ATP syntáza - ATP synthase
Bylo navrženo, aby tento článek byl sloučeny s F-ATPáza. (Diskutujte) Navrhováno od února 2020. |
| ATP syntáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
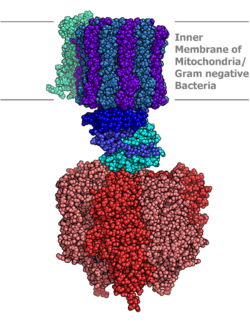 Molekulární model ATP syntázy stanoven pomocí Rentgenová krystalografie. Stator se zde nezobrazuje. | |||||||||
| Identifikátory | |||||||||
| EC číslo | 7.1.2.2 | ||||||||
| Číslo CAS | 9000-83-3 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
ATP syntáza je enzym který katalyzuje tvorbu molekuly akumulující energii adenosintrifosfát (ATP) pomocí adenosindifosfát (ADP) a anorganické fosfát (Stri). Celková reakce katalyzovaná ATP syntázou je:
- ADP + Pi + 3H+ven ⇌ ATP + H2O + 3H+v
Tvorba ATP z ADP a Pi je energeticky nepříznivý a normálně by postupoval opačným směrem. Aby byla tato reakce poháněna kupředu, syntéza ATP spojuje syntézu ATP během buněčné dýchání do elektrochemický gradient vytvořen rozdílem v proton (H+) koncentrace napříč vnitřní mitochondriální membrána v eukaryoty nebo plazmatická membrána v bakteriích. V době fotosyntéza v rostlinách je ATP syntetizován ATP syntázou pomocí protonového gradientu vytvořeného v tylakoidní lumen přes tylakoidní membránu a do chloroplastové stroma.
Eukaryotické ATP syntázy jsou F-ATPázy, běží "v opačném směru" pro ATPáza. Tento článek se zabývá hlavně tímto typem. F-ATPáza se skládá ze dvou hlavních podjednotek, FÓ a F1, který má mechanismus rotačního motoru umožňující produkci ATP.[1][2] Kvůli své rotující podjednotce je ATP syntáza a molekulární stroj.
Nomenklatura
F1 zlomek odvozuje svůj název od výrazu „Frakce 1“ a FÓ (psaný jako dolní index "o", ne "nula") odvozuje svůj název od toho, že je vazbou zlomek pro oligomycin, typ přirozeně odvozeného antibiotika, které je schopné inhibovat FÓ jednotka ATP syntázy.[3][4] Tyto funkční oblasti se skládají z různých proteinových podjednotek - viz tabulky. Tento enzym se používá při syntéze ATP prostřednictvím aerobního dýchání.
Struktura a funkce


Nachází se v tylakoidní membrána a vnitřní mitochondriální membrána, ATP syntáza se skládá ze dvou oblastí FÓ a F1. FÓ způsobí rotaci F1 a je vyroben z c-kruhu a podjednotek a, dva b, F6. F1 je tvořen z podjednotek α, β, γ, δ. F1 má ve vodě rozpustnou část, která může hydrolyzovat ATP. FÓ na druhé straně má hlavně hydrofobní oblasti. FÓ F1 vytváří cestu pro pohyb protonů přes membránu.[7]
F1 kraj
F1 část ATP syntázy je hydrofilní a je odpovědný za hydrolýzu ATP. F1 jednotka vyčnívá do prostoru mitochondriální matice. Podjednotky α a β tvoří hexamer se 6 vazebnými místy. Tři z nich jsou katalyticky neaktivní a vážou ADP.
Další tři podjednotky katalyzují syntézu ATP. Druhý F1 podjednotky γ, δ, ε jsou součástí mechanismu rotačního motoru (rotor / náprava). γ podjednotka umožňuje, aby β procházel konformačními změnami (tj. uzavřenými, napůl otevřenými a otevřenými stavy), které umožňují navázání a uvolnění ATP po syntéze. F1 částice je velká a lze ji vidět v transmisním elektronovém mikroskopu negativním barvením.[8] Jedná se o částice o průměru 9 nm, které pepřují vnitřní mitochondriální membránu.
| Podjednotka | Lidský gen | Poznámka |
|---|---|---|
| alfa | ATP5A1, ATPAF2 | |
| beta | ATP5B, ATPAF1, C16orf7 | |
| gama | ATP5C1 | |
| delta | ATP5D | Mitochondriální „delta“ je bakteriální / chloroplastický epsilon. |
| epsilon | ATP5E | Unikátní pro mitochondrie. |
| OSCP | ATP5O | V bakteriální a chloroplastické verzi se nazývá „delta“. |
FÓ kraj

FÓ je voda nerozpustný protein s osmi podjednotkami a transmembránovým prstencem. Prsten má tetramer tvar s a smyčka šroubovice spirální protein, který prochází konformačními změnami, když protonuje a deprotonuje, tlačí sousední podjednotky do rotace, což způsobuje otáčení FÓ což pak také ovlivňuje konformaci F1, což má za následek přepnutí stavů podjednotek alfa a beta. FÓ oblast ATP syntázy je protonový pór, který je zality v mitochondriální membráně. Skládá se ze tří hlavních podjednotek, a, b a c. Šest c podjednotek tvoří rotorový kruh a podjednotka b tvoří stopku spojující s F1 OSCP, který zabraňuje rotaci αβ hexameru. Podjednotka a spojuje b s c kroužkem.[11] Lidé mají šest dalších podjednotek, d, E, F, G, F6, a 8 (nebo A6L). Tato část enzymu je umístěna ve vnitřní mitochondriální membráně a spojuje translokaci protonů s rotací, což způsobuje syntézu ATP v F1 kraj.
U eukaryot, mitochondriální FÓ tvoří dimery ohýbající členy. Tyto dimery se samy uspořádají do dlouhých řad na konci cristae, možná první krok vzniku cristae.[12] Atomový model pro dimerní kvasinky FÓ oblast byla stanovena kryo-EM při celkovém rozlišení 3,6 Á.[13]
| Podjednotka | Lidský gen |
|---|---|
| A | MT-ATP6, MT-ATP8 |
| b | ATP5F1 |
| C | ATP5G1, ATP5G2, ATP5G3 |
Vazební model


V letech 1960 až 1970, Paul Boyer, a UCLA Profesor, vyvinul teorii mechanismu vazebné změny nebo klopného obvodu, který předpokládal, že syntéza ATP závisí na konformační změně ATP syntázy generované rotací gamma podjednotky. Výzkumná skupina John E. Walker, pak na MRC Laboratoř molekulární biologie v Cambridge, krystalizoval F1 katalytická doména ATP syntázy. Struktura, v době největší známé asymetrické proteinové struktury, naznačovala, že Boyerův model rotační katalýzy byl v podstatě správný. Pro objasnění toho sdíleli Boyer a Walker polovinu roku 1997 Nobelova cena za chemii.
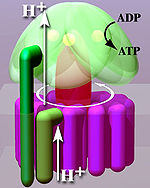
Krystalová struktura F1 ukázaly střídavé alfa a beta podjednotky (3 z každého), uspořádané jako oranžové segmenty kolem rotující asymetrické gama podjednotky. Podle současného modelu syntézy ATP (známého jako střídavý katalytický model) transmembránový potenciál vytvořený (H +) protonovými kationty dodávanými elektronovým transportním řetězcem pohání (H +) protonové kationty z mezimembránového prostoru přes membránu přes FÓ oblast ATP syntázy. Část FÓ (prsten z c-podjednotky ) otáčí se když protony procházejí membránou. The c-kroužek je pevně spojen s asymetrickou centrální stopkou (skládající se převážně z gama podjednotky), což způsobuje její rotaci v alfa3beta3 vypnuto1 což způsobí, že 3 vazebná místa pro katalytické nukleotidy procházejí řadou konformačních změn, které vedou k syntéze ATP. Major F1 podjednotkám je zabráněno v rotaci v soucitu s centrálním stonkovým rotorem obvodovým stonkem, který se připojuje k alfa3beta3 do nerotující části FÓ. Struktura intaktní ATP syntázy je v současné době známá při nízkém rozlišení od elektronová kryomikroskopie (kryo-EM) studie komplexu. Kryo-EM model ATP syntázy naznačuje, že periferní stonek je flexibilní struktura, která obklopuje komplex, když se připojuje k F1 do FÓ. Za správných podmínek může být enzymová reakce provedena také obráceně, s hydrolýzou ATP čerpání protonů přes membránu.
Mechanismus změny vazby zahrnuje aktivní místo cyklování β podjednotky mezi třemi stavy.[14] Ve "volném" stavu vstupují ADP a fosfát do aktivního místa; v sousedním diagramu je to zobrazeno růžově. Enzym poté prochází změnou tvaru a nutí tyto molekuly k sobě, přičemž aktivní místo ve výsledném „těsném“ stavu (zobrazeno červeně) váže nově produkovanou molekulu ATP s velmi vysokou afinita. Nakonec se aktivní místo cykluje zpět do otevřeného stavu (oranžové), uvolňuje ATP a váže více ADP a fosfátů, připraveno na další cyklus výroby ATP.[15]
Fyziologická role
Stejně jako ostatní enzymy, aktivita F1FÓ ATP syntáza je reverzibilní. Dostatečně velké množství ATP způsobí, že vytvoří transmembránu proton spád, to se používá k fermentaci bakterií, které nemají elektronový transportní řetězec, ale spíše hydrolyzují ATP, aby vytvořily protonový gradient, který používají k řízení bičíky a transport živin do buňky.
V dýchání bakterie za fyziologických podmínek běží ATP syntáza obecně opačným směrem a vytváří ATP při používání hnací síla protonu vytvořil elektronový transportní řetězec jako zdroj energie. Celkový proces vytváření energie tímto způsobem se nazývá oxidační fosforylace Stejný proces probíhá v mitochondrie, kde je ATP syntáza umístěna ve vnitřní mitochondriální membráně a F1-část projektů do mitochondriální matice. Spotřeba ATP ATP-syntázou pumpuje protonové kationty do matrice.
Vývoj
The vývoj ATP syntázy se považuje za modulární, čímž se spojily dvě funkčně nezávislé podjednotky a získaly nové funkce.[16][17] Zdá se, že k této asociaci došlo na počátku evoluční historie, protože ve všech královstvích života je v podstatě stejná struktura a aktivita enzymů ATP syntázy.[16] F-ATP syntáza vykazuje vysokou funkční a mechanickou podobnost s V-ATPáza.[18] Avšak zatímco F-ATP syntáza generuje ATP využitím protonového gradientu, V-ATPáza generuje protonový gradient na úkor ATP a generuje hodnoty pH již od 1.[19]
F1 Oblast také vykazuje významnou podobnost s hexamerickou DNA helikázy (zejména Rho faktor ) a celá oblast enzymu vykazuje určitou podobnost s H+
- napájen T3SS nebo bičíkový motor komplexy.[18][20][21] Α3β3 hexamer z F1 oblast vykazuje významnou strukturní podobnost s hexamerickými DNA helikázami; oba tvoří prstenec s trojnásobnou rotační symetrií se středním pórem. Oba mají role závislé na relativní rotaci makromolekuly v póru; DNA helikázy používají spirálovitý tvar DNA k řízení jejich pohybu podél molekuly DNA a k detekci nadšroubovice, zatímco α3β3 hexamer používá konformační změny prostřednictvím rotace y podjednotky k řízení enzymatické reakce.[22]
The H+
motor F.Ó částice vykazuje velkou funkční podobnost s částicemi H+
motory, které pohánějí bičíky.[18] Oba mají prsten mnoha malých alfa-šroubovicových proteinů, které rotují vzhledem k blízkým stacionárním proteinům pomocí a H+
potenciální gradient jako zdroj energie. Tento odkaz je však nejasný, protože celková struktura bičíkových motorů je mnohem složitější než u FÓ částice a kruh s asi 30 rotujícími proteiny je mnohem větší než 10, 11 nebo 14 spirálových proteinů v FÓ komplex. Novější strukturální údaje však ukazují, že prsten a stopka jsou strukturně podobné F1 částice.[21]

Teorie modulární evoluce původu ATP syntázy naznačuje, že dvě podjednotky s nezávislou funkcí, DNA helikáza s aktivitou ATPázy a H+
motor, byli schopni se vázat, a rotace motoru poháněla aktivitu ATPázy helikázy v opačném směru.[16][22] Tento komplex poté vyvinul větší účinnost a nakonec se vyvinul do dnešních složitých syntáz ATP. Alternativně DNA helikáza /H+
motorový komplex možná měl H+
aktivita pumpy s aktivitou ATPázy helikázy pohánějící H+
motor v opačném směru.[16] To se mohlo vyvinout k provedení reverzní reakce a působení jako ATP syntáza.[17][23][24]
Inhibitory
Byly objeveny různé přírodní a syntetické inhibitory ATP syntázy.[25] Ty byly použity ke zkoumání struktury a mechanismu ATP syntázy. Některé mohou mít terapeutické použití. Existuje několik tříd inhibitorů ATP syntázy, včetně inhibitorů peptidů, polyfenolických fytochemikálií, polyketidů, organických sloučenin cínu, polyenových derivátů α-pyronu, kationtových inhibitorů, analogů substrátu, modifikátorů aminokyselin a dalších různých chemikálií.[25] Mezi nejčastěji používané inhibitory ATP syntázy patří oligomycin a DCCD.
V různých organismech
Bakterie
E-coli ATP syntáza je nejjednodušší známá forma ATP syntázy s 8 různými typy podjednotek.[11]
Bakteriální F-ATPázy mohou příležitostně působit obráceně a přeměňovat je na ATPázu.[26] Některé bakterie nemají žádnou F-ATPázu, používají ATPázu typu A / V obousměrně.[9]
Droždí
Kvasinková ATP syntáza je jednou z nejlépe studovaných eukaryotických ATP syntáz; a pět F.1, osm F.Ó Bylo identifikováno sedm podjednotek a sedm asociovaných proteinů.[7] Většina z těchto proteinů má homology v jiných eukaryotech.[27][28][29][30]
Rostlina
V rostlinách je ATP syntáza také přítomna v chloroplasty (CF1FÓ-ATP syntáza). Enzym je integrován do thylakoid membrána; CF1- část se drží stroma, kde temné reakce fotosyntézy (nazývané také reakce nezávislé na světle nebo Calvinův cyklus ) a probíhá syntéza ATP. Celková struktura a katalytický mechanismus chloroplastové ATP syntázy jsou téměř stejné jako u bakteriálního enzymu. V chloroplastech však hnací síla protonu je generován nikoli dýchacím elektronovým transportním řetězcem, ale primárními fotosyntetickými proteiny. Syntáza má 40-aa inzert v gama podjednotce, aby inhibovala nehospodárnou aktivitu při setmění.[31]
Savec
ATP syntáza izolovaná z hovězího dobytka (Bos taurus) srdeční mitochondrie je z hlediska biochemie a struktury nejlépe charakterizovanou ATP syntázou. Hovězí srdce se používá jako zdroj enzymu kvůli vysoké koncentraci mitochondrií v srdečním svalu. Jejich geny mají blízkou homologii s lidskými ATP syntázami.[32][33][34]
Lidské geny, které kódují složky ATP syntáz:
- ATP5A1
- ATP5B
- ATP5C1, ATP5D, ATP5E, ATP5F1, ATP5G1, ATP5G2, ATP5G3, ATP5H, ATP5I, ATP5J, ATP5J2, ATP5L, ATP5O
- MT-ATP6, MT-ATP8
Ostatní eukaryoty
Eukaryoty patřící do některých odlišných linií mají velmi speciální organizace ATP syntázy. A Euglenozoa ATP syntáza tvoří dimer s F ve tvaru bumerangu1 hlava jako ostatní mitochondriální ATP syntázy, ale FÓ subkomplex má mnoho jedinečných podjednotek. Využívá to kardiolipin. Inhibiční IF1 také se váže odlišně, způsobem sdíleným s Trypanosomatida.[35]
Archaea
Archaea obecně nemá F-ATPázu. Místo toho syntetizují ATP pomocí A-ATPázy / syntázy, rotačního stroje, který je strukturálně podobný V-ATPáza ale hlavně funguje jako ATP syntáza.[26] Podobně jako bakterie F-ATPáza se předpokládá, že také funguje jako ATPáza.[9]
Viz také
- ATP10 protein potřebné pro montáž FÓ sektor mitochondriálního komplexu ATPázy.
- Chloroplast
- Řetězec přenosu elektronů
- Flavoprotein
- Mitochondrie
- Oxidační fosforylace
- P-ATPáza
- Protonová pumpa
- Rotující lokomoce v živých systémech
- Transmembránová ATPáza
- V-ATPáza
Reference
- ^ Okuno D, Iino R, Noji H (duben 2011). "Rotace a struktura FOF1-ATP syntázy". Journal of Biochemistry. 149 (6): 655–64. doi:10.1093 / jb / mvr049. PMID 21524994.
- ^ Junge W, Nelson N (červen 2015). Msgstr "ATP syntáza". Roční přehled biochemie. 84: 631–57. doi:10.1146 / annurev-biochem-060614-034124. PMID 25839341.
- ^ Kagawa Y, Racker E (květen 1966). „Částečné rozlišení enzymů katalyzujících oxidativní fosforylaci. 8. Vlastnosti faktoru, který uděluje citlivost na oligomycin mitochondriální adenosintrifosfatáze“. The Journal of Biological Chemistry. 241 (10): 2461–6. PMID 4223640.
- ^ Mccarty RE (listopad 1992). „Pohled rostlinného biochemika na H+
-ATPázy a ATP syntázy ". The Journal of Experimental Biology. 172 (Pt 1): 431–441. PMID 9874753. - ^ PDB: 5ARA; Zhou A, Rohou A, Schep DG, Bason JV, Montgomery MG, Walker JE, Grigorieff N, Rubinstein JL (říjen 2015). "Struktura a konformační stavy hovězí mitochondriální ATP syntázy pomocí kryo-EM". eLife. 4: e10180. doi:10.7554 / eLife.10180. PMC 4718723. PMID 26439008.
- ^ Goodsell D (prosinec 2005). „ATP Synthase“. Molekula měsíce. doi:10.2210 / rcsb_pdb / mom_2005_12.
- ^ A b Velours J, Paumard P, Soubannier V, Spannagel C, Vaillier J, Arselin G, Graves PV (květen 2000). „Organizace kvasinkové ATP syntázy F (0): studie založená na cysteinových mutantech, thiolové modifikaci a síťovacích činidlech“. Biochimica et Biophysica Acta (BBA) - bioenergetika. 1458 (2–3): 443–56. doi:10.1016 / S0005-2728 (00) 00093-1. PMID 10838057.
- ^ Fernandez Moran H, Oda T, Blair PV, Green DE (červenec 1964). "Makromolekulární opakující se jednotka mitochondriální struktury a funkce. Korelované elektronové mikroskopické a biochemické studie izolovaných mitochondrií a submochondriálních částic hovězího srdečního svalu". The Journal of Cell Biology. 22 (1): 63–100. doi:10.1083 / jcb.22.1.63. PMC 2106494. PMID 14195622.
- ^ A b C Stewart AG, Laming EM, Sobti M, Stock D (duben 2014). "Rotační ATPázy - dynamické molekulární stroje". Aktuální názor na strukturní biologii. 25: 40–8. doi:10.1016 / j.sbi.2013.11.013. PMID 24878343.
- ^ PDB: 1VZS; Carbajo RJ, Silvester JA, Runswick MJ, Walker JE, Neuhaus D (2004). "Struktura řešení podjednotky F (6) z oblasti periferní stopky ATP syntázy z mitochondrií bovinního srdce". Journal of Molecular Biology. 342 (2): 593–603. doi:10.1016 / j.jmb.2004.07.013. PMID 15327958.
- ^ A b Ahmad Z, Okafor F, Laughlin TF (2011). „Role nabitých reziduí v katalytických lokalitách ATP syntázy Escherichia coli“. Journal of Amino Acids. 2011: 1–12. doi:10.4061/2011/785741. PMC 3268026. PMID 22312470.
- ^ Blum TB, Hahn A, Meier T, Davies KM, Kühlbrandt W (březen 2019). „Dimery mitochondriální ATP syntázy indukují zakřivení membrány a samy se skládají do řádků“. Sborník Národní akademie věd Spojených států amerických. 116 (10): 4250–4255. doi:10.1073 / pnas.1816556116. PMC 6410833. PMID 30760595.
- ^ Guo H, Bueler SA, Rubinstein JL (listopad 2017). "Atomový model pro dimerní FÓ oblast mitochondriální ATP syntázy ". Věda. 358 (6365): 936–40. Bibcode:2017Sci ... 358..936G. doi:10.1126 / science.aao4815. PMC 6402782. PMID 29074581.
- ^ Gresser MJ, Myers JA, Boyer PD (říjen 1982). "Katalytická kooperativita mitochondriálního F z hovězího srdce."1 adenosintrifosfatáza. Korelace počáteční rychlosti, vázaného meziproduktu a měření výměny kyslíku se střídavým modelem se třemi místy ". The Journal of Biological Chemistry. 257 (20): 12030–8. PMID 6214554.
- ^ Nakamoto RK, Baylis Scanlon JA, Al-Shawi MK (srpen 2008). „Rotační mechanismus ATP syntázy“. Archivy biochemie a biofyziky. 476 (1): 43–50. doi:10.1016 / j.abb.2008.05.004. PMC 2581510. PMID 18515057.
- ^ A b C d Doering C, Ermentrout B, Oster G (prosinec 1995). "Rotační DNA motory". Biofyzikální deník. 69 (6): 2256–67. Bibcode:1995 BpJ .... 69,2256D. doi:10.1016 / S0006-3495 (95) 80096-2. PMC 1236464. PMID 8599633.
- ^ A b Crofts A. „Přednáška 10: ATP syntáza“. Vědy o životě na University of Illinois v Urbana – Champaign.
- ^ A b C „ATP Synthase“. Databáze InterPro.
- ^ Beyenbach KW, Wieczorek H (únor 2006). „H-ATPáza typu V: molekulární struktura a funkce, fyziologické role a regulace“. The Journal of Experimental Biology. 209 (Pt 4): 577–89. doi:10.1242 / jeb.02014. PMID 16449553.
- ^ Skordalakes E, Berger JM (červenec 2003). "Struktura terminátoru transkripce Rho: mechanismus rozpoznávání mRNA a načítání helikázy". Buňka. 114 (1): 135–46. doi:10.1016 / S0092-8674 (03) 00512-9. PMID 12859904. S2CID 5765103.
- ^ A b Imada K, Minamino T, Uchida Y, Kinoshita M, Namba K (březen 2016). „Pohled na export bičíku typu III odhalený složitou strukturou ATPázy typu III a jejím regulátorem“. Sborník Národní akademie věd Spojených států amerických. 113 (13): 3633–8. Bibcode:2016PNAS..113,3633I. doi:10.1073 / pnas.1524025113. PMC 4822572. PMID 26984495.
- ^ A b Martinez LO, Jacquet S, Esteve JP, Rolland C, Cabezón E, Champagne E a kol. (Leden 2003). „Ektopický beta řetězec ATP syntázy je receptorem apolipoproteinu A-I při jaterní HDL endocytóze“. Příroda. 421 (6918): 75–9. Bibcode:2003 Natur.421 ... 75 mil. doi:10.1038 / nature01250. PMID 12511957. S2CID 4333137.
- ^ Cross RL, Taiz L (leden 1990). „Duplikace genů jako prostředek ke změně poměrů H + / ATP během vývoje FÓF1 ATPázy a syntázy ". FEBS Dopisy. 259 (2): 227–9. doi:10.1016 / 0014-5793 (90) 80014-a. PMID 2136729. S2CID 32559858.
- ^ Cross RL, Müller V (říjen 2004). „Vývoj A-, F- a V-typu ATP syntáz a ATPáz: obrácení funkce a změny v H + / ATP vazebném poměru“. FEBS Dopisy. 576 (1–2): 1–4. doi:10.1016 / j.febslet.2004.08.065. PMID 15473999. S2CID 25800744.
- ^ A b Hong S, Pedersen PL (prosinec 2008). „ATP syntáza a působení inhibitorů využívaných ke studiu jejích rolí v oblasti lidského zdraví, nemocí a dalších vědeckých oblastí“. Recenze mikrobiologie a molekulární biologie. 72 (4): 590–641, obsah. doi:10.1128 / MMBR.00016-08. PMC 2593570. PMID 19052322.
- ^ A b Kühlbrandt W, Davies KM (leden 2016). „Rotary ATPases: New Twist to an Ancient Machine“. Trendy v biochemických vědách. 41 (1): 106–116. doi:10.1016 / j.tibs.2015.10.006. PMID 26671611.
- ^ Devenish RJ, Prescott M, Roucou X, Nagley P (květen 2000). „Pohledy na sestavení a fungování ATP syntázy pomocí molekulárně genetické manipulace podjednotek komplexu kvasinkových mitochondriálních enzymů“. Biochimica et Biophysica Acta (BBA) - bioenergetika. 1458 (2–3): 428–42. doi:10.1016 / S0005-2728 (00) 00092-X. PMID 10838056.
- ^ Kabaleeswaran V, Puri N, Walker JE, Leslie AG, Mueller DM (listopad 2006). „Nové rysy rotačního katalytického mechanismu odhalené ve struktuře kvasinek F1 ATPáza ". Časopis EMBO. 25 (22): 5433–42. doi:10.1038 / sj.emboj.7601410. PMC 1636620. PMID 17082766.
- ^ Stock D, Leslie AG, Walker JE (listopad 1999). "Molekulární architektura rotačního motoru v ATP syntáze". Věda. 286 (5445): 1700–5. doi:10.1126 / science.286.5445.1700. PMID 10576729.
- ^ Liu S, Charlesworth TJ, Bason JV, Montgomery MG, Harbor ME, Fearnley IM, Walker JE (květen 2015). "Čištění a charakterizace komplexů ATP syntázy z mitochondrií čtyř druhů hub". Biochemical Journal. 468 (1): 167–175. doi:10.1042 / BJ20150197. PMC 4422255. PMID 25759169.
- ^ Hahn A, Vonck J, Mills DJ, Meier T, Kühlbrandt W (květen 2018). "Struktura, mechanismus a regulace chloroplastové ATP syntázy". Věda. 360 (6389): eaat4318. doi:10.1126 / science.aat4318. PMC 7116070. PMID 29748256.
- ^ Abrahams JP, Leslie AG, Lutter R, Walker JE (srpen 1994). „Struktura při rozlišení 2,8 A F1-ATPázy z mitochondrií bovinního srdce“. Příroda. 370 (6491): 621–8. Bibcode:1994 Natur.370..621A. doi:10.1038 / 370621a0. PMID 8065448. S2CID 4275221.
- ^ Gibbons C, Montgomery MG, Leslie AG, Walker JE (listopad 2000). "Struktura centrálního stonku v bovinní F (1) -ATPáze v rozlišení 2,4 A". Přírodní strukturní biologie. 7 (11): 1055–61. doi:10.1038/80981. PMID 11062563. S2CID 23229994.
- ^ Menz RI, Walker JE, Leslie AG (srpen 2001). "Struktura hovězí mitochondriální F (1) -ATPázy s nukleotidem vázaným na všechna tři katalytická místa: důsledky pro mechanismus rotační katalýzy". Buňka. 106 (3): 331–41. doi:10.1016 / s0092-8674 (01) 00452-4. PMID 11509182. S2CID 1266814.
- ^ Mühleip A, McComas SE, Amunts A (listopad 2019). "Struktura mitochondriální ATP syntázy s navázaným nativním kardiolipinem". eLife. 8: e51179. doi:10,7554 / eLife.51179. PMC 6930080. PMID 31738165. Shrnutí ležel – eLife.
Další čtení
- Nick Lane: Zásadní otázka: Energie, evoluce a počátky složitého života, Ww Norton, 2015-07-20, ISBN 978-0393088816 (Link points to Figure 10 showing model of ATP synthase)
externí odkazy
- Boris A. Feniouk: „ATP syntáza - skvělý molekulární stroj“
- Dobře ilustrované Přednáška ATP syntázy Antony Crofts z University of Illinois v Urbana – Champaign.
- Proton a sodík translokující ATPázy typu F, typu V a typu A v databázi OPM
- Nobelova cena za chemii 1997 Paul D. Boyer a John E. Walker za enzymatický mechanismus syntézy ATP; a Jens C. Skou za objev iontového transportního enzymu, Na+
, K.+
-ATPase. - Harvardská multimediální produkční stránka - videa - Animace syntézy ATP
- David Goodsell: "ATP Synthase - Molekula měsíce"