PIM2 (gen) - PIM2 (gene)





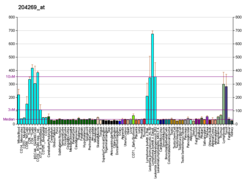
Serin / threonin-protein kináza Pim-2 je enzym že u lidí je kódován PIM2 .[5][6]
PIM2 nebo Provirální integrace viru Moloney 2 je serin / threonin kináza který má role v růst buněk šíření, apoptóza a regulace kaskády přenosu signálu.[7]
Struktura
Doposud byla většina strukturních informací náležejících k rodině PIM kináz omezena na PIM1. Výsledkem je, že většina úsilí o vývoj inhibitorů směřovala také k PIM1. PIM2 sdílí 55% sekvenční identitu s PIM1 a struktura PIM2 do značné míry souvisí s PIM1.[8] Stejně jako PIM1, PIM2 ukazuje bi-lobal kinázovou architekturu s konstitutivně aktivní uzavřenou konformací. Hlavní řetězec obou molekuly je totožný s výjimkou dvou flexibilních oblastí v N-terminálním laloku.

Nejvýznamnějším strukturálním rozdílem mezi PIM1 a PIM2 je absence koncové šroubovice αJ v PIM2. Posledních 23 zbytků PIM2 se zcela liší od PIM1, protože PIM2 obsahuje 6 prolin zbytky v této oblasti a nepředpokládá se, že tvoří stejné terciární struktury. Výsledkem je, že absence interakcí přítomných v této oblasti může zvýšit flexibilitu PIM2 v laloku N-terminální kinázy a přispět k neuspořádaným oblastem struktury PIM2.[9]
Funkce
PIM2 je exprimován s vysokými hladinami v mozek a lymfoidní buňky. Sloučenina PIM1-3 knockout myši kteří přežili perinatální období, vykazovali velké zmenšení velikosti těla. To naznačuje, že PIM enzymy jsou důležité pro růst těla.[10] Pokusy naznačují, že PIM1 a PIM2 jsou nezbytné pro proliferaci a přežití lymfocytů závislou na cytokinech.[11] Experimenty s transgenní myši s indukovaným lymfomy odhalil zvýšené hladiny PIM2 jako častou, ale pozdní událost v roce 2006 tumorigeneze.[12]
Pokusy provedené na jaderném faktoru κB (NFκB ) jaderná translokace u člověka perineurální invaze (PNI) odhalily, že up-regulace NFκB a jeho následného cíle, PIM2, byly součástí antiapoptotické signalizační kaskády, která je spojena s rakovinné buňky v PNI. Tato kaskáda může regulovat inhibici apoptózy. Studie také ukázala, že zvýšené hladiny PIM2 byly spojeny s PNI.[13] PIM2 kináza se proto ukázala jako klíčový lékový cíl k obnovení apoptózy u lidských rezistentních rakovin.[14][15][16]
Mechanismus
V hlášení krystalové struktury, PIM1 a PIM2 předpokládají aktivní konformaci. Typicky, kinázy “ aktivní stav je charakterizován přítomností konzervovaného lysin, konformace uzavřeného laloku a dobře strukturovaný aktivační segment. Aktivační segment často vyžaduje fosforylace aby tam bylo katalytická aktivita. Jakmile je fosforylován, aktivní segment se složí na spodní lalok a reorganizuje peptid - vazebné místo, které následně vede k enzymatické aktivaci. PIM kinázy jsou však katalyticky aktivní bez fosforylace. Krystalové struktury ukazují, že nefosforylovaný aktivační segment tvoří mnoho polárních interakcí s lalokem dolní kinázy, který stabilizuje aktivní konformaci. Zatímco PIM kináza ano autofosforylát, funkční důsledky nejsou známy.[17]
Inhibitory
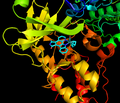
Inhibitor působí jako ATP mimetikum v ATP vazebné kapse

Staurosporin a sloučenina ruthenia 12

Obecná struktura inhibitoru organoruthenia



PIM2 (spolu s PIM1) má jedinečnou vázací kapsu pro ATP s kloubovou oblastí, což z něj dělá atraktivní cíl pro silný PIM s malou molekulou inhibitory kinázy.[18]
Mnoho inhibitorů je často selektivnějších pro PIM1 a PIM3 než PIM2. Jinými slovy, PIM2 je obvykle inhibován s mnohem nižší účinností. Strukturální modely zatím tento jev nedokážou vysvětlit. Mohlo by to však souviset s rozdíly v dynamických vlastnostech různých PIM izoformy.
V sérii organoruthenium sloučeniny [19] na základě a Staurosporin lešení[20] sloučenina 12 poskytla téměř úplnou inhibici při koncentraci 10 nM. Proti PIM1 však byl o něco méně účinný.
SAR naznačuje, že přidání potenciálních vodíkových vazebných skupin v polohách R1 a R2 dramaticky zvyšuje účinnost proti oběma kinázám. Podobná substituce polohy R3 byla méně účinná a substituce halogenem byla ještě rušivější.
Klinický význam
Ve studii se 48 pacienty, kteří měli non-Hodgkinův lymfom (NHL) a lymfocytární leukémie, exprese hPim-2 byla analyzována pomocí hybridizace in situ, kvantitativní RT-PCR a FACS analýza. Studie prokázaly vyšší hladiny exprese v NHL oproti normálním lymfocytům i v chronická lymfocytární leukémie přes normální B-buňky.[21]
Zvýšené hladiny PIM2 byly také nalezeny v primárních výbuchech z Akutní myeloidní leukémie pacientů. PIM2 může být důležitou kinázou při fosforylaci 4E-BP1. Konstitutivní fosforylace 4E-BP1 se běžně vyskytuje u rakoviny a přispívá k omezené translaci transkriptů souvisejících s malignitou, mezi které patří c-Myc a Cyclin D. Vyřazení PIM2 pomocí iRNA silně snížilo akumulaci onkogenních proteinů.[22] Ve výsledku může být PIM2 atraktivním cílem pro akutní myeloidní leukémii.
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000102096 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000031155 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Baytel D, Shalom S, Madgar I, Weissenberg R, Don J (prosinec 1998). „Lidský proto-onkogen Pim-2 a jeho testikulární exprese“. Biochim Biophys Acta. 1442 (2–3): 274–85. doi:10.1016 / s0167-4781 (98) 00185-7. PMID 9804974.
- ^ „Entrez Gene: PIM2 pim-2 onkogen“.
- ^ Alvarado Y, Giles FJ, Swords RT (2012). "PIM kinázy v hematologických rakovinách". Expert Rev Hematol. 5 (1): 81–96. doi:10,1586 / ehm.11,69. PMID 22272708.
- ^ Debreczeni JE, Bullock AN, Atilla GE, Williams DS, Bregman H, Knapp S, Meggers E (únor 2006). „Rutheniové polo sendvičové komplexy navázané na protein kinázu Pim-1“. Angewandte Chemie. 45 (10): 1580–5. doi:10.1002 / anie.200503468. PMID 16381041.
- ^ Bullock AN, Russo S, Amos A, Pagano N, Bregman H, Debreczeni JE, Lee WH, von Delft F, Meggers E, Knapp S (2005). "Krystalová struktura PIM2 kinázy v komplexu s inhibitorem organoruthenia". PLOS ONE. 4 (10): e7112. doi:10.1371 / journal.pone.0007112. PMC 2743286. PMID 19841674.
- ^ Mikkers H, Nawijn M, Allen J, Brouwers C, Verhoeven E, Jonkers J, Berns A (2004). „Myši nedostatečné pro všechny PIM kinázy vykazují zmenšenou velikost těla a zhoršenou reakci na hematopoetické růstové faktory“. Mol Cell Biol. 24 (13): 6104–6115. doi:10.1128 / MCB.24.13.6104-6115.2004. PMC 480904. PMID 15199164.
- ^ Tahvanainen J, Kyläniemi MK, Kanduri K, Gupta B, Lähteenmäki H, Kallonen T, Rajavuori A, Rasool O, Koskinen PJ, Rao KV, Lähdesmäki H, Lahesmaa R (2013). „Provirální integrační web pro kinázy viru moloney myší leukémie (PIM) podporuje diferenciaci buněk lidského T pomocníka 1“. The Journal of Biological Chemistry. 288 (5): 3048–3058. doi:10,1074 / jbc.M112.361709. PMC 3561529. PMID 23209281.
- ^ Breuer ML, Cuypers HT, Berns A (březen 1989). „Důkazy o zapojení pim-2, nového společného provirového inzertního místa, do progrese lymfomů“. Časopis EMBO. 8 (3): 743–8. doi:10.1002 / j.1460-2075.1989.tb03434.x. PMC 400870. PMID 2721500.
- ^ Ayala GE, Dai H, Ittmann M, Li R, Powell M, Frolov A a kol. (Září 2004). „Mechanismy růstu a přežití spojené s perineurální invazí u rakoviny prostaty“. Výzkum rakoviny. 64 (17): 6082–90. doi:10.1158 / 0008-5472.CAN-04-0838. PMID 15342391.
- ^ Giles F (2005). "Inhibitor Pim kinázy, prosím". Krev. 105 (11): 4158–4159. doi:10.1182 / krev-2005-03-1150.
- ^ Malone T, Schäfer L, Simon N, Heavey S, Cuffe S, Finn S a kol. (Březen 2020). „Současné perspektivy zaměřené na PIM kinázy k překonání mechanismů rezistence na léky a imunitní úniky u rakoviny“. Farmakologie a terapeutika. 207: 107454. doi:10.1016 / j.pharmthera.2019.107454. PMID 31836451.
- ^ Luszczak S, Kumar C, Sathyadevan VK, Simpson BS, Gately KA, Whitaker HC, Heavey S (2020). „Inhibice PIM kinázy: společné terapeutické přístupy u rakoviny prostaty“. Transdukce signálu a cílená terapie. 5: 7. doi:10.1038 / s41392-020-0109-r. PMC 6992635. PMID 32025342.
- ^ Brault L, Gasser C, Bracher F, Huber K, Knapp S, Schwaller J (2010). "PIM serin / threonin kinázy v patogenezi a terapii hematologických malignit a pevných nádorů". Haematologica. 95 (6): 1004–1015. doi:10 3324 / haematol.2009.017079. PMC 2878801. PMID 20145274.
- ^ Pogacic V, Bullock AN, Fedorov O, Filippakopoulos P, Gasser C, Biondi A, Meyer-Monard S, Knapp S, Schwaller J (červenec 2007). „Strukturální analýza identifikuje imidazo [1,2-b] pyridaziny jako inhibitory PIM kinázy s antileukemickou aktivitou in vitro“. Výzkum rakoviny. 67 (14): 6916–24. doi:10.1158 / 0008-5472.CAN-07-0320. PMID 17638903.
- ^ Pagano N, Maksimoska J, Bregman H, Williams DS, Webster RD, Xue F, Meggers E (duben 2007). „Rutheniové polo sendvičové komplexy jako inhibitory proteinkinázy: derivatizace pyridokarbazolového farmakoforového ligandu“. Organická a biomolekulární chemie. 5 (8): 1218–27. doi:10.1039 / b700433h. PMID 17406720.
- ^ Meggers E (červen 2007). "Zkoumání biologicky relevantního chemického prostoru s kovovými komplexy". Aktuální názor na chemickou biologii. 11 (3): 287–92. doi:10.1016 / j.cbpa.2007.05.013. PMID 17548234.
- ^ Cohen AM, Grinblat B, Bessler H, Kristt D, Kremer A, Schwartz A, Halperin M, Shalom S, Merkelová D, Don J (2004). "Zvýšená exprese genu hPim-2 v lidské chronické lymfocytární leukémii a nehodgkinském lymfomu". Leukémie a lymfom. 45 (5): 951–955. doi:10.1080/10428190310001641251. PMID 15291354.
- ^ Moerke NJ, Aktas H, Chen H, Cantel S, Reibarkh MY, Fahmy A, Gross JD, Degterev A, Yuan J, Chorev M, Halperin JA, Wagner G (leden 2007). "Inhibice interakce mezi faktory iniciace translace eIF4E a eIF4G s malou molekulou". Buňka. 128 (2): 257–67. doi:10.1016 / j.cell.2006.11.046. PMID 17254965.
Další čtení
- Yan B, Zemskova M, držitel S, Chin V, Kraft A, Koskinen PJ, Lilly M (2003). „PIM-2 kináza fosforyluje BAD na serinu 112 a reverzuje buněčnou smrt vyvolanou BAD“. J. Biol. Chem. 278 (46): 45358–67. doi:10,1074 / jbc.M307933200. PMID 12954615.
- Cohen AM, Grinblat B, Bessler H, Kristt D, Kremer A, Schwartz A, Halperin M, Shalom S, Merkelová D, Don J (2004). "Zvýšená exprese genu hPim-2 u lidské chronické lymfocytární leukémie a non-Hodgkinova lymfomu". Leuku. Lymfom. 45 (5): 951–5. doi:10.1080/10428190310001641251. PMID 15291354.
- Hammerman PS, Fox CJ, Cinalli RM, Xu A, Wagner JD, Lindsten T, Thompson CB (2005). „Transformace lymfocytů pomocí Pim-2 závisí na aktivaci nukleárního faktoru-kappaB“. Cancer Res. 64 (22): 8341–8. doi:10.1158 / 0008-5472.CAN-04-2284. PMID 15548703.
- Rainio EM, Ahlfors H, Carter KL, Ruuska M, Matikainen S, Kieff E, Koskinen PJ (2005). „Pim kinázy jsou během infekce virem Epstein-Barrové nadměrně regulovány a zvyšují aktivitu EBNA2“. Virologie. 333 (2): 201–6. doi:10.1016 / j.virol.2005.01.001. PMID 15721354.
- Dai H, Li R, Wheeler T, Diaz de Vivar A, Frolov A, Tahir S, Agoulnik I, Thompson T, Rowley D, Ayala G (2005). „Regulace Pim-2: biologické důsledky spojené s progresí onemocnění a perinuerální invazí u rakoviny prostaty“. Prostata. 65 (3): 276–86. doi:10.1002 / pros.20294. PMID 16015593.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (2005). „Směrem k mapě lidské interakční sítě protein-protein v měřítku proteomu“. Příroda. 437 (7062): 1173–8. doi:10.1038 / nature04209. PMID 16189514.
- Macdonald A, Campbell DG, Toth R, McLauchlan H, Hastie CJ, Arthur JS (2006). „Pim kinázy fosforylují více míst na Bad a podporují vazbu 14-3-3 a disociaci z Bcl-XL“. BMC Cell Biol. 7: 1. doi:10.1186/1471-2121-7-1. PMC 1368972. PMID 16403219.
externí odkazy
- Přehled všech strukturálních informací dostupných v PDB pro UniProt: Q9P1W9 (Serin / threonin-protein kináza pim-2) na PDBe-KB.
Galerie PDB | |
|---|---|
|
