.[9] Jako většina napěťově řízené iontové kanály, α-podjednotka se skládá ze 4 podjednotek. Každá podjednotka je tvořena 6 alfa-helikálními, transmembránovými doménami, které procházejí membránou (očíslované S1-S6). Podjednotky S1-S4 tvoří napěťový senzor, zatímco podjednotky S5-S6 tvoří selektivní filtr.[10] Ke snímání napětí buňky obsahují helixy S1-S3 mnoho negativně nabitých aminokyselin aminokyselin, zatímco helixy S4 obsahují většinou pozitivně nabité aminokyseliny s P-smyčka připojení šroubovice S4 k S5. Po doménách S1-6 existuje šest domén C, které se skládají ze dvou EF-ruční motivy (C1-2 a C3-4) a doména Pre-IQ (C5) a IQ doména (C6). Na N-konci jsou také dva motivy EF ruky. N i C konec jsou v cytosolickém prostoru, přičemž C-konec je mnohem delší než N-konec.[11]
Je známo, že podjednotka β má čtyři izoformy (β1-β4) k regulaci funkcí kanálu a je připojen k α1 prostřednictvím linkeru α1 I a II v cytosolu na β α1-kapsa na vázání (ABP).[7][12] Každá izoforma obsahuje a src homologie 3 domény (SH3) a guanylát-kináza jako domian (GK), které jsou odděleny doménou HOOK a třemi nestrukturovanými oblastmi.[12]
Podjednotky α2 a δ jsou vzájemně spojeny disulfidovými vazbami (někdy známými jako podjednotka α2δ) a interagují s α1.[7] mají čtyři známé izoformy zvané α2δ-1 až α2δ-2 a obsahují doménu von Willebrand A (VWA) a Mezipaměť domény. Oblast α2 je v extracelulárním prostoru, zatímco oblast δ je v buněčné membráně a bylo vidět, že je ukotvena pomocí glykosylfosfatidylinositol (GPI) kotva.[12]
Γ podjednotka má osm izoforem (γ1-γ8) a je připojena k podjednotce α1 a byla nalezena pouze ve svalových buňkách v kanálech CaV1.1 a CaV1.2.[12] O γ podjednotce není známo mnoho, ale souvisí to s interakcemi v hydrofobních silách.[3]
Mechanismus
K otevření pórů v kalciových kanálech typu L dochází v podjednotce α1. Když se membrána depolarizuje, šroubovice S4 se pohybuje spojkami S4 a S5 na cytoplazmatické konce šroubovic S5 a S6. Tím se otevře aktivační brána který je tvořen vnitřní stranou spirál S6 v podjednotce α1.[11]
Nejvýznamnějším způsobem autoinhibice vápníkových kanálů typu L je komplex Ca + 2 / Cam.[11] Jak se póry otevírají a způsobují příliv vápníku, váže se na něj vápník klimodulin a pak interaguje se smyčkou, která spojuje sousední EF-ruční motivy a způsobí konformační změnu motivu EF ruky, takže interaguje s póry a způsobí rychlou inhibici v kanálu.[6] Stále se diskutuje o tom, kde a jak póry a EF ruka interagují. Hydrofobní kapsy v komplexu Ca + 2 / Cam se budou také vázat na tři části IQ doména známé jako „aromatické kotvy“.[11] Komplex CA + 2 / Cam má vysokou afinitu k vápníkovým kanálům typu L, což mu umožňuje blokovat, i když je v buňce přítomno nízké množství vápníku. Pór se nakonec uzavře, když se buňka repolarizuje a způsobí konformační změnu v kanálu, aby se dostal do uzavřené konformace.
Inhibice a modulace
Jednou z nejuznávanějších charakteristik vápníkového kanálu typu L je jeho jedinečná citlivost na 1,4-dihydropyridiny (DHP).[3] Na rozdíl od jiných napěťově řízených vápníkových kanálů jsou vápníkové kanály typu L odolné vůči ⍵-CT X (GVIA) a itory-AG A (IVA) inhibiční léky.[3]
Dobře pozorovaná forma modulace je způsobena alternativní sestřih. Běžnou formou modulace z alternativního sestřihu je C-terminální modulátor (CTM). Má pozitivně nabitou a-šroubovici na C-konci zvanou DCRD a záporně nabitou šroubovici hned za IQ motivem (místo interakce CaM) zvaným PCRD. Tyto dvě šroubovice mohou tvořit strukturu, která se váže konkurenčně Vačka snížit pravděpodobnost otevřeného stavu a snížit inhibici závislou na vápníku (CDI).[7]
Alternativní sestřih je také vidět na β podjednotkách, aby se vytvořily různé izoformy dát kanálům různé vlastnosti kvůli palmitoylace[6] a Úpravy RNA.[7] Jiné formy modulace na p podjednotce zahrnují zvýšení nebo snížení exprese podjednotky. To je způsobeno skutečností, že β podjednotky zvyšují pravděpodobnost otevření kanálu, aktivitu v plazmatické membráně a antagonizují ubikvitinace kanálu.[6]
Vápníkové kanály typu L jsou také modulovány Receptory spojené s G proteinem a adrenergní nervový systém.[6] Protein kináza A (PKA) aktivovaný kaskádou receptorů spřažených s G proteinem může fosforylovat vápníkové kanály typu L poté, co kanály vytvoří signální komplex s A-kinázové kotvící proteiny (AKAP) , zvýšit proud vápníku kanálem, zvýšit pravděpodobnost otevřeného stavu a zrychlené období zotavení. Aktivováno Fosfolipáza C (PLC) z receptorů spřažených s G proteinem může rozložit polyfosfoinositidy a snížit kanály vápníkového proudu o 20% - 30%.[7]
Bylo zjištěno, že adrenergní nervový systém moduluje vápníkové kanály typu L štěpením C-koncového fragmentu, když je stimulován β-adrenergní receptor ke zvýšení aktivace kanálů.[6]
Geny
Viz také
Reference
- ^ A b Felizola SJ, Maekawa T, Nakamura Y, Satoh F, Ono Y, Kikuchi K a kol. (Říjen 2014). "Napěťově řízené vápníkové kanály v lidském nadledvinu a primárním aldosteronismu". The Journal of Steroid Biochemistry and Molecular Biology. 144 Pt B (část B): 410–6. doi:10.1016 / j.jsbmb.2014.08.012. PMID 25151951. S2CID 23622821.
- ^ Rossier MF (2016). „Vápníkový kanál typu T: Privilegovaná brána pro vstup vápníku a kontrolu nad steroidogenezí nadledvin“. Hranice v endokrinologii. 7: 43. doi:10.3389 / fendo.2016.00043. PMC 4873500. PMID 27242667.
- ^ A b C d E F G Lipscombe D, Helton TD, Xu W (listopad 2004). "Vápníkové kanály typu L: nízká hladina". Journal of Neurophysiology. 92 (5): 2633–41. doi:10.1152 / jn.00486.2004. PMID 15486420.
- ^ Yamakage M, Namiki A (únor 2002). „Vápníkové kanály - základní aspekty jejich struktury, funkce a kódování genů; anestetické působení na kanály - přehled“. Canadian Journal of Anesthesia. 49 (2): 151–64. doi:10.1007 / BF03020488. PMID 11823393.
- ^ Hughes A (2017). "Blokátory kalciového kanálu". In Bakris G, Sorrentino M (eds.). Hypertenze: společník Braunwaldovy choroby srdce (Třetí vydání.). Philadelphia, PA: Elsevier Health Sciences. str. 242–253. ISBN 9780323508766. OCLC 967938982.
- ^ A b C d E F G Dolphin AC (říjen 2018). „Napěťově řízené vápníkové kanály: jejich objev, funkce a význam jako lékové cíle“. Pokroky v mozku a neurovědě. 2: 2398212818794805. doi:10.1177/2398212818794805. PMC 6179141. PMID 30320224.
- ^ A b C d E F Striessnig J, Pinggera A, Kaur G, Bock G, Tuluc P (březen 2014). „2+ kanálů v srdci a mozku“. Wiley Interdisciplinary Reviews: Membrane Transport and Signaling. 3 (2): 15–38. doi:10,1002 / hm. 102. PMC 3968275. PMID 24683526.
- ^ A b Bodi I, Mikala G, Koch SE, Akhter SA, Schwartz A (prosinec 2005). „Kalciový kanál typu L v srdci: rytmus pokračuje“. The Journal of Clinical Investigation. 115 (12): 3306–17. doi:10,1172 / JCI27167. PMC 1297268. PMID 16322774.
- ^ "Napěťově řízené vápníkové kanály | Úvod | BPS / IUPHAR Průvodce po farmakologii". www.guidetopharmacology.org. Citováno 2019-11-28.
- ^ Catterall WA, Perez-Reyes E, Snutch TP, Striessnig J (prosinec 2005). „International Union of Pharmacology. XLVIII. Nomenklatura a strukturně-funkční vztahy napěťově řízených vápníkových kanálů“. Farmakologické recenze. 57 (4): 411–25. doi:10.1124 / pr.57.4.5. PMID 16382099. S2CID 10386627.
- ^ A b C d Wahl-Schott C, Baumann L, Cuny H, Eckert C, Griessmeier K, Biel M (říjen 2006). "Vypnutí deaktivace závislé na vápníku v vápníkových kanálech typu L pomocí autoinhibiční domény". Sborník Národní akademie věd Spojených států amerických. 103 (42): 15657–62. Bibcode:2006PNAS..10315657W. doi:10.1073 / pnas.0604621103. PMC 1622877. PMID 17028172.
- ^ A b C d Shaw RM, Colecraft HM (květen 2013). "Cílení vápníkových kanálů typu L a místní signalizace v srdečních myocytech". Kardiovaskulární výzkum. 98 (2): 177–86. doi:10.1093 / cvr / cvt021. PMC 3633156. PMID 23417040.
Další čtení
- Takahashi K, Hayashi S, Miyajima M, Omori M, Wang J, Kaihara K a kol. (Květen 2019). „Kalciový kanál typu L moduluje mechanosenzitivitu buněčné linie kardiomyocytů H9c2“. Buněčný vápník. 79: 68–74. doi:10.1016 / j.ceca.2019.02.008. PMID 30836292.
| Tento článek o gen na lidský chromozom 11 je pahýl. Wikipedii můžete pomoci pomocí rozšiřovat to. |
Tento článek včlení text z United States National Library of Medicine, který je v veřejná doména.
externí odkazy
- „Napěťově řízené vápníkové kanály“. Databáze IUPHAR receptorů a iontových kanálů. Mezinárodní unie základní a klinické farmakologie.
- Typ L + vápník + kanál v americké národní lékařské knihovně Lékařské předměty (Pletivo)