Leucin - Leucine
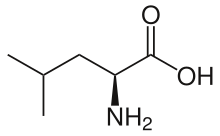 L-Leucin | |
 | |
| Jména | |
|---|---|
| Název IUPAC Leucin | |
| Ostatní jména Kyselina 2-amino-4-methylpentanová | |
| Identifikátory | |
3D model (JSmol ) | |
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| DrugBank | |
| Informační karta ECHA | 100.000.475 |
| KEGG | |
PubChem CID | |
| UNII | |
Řídicí panel CompTox (EPA) | |
| |
| |
| Vlastnosti | |
| C6H13NÓ2 | |
| Molární hmotnost | 131.175 g · mol−1 |
| Kyselost (strK.A) | 2,36 (karboxyl), 9,60 (amino)[1] |
| -84.9·10−6 cm3/ mol | |
| Stránka s doplňkovými údaji | |
| Index lomu (n), Dielektrická konstanta (εr), atd. | |
Termodynamické data | Fázové chování pevná látka - kapalina - plyn |
| UV, IR, NMR, SLEČNA | |
Pokud není uvedeno jinak, jsou uvedeny údaje o materiálech v nich standardní stav (při 25 ° C [77 ° F], 100 kPa). | |
| Reference Infoboxu | |
Leucin (symbol Leu nebo L)[2] je esenciální aminokyselina který se používá v biosyntéza z bílkoviny. Leucin je α-aminokyselina, což znamená, že obsahuje α-aminoskupina (který je v protonované -NH3+ za biologických podmínek), α-skupina karboxylové kyseliny (který je v deprotonovaném −COO− forma za biologických podmínek) a postranní řetězec isobutylová skupina, což je a nepolární alifatický aminokyselina. to je nezbytný u lidí, což znamená, že si ho tělo nedokáže syntetizovat: musí být získáno ze stravy. Lidské zdroje výživy jsou potraviny, které obsahují bílkoviny, jako jsou maso, mléčné výrobky, sójové výrobky a fazole a jiné luštěniny. to je kódovaný podle kodony UUA, UUG, CUU, CUC, CUA a CUG.
Jako valin a isoleucin, leucin je a aminokyselina s rozvětveným řetězcem. Primární konečné metabolické produkty metabolismu leucinů jsou acetyl-CoA a acetoacetát; je tedy výhradně jedním ze dvou ketogenní aminokyseliny, s lysin být tím druhým.[3] Je to nejdůležitější ketogenní aminokyselina u lidí.[4]
Leucin a kyselina p-hydroxy-p-methylmáselná, menší leucin metabolit, exponát farmakologická aktivita u lidí a bylo prokázáno, že propagují biosyntéza bílkovin přes fosforylace z mechanický cíl rapamycinu (mTOR).[5][6]
Dietní leucin
Jako potravinářská přídatná látka, L-leucin má Číslo E. E641 a je klasifikován jako a zvýrazňovač chuti.[7]
Požadavky
Rada pro výživu a výživu (FNB) Amerického lékařského institutu stanovila doporučené dietní dávky (RDA) pro esenciální aminokyseliny v roce 2002. Pro leucin, pro dospělé ve věku 19 let a starší, 42 mg / kg tělesné hmotnosti / den.[8]
Zdroje
| Jídlo | g / 100 g |
|---|---|
| Syrovátková bílkovina koncentrát, suchý prášek | 10.0-12.0 |
| Sójový protein koncentrát, suchý prášek | 7.5-8.5 |
| Hráškový protein koncentrát, suchý prášek | 6.6 |
| Sójové boby, zralá semena, pražená, solená | 2.87 |
| Konopí loupané semeno | 2.16 |
| Hovězí, kulatý, horní kulatý, syrový | 1.76 |
| Arašídy | 1.67 |
| Ryba, losos, růžový, syrový | 1.62 |
| Pšeničné klíčky | 1.57 |
| Mandle | 1.49 |
| Kuře, brojlery nebo fritézy, stehno, syrové | 1.48 |
| Slepičí vejce, žloutek, syrový | 1.40 |
| Oves | 1.28 |
| Edamame (sójové boby, zelené, syrové) | 0.93 |
| Fazole, pinto, vařené | 0.78 |
| Čočka, vařené | 0.65 |
| Cizrna, vařené | 0.63 |
| Kukuřice, žlutá | 0.35 |
| Kravské mléko, celé, 3,25% mléčného tuku | 0.27 |
| Rýže, hnědá, středně zrnitá, vařená | 0.19 |
| Mléko, člověk, zralý, tekutý | 0.10 |
Zdravé efekty
Jako doplněk stravy Bylo zjištěno, že leucin zpomaluje degradaci svalové tkáně zvýšením syntézy svalových proteinů u starších krys.[10] Výsledky srovnávacích studií jsou však konfliktní. Dlouhodobá suplementace leucinem nezvyšuje svalovou hmotu ani sílu u zdravých starších mužů.[11] Je zapotřebí více studií, nejlépe studií založených na objektivním náhodném vzorku společnosti. Faktory, jako je výběr životního stylu, věk, pohlaví, strava, cvičení atd., Musí být zohledněny v analýzách, aby se izolovaly účinky doplňkového leucinu jako samostatného přípravku nebo pokud se užívají s jinými aminokyseliny s rozvětveným řetězcem (BCAA). Do té doby nelze doplňkový leucin doplňovat jako hlavní důvod svalového růstu nebo optimální údržby pro celou populaci.
L-leucin i D-leucin chrání myši před záchvaty.[12] D-leucin také ukončí záchvaty u myší po nástupu záchvatové aktivity, přinejmenším stejně účinně jako diazepam a bez sedativních účinků.[12] Snížený dietní příjem L-leucinu podporuje u myší adipozitu.[13] Vysoké hladiny leucinu v krvi jsou spojeny s inzulínovou rezistencí u lidí, myší a hlodavců.[14] Může to být způsobeno stimulací leucinu mTOR signalizace.[15] Dietní omezení leucinů a dalších BCAA může zvrátit obezitu vyvolanou stravou u myší divokého typu zvýšením energetického výdeje a může omezit nárůst tukové hmoty u hyperfagických potkanů.[16][17]
Bezpečnost
Toxicita pro leucin, jak je vidět u dekompenzované onemocnění moči javorovým sirupem, způsobuje delirium a neurologické kompromisy a může být život ohrožující.[Citace je zapotřebí ]
Vysoký příjem leucinů může způsobit nebo zhoršit příznaky pelagra u lidí s nízkou niacin status, protože narušuje převod L-tryptofan na niacin.[18]
Leucin v dávce vyšší než 500 mg / kg / den byl pozorován u hyperamonémie.[19] Jako takový lze neoficiálně navrhnout tolerovatelnou horní úroveň příjmu (UL) pro leucin u zdravých dospělých mužů v dávce 500 mg / kg / den nebo 35 g / den za akutních dietních podmínek.[19][20]
Farmakologie
Farmakodynamika
Leucin je a dietní aminokyselina se schopností přímo stimulovat myofibrilární sval proteosyntéza.[21] Tento účinek leucinu vyplývá z jeho role jako aktivátoru mechanický cíl rapamycinu (mTOR),[6] A serin-threoninová protein kináza který reguluje biosyntéza bílkovin a růst buněk. Aktivace mTOR leucinem je zprostředkována prostřednictvím Hadrové GTPasy,[22][23][24] leucin vazba na leucyl-tRNA syntetáza,[22][23] vazba leucinu na sestrin 2,[25][26][27] a případně další mechanismy.
Metabolismus u lidí
Metabolismus leucinů u lidí |

{kind=link}
Metabolismus leucinů se vyskytuje u mnoha tkáně v lidském těle; nicméně, většina dietní leucin je metabolizována v játra, tuková tkáň, a svalová tkáň.[nutná lékařská citace ] Tuk a svalová tkáň používají při tvorbě leucin steroly a další sloučeniny.[nutná lékařská citace ] Kombinované užívání leucinu v těchto dvou tkáních je sedmkrát vyšší než v játrech.[33]
U zdravých jedinců přibližně 60% stravy L-leucin se metabolizuje po několika hodinách, zhruba 5% (2–10% rozsah) dietní L-leucin se převádí na kyselina p-hydroxy-p-methylmáselná (HMB).[31][34][32] Kolem 40% stravy L-leucin je převeden na acetyl-CoA, který se následně používá při syntéze dalších sloučenin.[32]
Drtivá většina z L-leucin metabolismus je zpočátku katalyzován aminotransferáza s rozvětveným řetězcem produkující enzym a-ketoisocaproate (α-KIC).[31][32] α-KIC je většinou metabolizován mitochondriální enzym rozvětvený řetězec α-ketokyselina dehydrogenáza, který jej převede na isovaleryl-CoA.[31][32] Isovaleryl-CoA je následně metabolizován isovaleryl-CoA dehydrogenáza a převedeny na MC-CoA, který se používá při syntéze acetyl-CoA a dalších sloučenin.[32] V době nedostatek biotinu, HMB lze syntetizovat z MC-CoA přes enoyl-CoA hydratáza a neznámý thioesteráza enzym,[28][29][35] které konvertují MC-CoA do HMB-CoA a HMB-CoA do HMB.[29] Relativně malé množství α-KIC se metabolizuje v játra podle cytosolický enzym 4-hydroxyfenylpyruvát dioxygenáza (KIC dioxygenáza), která převádí α-KIC na HMB.[31][32][36] U zdravých jedinců tato vedlejší cesta - která zahrnuje přeměnu L-leucin na α-KIC a pak HMB - je převládající cestou syntézy HMB.[31][32]
Malý zlomek L-leucin metabolismus - méně než 5% ve všech tkáních kromě testy kde představuje přibližně 33% - je zpočátku katalyzován leucinaminomutáza, produkující β-leucin, který je následně metabolizován na β-ketoisocaproate (β-KIC), p-ketoizokaproyl-CoA a poté acetyl-CoA řadou necharakterizovaných enzymů.[32][37]
Metabolismus HMB je katalyzován necharakterizovaným enzymem, který jej převádí na p-hydroxy p-methylbutyryl-CoA (HMB-CoA).[28][32] HMB-CoA je metabolizován buď enoyl-CoA hydratáza nebo jiný necharakterizovaný enzym produkující p-methylkrotonyl-CoA (MC-CoA) nebo hydroxymethylglutaryl-CoA (HMG-CoA).[31][32] MC-CoA se pak převádí enzymem methylkrotonyl-CoA karboxyláza na methylglutaconyl-CoA (MG-CoA), který je následně převeden na HMG-CoA podle methylglutakonyl-CoA hydratáza.[31][32][37] HMG-CoA se poté štěpí na acetyl-CoA a acetoacetát podle HMG-CoA lyáza nebo se používá při výrobě cholesterolu prostřednictvím mevalonátová cesta.[31][32]
Syntéza v nehumánních organismech
Leucin je esenciální aminokyselina ve stravě zvířat, protože k jeho syntéze chybí úplná enzymatická cesta de novo z potenciálních prekurzorových sloučenin. V důsledku toho je musí přijímat, obvykle jako součást bílkovin. Rostliny a mikroorganismy syntetizují leucin z kyselina pyrohroznová s řadou enzymů:[38]
- Acetolaktát syntáza
- Isomero kyseliny acetohydroxylovéreduktáza
- Dihydroxykyselina dehydratáza
- a-isopropylmalát syntáza
- a-isopropylmalát izomeráza
- Leucin aminotransferáza
Syntéza malé hydrofobní aminokyseliny valin zahrnuje také počáteční část této cesty.
Chemie

Leucin je aminokyselina s rozvětveným řetězcem (BCAA), protože obsahuje alifatický postranní řetězec, který není lineární.
Racemický leucin byl podroben cirkulárně polarizovaný synchrotronové záření lépe porozumět původu biomolekulární asymetrie. Bylo vyvoláno enantiomerní zvýšení o 2,6%, což naznačuje možný fotochemický původ biomolekul. homochirality.[39]
Viz také
- Leuciny, izomery a deriváty leucinu
- Leucinový zip, společný motiv v proteinech transkripčního faktoru
Poznámky
- ^ Tato reakce je katalyzována neznámou thioesteráza enzym.[28][29]
Reference
- ^ Dawson, R.M.C. a kol., Data pro biochemický výzkum, Oxford, Clarendon Press, 1959.
- ^ „Nomenklatura a symbolika pro aminokyseliny a peptidy“. Smíšená komise IUPAC-IUB pro biochemickou nomenklaturu. 1983. Archivovány od originál dne 9. října 2008. Citováno 5. března 2018.
- ^ Ferrier, Denise R. (24. května 2013). Biochemie. Lippincott Williams & Wilkins. ISBN 9781451175622.
- ^ Cynober, Luc A. (13. listopadu 2003). Metabolické a terapeutické aspekty aminokyselin v klinické výživě, druhé vydání. CRC Press. p. 101. ISBN 9780203010266.
- ^ Silva VR, Belozo FL, Micheletti TO, Conrado M, Stout JR, Pimentel GD, Gonzalez AM (září 2017). „Doplnění volné kyseliny β-hydroxy-β-methylbutyrát může zlepšit regeneraci a přizpůsobení svalů po tréninku na odpor: systematický přehled.“ Výzkum výživy. 45: 1–9. doi:10.1016 / j.nutres.2017.07.008. hdl:11449/170023. PMID 29037326.
Mechanismy účinku HMB se obecně považují za související s jeho účinkem jak na syntézu svalových bílkovin, tak na rozpad svalových proteinů (obrázek 1) [2, 3]. Zdá se, že HMB stimuluje syntézu svalových proteinů prostřednictvím upregulace savčího / mechanického cíle rapamycinového komplexu 1 (mTORC1), signální kaskády zapojené do koordinace iniciace translace svalové proteinové syntézy [2, 4]. HMB může mít navíc antagonistické účinky na dráhu ubikvitin-proteazom, systém, který degraduje intracelulární proteiny [5, 6]. Důkazy také naznačují, že HMB podporuje myogenní proliferaci, diferenciaci a fúzi buněk [7]. ... Ukázalo se, že exogenní podávání HMB-FA zvyšuje intramuskulární anabolickou signalizaci, stimuluje syntézu svalových bílkovin a oslabuje rozpad svalových bílkovin u lidí [2].
- ^ A b Wilkinson DJ, Hossain T, Hill DS, Phillips BE, Crossland H, Williams J, Loughna P, Churchward-Venne TA, Breen L, Phillips SM, Etheridge T, Rathmacher JA, Smith K, Szewczyk NJ, Atherton PJ (červen 2013) . „Účinky leucinu a jeho metabolitu β-hydroxy-β-methylbutyrátu na metabolismus bílkovin lidského kosterního svalstva“. The Journal of Physiology. 591 (11): 2911–2923. doi:10.1113 / jphysiol.2013.253203. PMC 3690694. PMID 23551944.
Stimulace MPS prostřednictvím signalizace mTORc1 po expozici HMB je v souladu s preklinickými studiemi (Eley et al. 2008). ... Kromě toho byla zřejmá divergence v amplitudě fosforylace pro 4EBP1 (u Thr37 / 46 a Ser65 / Thr70) a p70S6K (Thr389) v reakci na Leu i HMB, přičemž druhá vykazovala výraznější a trvalejší fosforylaci. ... Nicméně vzhledem k tomu, že celková odpověď MPS byla podobná, tento rozdíl v buněčné signalizaci se neprojevil ve statisticky rozlišitelných anabolických účincích v našem primárním měřítku výsledku MPS. ... Je zajímavé, že i když orálně dodávaný HMB neprodukoval žádné zvýšení plazmatického inzulínu, způsobil pokles MPB (-57%). Za normálních okolností se postprandiální poklesy MPB (o ~ 50%) připisují účinkům inzulínu šetřícího dusík, protože podávání inzulinu v postabsorpčních koncentracích (5 μU ml−1) při nepřetržitém infuzi AA (18 g h−1) nepotlačil MPB (Greenhaff et al. 2008), a proto jsme se rozhodli neměřit MPB ve skupině Leu kvůli očekávané hyperinzulinémii (obr. 3C). HMB tedy snižuje MPB způsobem podobným, ale nezávislým na inzulínu. Tato zjištění jsou v souladu se zprávami o antikatabolických účincích HMB potlačujících MPB v preklinických modelech prostřednictvím oslabení proteolýzy zprostředkované proteazomálně v reakci na LPS (Eley et al. 2008).
- ^ Winter, Ruth (2009). Slovník spotřebitelů potravinářských přídatných látek (7. vydání). New York: Three Rivers Press. ISBN 978-0307408921.
- ^ Lékařský ústav (2002). „Proteiny a aminokyseliny“. Referenční dietní příjem energie, sacharidů, vlákniny, tuků, mastných kyselin, cholesterolu, bílkovin a aminokyselin. Washington, DC: Národní akademie Press. 589–768.
- ^ Národní databáze živin pro standardní referenci. Americké ministerstvo zemědělství. Archivovány od originál dne 3. března 2015. Citováno 16. září 2009.
- ^ L. Combaret a kol. Centrum pro výzkum lidské výživy v Clermont-Ferrand (2005). „Dieta doplněná leucinem obnovuje defektní postprandiální inhibici proteolýzy závislé na proteazomu ve věku kosterního svalu krysy“. Journal of Physiology Volume 569, číslo 2, s. 1. 489-499. 569 (Pt 2): 489–99. doi:10.1113 / jphysiol.2005.098004. PMC 1464228. PMID 16195315. Archivovány od originál dne 16. května 2008. Citováno 25. března 2008.
- ^ Verhoeven S, Vanschoonbeek K, Verdijk LB, Koopman R, Wodzig WK, Dendale P, van Loon LJ (květen 2009). „Dlouhodobá suplementace leucinem nezvyšuje svalovou hmotu ani sílu u zdravých starších mužů“. American Journal of Clinical Nutrition. 89 (5): 1468–75. doi:10.3945 / ajcn.2008.26668. PMID 19321567.
- ^ A b Hartman AL, Santos P, O'Riordan KJ, Stafstrom CE, Marie Hardwick J (říjen 2015). „Silné účinky D-leucinu proti záchvatům“. Neurobiologie nemocí. 82: 46–53. doi:10.1016 / j.nbd.2015.05.013. PMC 4640989. PMID 26054437.
- ^ Fontana L, Cummings NE, Arriola Apelo SI, Neuman JC, Kasza I, Schmidt BA, Cava E, Spelta F, Tosti V, Syed FA, Baar EL, Veronese N, Cottrell SE, Fenske RJ, Bertozzi B, Brar HK, Pietka T, Bullock AD, Figenshau RS, Andriole GL, Merrins MJ, Alexander CM, Kimple ME, Lamming DW (červenec 2016). „Snížená spotřeba aminokyselin s rozvětveným řetězcem zlepšuje metabolické zdraví“. Zprávy buněk. 16 (2): 520–530. doi:10.1016 / j.celrep.2016.05.092. PMC 4947548. PMID 27346343.
- ^ Lynch CJ, Adams SH (prosinec 2014). „Aminokyseliny s rozvětveným řetězcem v metabolické signalizaci a inzulínové rezistenci“. Recenze přírody. Endokrinologie. 10 (12): 723–36. doi:10.1038 / nrendo.2014.171. PMC 4424797. PMID 25287287.
- ^ Caron A, Richard D, Laplante M (2015). "Role komplexů mTOR v metabolismu lipidů". Každoroční přehled výživy. 35: 321–48. doi:10.1146 / annurev-nutr-071714-034355. PMID 26185979.
- ^ Cummings NE, Williams EM, Kasza I, Konon EN, Schaid MD, Schmidt BA, Poudel C, Sherman DS, Yu D, Arriola Apelo SI, Cottrell SE, Geiger G, Barnes ME, Wisinski JA, Fenske RJ, Matkowskyj KA, Kimple ME, Alexander CM, Merrins MJ, Lamming DW (prosinec 2017). „Obnova metabolického zdraví snížením spotřeby aminokyselin s rozvětveným řetězcem“. The Journal of Physiology. 596 (4): 623–645. doi:10.1113 / JP275075. PMC 5813603. PMID 29266268.
- ^ White PJ, Lapworth AL, An J, Wang L, McGarrah RW, Stevens RD, Ilkayeva O, George T, Muehlbauer MJ, Bain JR, Trimmer JK, Brosnan MJ, Rolph TP, Newgard CB (červenec 2016). „Omezení aminokyselin s rozvětveným řetězcem u potkanů Zucker-mastných zvyšuje citlivost svalového inzulínu zvýšením účinnosti oxidace mastných kyselin a exportu acyl-glycinu“. Molekulární metabolismus. 5 (7): 538–51. doi:10.1016 / j.molmet.2016.04.006. PMC 4921791. PMID 27408778.
- ^ Badawy AA, Lake SL, Dougherty DM (2014). „Mechanismy pelagragenního účinku leucinu: stimulace oxidace jaterního tryptofanu podáváním aminokyselin s rozvětveným řetězcem zdravým lidským dobrovolníkům a úloha bez plazmatického tryptofanu a celkových kynureninů“. International Journal of Tryptophan Research. 7: 23–32. doi:10.4137 / IJTR.S18231. PMC 4259507. PMID 25520560.
- ^ A b Elango R, Chapman K, Rafii M, Ball RO, Pencharz PB (říjen 2012). „Stanovení přijatelné horní úrovně příjmu leucinu v akutních dietních studiích u mladých mužů“. American Journal of Clinical Nutrition. 96 (4): 759–67. doi:10.3945 / ajcn.111.024471. PMID 22952178.
Bylo pozorováno významné zvýšení koncentrací amoniaku v krvi nad normální hodnoty, plazmatických koncentrací leucinů a vylučování leucinů močí při příjmu leucinu> 500 mg · kg · 1 · d · 1. Oxidace l- [l-³³C] -leucinů vyjádřená jako značkovací stopová oxidace v dechu (F1³CO₂), oxidace leucinu a oxidace kyseliny α-ketoizokapronové (KIC) vedla k různým výsledkům: plató ve F1³CO₂ pozorované po 500 mg · kg Při oxidaci leucinu nebylo pozorováno žádné jasné plató a oxidace KIC se objevila na plató po 750 mg.kg-1. D1. Na základě plazmatických a močových proměnných lze jako opatrný odhad za akutních dietních podmínek navrhnout UL pro leucin u zdravých dospělých mužů v dávce 500 mg · kg · 1 · d · 1 nebo ~ 35 g / d.
- ^ Rasmussen B, Gilbert E, Turki A, Madden K, Elango R (červenec 2016). „Stanovení bezpečnosti doplňování leucinů u zdravých starších mužů“. Aminokyseliny. 48 (7): 1707–16. doi:10.1007 / s00726-016-2241-0. PMID 27138628. S2CID 3708265.
horní limit pro příjem leucinu u zdravých starších osob by mohl být stanoven podobně jako u mladých mužů na 500 mg kg-1 den-1 nebo ~ 35 g / den pro osobu vážící 70 kg
- ^ Etzel MR (duben 2004). „Výroba a použití mléčných proteinových frakcí“. The Journal of Nutrition. 134 (4): 996S – 1002S. doi:10.1093 / jn / 134.4.996S. PMID 15051860.
- ^ A b Kim JH, Lee C, Lee M, Wang H, Kim K, Park SJ, Yoon I, Jang J, Zhao H, Kim HK, Kwon NH, Jeong SJ, Yoo HC, Kim JH, Yang JS, Lee MY, Lee CW `` Yun J, Oh SJ, Kang JS, Martinis SA, Hwang KY, Guo M, Han G, Han JM, Kim S (září 2017). „Kontrola leucin-dependentní dráhy mTORC1 chemickým zásahem leucyl-tRNA syntetázy a RagD interakce“. Příroda komunikace. 8 (1): 732. Bibcode:2017NatCo ... 8..732K. doi:10.1038 / s41467-017-00785-0. PMC 5622079. PMID 28963468.
- ^ A b Jewell JL, Russell RC, Guan KL (březen 2013). „Aminokyselina signalizující před mTOR“. Nature Reviews Molecular Cell Biology. 14 (3): 133–9. doi:10.1038 / nrm3522. PMC 3988467. PMID 23361334.
- ^ Sancak Y, Peterson TR, Shaul YD, Lindquist RA, Thoreen CC, Bar-Peled L, Sabatini DM (červen 2008). „Rag GTPasy váží raptor a zprostředkovávají aminokyselinovou signalizaci na mTORC1“. Věda. 320 (5882): 1496–501. Bibcode:2008Sci ... 320.1496S. doi:10.1126 / science.1157535. PMC 2475333. PMID 18497260.
- ^ Wolfson RL, Chantranupong L, Saxton RA, Shen K, Scaria SM, Cantor JR, Sabatini DM (leden 2016). „Sestrin2 je leucinový senzor pro dráhu mTORC1“. Věda. 351 (6268): 43–8. Bibcode:2016Sci ... 351 ... 43W. doi:10.1126 / science.aab2674. PMC 4698017. PMID 26449471.
- ^ Saxton RA, Knockenhauer KE, Wolfson RL, Chantranupong L, Pacold ME, Wang T, Schwartz TU, Sabatini DM (leden 2016). „Strukturální základna pro snímání leucinů cestou Sestrin2-mTORC1“. Věda. 351 (6268): 53–8. Bibcode:2016Sci ... 351 ... 53S. doi:10.1126 / science.aad2087. PMC 4698039. PMID 26586190.
- ^ Chantranupong L, Wolfson RL, Orozco JM, Saxton RA, Scaria SM, Bar-Peled L, Spooner E, Isasa M, Gygi SP, Sabatini DM (říjen 2014). „Sestriny interagují s GATOR2, aby negativně regulovaly dráhu snímání aminokyselin před mTORC1“. Zprávy buněk. 9 (1): 1–8. doi:10.1016 / j.celrep.2014.09.014. PMC 4223866. PMID 25263562.
- ^ A b C „KEGG reakce: R10759“. Kyoto Encyclopedia of Genes and Genomes. Laboratoře Kanehisa. Archivováno z původního dne 1. července 2016. Citováno 24. června 2016.
- ^ A b C Mock DM, Stratton SL, Horvath TD, Bogusiewicz A, Matthews NI, Henrich CL, Dawson AM, Spencer HJ, Owen SN, Boysen G, Moran JH (listopad 2011). „Vylučování 3-hydroxyisovalerové kyseliny a 3-hydroxyisovalerylkarnitinu močí se zvyšuje u lidí s nedostatkem biotinu v reakci na výzvu leucinů“. primární zdroj. The Journal of Nutrition. 141 (11): 1925–1930. doi:10.3945 / jn.111.146126. PMC 3192457. PMID 21918059.
Snížená aktivita MCC zhoršuje katalýzu zásadního kroku v mitochondriálním katabolismu BCAA leucinů. Metabolické poškození odvádí methylkrotonyl CoA na 3-hydroxyisovaleryl CoA v reakci katalyzované enoyl-CoA hydratázou (22, 23). Akumulace 3-hydroxyisovaleryl CoA může inhibovat buněčné dýchání buď přímo, nebo prostřednictvím účinků na poměry acyl CoA: volný CoA, pokud nedojde k dalšímu metabolismu a detoxikaci 3-hydroxyisovaleryl CoA (22). Přenos na karnitin 4 karnitin acyl-CoA transferázami distribuovanými v subcelulárních kompartmentech pravděpodobně slouží jako důležitý rezervoár pro acylové skupiny (39–41). 3-Hydroxyisovaleryl CoA je pravděpodobně detoxikován karnitin-acetyltransferázou produkující 3HIA-karnitin, který je transportován přes vnitřní mitochondriální membránu (a tedy účinně z mitochondrií) přes translokázu karnitin-acylkarnitin (39). Předpokládá se, že 3HIA-karnitin je buď přímo deacylován hydrolázou na 3HIA, nebo podstoupí druhou výměnu CoA, aby znovu vytvořil 3-hydroxyisovaleryl CoA, následovaný uvolněním 3HIA a volného CoA thioesterázou.
- ^ A b Wilson JM, Fitschen PJ, Campbell B, Wilson GJ, Zanchi N, Taylor L, Wilborn C, Kalman DS, Stout JR, Hoffman JR, Ziegenfuss TN, Lopez HL, Kreider RB, Smith-Ryan AE, Antonio J (únor 2013) . „Stanoviště mezinárodní společnosti pro sportovní výživu: beta-hydroxy-beta-methylbutyrát (HMB)“. Journal of the International Society of Sports Nutrition. 10 (1): 6. doi:10.1186/1550-2783-10-6. PMC 3568064. PMID 23374455.
- ^ A b C d E F G h i j k l m n Kohlmeier M (květen 2015). "Leucin". Metabolismus živin: Struktury, funkce a geny (2. vyd.). Akademický tisk. 385–388. ISBN 978-0-12-387784-0. Archivováno z původního dne 22. března 2018. Citováno 6. června 2016.
Energetické palivo: Nakonec se většina Leu rozloží a poskytuje přibližně 6,0 kcal / g. Asi 60% požitého Leu se oxiduje během několika hodin ... Ketogeneze: Významná část (40% požité dávky) se přemění na acetyl-CoA a tím přispívá k syntéze ketonů, steroidů, mastných kyselin a dalších sloučeniny
Obrázek 8.57: Metabolismus L-leucin Archivováno 22. března 2018 v Wayback Machine - ^ Rosenthal J, Angel A, Farkas J (únor 1974). „Metabolický osud leucinu: významný prekurzor sterolu v tukové tkáni a svalech“. Dopoledne. J. Physiol. 226 (2): 411–8. doi:10.1152 / ajplegacy.1974.226.2.411. PMID 4855772.
- ^ Brioche T, Pagano AF, Py G, Chopard A (srpen 2016). „Svalová ztráta a stárnutí: experimentální modely, infiltrace tuků a prevence“ (PDF). Molekulární aspekty medicíny. 50: 56–87. doi:10.1016 / j.mam.2016.04.006. PMID 27106402.
Závěrem lze říci, že léčba HMB se jasně jeví jako bezpečná silná strategie proti sarkopenii a obecněji proti úbytku svalů, protože HMB zlepšuje svalovou hmotu, svalovou sílu a fyzický výkon. Zdá se, že HMB je schopen působit na tři ze čtyř hlavních mechanismů zapojených do svalové dekondice (obrat bílkovin, apoptóza a regenerační proces), zatímco u hypotézy se předpokládá, že silně ovlivní čtvrtý (mitochondriální dynamika a funkce). HMB je navíc levná (~ 30–50 USD za měsíc při 3 g denně) a může předcházet osteopenii (Bruckbauer a Zemel, 2013; Tatara, 2009; Tatara a kol., 2007, 2008, 2012) a snižovat kardiovaskulární rizika (Nissen et al., 2000). Ze všech těchto důvodů by se HMB měla běžně používat v podmínkách úbytku svalů, zejména u starších lidí. ... 3 g CaHMB užívaných třikrát denně (pokaždé 1 g) je optimální dávkování, které umožňuje kontinuální biologickou dostupnost HMB v těle (Wilson et al., 2013)
- ^ „KEGG reakce: R04137“. Kyoto Encyclopedia of Genes and Genomes. Laboratoře Kanehisa. Archivováno z původního dne 1. července 2016. Citováno 24. června 2016.
- ^ "Homo sapiens: 4-hydroxyfenylpyruvát dioxygenázová reakce". MetaCyc. SRI International. 20. srpna 2012. Citováno 6. června 2016.
- ^ A b "Leucinový metabolismus". BRENDA. Technische Universität Braunschweig. Archivovány od originál dne 17. srpna 2016. Citováno 12. srpna 2016.
- ^ Nelson, D.L .; Cox, M. M. „Lehninger, Principles of Biochemistry“ 3. vydání. Worth Publishing: New York, 2000. ISBN 1-57259-153-6.
- ^ Meierhenrich: Aminokyseliny a asymetrie života, Springer-Verlag, 2008, ISBN 978-3-540-76885-2.
{kind=link}
{kind=link}
externí odkazy
| Obecná témata |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Podle vlastností |
| ||||||||||
| |||||||||||