Enzymatická katalýza - Enzyme catalysis
Enzymatická katalýza je nárůst v hodnotit a proces podle a biologická molekula „enzym ". Většina enzymů jsou bílkoviny a většina takových procesů jsou chemické reakce. V enzymu obvykle probíhá katalýza v lokalizovaném místě, které se nazývá Aktivní stránky.
Většina enzymy jsou vyrobeny převážně z proteinů, buď jediného proteinového řetězce nebo mnoha takových řetězců v a komplex více podjednotek. Enzymy často také obsahují nebílkovinné složky, jako jsou ionty kovů nebo specializované organické molekuly známé jako kofaktor (např. adenosintrifosfát ). Mnoho kofaktorů jsou vitamíny a jejich role vitaminů přímo souvisí s jejich použitím při katalýze biologických procesů v metabolismu. Katalýza z biochemické reakce v buňka je životně důležitý, protože mnoho, ale ne všechny metabolicky esenciální reakce, mají při katalýze velmi nízké rychlosti. Jedním z faktorů evoluce bílkovin je optimalizace těchto katalytických aktivit, i když pouze nejdůležitější enzymy pracují poblíž limitů katalytické účinnosti a mnoho enzymů není zdaleka optimální. Mezi důležité faktory při enzymové katalýze patří obecná kyselá a bazická katalýza, orbitální řízení, entropické omezení, orientační účinky (tj. Klíčová a klíčová katalýza), jakož i pohybové účinky zahrnující dynamika bílkovin[1]
Mechanismy enzymové katalýzy se liší, ale všechny jsou v zásadě podobné jiným typům chemická katalýza v tom, že rozhodujícím faktorem je snížení energetické bariéry, která odděluje reaktanty od produktů[2]. Snížení o aktivační energie (EA) zvyšuje podíl molekul reaktantů, které mohou překonat tuto bariéru a vytvořit produkt. Důležitým principem je, že protože pouze snižují energetické bariéry mezi produkty a reaktanty, enzymy vždy katalyzují reakce v obou směrech a nemohou řídit reakci vpřed nebo ovlivnit rovnovážnou pozici - pouze rychlost, s níž je dosaženo. Stejně jako u jiných katalyzátorů není enzym reakcí spotřebován nebo měněn (jako substrát), ale je recyklován tak, že jediný enzym provádí mnoho cyklů katalýzy.
Vyvolaný střih
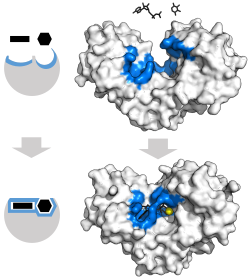
Klasický model pro enzymPodklad interakce je model indukovaného přizpůsobení.[3] Tento model navrhuje, že počáteční interakce mezi enzymem a substrátem je relativně slabá, ale že tyto slabé interakce rychle vyvolávají konformační změny v enzymu, který posiluje vazbu.
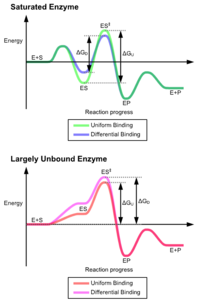
Výhody mechanismu indukovaného přizpůsobení vznikají v důsledku stabilizačního účinku silné vazby enzymu. Existují dva různé mechanismy vazby na substrát: uniformní vazba, která má silnou vazbu na substrát, a diferenciální vazba, která má silnou vazbu na přechodový stav. Stabilizační účinek rovnoměrné vazby zvyšuje jak vazebnou afinitu substrátu, tak přechodového stavu, zatímco diferenciální vazba zvyšuje pouze vazebnou afinitu přechodového stavu. Oba jsou používány enzymy a byly evolučně vybrány tak, aby se minimalizovala aktivační energie reakce. Enzymy, které jsou nasycené, tj. Mají vysoce afinitní vazbu na substrát, vyžadují diferenciální vazbu, aby se snížila energie aktivace, zatímco malé enzymy nevázané na substrát mohou používat buď diferenciální nebo jednotnou vazbu.[4]
Tyto účinky vedly k tomu, že většina proteinů používá mechanismus diferenciální vazby ke snížení energie aktivace, takže většina substrátů má vysokou afinitu k enzymu v přechodném stavu. Diferenciální vazba se provádí mechanismem indukovaného přizpůsobení - substrát se nejprve váže slabě, poté enzym mění konformaci, zvyšuje afinitu k přechodnému stavu a stabilizuje jej, čímž snižuje aktivační energii k jeho dosažení.
Je však důležité objasnit, že koncept indukovaného přizpůsobení nelze použít k racionalizaci katalýzy. To znamená, že chemická katalýza je definována jako redukce EA‡ (když je systém již v ES‡) ve vztahu k EA‡ v nekatalyzované reakci ve vodě (bez enzymu). Indukované přizpůsobení pouze naznačuje, že bariéra je nižší v uzavřené formě enzymu, ale neříká nám, co je důvodem redukce bariéry.
Indukované přizpůsobení může být prospěšné pro věrnost molekulárního rozpoznávání v přítomnosti konkurence a šumu prostřednictvím konformační korektury mechanismus.[5]
Mechanismy alternativní reakční cesty
Tyto konformační změny také přinášejí katalytické zbytky v Aktivní stránky blízké chemickým vazbám v substrátu, které se při reakci změní. Po navázání jeden nebo více mechanismů katalýzy snižuje energii reakce přechodový stav, poskytnutím alternativní chemické cesty pro reakci. Existuje šest možných mechanismů katalýzy „přes bariéru“, jakož i mechanismů „přes bariéru“:
Blízkost a orientace
Interakce enzym-substrát srovnávají reaktivní chemické skupiny a drží je blízko sebe v optimální geometrii, což zvyšuje rychlost reakce. Tím se snižuje entropie reaktantů, a tak činí adiční nebo přenosové reakce méně nepříznivými, protože snížení celkové entropie, když se dva reaktanty stanou jediným produktem. Jedná se však o obecný účinek, který se projevuje v nepřidávacích nebo přenosových reakcích, kde k nim dochází v důsledku zvýšení „účinné koncentrace“ činidel. To se rozumí, když se vezme v úvahu, jak zvýšení koncentrace vede ke zvýšení reakční rychlosti: v podstatě, když jsou reaktanty koncentrovanější, častěji se srazí, a tak reagují častěji. V případě enzymové katalýzy vazba činidel na enzym omezuje konformační prostor reaktantů a udržuje je ve „správné orientaci“ a blízko sebe, takže kolidují častěji a se správnou geometrií, aby usnadnily požadovaná reakce. „Efektivní koncentrace“ je koncentrace, kterou by reaktant musel být, aby byl volný v roztoku, aby mohl zažít stejnou kolizní frekvenci. Takové teoretické efektivní koncentrace jsou často nefyzikální a ve skutečnosti je nemožné je realizovat - což svědčí o velké katalytické síle mnoha enzymů s masivním nárůstem rychlosti v nekatalyzovaném stavu.
| Například: |
| Podobné reakce proběhnou mnohem rychleji, pokud je reakce intramolekulární. |
 |
| Efektivní koncentrace acetát v intramolekulární reakci lze odhadnout jako k2/ k1 = 2 x 105 Molární. |
Situace však může být složitější, protože moderní výpočetní studie prokázaly, že tradiční příklady účinků blízkosti nelze přímo spojit s entropickými účinky enzymu.[6][7][8] Také původní entropický návrh[9] bylo zjištěno, že do značné míry nadhodnocuje příspěvek orientační entropie ke katalýze.[10]
Dárci nebo akceptory protonů
Dárci a akceptory protonů, tj. kyseliny a základna může darovat a přijímat protony za účelem stabilizace vývojových nábojů v přechodném stavu. To souvisí s celkovým principem katalýzy, spočívajícím ve snižování energetických bariér, protože přechodové stavy jsou obecně vysokoenergetické stavy a jejich stabilizací se tato vysoká energie snižuje, čímž se bariéra snižuje. Klíčovým rysem enzymové katalýzy u mnoha nebiologických katalýz je to, že při stejné reakci lze kombinovat jak kyselou, tak bazickou katalýzu. V mnoha abiotických systémech mohou kyseliny (velké [H +]) nebo báze (velké koncentrace H + klesá nebo druhy s elektronovými páry) zvýšit rychlost reakce; ale prostředí samozřejmě může mít pouze jedno celkové pH (měřítko kyselosti nebo zásaditosti (zásaditosti)). Vzhledem k tomu, že enzymy jsou velké molekuly, mohou umístit jak kyselé skupiny, tak bazické skupiny do svého aktivního místa, aby interagovaly se svými substráty, a používají oba režimy nezávisle na objemovém pH.
K aktivaci se často používá obecná kyselá nebo bazická katalýza nukleofil a / nebo elektrofil nebo stabilizovat odcházející skupiny. V aktivním místě se používá mnoho aminokyselin s kyselými nebo zásaditými skupinami, jako je kyselina glutamová a aspartová, histidin, cystin, tyrosin, lysin a arginin, stejně jako serin a threonin. Kromě toho se často používá peptidový hlavní řetězec s karbonylovými a amidovými N skupinami. Cystine a Histidin jsou velmi často zapojeni, protože oba mají pKa téměř neutrální pH a proto může přijímat i darovat protony.
Mnoho reakčních mechanismů zahrnujících acidobazickou katalýzu předpokládá podstatně pozměněnou pKa. Tato změna pKa je možná prostřednictvím místního prostředí zbytku[Citace je zapotřebí ].
| Podmínky | Kyseliny | Základny |
|---|---|---|
| Hydrofobní prostředí | Zvyšte pKa | Snížení pKa |
| Sousední zbytky podobného náboje | Zvyšte pKa | Snížení pKa |
| Solný most (a vodík tvorba vazby) | Snížení pKa | Zvyšte pKa |
pKa může být také významně ovlivněna okolním prostředím, a to do té míry, že zbytky, které jsou zásadité v roztoku, mohou působit jako donory protonů a naopak.
| Například: |
| Katalytická triáda a serinová proteáza |
 |
| Počáteční krok katalytického mechanismu serinové proteázy zahrnuje histidin aktivního místa přijímající proton ze serinového zbytku. To připravuje serin jako nukleofil k útoku na amidovou vazbu substrátu. Tento mechanismus zahrnuje darování protonu ze serinu (báze, pKa 14) histidinu (kyselina, pKa 6), což je možné díky místnímu prostředí bází. |
Je důležité objasnit, že modifikace pKa je čistou součástí elektrostatického mechanismu.[11] Kromě toho je katalytický účinek výše uvedeného příkladu spojen hlavně se snížením pKa oxyaniontu a zvýšením pKa histidinu, zatímco přenos protonů ze serinu na histidin není významně katalyzován, protože není bariéra určující míru.[12]
Elektrostatická katalýza
Stabilizace nabitých přechodových stavů může být také pomocí zbytků ve formování aktivního místa iontové vazby (nebo částečné interakce iontového náboje) s meziproduktem. Tyto vazby mohou buď pocházet kyselé nebo základní boční řetězy nalezené na aminokyseliny jako lysin, arginin, kyselina asparagová nebo kyselina glutamová nebo pocházejí z kovu kofaktory jako zinek. Kovové ionty jsou obzvláště účinné a mohou dostatečně snížit pKa vody, aby se stal účinným nukleofilem.
Systematické počítačové simulační studie prokázaly, že elektrostatické jevy zdaleka nejvíce přispívají ke katalýze.[11] Může zvýšit rychlost reakce až o faktor 107.[13] Zejména bylo zjištěno, že enzym poskytuje prostředí, které je polárnější než voda, a že iontové přechodové stavy jsou stabilizovány pevnými dipóly. To se velmi liší od stabilizace přechodového stavu ve vodě, kde molekuly vody musí platit „reorganizační energií“.[14] Za účelem stabilizace iontových a nabitých stavů. Katalýza je tedy spojena se skutečností, že polární skupiny enzymu jsou předorganizovány [15]
Ukázalo se, že velikost elektrostatického pole vyvíjeného aktivním místem enzymu vysoce koreluje se zvýšením katalytické rychlosti enzymu[16][17]
Vazba substrátu obvykle vylučuje vodu z aktivního místa, čímž snižuje místní dielektrickou konstantu na konstantu organického rozpouštědla. To posiluje elektrostatické interakce mezi nabitými / polárními substráty a aktivními místy. Studie navíc ukázaly, že distribuce nábojů kolem aktivních míst jsou uspořádána tak, aby stabilizovala přechodové stavy katalyzovaných reakcí. V několika enzymech tyto distribuce náboje zjevně slouží k vedení polárních substrátů k jejich vazebným místům, takže rychlosti těchto enzymatických reakcí jsou větší než jejich zdánlivé limity řízené difúzí[Citace je zapotřebí ].
| Například: |
| Karboxypeptidáza katalytický mechanismus |
 |
| Čtyřboký meziprodukt je stabilizován částečnou iontovou vazbou mezi Zn2+ iont a negativní náboj na kyslíku. |
Kovalentní katalýza
Kovalentní katalýza zahrnuje substrát tvořící přechodný jev kovalentní vazba se zbytky v aktivním místě enzymu nebo s kofaktorem. To přidává do reakce další kovalentní meziprodukt a pomáhá snížit energii pozdějších přechodových stavů reakce. Kovalentní vazba musí být v pozdější fázi reakce přerušena, aby se enzym regeneroval. Tento mechanismus využívá katalytická triáda enzymů, jako je proteázy jako chymotrypsin a trypsin, kde se tvoří meziprodukt acyl-enzym. Alternativní mechanismus je Schiff základna formace pomocí bezplatného amin od a lysin reziduum, jak je vidět v enzymu aldoláza v průběhu glykolýza.
Některé enzymy využívají neaminokyselinu kofaktory jako pyridoxal fosfát (PLP) nebo thiamin pyrofosfát (TPP) za vzniku kovalentních meziproduktů s molekulami reaktantů.[18][19] Tyto kovalentní meziprodukty fungují tak, aby snižovaly energii pozdějších přechodových stavů, podobně jako kovalentní meziprodukty vytvořené s aminokyselinovými zbytky aktivního místa umožňují stabilizaci, ale schopnosti kofaktorů umožňují enzymům provádět reakce, které samotné aminokyselinové zbytky nemohou. Enzymy využívající takové kofaktory zahrnují enzym závislý na PLP aspartát transamináza a enzym závislý na TPP pyruvátdehydrogenáza.[20][21]
Místo snížení aktivační energie pro reakční cestu poskytuje alternativu kovalentní katalýza cesta pro reakci (prostřednictvím kovalentního meziproduktu) a tak se liší od skutečné katalýzy.[11] Například energetika kovalentní vazby na molekulu serinu v chymotrypsinu by měla být srovnávána s dobře známou kovalentní vazbou na nukleofil v reakci nekatalyzovaného roztoku. Skutečný návrh kovalentní katalýzy (kde je bariéra nižší než odpovídající bariéra v roztoku) by vyžadoval například částečnou kovalentní vazbu k přechodnému stavu pomocí skupiny enzymů (např. Velmi silná vodíková vazba) a tak účinky významně nepřispívají ke katalýze.
Katalýza kovovými ionty
Kovový iont v aktivním místě se účastní katalýzy koordinací stabilizace náboje a stínění. Díky kladnému náboji kovu lze pomocí kovových iontů stabilizovat pouze záporné náboje.[22] Kovové ionty jsou však při biologické katalýze výhodné, protože nejsou ovlivněny změnami pH.[23] Kovové ionty mohou také působit na ionizaci vody působením jako Lewisova kyselina.[24] Kovové ionty mohou být také činiteli oxidace a redukce.[25]
Bond kmen
Toto je hlavní účinek indukované vazby fit, kde je afinita enzymu k přechodnému stavu větší než k samotnému substrátu. To indukuje strukturální přesmyky, které napínají vazby substrátu do polohy blíže ke konformaci přechodového stavu, čímž se snižuje energetický rozdíl mezi substrátem a přechodovým stavem a pomáhá se katalyzovat reakce.
Účinek přetvoření je ve skutečnosti spíše destabilizačním účinkem než základním přechodovým stavem.[11][26][stránka potřebná ] Enzymy jsou navíc velmi flexibilní a nemohou aplikovat účinek velkého kmene.[27]
Kromě vazebného kmene v substrátu může být vazebný kmen indukován také v samotném enzymu, aby aktivoval zbytky v aktivním místě.
| Například: |
| Substrát, vázaný substrát a konformace přechodového stavu lysozym. |
| Substrát, po navázání, je zkreslen z konformace polovičního křesla hexózového kruhu (kvůli sterické zábraně s aminokyselinami proteinu nutícími ekvatoriální c6 být v axiální poloze) do konformační křesla[28][stránka potřebná ] |
Kvantové tunelování
Tyto tradiční mechanismy „přes bariéru“ byly v některých případech zpochybněny modely a pozorováním mechanismů „přes bariéru“ (kvantové tunelování ). Některé enzymy pracují s kinetikou, která je rychlejší než to, co by předpovídal klasický ΔG‡. V modelech „skrz bariéru“ může proton nebo elektron tunelovat aktivačními bariérami.[29][30] Kvantové tunelování protonů bylo pozorováno v roce tryptamin oxidace aromatická aminodehydrogenáza.[31]
Zdá se, že kvantové tunelování neposkytuje významnou katalytickou výhodu, protože příspěvky tunelování jsou podobné v katalyzovaných i nekatalyzovaných reakcích v roztoku.[30][32][33][34] Příspěvek tunelování (obvykle zvýšení rychlostních konstant o faktor ~ 1000[31] ve srovnání s rychlostí reakce pro klasickou cestu „přes bariéru“) je pravděpodobně zásadní pro životaschopnost biologických organismů. To zdůrazňuje obecný význam tunelovacích reakcí v biologii.
V letech 1971-1972 byl formulován první kvantově-mechanický model enzymové katalýzy.[35][36][je potřeba zdroj třetí strany ]
Aktivní enzym
Vazebnou energii komplexu enzym-substrát nelze považovat za vnější energii, která je nezbytná pro aktivaci substrátu. Enzym s vysokým obsahem energie může nejprve přenášet určitou specifickou energetickou skupinu X1 z katalytického místa enzymu na konečné místo prvního vázaného reaktantu, pak další skupiny X2 z druhého vázaného reaktantu (nebo z druhé skupiny jediného reaktantu) musí být přenesen na aktivní místo, aby se dokončila přeměna substrátu na produkt a regeneraci enzymu.[37]
Můžeme prezentovat celou enzymatickou reakci jako dvě vazebné reakce:
(1)

(2)

Je to vidět z reakce (1), že skupina X1 aktivního enzymu se objeví v produktu kvůli možnosti výměnné reakce uvnitř enzymu, aby se zabránilo jak elektrostatické inhibici, tak odpuzování atomů. Představujeme tedy aktivní enzym jako silný reaktant enzymatické reakce. Reakce (2) ukazuje neúplnou přeměnu substrátu, protože jeho skupina X2 zůstává uvnitř enzymu. Tento přístup jako myšlenka dříve navrhoval spoléhat se na hypotetické extrémně vysoké enzymatické přeměny (katalyticky dokonalý enzym).[38]
Klíčovým bodem pro ověření současného přístupu je, že katalyzátorem musí být komplex enzymu s přenosovou skupinou reakce. Tento chemický aspekt je podporován dobře prostudovanými mechanismy několika enzymatických reakcí. Zvažte reakci hydrolýzy peptidové vazby katalyzovanou čistým proteinem α-chymotrypsinem (enzym působící bez kofaktoru), který je dobře studovaným členem rodiny serinových proteáz, viz.[39]
Výsledky experimentu pro tuto reakci uvádíme ve dvou chemických krocích:
(3)

(4)

kde S1 je polypeptid, P1 a P2 jsou produkty. První chemický krok (3) zahrnuje tvorbu kovalentního meziproduktu acyl-enzymu. Druhý krok (4) je krok deacylace. Je důležité si uvědomit, že skupina H +, která se původně nacházela na enzymu, ale ne ve vodě, se objevuje v produktu před krokem hydrolýzy, proto ji lze považovat za další skupinu enzymatické reakce.
Reakce (3) ukazuje, že enzym působí jako silný reaktant reakce. Podle navrhovaného konceptu transport H z enzymu podporuje první konverzi reaktantů, rozpad první počáteční chemické vazby (mezi skupinami P1 a P2). Krok hydrolýzy vede k rozpadu druhé chemické vazby a regeneraci enzymu.
Navrhovaný chemický mechanismus nezávisí na koncentraci substrátů nebo produktů v médiu. Posun v jejich koncentraci však způsobuje hlavně změny volné energie v prvním a posledním kroku reakcí (1) a (2) v důsledku změn ve volném energetickém obsahu každé molekuly, ať už S nebo P, ve vodném roztoku. Tento přístup je v souladu s následujícím mechanismem svalové kontrakce. Posledním krokem hydrolýzy ATP v kosterním svalu je uvolňování produktu způsobené asociací hlav myosinu s aktinem.[40] Uzavření rozštěpu vázajícího aktin během asociační reakce je strukturálně spojeno s otevřením kapsy vázající nukleotidy na aktivním místě myosinu.[41]
Závěrečné kroky hydrolýzy ATP zahrnují zejména rychlé uvolňování fosfátu a pomalé uvolňování ADP.[42][43]Uvolňování fosfátového aniontu z vázaného aniontu ADP do vodného roztoku lze považovat za exergonickou reakci, protože fosfátový anion má nízkou molekulovou hmotnost.
Dospíváme tedy k závěru, že primární uvolňování anorganického fosfátu H je2PO4− vede k transformaci významné části volné energie hydrolýzy ATP na kinetickou energii solvatovaného fosfátu, produkující aktivní streaming. Tento předpoklad lokální mechanicko-chemické transdukce je v souladu s Tiroshovým mechanismem svalové kontrakce, kde svalová síla pochází z integrovaného působení aktivního proudu vytvořeného hydrolýzou ATP.[44][45]
Příklady katalytických mechanismů
Ve skutečnosti většina enzymových mechanismů zahrnuje kombinaci několika různých typů katalýzy.
Triosa fosfát izomeráza
Triosa fosfát izomeráza (ES 5.3.1.1 ) katalyzuje reverzibilní přeměnu obou triose fosfáty izomery dihydroxyaceton fosfát a D-glyceraldehyd-3-fosfát.
Trypsin
Trypsin (ES 3.4.21.4 ) je serinová proteáza který štěpí protein substráty po lysin nebo arginin zbytky pomocí a katalytická triáda provést kovalentní katalýzu a oxyanionový otvor stabilizovat hromadění náboje na přechodové stavy.
Aldoláza
Aldoláza (ES 4.1.2.13 ) katalyzuje rozpad fruktóza 1,6-bisfosfát (F-1,6-BP) do glyceraldehyd-3-fosfát a dihydroxyaceton fosfát (DHAP ).
Difuzivita enzymu
Příchod studie s jednou molekulou vedl v roce 2010 k pozorování, že pohyb nevazovaných enzymů se zvyšuje se zvyšující se koncentrací substrátu a zvyšuje se entalpie reakce.[46] Následná pozorování naznačují, že tento nárůst v difuzivita je poháněn přechodným vytěsňováním enzymu těžiště, což má za následek „zpětný ráz, který pohání enzym“.[47]
Podobnost reakce
Podobnost mezi enzymatickými reakcemi (ES ) lze vypočítat pomocí změn vazeb, reakčních center nebo metrik spodní stavby (EC-BLAST ).[48]
Viz také
- Katalytická triáda
- Stanovení enzymu
- Kinetika enzymů
- Enzymová promiskuita
- Dynamika proteinů
- Pseudoenzymy, jejichž všudypřítomnost navzdory jejich katalytické nečinnosti naznačuje omic Důsledky
- Kvantové tunelování
- Mapa proteolýzy
- Čas vyřešil krystalografii
Reference
- ^ Kamerlin, S. C .; Warshel, A (2010). „Na úsvitu 21. století: Je dynamika chybějícím článkem pro pochopení enzymové katalýzy?“. Proteiny: struktura, funkce a bioinformatika. 78 (6): 1339–75. doi:10,1002 / prot.22654. PMC 2841229. PMID 20099310.
- ^ Srinivasan, Bharath (27. září 2020). „Slova rady: výuka kinetiky enzymů“. FEBS Journal. doi:10.1111 / febs.15537. ISSN 1742-464X.
- ^ Koshland DE (Únor 1958). „Aplikace teorie specificity enzymů na syntézu proteinů“. Proc. Natl. Acad. Sci. USA 44 (2): 98–104. Bibcode:1958PNAS ... 44 ... 98K. doi:10.1073 / pnas.44.2.98. PMC 335371. PMID 16590179.

- ^ Anslyn, E.V .; Dougherty, D.A. (2006). Moderní fyzikální organická chemie. University Science Books. ISBN 978-1-891389-31-3.
- ^ Savir Y; Tlustý T (2007). Scalas, Enrico (ed.). „Konformační korektury: Dopad konformačních změn na specifičnost molekulárního rozpoznávání“ (PDF). PLOS One. 2 (5): e468. Bibcode:2007PLoSO ... 2..468S. doi:10.1371 / journal.pone.0000468. PMC 1868595. PMID 17520027. Archivovány od originál (PDF) dne 14. května 2011. Citováno 22. srpna 2010.
- ^ Stanton, R.V .; Perakyla, M .; Bakowies, D .; Kollman, P.A. (1998). „Kombinované výpočty ab initio a volné energie ke studiu reakcí v enzymech a roztoku: hydrolýza amidů v trypsinu a vodném roztoku“. J. Am. Chem. Soc. 120 (14): 3448–3457. doi:10.1021 / ja972723x.
- ^ Kuhn, B .; Kollman, P.A. (2000). „Výpočty QM-FE a molekulární dynamiky na katechol-O-methyltransferáze: volná energie aktivace v enzymu a ve vodném roztoku a regioselektivita reakce katalyzované enzymem“. J. Am. Chem. Soc. 122 (11): 2586–2596. doi:10.1021 / ja992218v.
- ^ Bruice, T.C .; Lightstone, FC (1999). „Příspěvky základního stavu a přechodového stavu k sazbám intramolekulárních a enzymatických reakcí“. Acc. Chem. Res. 32 (2): 127–136. doi:10.1021 / ar960131y.
- ^ Page, M. I.; Jencks, W.P. (1971). „Entropické příspěvky k rychlosti zrychlení v enzymatických a intramolekulárních reakcích a chelátový efekt“. Proc. Natl. Acad. Sci. USA. 68 (8): 1678–1683. Bibcode:1971PNAS ... 68.1678P. doi:10.1073 / pnas.68.8.1678. PMC 389269. PMID 5288752.
- ^ Warshel, A .; Parson, W.W. (2001). „Dynamika biochemických a biofyzikálních reakcí: Pohled z počítačových simulací“. Kvart. Biophys. 34 (4): 563–679. doi:10.1017 / s0033583501003730. PMID 11852595.
- ^ A b C d Warshel, A .; Sharma, P.K .; Kato, M .; Xiang, Y .; Liu, H .; Olsson, M.H.M. (2006). "Elektrostatický základ enzymové katalýzy". Chem. Rev. 106 (8): 3210–3235. doi:10.1021 / cr0503106. PMID 16895325.
- ^ Warshel, A .; Naray-Szabo, G .; Sussman, F .; Hwang, J.-K. (1989). "Jak skutečně fungují serinové proteázy?". Biochemie. 28 (9): 3629–37. doi:10.1021 / bi00435a001. PMID 2665806.
- ^ Voet, Donalde. (policista 2011). Biochemie. John Wiley & Sons. OCLC 808679090. Zkontrolujte hodnoty data v:
| datum =(Pomoc) - ^ Marcus, R. A. (1965). „K teorii reakcí přenosu elektronů. VI. Jednotné zpracování homogenních a elektrodových reakcí“ (PDF). J. Chem. Phys. 43 (2): 679–701. Bibcode:1965JChPh..43..679M. doi:10.1063/1.1696792.
- ^ Warshel, A (1978). „Energetika enzymové katalýzy“. Proc. Natl. Acad. Sci. USA. 75 (11): 5250–5254. Bibcode:1978PNAS ... 75,5250 W.. doi:10.1073 / pnas.75.11.5250. PMC 392938. PMID 281676.
- ^ „Jak fungují enzymy“
- ^ „EXTRÉMNÍ KATALÝZA VÝKONU ELEKTRICKÝCH POLÍ NA AKTIVNÍM STRÁNKÁCH KETOSTEROIDOVÉ izomery“, [1]
- ^ Toney, M. D. "Specifičnost reakce v pyridoxalových enzymech." Archivy biochemie a biofyziky (2005) 433: 279-287
- ^ Informační centrum o mikroživinách, Oregonská státní univerzita
- ^ Voet, Donald; Judith Voet (2004). Biochemie. John Wiley & Sons Inc. str.986–989. ISBN 978-0-471-25090-6.
- ^ Voet, Donald; Judith Voet (2004). Biochemie. John Wiley & Sons Inc. str.604–606. ISBN 978-0-471-25090-6.
- ^ Piccirilli, Joseph A .; Vyle, Joseph S .; Caruthers, Marvin H .; Cech, Thomas R. (7. ledna 1993). "Katalýza kovovými ionty v ribozymové reakci Tetrahymena". Příroda. 361 (6407): 85–88. Bibcode:1993Natur.361 ... 85P. doi:10.1038 / 361085a0. PMID 8421499. S2CID 4326584.
- ^ BENDER, MYRON L. (1. ledna 1962). "Kovová iontová katalýza nukleofilních organických reakcí v roztoku". Reakce koordinovaných ligandů. Pokroky v chemii. 37. AMERICKÁ CHEMICKÁ SPOLEČNOST. 19–36. doi:10.1021 / ba-1963-0037.ch002. ISBN 978-0841200388.
- ^ Fife, Thomas H .; Przystas, Theodore J. (1. února 1985). „Katalýza dvojmocných kovových iontů při hydrolýze esterů kyseliny pikolinové. Kovové ionty podporovaly hydroxidové ionty a reakce katalyzované vodou“. Journal of the American Chemical Society. 107 (4): 1041–1047. doi:10.1021 / ja00290a048. ISSN 0002-7863.
- ^ Stadtman, E. R. (1. ledna 1990). „Oxidace proteinů katalyzovaná ionty kovů: biochemický mechanismus a biologické důsledky“. Radikální biologie a medicína zdarma. 9 (4): 315–325. doi:10.1016/0891-5849(90)90006-5. ISSN 0891-5849. PMID 2283087.
- ^ Jencks, William P. (1987) [1969]. Katalýza v chemii a enzymologii. Série McGraw-Hill v pokročilé chemii (dotisk ed.). New York: Dover Publications. ISBN 9780486654607.
- ^ Warshel, A .; Levitt, M. (1976). „Teoretické studie enzymatických reakcí: dielektrická elektrostatická a sterická stabilizace karboniového iontu při reakci lyzozymu“. Journal of Molecular Biology. 103 (2): 227–49. doi:10.1016/0022-2836(76)90311-9. PMID 985660.
- ^
, který má podobný tvar jako přechodový stav. - ^ Garcia-Viloca, M; Gao, J; Karplus, M; Truhlar, DG (2004). "Jak fungují enzymy: analýza pomocí teorie moderní rychlosti a počítačových simulací". Věda. 303 (5655): 186–95. Bibcode:2004Sci ... 303..186G. doi:10.1126 / science.1088172. PMID 14716003. S2CID 17498715.
- ^ A b Olsson, MH; Siegbahn, PE; Warshel, A (2004). „Simulace účinku velkého kinetického izotopu a teplotní závislosti přenosu atomu vodíku v lipoxygenáze“. Journal of the American Chemical Society. 126 (9): 2820–8. doi:10.1021 / ja037233l. PMID 14995199.
- ^ A b Masgrau, L; Roujeinikova, A; Johannissen, LO; Hothi, P; Basran, J; Ranaghan, KE; Mulholland, AJ; Sutcliffe, MJ; et al. (2006). "Atomový popis enzymové reakce, v níž dominuje protonové tunelování". Věda. 312 (5771): 237–41. Bibcode:2006Sci ... 312..237M. doi:10.1126 / science.1126002. PMID 16614214. S2CID 27201250.
- ^ Hwang, J.-K .; Warshel, A. (1996). "Jak důležité jsou kvantově mechanické jaderné pohyby v enzymové katalýze". J. Am. Chem. Soc. 118 (47): 11745–11751. doi:10.1021 / ja962007f.
- ^ Ball, P. (2004). „Enzymy: Náhodou nebo záměrně?“. Příroda. 431 (7007): 396–397. Bibcode:2004 Natur.431..396B. doi:10.1038 / 431396a. PMID 15385982. S2CID 228263.
- ^ Olsson, M.H.M .; Parson, W.W .; Warshel, A. (2006). "Dynamické příspěvky k enzymové katalýze: kritické testy populární hypotézy". Chem. Rev. 106 (5): 1737–1756. doi:10.1021 / cr040427e. PMID 16683752.
- ^ Volkenshtein M.V., Dogonadze R.R., Madumarov A.K., Urushadze Z.D., Kharkats Yu.I. Teorie enzymové katalýzy.- Molekuliarnaya Biologia, Moskva, 6, 1972, 431-439
- ^ Volkenshtein M.V., Dogonadze R.R., Madumarov A.K., Urushadze Z.D., Kharkats Yu.I. Elektronické a konformační interakce při enzymové katalýze. In: E.L. Andronikashvili (vyd.), Konformatsionnie Izmenenia Biopolimerov v Rastvorakh, nakladatelství „Nauka“, Moskva, 1973, 153-157
- ^ Foigel, Alexander G. (2011). „Je enzym silným reaktantem biochemické reakce?“. Molekulární a buněčná biochemie. 352 (1–2): 87–9. doi:10.1007 / s11010-011-0742-4. PMID 21318350. S2CID 11133081.
- ^ Fogel, A.G. (1982). "Kooperativita enzymatických reakcí a molekulární aspekty transdukce energie". Mol. Buňka. Biochem. 47 (1): 59–64. doi:10.1007 / bf00241567. PMID 7132966. S2CID 21790380.
- ^ Hengge, AC; Stein, RL (2004). "Role konformační mobility proteinů v enzymové katalýze: acylace alfa-chymotrypsinu specifickými peptidovými substráty". Biochemie. 43 (3): 742–747. doi:10.1021 / bi030222k. PMID 14730979.
- ^ Lymn, RW; Taylor, EW. (1971). "Mechanismus hydrolýzy adenosintrifosfátu aktomyosinem". Biochemie. 10 (25): 4617–4624. doi:10.1021 / bi00801a004. PMID 4258719.
- ^ Holmes, KC; Angert, I; Kull, FG; Jahn, W; Schroder, RR. (2003). „Elektronová kryomikroskopie ukazuje, jak silná vazba myosinu na aktin uvolňuje nukleotid“. Příroda. 425 (6956): 423–427. Bibcode:2003 Natur.425..423H. doi:10.1038 / nature02005. PMID 14508495. S2CID 2686184.
- ^ Siemankowski, RF; Wiseman, MO; Bílá, HD. (1985). „Disociace ADP z dílčího fragmentu aktomyosinu 1 je dostatečně pomalá, aby omezila rychlost nezatíženého zkrácení ve svalech obratlovců“. Proc. Natl. Acad. Sci. USA. 82 (3): 658–662. Bibcode:1985PNAS ... 82..658S. doi:10.1073 / pnas.82.3.658. PMC 397104. PMID 3871943.
- ^ Bílá, HD; Belknap, B; Webb, MR. (1997). „Kinetika štěpení nukleosid trifosfátu a kroky uvolňování fosfátu přidruženým králičím skeletálním aktomyosinem, měřeno pomocí nové fluorescenční sondy pro fosfát“. Biochemie. 36 (39): 11828–11836. doi:10.1021 / bi970540h. PMID 9305974.
- ^ Tirosh, R; Nízká, WZ; Oplatka, A. (1990). „Translační pohyb aktinových vláken v přítomnosti těžkého meromyosinu a MgATP měřený Dopplerovým rozšířením rozptylu laserového světla“. Biochim. Biophys. Acta. 1037 (3): 274–280. doi:10.1016 / 0167-4838 (90) 90025-b. PMID 2178685.
- ^ Tirosh, R. (2006). „Balistické protony a mikrovlnné vodní roztoky (soliton) v bioenergetických transformacích“. Int. J. Mol. Sci. 7 (9): 320–345. doi:10.3390 / i7090320.
- ^ Muddana, Hari S .; Sengupta, Samudra; Mallouk, Thomas E .; et al. (28. ledna 2010). „Substrátová katalýza zvyšuje difúzi jednoho enzymu“. Journal of the American Chemical Society. 132 (7): 2110–1. doi:10.1021 / ja908773a. PMC 2832858. PMID 20108965.

- ^ Riedel, Clement; Gabizon, Ronen; Wilson, Christian A. M .; et al. (8. ledna 2015). „Teplo uvolněné během katalytického obratu zvyšuje difúzi enzymu“. Příroda. 517 (7533): 227–30. Bibcode:2015Natur.517..227R. doi:10.1038 / příroda14043. PMC 4363105. PMID 25487146. Shrnutí ležel – Příroda: Zprávy a pohledy (8. ledna 2015).
- ^ Rahman, SA; Cuesta, SM; Furnham, N; Holliday, GL; Thornton, JM (2014). „EC-BLAST: nástroj pro automatické vyhledávání a porovnávání enzymových reakcí“. Přírodní metody. 11 (2): 171–174. doi:10.1038 / nmeth.2803. PMC 4122987. PMID 24412978.
Další čtení
- Alan Fersht, Struktura a mechanismus ve vědě o bílkovinách: Průvodce enzymovou katalýzou a skládáním bílkovin. W. H. Freeman, 1998. ISBN 0-7167-3268-8
- Vyhrazené vydání Filozofické transakce B na kvantové katalýze v enzymech volně dostupné.[trvalý mrtvý odkaz ]
externí odkazy
 Média související s Enzymatická katalýza na Wikimedia Commons
Média související s Enzymatická katalýza na Wikimedia Commons