Vývoj - Evolution

Vývoj je změna v dědičný charakteristiky biologické populace po sobě jdoucích generace.[1][2] Tyto vlastnosti jsou výrazy z geny které se předávají z rodiče na potomka během reprodukce. V důsledku toho obvykle v rámci dané populace existují různé charakteristiky mutace, genetická rekombinace a další zdroje genetická variace.[3] Evoluce nastává, když evoluční procesy jako přírodní výběr (počítaje v to sexuální výběr ) a genetický drift jednat podle této variace, což vede k tomu, že se určité charakteristiky stanou v populaci běžnějšími nebo vzácnějšími.[4] Právě tento proces evoluce dal vzniknout biologická rozmanitost na všech úrovních biologická organizace, včetně úrovní druh, individuální organismy a molekuly.[5][6]
The vědecká teorie evoluce přirozeným výběrem byla koncipována nezávisle na Charles Darwin a Alfred Russel Wallace v polovině 19. století a byl podrobně popsán v Darwinově knize O původu druhů.[7] Evoluce přirozeným výběrem byla poprvé demonstrována pozorováním, že se často produkuje více potomků, než může přežít. Poté následují tři pozorovatelné fakta o živých organismech: (1) vlastnosti se u jednotlivců liší podle morfologie, fyziologie a chování (fenotypová variace ), (2) různé vlastnosti udělují různé míry přežití a reprodukce (rozdíl zdatnost ) a (3) vlastnosti lze předávat z generace na generaci (dědičnost fitness).[8] V následujících generacích je tedy pravděpodobnější, že členové populace budou nahrazeni potomci rodičů s příznivými vlastnostmi, které jim umožnily přežít a rozmnožovat se ve svých příslušných prostředí. Na počátku 20. století jiné konkurenční myšlenky evoluce jako mutace a ortogeneze byly vyvrácen jako moderní syntéza smířený Darwinovská evoluce s klasická genetika, který stanovil adaptivní evoluce jako způsobené přirozeným výběrem, který působí Mendelian genetická variace.[9]
Všechno život na Zemi sdílí a poslední univerzální společný předek (LUCA)[10][11][12] který žil přibližně před 3,5–3,8 miliardami let.[13] The fosilní záznam zahrnuje postup od počátku biogenní grafit,[14] na mikrobiální mat fosilie,[15][16][17] zkamenělé mnohobuněčné organismy. Stávající vzorce biologické rozmanitosti byly formovány opakovanými formacemi nových druhů (speciace ), změny v rámci druhů (anageneze ) a ztráta druhů (zánik ) skrze evoluční historie života na Zemi.[18] Morfologické a biochemické vlastnosti jsou si více podobné u druhů, které sdílejí více nedávný společný předek a lze jej použít k rekonstrukci fylogenetické stromy.[19][20]
Evoluční biologové pokračovali ve studiu různých aspektů evoluce formováním a testováním hypotézy stejně jako konstrukční teorie založené na důkaz z pole nebo laboratoře a na datech generovaných metodami matematická a teoretická biologie. Jejich objevy ovlivnily nejen vývoj biologie ale mnoho dalších vědeckých a průmyslových oborů, včetně zemědělství, lék a počítačová věda.[21]
Historie evolučního myšlení




Klasické časy
Návrh, že jeden typ organismu by mohl sestoupit z jiného typu, se vrací k některým z prvních předsokratovské řecký filozofové, jako Anaximander a Empedokles.[23] Takové návrhy přežily až do římských časů. The básník a filozof Lucretius následoval Empedokla v jeho mistrovském díle De rerum natura (O povaze věcí).[24][25]
Středověký
Na rozdíl od nich materialistický pohledy, Aristotelismus považoval všechny přirozené věci za aktualizace stálých přírodních možností, známých jako formuláře.[26][27] To bylo součástí středověku teleologický porozumění Příroda ve kterém všechny věci mají hrát zamýšlenou roli v a božský vesmírný objednat. Variace této myšlenky se staly standardním chápáním Středověk a byly integrovány do křesťan Učení, ale Aristoteles nepožadoval, aby skutečné typy organismů vždy odpovídaly jeden za druhým přesným metafyzickým formám, a konkrétně uvedl příklady toho, jak mohou nové druhy živých věcí vzniknout.[28]
Předdarwinovský
V 17. století nový metoda z moderní věda odmítl aristotelský přístup. Hledalo vysvětlení přírodních jevů, pokud jde o fyzikální zákony které byly stejné pro všechny viditelné věci a které nevyžadovaly existenci jakýchkoli pevných přírodních kategorií nebo božského kosmického řádu. Tento nový přístup se však pomalu zakořenil v biologických vědách, poslední bašte konceptu stálých přírodních typů. John Ray použil jeden z dříve obecnějších pojmů pro stálé přírodní typy, „druhy“, na druhy rostlin a zvířat, ale každý druh živého tvora striktně identifikoval jako druh a navrhl, aby každý druh mohl být definován rysy, které si udržovaly generaci po generaci.[29] Biologická klasifikace zavedená Carl Linné v roce 1735 výslovně uznal hierarchickou povahu druhových vztahů, ale přesto považoval druh za ustálený podle božského plánu.[30]
jiný přírodovědci této doby spekulovalo o evoluční změně druhů v průběhu času podle přírodních zákonů. V roce 1751 Pierre Louis Maupertuis psal o přírodních modifikacích vyskytujících se během reprodukce a hromadících se po mnoho generací za účelem produkce nových druhů.[31] Georges-Louis Leclerc, hrabě de Buffon navrhl, že by druhy mohly degenerovat do různých organismů, a Erazmus Darwin navrhl, aby všechna teplokrevná zvířata mohla pocházet z jediného mikroorganismu (nebo „vlákna“).[32] První plnohodnotné evoluční schéma bylo Jean-Baptiste Lamarck "transmutační" teorie z roku 1809,[33] který předpokládal spontánní generace neustále produkují jednoduché formy života, které vyvinuly větší složitost v paralelních liniích s inherentní progresivní tendencí, a předpokládají, že na místní úrovni se tyto linie přizpůsobily prostředí tím, že zdědily změny způsobené jejich používáním nebo nepoužíváním u rodičů.[34][35] (Druhý proces byl později nazýván Lamarckismus.)[34][36][37][38] Tyto myšlenky byli odsouzeni zavedenými přírodovědci jako spekulace postrádající empirickou podporu. Zejména, Georges Cuvier trval na tom, že druhy spolu nesouvisí a nejsou pevně stanoveny, přičemž jejich podobnosti odrážejí božský design pro funkční potřeby. Mezitím Ray vytvořil myšlenky benevolentního designu William Paley do Přirozená teologie nebo důkazy o existenci a vlastnostech božstva (1802), který navrhl složité úpravy jako důkaz božského designu a který obdivoval Charles Darwin.[39][40][41]
Darwinovská revoluce
Zásadní zlom od konceptu konstantních typologických tříd nebo typů v biologii přišel s evoluční teorií prostřednictvím přirozeného výběru, kterou formuloval Charles Darwin z hlediska variabilních populací. Darwin použil výraz „sestup s úpravou„spíše než„ vývoj “.[42] Částečně ovlivněno Esej o principu populace (1798) od Thomas Robert Malthus Darwin poznamenal, že populační růst povede k „boji o existenci“, ve kterém budou převládat příznivé variace, zatímco ostatní zahynou. V každé generaci mnoho potomků nedokáže přežít do věku reprodukce kvůli omezeným zdrojům. To by mohlo vysvětlit rozmanitost rostlin a zvířat od společného původu prostřednictvím působení přírodních zákonů stejným způsobem pro všechny typy organismů.[43][44][45][46] Darwin rozvinul svou teorii „přirozeného výběru“ od roku 1838 a později na toto téma psal „velkou knihu“ Alfred Russel Wallace poslal mu verzi prakticky stejné teorie v roce 1858. Jejich samostatné papíry byly předloženy společně na zasedání 1858 Linnean Society of London.[47] Na konci roku 1859 vyšlo Darwinovo vydání jeho „abstraktu“ jako O původu druhů vysvětlil přirozený výběr podrobně a způsobem, který vedl ke stále širšímu přijetí Darwinovy koncepty evoluce na úkor alternativní teorie. Thomas Henry Huxley aplikoval Darwinovy myšlenky na člověka pomocí paleontologie a srovnávací anatomie poskytnout silné důkazy o tom, že lidé a lidoopi sdíleli společný původ. Některé z toho byly vyrušeny, protože z toho vyplývalo, že lidé v USA nemají zvláštní místo vesmír.[48]
Pangenesis a dědičnost
Mechanismy reprodukční dědičnosti a původ nových vlastností zůstaly záhadou. Za tímto účelem rozvinul Darwin svou prozatímní teorii pangenesis.[49] V roce 1865 Gregor Mendel uvedl, že vlastnosti byly zděděny předvídatelným způsobem prostřednictvím nezávislý sortiment a segregace prvků (později známé jako geny). Mendelovy zákony dědičnosti nakonec nahradily většinu Darwinovy teorie pangeneze.[50] August Weismann udělal důležitý rozdíl mezi zárodečné buňky které vedou k gamety (jako spermie a vaječné buňky ) a somatické buňky těla, což ukazuje, že dědičnost prochází pouze zárodečnou linií. Hugo de Vries spojil Darwinovu teorii pangeneze s Weismannovým rozlišením zárodečných / soma buněk a navrhl, aby Darwinovy pangeny byly soustředěny v buněčné jádro a když se vyjádřili, mohli se přestěhovat do cytoplazma změnit buňka struktura. De Vries byl také jedním z výzkumníků, kteří Mendelovu práci dobře poznali, protože věřil, že Mendelianovy rysy odpovídají přenosu dědičných variací podél zárodečné linie.[51] Aby vysvětlil, jak vznikají nové varianty, vyvinul se de Vries teorie mutací což vedlo k dočasné roztržce mezi těmi, kteří akceptovali darwinovskou evoluci, a biometriky, kteří se spojili s de Vriesem.[35][52][53] Ve třicátých letech minulého století průkopníci v oblasti populační genetika, jako Ronald Fisher, Sewall Wright a J. B. S. Haldane položil základy evoluce na robustní statistickou filozofii. Falešný rozpor mezi Darwinovou teorií, genetickými mutacemi a Mendelovo dědictví byl tedy smířen.[54]
„Moderní syntéza“
Ve 20. a 30. letech 20. století tzv moderní syntéza propojil přirozený výběr a populační genetiku založenou na mendelovském dědictví do jednotné teorie, která se obecně vztahovala na jakékoli odvětví biologie. Moderní syntéza vysvětlila vzorce pozorované napříč druhy v populacích fosilní přechody v paleontologii a komplexní buněčné mechanismy v vývojová biologie.[35][55] Zveřejnění struktury DNA podle James Watson a Francis Crick s příspěvkem Rosalind Franklin v roce 1953 prokázal fyzický mechanismus dědičnosti.[56] Molekulární biologie lepší porozumění vztahu mezi genotyp a fenotyp. Pokroky byly provedeny také ve fylogenetice systematičnost, mapování přechodu znaků do srovnávacího a testovatelného rámce zveřejněním a používáním evoluční stromy.[57][58] V roce 1973 evoluční biolog Theodosius Dobžanský napsal to "nic v biologii nemá smysl, kromě evolučního světla „, protože to přineslo na světlo vztahy souvislostí toho, co se nejprve zdálo nesouvislé skutečnosti v přírodní historii vysvětlující soubor znalostí, který popisuje a předpovídá mnoho pozorovatelných faktů o životě na této planetě.[59]
Další syntézy
Od té doby byla moderní syntéza dále rozšířena o vysvětlení biologických jevů v celé a integrační škále biologická hierarchie, od genů k druhům.[60] Jedno rozšíření, známé jako evoluční vývojová biologie a neformálně nazývané „evo-devo“, zdůrazňuje, jak změny mezi generacemi (evoluce) působí na vzorce změn v jednotlivých organismech (rozvoj ).[61][62][63] Od počátku 21. století a ve světle objevů provedených v posledních desetiletích někteří biologové argumentovali pro rozšířená evoluční syntéza, což by vysvětlovalo účinky režimů negenetické dědičnosti, jako např epigenetika, rodičovské účinky, ekologické dědictví a kulturní dědictví, a vyvíjitelnost.[64][65]
Dědičnost
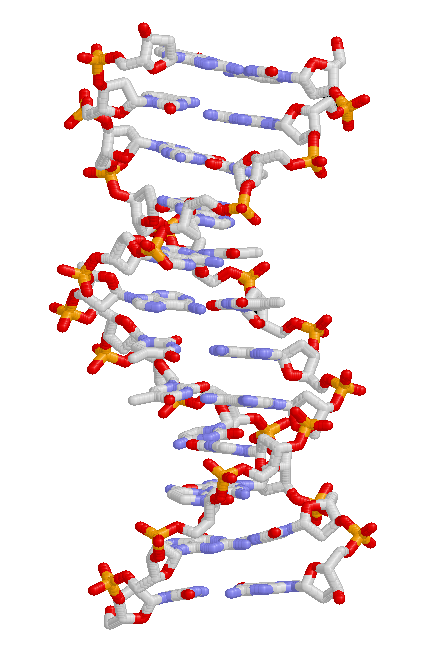
Evoluce v organismech nastává prostřednictvím změn dědičných znaků - zděděných vlastností organismu. Například u lidí barva očí je zděděná vlastnost a jednotlivec může zdědit „vlastnost hnědých očí“ od jednoho ze svých rodičů.[66] Zděděné vlastnosti jsou řízeny geny a kompletní sadou genů v organismu genom (genetický materiál) se nazývá jeho genotyp.[67]
Úplná sada pozorovatelných znaků, které tvoří strukturu a chování organismu, se nazývá jeho fenotyp. Tyto vlastnosti pocházejí z interakce jeho genotypu s prostředím.[68] Ve výsledku není mnoho aspektů fenotypu organismu zděděno. Například, opálený kůže pochází z interakce mezi genotypem člověka a slunečním světlem; opálení tedy nejsou předávána dětem lidí. Někteří lidé se však opálí snadněji než jiní, kvůli rozdílům v genotypových variacích; pozoruhodným příkladem jsou lidé se zděděnou vlastností albinismus, kteří se vůbec neopálí a jsou velmi citliví na úžeh.[69]
Dědičné vlastnosti se předávají z jedné generace na druhou prostřednictvím DNA, a molekula který kóduje genetickou informaci.[67] DNA je dlouhá biopolymer složený ze čtyř typů základen. Sekvence bází podél konkrétní molekuly DNA specifikuje genetickou informaci podobným způsobem jako posloupnost písmen vysvětlujících větu. Než se buňka rozdělí, zkopíruje se DNA, takže každá z výsledných dvou buněk zdědí sekvenci DNA. Části molekuly DNA, které specifikují jednu funkční jednotku, se nazývají geny; různé geny mají různé sekvence bází. V buňkách tvoří dlouhé řetězce DNA kondenzované struktury zvané chromozomy. Specifické umístění sekvence DNA v chromozomu je známé jako a místo. Pokud se sekvence DNA v lokusu mezi jednotlivci liší, různé formy této sekvence se nazývají alely. Sekvence DNA se mohou měnit prostřednictvím mutací a vytvářet nové alely. Pokud dojde k mutaci v genu, nová alela může ovlivnit vlastnost, kterou gen řídí, a změnit tak fenotyp organismu.[70] I když tato jednoduchá korespondence mezi alelou a znakem v některých případech funguje, většina znaků je složitější a je řízena kvantitativní znak loci (více interagujících genů).[71][72]
Nedávné nálezy potvrdily důležité příklady dědičných změn, které nelze vysvětlit změnami v pořadí nukleotidy v DNA. Tyto jevy jsou klasifikovány jako epigenetický dědické systémy.[73] Methylace DNA označení chromatin, soběstačné metabolické smyčky, umlčování genů pomocí Interference RNA a trojrozměrný konformace z bílkoviny (jako priony ) jsou oblasti, kde byly na úrovni organismu objeveny epigenetické dědičné systémy.[74][75] Vývojoví biologové naznačují, že komplexní interakce v genetické sítě a komunikace mezi buňkami může vést k dědičným variacím, které mohou podložit některé mechaniky vývojová plasticita a kanalizace.[76] Dědičnost se může vyskytnout i v ještě větších měřítcích. Například ekologické dědictví prostřednictvím procesu konstrukce výklenku je definována pravidelnou a opakovanou činností organismů v jejich prostředí. To generuje dědictví efektů, které se mění a vracejí se do režimu výběru následujících generací. Potomci dědí geny plus vlastnosti prostředí generované ekologickými akcemi předků.[77] Mezi další příklady dědičnosti v evoluci, které nejsou pod přímou kontrolou genů, patří dědičnost kulturní rysy a symbiogeneze.[78][79]
Zdroje variací


Evoluce může nastat, pokud je toho dost genetická variace v rámci populace. Variace pochází z mutací v genomu a jejich přeskupení sexuální reprodukce a migrace mezi populacemi (tok genů ). Navzdory neustálému zavádění nových variací prostřednictvím mutací a toku genů je většina genomu druhu stejná u všech jedinců tohoto druhu.[80] I relativně malé rozdíly v genotypu však mohou vést k dramatickým rozdílům ve fenotypu: například šimpanzi a lidé se liší pouze asi v 5% jejich genomů.[81]
Fenotyp jednotlivého organismu je výsledkem jeho genotypu i vlivu prostředí, ve kterém žil. Podstatná část fenotypové variace v populaci je způsobena genotypovou variací.[72] Moderní evoluční syntéza definuje evoluci jako změnu v čase v této genetické variantě. Frekvence jedné konkrétní alely bude více či méně převládající ve srovnání s jinými formami tohoto genu. Variace zmizí, když nová alela dosáhne bodu fixace —Když buď zmizí z populace, nebo úplně nahradí alelu předků.[82]
Před objevením Mendelovy genetiky byla jedna společná hypotéza smíšené dědictví. Ale s kombinováním dědičnosti by se genetická variace rychle ztratila, což by učinilo evoluci přirozeným výběrem nepravděpodobnou. The Hardy-Weinbergův princip poskytuje řešení toho, jak se udržuje variace v populaci s Mendelianovou dědičností. Četnosti alel (variace v genu) zůstanou konstantní, pokud nedojde k selekci, mutaci, migraci a genetickému driftu.[83]
Mutace
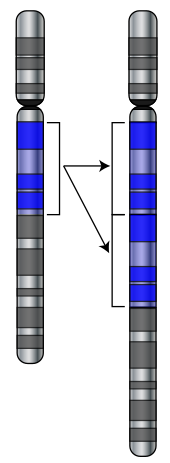
Mutace jsou změny v sekvenci DNA genomu buňky. Pokud dojde k mutacím, mohou změnit produkt genu, nebo zabránit fungování genu, nebo nemají žádný účinek. Na základě studií za běhu Drosophila melanogaster, bylo navrženo, že pokud mutace změní protein produkovaný genem, bude to pravděpodobně škodlivé, přičemž přibližně 70% těchto mutací má škodlivé účinky a zbytek je neutrální nebo slabě prospěšný.[84]
Mutace mohou zahrnovat velké části chromozomu duplikováno (obvykle do genetická rekombinace ), který může zavést další kopie genu do genomu.[85] Extra kopie genů jsou hlavním zdrojem suroviny potřebné pro vývoj nových genů.[86] To je důležité, protože většina nových genů se vyvíjí uvnitř genové rodiny z již existujících genů, které sdílejí společné předky.[87] Například člověk oko používá čtyři geny k vytvoření struktur, které snímají světlo: tři pro barevné vidění a jeden pro noční vidění; všechny čtyři pocházejí z jediného genu předků.[88]
Nové geny lze generovat z genu předků, když duplicitní kopie mutuje a získá novou funkci. Tento proces je jednodušší, jakmile byl gen duplikován, protože zvyšuje nadbytek systému; jeden gen v páru může získat novou funkci, zatímco druhá kopie nadále plní svoji původní funkci.[89][90] Jiné typy mutací mohou dokonce generovat zcela nové geny z dříve nekódující DNA.[91][92]
Generování nových genů může také zahrnovat duplikování malých částí několika genů, přičemž tyto fragmenty se poté rekombinují a vytvoří nové kombinace s novými funkcemi.[93][94] Když jsou nové geny shromážděny z míchání již existujících částí, domén fungují jako moduly s jednoduchými nezávislými funkcemi, které lze vzájemně kombinovat a vytvářet nové kombinace s novými a komplexními funkcemi.[95] Například, polyketid syntázy jsou velké enzymy které dělají antibiotika; obsahují až sto nezávislých domén, z nichž každá katalyzuje jeden krok v celém procesu, jako krok v montážní lince.[96]
Sex a rekombinace
v nepohlavní organismy, geny jsou zděděny společně, nebo propojeno, protože se během reprodukce nemohou mísit s geny jiných organismů. Naproti tomu potomci sexuální organismy obsahují náhodné směsi chromozomů jejich rodičů, které jsou vytvářeny prostřednictvím nezávislého sortimentu. V souvisejícím procesu zvaném homologní rekombinace si pohlavní organismy vyměňují DNA mezi dvěma odpovídajícími chromozomy.[97] Rekombinace a opětovné řazení nemění frekvence alel, ale naopak mění, které alely jsou navzájem spojeny, a produkují potomky s novými kombinacemi alel.[98] Pohlaví obvykle zvyšuje genetickou variabilitu a může zvyšovat rychlost evoluce.[99][100]

Dvojnásobnou cenu sexu poprvé popsal John Maynard Smith.[101] První cenou je, že u sexuálně dimorfních druhů může rodit mláďata pouze jedno ze dvou pohlaví. Tato cena se nevztahuje na hermafroditické druhy, jako je většina rostlin a mnoho dalších bezobratlých. Druhá cena spočívá v tom, že každý jedinec, který se pohlavně rozmnožuje, může přenést pouze 50% svých genů na jakéhokoli jednotlivého potomka, přičemž při předávání každé nové generace se přenáší ještě méně.[102] Přesto je sexuální reprodukce častějším prostředkem reprodukce mezi eukaryoty a mnohobuněčnými organismy. The Hypotéza Rudé královny byl použit k vysvětlení významu sexuální reprodukce jako prostředku umožňujícího neustálý vývoj a adaptaci v reakci na koevoluce s jinými druhy ve stále se měnícím prostředí.[102][103][104][105]
Tok genů
Tok genů je výměna genů mezi populacemi a mezi druhy.[106] Může to tedy být zdroj variací, který je pro populaci nebo druh nový. Tok genů může být způsoben pohybem jedinců mezi samostatnými populacemi organismů, což může být způsobeno pohybem myší mezi vnitrozemskými a pobřežními populacemi nebo pohybem pyl mezi populacemi trav odolnými vůči těžkým kovům a citlivým na těžké kovy.
Přenos genů mezi druhy zahrnuje tvorbu hybridní organismy a horizontální přenos genů. Horizontální přenos genů je přenos genetického materiálu z jednoho organismu do jiného organismu, který není jeho potomkem; toto je nejběžnější mezi bakterie.[107] V medicíně to přispívá k šíření odolnost proti antibiotikům jako když jedna bakterie získá geny rezistence, může je rychle přenést na jiné druhy.[108] Horizontální přenos genů z bakterií na eukaryota, jako jsou kvasinky Saccharomyces cerevisiae a nosatce fazole adzuki Callosobruchus chinensis vyskytl se.[109][110] Příkladem větších převodů je eukaryotický bdelloid rotifers, které obdržely řadu genů z bakterií, houby a rostliny.[111] Viry může také přenášet DNA mezi organismy, což umožňuje přenos genů i napříč biologické domény.[112]
K velkému přenosu genů došlo také mezi předky eukaryotické buňky a bakterie, během akvizice chloroplasty a mitochondrie. Je možné, že samotné eukaryoty pocházejí z horizontálních genových přenosů mezi bakteriemi a archaea.[113]
Mechanismy

Od a neodarwinovský perspektiva, k evoluci dochází, když dojde ke změnám ve frekvencích alel v populaci křížících se organismů,[83] například alela černé barvy v populaci můr je stále běžnější. Mechanismy, které mohou vést ke změnám ve frekvencích alel, zahrnují přirozený výběr, genetický drift, tok genů a zkreslení mutací.
Přírodní výběr
Evoluce pomocí přirozeného výběru je proces, při kterém se vlastnosti, které zvyšují přežití a reprodukci, stávají běžnějšími v následujících generacích populace. Často se tomu říká „samozřejmý“ mechanismus, protože nutně vyplývá ze tří jednoduchých faktů:[8]
- Varianty existují v populacích organismů s ohledem na morfologii, fyziologii a chování (fenotypová variace).
- Různé rysy udělují různou míru přežití a reprodukce (diferenciální zdatnost).
- Tyto vlastnosti lze předávat z generace na generaci (dědičnost zdatnosti).
Produkuje se více potomků, než kolik jich může přežít, a tyto podmínky produkují konkurenci mezi organismy o přežití a reprodukci. V důsledku toho organismy se znaky, které jim dávají výhodu nad svými konkurenty, s větší pravděpodobností přenesou své vlastnosti na další generaci než organismy se znaky, které jim nepřiznávají výhodu.[114] Tento teleonomie je kvalita, kterou proces přirozeného výběru vytváří a zachovává vlastnosti, které jsou zdánlivě vybavené pro funkční role, které plní.[115] Důsledky výběru zahrnují nenáhodné páření[116] a genetické stopování.
Ústředním konceptem přirozeného výběru je evoluční fitness organismu.[117] Fitness se měří schopností organismu přežít a rozmnožovat se, což určuje velikost jeho genetického přínosu pro další generaci.[117] Fitness však není stejný jako celkový počet potomků: místo toho je fitness indikován podílem následujících generací, které nesou geny organismu.[118] Například pokud by organismus mohl dobře přežít a rychle se množit, ale jeho potomci byli příliš malí a slabí na to, aby přežili, tento organismus by malým genetickým přínosem pro budoucí generace a měl by tak nízkou kondici.[117]
Pokud alela zvyšuje kondici více než ostatní alely tohoto genu, pak se s každou generací bude tato alela v populaci běžnější. O těchto vlastnostech se říká, že jsou „vybrány pro. "Příklady vlastností, které mohou zvýšit kondici, jsou lepší přežití a vyšší úrodnost. Naopak nižší kondice způsobená méně prospěšnou nebo škodlivou alelou vede k tomu, že se tato alela stává vzácnější - jsou vybrány proti."[119] Důležité je, že vhodnost alely není pevnou charakteristikou; pokud se prostředí změní, mohou se dříve neutrální nebo škodlivé vlastnosti stát prospěšnými a dříve prospěšné vlastnosti se mohou stát škodlivými.[70] I když se však směr výběru obrátí tímto způsobem, vlastnosti, které byly ztraceny v minulosti, se nemusí znovu vyvíjet ve stejné podobě (viz Dollov zákon ).[120][121] Opětovná aktivace spících genů, pokud však nebyly z genomu odstraněny a byly potlačeny jen možná po stovky generací, může vést k opětovnému výskytu znaků, o nichž se předpokládá, že jsou ztraceny jako zadní nohy v delfíni, zuby dovnitř kuřata, křídla v bezkřídlé tyči hmyz, ocasy a další bradavky u lidí atd.[122] „Návraty“, jako jsou tyto, jsou známé jako atavismy.

· Graf 1 ukazuje směrový výběr, ve kterém jediný extrém fenotyp je zvýhodněný.
· Graf 2 zobrazuje stabilizující výběr, kde je přechodný fenotyp upřednostňován před extrémními vlastnostmi.
· Graf 3 ukazuje rušivý výběr, kde jsou upřednostňovány extrémní fenotypy před meziproduktem.
Přirozený výběr v populaci pro vlastnost, která se může lišit v rozsahu hodnot, jako je například výška, lze rozdělit do tří různých typů. První je směrový výběr, což je posun průměrné hodnoty znaku v čase - například organismy pomalu rostou.[123] Za druhé, rušivý výběr je výběr extrémních hodnot vlastností a často vede k dvě různé hodnoty se stávají nejčastějšími, s výběrem oproti průměrné hodnotě. To by bylo, když by výhodu měly buď nízké nebo vysoké organismy, ale ne ty střední. Nakonec dovnitř stabilizující výběr na obou koncích je výběr proti extrémním hodnotám vlastností, což způsobí pokles rozptyl kolem průměrné hodnoty a menší rozmanitosti.[114][124] To by například způsobilo, že organismy budou mít nakonec podobnou výšku.
Přirozený výběr obecně dělá z přírody měřítko, proti kterému jednotlivci a jednotlivé rysy pravděpodobně přežijí. „Příroda“ v tomto smyslu odkazuje na ekosystém, to znamená systém, ve kterém organismy interagují s každým dalším prvkem, fyzický stejně jako biologický ve svém místním prostředí. Eugene Odum, zakladatel společnosti ekologie, definoval ekosystém jako: „Jakákoli jednotka, která zahrnuje všechny organismy ... v dané oblasti interagující s fyzickým prostředím, takže tok energie vede k jasně definované trofické struktuře, biotické rozmanitosti a materiálovým cyklům (tj. výměně materiálů mezi živými a neživými částmi) v systému .... "[125] Každá populace v ekosystému zaujímá odlišné výklenek nebo pozice s odlišnými vztahy k ostatním částem systému. Tyto vztahy zahrnují životní historii organismu, jeho postavení v potravní řetězec a jeho geografický rozsah. Toto široké chápání přírody umožňuje vědcům vymezit specifické síly, které společně tvoří přirozený výběr.
Přirozený výběr může působit na různé úrovně organizace, jako jsou geny, buňky, jednotlivé organismy, skupiny organismů a druhů.[126][127][128] Výběr může působit na více úrovních současně.[129] Příkladem selekce pod úrovní jednotlivého organismu jsou tzv. Geny transpozice, které se mohou replikovat a šířit po genomu.[130] Výběr na úrovni nad jednotlivcem, například výběr skupiny, může umožnit vývoj spolupráce.[131]
Genetické stopování
Rekombinace umožňuje oddělení alel na stejném řetězci DNA. Rychlost rekombinace je však nízká (přibližně dvě události na chromozom za generaci). Výsledkem je, že geny blízko sebe na chromozomu nemusí být vždy navzájem zamíchány a geny, které jsou blízko sebe, mají tendenci být zděděny společně, což je jev známý jako vazba.[132] Tato tendence se měří zjišťováním toho, jak často se dvě alely vyskytují společně na jednom chromozomu ve srovnání s očekávání, kterému se říká jejich vazebná nerovnováha. Sada alel, která se obvykle dědí ve skupině, se nazývá a haplotyp. To může být důležité, když je jedna alela v konkrétním haplotypu silně prospěšná: přirozený výběr může řídit a selektivní zametání což také způsobí, že ostatní alely v haplotypu budou v populaci běžnější; tento efekt se nazývá genetické stopování nebo genetický návrh.[133] Genetický návrh způsobený skutečností, že některé neutrální geny jsou geneticky spojeny s ostatními, které jsou pod selekcí, lze částečně zachytit vhodnou účinnou velikostí populace.[134]
Sexuální výběr
Zvláštním případem přirozeného výběru je sexuální výběr, což je výběr jakéhokoli znaku, který zvyšuje úspěšnost páření zvýšením atraktivity organismu pro potenciální kamarády.[135] Rysy, které se vyvinuly sexuálním výběrem, jsou zvláště výrazné u mužů několika druhů zvířat. Ačkoli jsou sexuálně upřednostňovány, rysy jako těžkopádné parohy, páření, velká velikost těla a jasné barvy často přitahují predaci, což ohrožuje přežití jednotlivých mužů.[136][137] Tato nevýhoda přežití je vyvážena vyšším reprodukčním úspěchem u mužů, který je vykazuje těžko falešný, sexuálně vybrané vlastnosti.[138]
Genetický drift

Genetický drift je náhodná fluktuace frekvence alel v populaci z jedné generace na druhou.[139] Pokud selektivní síly chybí nebo jsou relativně slabé, frekvence alel jsou stejně pravděpodobné drift nahoru nebo dolů v každé následující generaci, protože alely podléhají chyba vzorkování.[140] Tento drift se zastaví, když se alela nakonec zafixuje, a to buď zmizením z populace nebo úplným nahrazením ostatních alel. Genetický drift proto může eliminovat některé alely z populace pouze díky náhodě. I při absenci selektivních sil může genetický drift způsobit, že se dvě oddělené populace, které začínají se stejnou genetickou strukturou, driftují od sebe na dvě odlišné populace s různými sadami alel.[141]
The neutrální teorie molekulární evoluce navrhl, že většina evolučních změn je výsledkem fixace neutrální mutace genetickým driftem.[142] Proto je v tomto modelu většina genetických změn v populaci výsledkem konstantního mutačního tlaku a genetického driftu.[143] Tato forma neutrální teorie je nyní z velké části opuštěna, protože se nezdá, že by odpovídala genetické variabilitě viděné v přírodě.[144][145] Novější a lépe podporovanou verzí tohoto modelu je však téměř neutrální teorie kde mutace, která by byla efektivně neutrální u malé populace, nemusí být nutně neutrální u velké populace.[114] Jiné alternativní teorie navrhují, že genetický drift je převyšován jinými stochastickými silami v evoluci, jako je genetické stopování, známé také jako genetický návrh.[140][134][146]
Doba, po kterou se neutrální alela fixuje genetickým driftem, závisí na velikosti populace, u menších populací k fixaci dochází rychleji.[147] Počet jednotlivců v populaci není kritický, nýbrž opatření známé jako efektivní velikost populace.[148] Efektivní populace je obvykle menší než celková populace, protože bere v úvahu faktory, jako je úroveň příbuzenské plemenitby a stádium životního cyklu, ve kterém je populace nejmenší.[148] Efektivní velikost populace nemusí být stejná pro každý gen ve stejné populaci.[149]
Je obvykle obtížné měřit relativní význam výběru a neutrálních procesů, včetně driftu.[150] Srovnávací význam adaptivních a neadaptivních sil při řízení evolučních změn je oblastí aktuální výzkum.[151]
Tok genů
Tok genů zahrnuje výměnu genů mezi populacemi a mezi druhy.[106] Přítomnost nebo nepřítomnost toku genů zásadně mění průběh evoluce. Kvůli složitosti organismů se u každé ze dvou zcela izolovaných populací nakonec vyvine genetická nekompatibilita neutrálními procesy, jako v případě Bateson-Dobzhansky-Mullerův model, i když obě populace zůstávají v podstatě identické, pokud jde o jejich adaptaci na životní prostředí.
Pokud se vyvíjí genetická diferenciace mezi populacemi, tok genů mezi populacemi může zavést znaky nebo alely, které jsou pro místní populaci nevýhodné, a to může vést k tomu, že se v těchto populacích vytvoří organismy, které vyvinou mechanismy, které zabrání páření s geneticky vzdálenými populacemi, což nakonec povede k výskytu nových druhů . Výměna genetických informací mezi jednotlivci je tedy zásadně důležitá pro rozvoj Koncept biologických druhů (BSC).
Během vývoje moderní syntézy vyvinul Sewall Wright jeho teorie posunu rovnováhy, který považoval tok genů mezi částečně izolovanými populacemi za důležitý aspekt adaptivní evoluce.[152] Nedávno však došlo k zásadní kritice významu teorie posunu rovnováhy.[153]
Předpětí mutace
Mutační zaujatost je obvykle koncipován jako rozdíl v očekávaných rychlostech pro dva různé druhy mutací, např. zkreslení přechod-transverze, zkreslení GC-AT, zkreslení delece-vložení. To souvisí s myšlenkou vývojová zaujatost.
Haldane[154] a Fisher[155] tvrdil, že protože mutace je slabý tlak snadno překonatelný výběrem, tendence mutace by byly neúčinné, s výjimkou podmínek neutrálního vývoje nebo mimořádně vysokých rychlostí mutací. Tento argument opačných tlaků byl dlouho používán k odmítnutí možnosti vnitřních tendencí vývoje,[156] dokud molekulární éra nevyvolala obnovený zájem o neutrální evoluci.
Noboru Sueoka[157] a Ernst Freese[158] navrhl, že systematické předsudky v mutaci mohou být zodpovědné za systematické rozdíly ve složení genomového GC mezi druhy. Identifikace zkreslení GC E-coli kmen mutátorů v roce 1967,[159] spolu s návrhem neutrální teorie stanovil věrohodnost mutačních vysvětlení pro molekulární vzorce, které jsou nyní v literatuře molekulární evoluce běžné.
Například mutační předsudky jsou často vyvolávány v modelech použití kodonů.[160] Takové modely také zahrnují účinky selekce podle modelu mutace-výběru-driftu,[161] což umožňuje jak předsudky mutací, tak diferenciální výběr na základě účinků na překlad. Hypotézy zkreslení mutací hrály důležitou roli ve vývoji myšlení o vývoji složení genomu, včetně izochorů.[162] Different insertion vs. deletion biases in different taxony can lead to the evolution of different genome sizes.[163][164] The hypothesis of Lynch regarding genome size relies on mutational biases toward increase or decrease in genome size.
However, mutational hypotheses for the evolution of composition suffered a reduction in scope when it was discovered that (1) GC-biased gene conversion makes an important contribution to composition in diploid organisms such as mammals[165] and (2) bacterial genomes frequently have AT-biased mutation.[166]
Contemporary thinking about the role of mutation biases reflects a different theory from that of Haldane and Fisher. More recent work[156] showed that the original "pressures" theory assumes that evolution is based on standing variation: when evolution depends on the introduction of new alleles, mutational and developmental biases in introduction can impose biases on evolution without requiring neutral evolution or high mutation rates.
Several recent studies report that the mutations implicated in adaptation reflect common mutation biases[167][168][169] though others dispute this interpretation.[170]
Výsledky
Evolution influences every aspect of the form and behaviour of organisms. Most prominent are the specific behavioural and physical adaptations that are the outcome of natural selection. These adaptations increase fitness by aiding activities such as finding food, avoiding dravci or attracting mates. Organisms can also respond to selection by cooperating with each other, usually by aiding their relatives or engaging in mutually beneficial symbióza. In the longer term, evolution produces new species through splitting ancestral populations of organisms into new groups that cannot or will not interbreed.
These outcomes of evolution are distinguished based on time scale as makroevoluce versus microevolution. Macroevolution refers to evolution that occurs at or above the level of species, in particular speciation and extinction; whereas microevolution refers to smaller evolutionary changes within a species or population, in particular shifts in alela frekvence and adaptation.[172] In general, macroevolution is regarded as the outcome of long periods of microevolution.[173] Thus, the distinction between micro- and macroevolution is not a fundamental one—the difference is simply the time involved.[174] However, in macroevolution, the traits of the entire species may be important. For instance, a large amount of variation among individuals allows a species to rapidly adapt to new stanoviště, lessening the chance of it going extinct, while a wide geographic range increases the chance of speciation, by making it more likely that part of the population will become isolated. In this sense, microevolution and macroevolution might involve selection at different levels—with microevolution acting on genes and organisms, versus macroevolutionary processes such as výběr druhů acting on entire species and affecting their rates of speciation and extinction.[175][176][177]
A common misconception is that evolution has goals, long-term plans, or an innate tendency for "progress", as expressed in beliefs such as orthogenesis and evolutionism; realistically however, evolution has no long-term goal and does not necessarily produce greater complexity.[178][179][180] Ačkoli složité druhy have evolved, they occur as a side effect of the overall number of organisms increasing and simple forms of life still remain more common in the biosphere.[181] For example, the overwhelming majority of species are microscopic prokaryoty, which form about half the world's biomasa despite their small size,[182] and constitute the vast majority of Earth's biodiversity.[183] Simple organisms have therefore been the dominant form of life on Earth throughout its history and continue to be the main form of life up to the present day, with complex life only appearing more diverse because it is more noticeable.[184] Indeed, the evolution of microorganisms is particularly important to modern evolutionary research, since their rapid reproduction allows the study of experimental evolution and the observation of evolution and adaptation in real time.[185][186]
Přizpůsobování

Adaptation is the process that makes organisms better suited to their habitat.[187][188] Also, the term adaptation may refer to a trait that is important for an organism's survival. For example, the adaptation of koně ' teeth to the grinding of grass. By using the term přizpůsobování for the evolutionary process and adaptive trait for the product (the bodily part or function), the two senses of the word may be distinguished. Adaptations are produced by natural selection.[189] The following definitions are due to Theodosius Dobzhansky:
- Přizpůsobování is the evolutionary process whereby an organism becomes better able to live in its habitat or habitats.[190]
- Adaptedness is the state of being adapted: the degree to which an organism is able to live and reproduce in a given set of habitats.[191]
- An adaptive trait is an aspect of the developmental pattern of the organism which enables or enhances the probability of that organism surviving and reproducing.[192]
Adaptation may cause either the gain of a new feature, or the loss of an ancestral feature. An example that shows both types of change is bacterial adaptation to antibiotic selection, with genetic changes causing antibiotic resistance by both modifying the target of the drug, or increasing the activity of transporters that pump the drug out of the cell.[193] Other striking examples are the bacteria Escherichia coli evolving the ability to use kyselina citronová as a nutrient in a long-term laboratory experiment,[194] Flavobacterium evolving a novel enzyme that allows these bacteria to grow on the by-products of nylon výrobní,[195][196] and the soil bacterium Sfingobium evolving an entirely new metabolická cesta that degrades the synthetic pesticid pentachlorfenol.[197][198] An interesting but still controversial idea is that some adaptations might increase the ability of organisms to generate genetic diversity and adapt by natural selection (increasing organisms' evolvability).[199][200][201][202][203]
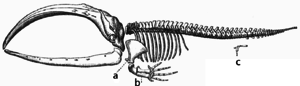
Adaptation occurs through the gradual modification of existing structures. Consequently, structures with similar internal organisation may have different functions in related organisms. This is the result of a single ancestral structure being adapted to function in different ways. The bones within netopýr wings, for example, are very similar to those in myši feet and primát hands, due to the descent of all these structures from a common mammalian ancestor.[205] However, since all living organisms are related to some extent,[206] even organs that appear to have little or no structural similarity, such as členovec, oliheň a obratlovců eyes, or the limbs and wings of arthropods and vertebrates, can depend on a common set of homologous genes that control their assembly and function; tomu se říká deep homology.[207][208]
During evolution, some structures may lose their original function and become vestigial structures.[209] Such structures may have little or no function in a current species, yet have a clear function in ancestral species, or other closely related species. Mezi příklady patří pseudogeny,[210] the non-functional remains of eyes in blind cave-dwelling fish,[211] wings in flightless birds,[212] the presence of hip bones in whales and snakes,[204] and sexual traits in organisms that reproduce via asexual reproduction.[213] Příklady vestigial structures in humans zahrnout wisdom teeth,[214] the kostrč,[209] the vermiformní příloha,[209] and other behavioural vestiges such as husí kůže[215][216] a primitivní reflexy.[217][218][219]
However, many traits that appear to be simple adaptations are in fact exaptations: structures originally adapted for one function, but which coincidentally became somewhat useful for some other function in the process.[220] One example is the African lizard Holaspis guentheri, which developed an extremely flat head for hiding in crevices, as can be seen by looking at its near relatives. However, in this species, the head has become so flattened that it assists in gliding from tree to tree—an exaptation.[220] Within cells, molecular machines such as the bacterial bičíky[221] a protein sorting machinery[222] evolved by the recruitment of several pre-existing proteins that previously had different functions.[172] Another example is the recruitment of enzymes from glykolýza a xenobiotický metabolismus to serve as structural proteins called krystaliny within the lenses of organisms' eyes.[223][224]
An area of current investigation in evolutionary developmental biology is the developmental basis of adaptations and exaptations.[225] This research addresses the origin and evolution of embryonální vývoj and how modifications of development and developmental processes produce novel features.[226] These studies have shown that evolution can alter development to produce new structures, such as embryonic bone structures that develop into the jaw in other animals instead forming part of the middle ear in mammals.[227] It is also possible for structures that have been lost in evolution to reappear due to changes in developmental genes, such as a mutation in chickens causing embryos to grow teeth similar to those of krokodýli.[228] It is now becoming clear that most alterations in the form of organisms are due to changes in a small set of conserved genes.[229]
Koevoluce

Interactions between organisms can produce both conflict and cooperation. When the interaction is between pairs of species, such as a patogen a a hostitel, or a predator and its prey, these species can develop matched sets of adaptations. Here, the evolution of one species causes adaptations in a second species. These changes in the second species then, in turn, cause new adaptations in the first species. This cycle of selection and response is called coevolution.[230] An example is the production of tetrodotoxin v drsný mlok and the evolution of tetrodotoxin resistance in its predator, the obyčejný podvazek had. In this predator-prey pair, an evoluční závod ve zbrojení has produced high levels of toxin in the newt and correspondingly high levels of toxin resistance in the snake.[231]
Spolupráce
Not all co-evolved interactions between species involve conflict.[232] Many cases of mutually beneficial interactions have evolved. For instance, an extreme cooperation exists between plants and the mykorhizní houby that grow on their roots and aid the plant in absorbing nutrients from the soil.[233] Tohle je reciproční relationship as the plants provide the fungi with sugars from fotosyntéza. Here, the fungi actually grow inside plant cells, allowing them to exchange nutrients with their hosts, while sending signály that suppress the plant imunitní systém.[234]
Coalitions between organisms of the same species have also evolved. Extrémním případem je eusocialita found in social insects, such as včely, termiti a mravenci, where sterile insects feed and guard the small number of organisms in a kolonie that are able to reproduce. On an even smaller scale, the somatic cells that make up the body of an animal limit their reproduction so they can maintain a stable organism, which then supports a small number of the animal's germ cells to produce offspring. Here, somatic cells respond to specific signals that instruct them whether to grow, remain as they are, or die. If cells ignore these signals and multiply inappropriately, their uncontrolled growth causes cancer.[235]
Such cooperation within species may have evolved through the process of příbuzný výběr, which is where one organism acts to help raise a relative's offspring.[236] This activity is selected for because if the pomáhání individual contains alleles which promote the helping activity, it is likely that its kin will taky contain these alleles and thus those alleles will be passed on.[237] Other processes that may promote cooperation include výběr skupiny, where cooperation provides benefits to a group of organisms.[238]
Speciace

Speciation is the process where a species diverges into two or more descendant species.[239]
There are multiple ways to define the concept of "species." The choice of definition is dependent on the particularities of the species concerned.[240] For example, some species concepts apply more readily toward sexually reproducing organisms while others lend themselves better toward asexual organisms. Despite the diversity of various species concepts, these various concepts can be placed into one of three broad philosophical approaches: interbreeding, ecological and phylogenetic.[241] The Biological Species Concept (BSC) is a classic example of the interbreeding approach. Defined by evolutionary biologist Ernst Mayr in 1942, the BSC states that "species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups."[242] Despite its wide and long-term use, the BSC like others is not without controversy, for example because these concepts cannot be applied to prokaryotes,[243] and this is called the druhový problém.[240] Some researchers have attempted a unifying monistic definition of species, while others adopt a pluralistic approach and suggest that there may be different ways to logically interpret the definition of a species.[240][241]
Barriers to reproduction between two diverging sexual populations are required for the populations to become new species. Gene flow may slow this process by spreading the new genetic variants also to the other populations. Depending on how far two species have diverged since their poslední společný předek, it may still be possible for them to produce offspring, as with horses and osli mating to produce muly.[244] Such hybrids are generally infertile. In this case, closely related species may regularly interbreed, but hybrids will be selected against and the species will remain distinct. However, viable hybrids are occasionally formed and these new species can either have properties intermediate between their parent species, or possess a totally new phenotype.[245] The importance of hybridisation in producing nový druh of animals is unclear, although cases have been seen in many types of animals,[246] s šedá rosnička being a particularly well-studied example.[247]
Speciation has been observed multiple times under both controlled laboratory conditions (see laboratory experiments of speciation ) and in nature.[248] In sexually reproducing organisms, speciation results from reproductive isolation followed by genealogical divergence. There are four primary geographic modes of speciation. The most common in animals is alopatrická speciace, which occurs in populations initially isolated geographically, such as by fragmentace stanovišť or migration. Selection under these conditions can produce very rapid changes in the appearance and behaviour of organisms.[249][250] As selection and drift act independently on populations isolated from the rest of their species, separation may eventually produce organisms that cannot interbreed.[251]
The second mode of speciation is peripatrická speciace, which occurs when small populations of organisms become isolated in a new environment. This differs from allopatric speciation in that the isolated populations are numerically much smaller than the parental population. Tady je founder effect causes rapid speciation after an increase in příbuzenské plemenitby increases selection on homozygotes, leading to rapid genetic change.[252]
The third mode is parapatrická speciace. This is similar to peripatric speciation in that a small population enters a new habitat, but differs in that there is no physical separation between these two populations. Instead, speciation results from the evolution of mechanisms that reduce gene flow between the two populations.[239] Generally this occurs when there has been a drastic change in the environment within the parental species' habitat. One example is the grass Anthoxanthum odoratum, which can undergo parapatric speciation in response to localised metal pollution from mines.[253] Here, plants evolve that have resistance to high levels of metals in the soil. Selection against interbreeding with the metal-sensitive parental population produced a gradual change in the flowering time of the metal-resistant plants, which eventually produced complete reproductive isolation. Selection against hybrids between the two populations may cause výztuž, which is the evolution of traits that promote mating within a species, as well as character displacement, which is when two species become more distinct in appearance.[254]

Finally, in sympatrická speciace species diverge without geographic isolation or changes in habitat. This form is rare since even a small amount of gene flow may remove genetic differences between parts of a population.[255] Generally, sympatric speciation in animals requires the evolution of both genetic differences a nonrandom mating, to allow reproductive isolation to evolve.[256]
One type of sympatric speciation involves křížení of two related species to produce a new hybrid species. This is not common in animals as animal hybrids are usually sterile. This is because during redukční dělení buněk the homologní chromozomy from each parent are from different species and cannot successfully pair. However, it is more common in plants because plants often double their number of chromosomes, to form polyploidy.[257] This allows the chromosomes from each parental species to form matching pairs during meiosis, since each parent's chromosomes are represented by a pair already.[258] An example of such a speciation event is when the plant species Arabidopsis thaliana a Arabidopsis arenosa crossbred to give the new species Arabidopsis suecica.[259] This happened about 20,000 years ago,[260] and the speciation process has been repeated in the laboratory, which allows the study of the genetic mechanisms involved in this process.[261] Indeed, chromosome doubling within a species may be a common cause of reproductive isolation, as half the doubled chromosomes will be unmatched when breeding with undoubled organisms.[262]
Speciation events are important in the theory of punctuated equilibrium, which accounts for the pattern in the fossil record of short "bursts" of evolution interspersed with relatively long periods of stasis, where species remain relatively unchanged.[263] In this theory, speciation and rapid evolution are linked, with natural selection and genetic drift acting most strongly on organisms undergoing speciation in novel habitats or small populations. As a result, the periods of stasis in the fossil record correspond to the parental population and the organisms undergoing speciation and rapid evolution are found in small populations or geographically restricted habitats and therefore rarely being preserved as fossils.[176]
Zánik

Extinction is the disappearance of an entire species. Extinction is not an unusual event, as species regularly appear through speciation and disappear through extinction.[264] Nearly all animal and plant species that have lived on Earth are now extinct,[265] and extinction appears to be the ultimate fate of all species.[266] These extinctions have happened continuously throughout the history of life, although the rate of extinction spikes in occasional mass události zániku.[267] The Událost vyhynutí křída – paleogen, during which the non-avian dinosaurs became extinct, is the most well-known, but the earlier Událost vyhynutí perm-trias was even more severe, with approximately 96% of all marine species driven to extinction.[267] The Událost vyhynutí holocénu is an ongoing mass extinction associated with humanity's expansion across the globe over the past few thousand years. Present-day extinction rates are 100–1000 times greater than the background rate and up to 30% of current species may be extinct by the mid 21st century.[268] Human activities are now the primary cause of the ongoing extinction event;[269][270] globální oteplování may further accelerate it in the future.[271] Despite the estimated extinction of more than 99 percent of all species that ever lived on Earth,[272][273] about 1 trillion species are estimated to be on Earth currently with only one-thousandth of one percent described.[274]
The role of extinction in evolution is not very well understood and may depend on which type of extinction is considered.[267] The causes of the continuous "low-level" extinction events, which form the majority of extinctions, may be the result of competition between species for limited resources (the princip konkurenčního vyloučení ).[61] If one species can out-compete another, this could produce species selection, with the fitter species surviving and the other species being driven to extinction.[127] The intermittent mass extinctions are also important, but instead of acting as a selective force, they drastically reduce diversity in a nonspecific manner and promote bursts of rapid evolution and speciation in survivors.[275]
Evoluční historie života
Původ života
The Země je stará asi 4,54 miliardy let.[276][277][278] The earliest undisputed evidence of life on Earth dates from at least 3.5 billion years ago,[13][279] Během Eoarchean Éra po geologii kůra začal tuhnout po dřívějším roztavení Hadean Eon. Microbial mat fossils have been found in 3.48 billion-year-old sandstone in Western Australia.[15][16][17] Other early physical evidence of a biogenic substance is graphite in 3.7 billion-year-old metasedimentární horniny discovered in Western Greenland[14] as well as "remains of biotický život " found in 4.1 billion-year-old rocks in Western Australia.[280][281] Commenting on the Australian findings, Stephen Blair Hedges wrote, "If life arose relatively quickly on Earth, then it could be common in the universe."[280][282] In July 2016, scientists reported identifying a set of 355 geny z poslední univerzální společný předek (LUCA) všech organismů žijících na Zemi.[283]
More than 99 percent of all species, amounting to over five billion species,[284] that ever lived on Earth are estimated to be extinct.[272][273] Estimates on the number of Earth's current species range from 10 million to 14 million,[285][286] of which about 1.9 million are estimated to have been named[287] and 1.6 million documented in a central database to date,[288] leaving at least 80 percent not yet described.
Highly energetic chemistry is thought to have produced a self-replicating molecule around 4 billion years ago, and half a billion years later the last common ancestor of all life existoval.[11] The current scientific consensus is that the complex biochemistry that makes up life came from simpler chemical reactions.[289][290] The beginning of life may have included self-replicating molecules such as RNA[291] and the assembly of simple cells.[292]
Společný sestup
All organisms on Earth are descended from a common ancestor or ancestral gene pool.[206][293] Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events.[294] The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities. Třetí, zakrnělé rysy with no clear purpose resemble functional ancestral traits. Fourth, organisms can be classified using these similarities into a hierarchy of nested groups, similar to a family tree.[295]
Modern research has suggested that, due to horizontální přenos genů, this "tree of life" may be more complicated than a simple branching tree since some genes have spread independently between distantly related species.[296][297] To solve this problem and others, some authors prefer to use the "Coral of life " as a metaphor or a mathematical model to illustrate the evolution of life. This view dates back to an idea briefly mentioned by Darwin but later abandoned.[298]
Minulé druhy také zanechaly záznamy o své evoluční historii. Fosílie spolu se srovnávací anatomií dnešních organismů tvoří morfologický nebo anatomický záznam.[299] Porovnáním anatomií moderních i vyhynulých druhů mohou paleontologové odvodit linie těchto druhů. Tento přístup je však nejúspěšnější pro organismy, které měly tvrdé části těla, jako jsou mušle, kosti nebo zuby. Dále, protože prokaryoty, jako jsou bakterie a archea, sdílejí omezený soubor běžných morfologií, jejich fosílie neposkytují informace o jejich předcích.
More recently, evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic set of nucleotides and aminokyseliny.[300] Vývoj molekulární genetika has revealed the record of evolution left in organisms' genomes: dating when species diverged through the molekulární hodiny produced by mutations.[301] For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 98% of their genomes and analysing the few areas where they differ helps shed light on when the common ancestor of these species existed.[302]
Evolution of life

Prokaryoti obývali Zemi před přibližně 3–4 miliardami let.[304][305] No obvious changes in morphology or cellular organisation occurred in these organisms over the next few billion years.[306] Eukaryotické buňky se objevily před 1,6–2,7 miliardami let. Další velká změna v buněčné struktuře nastala, když byly bakterie pohlceny eukaryotickými buňkami, v tzv. Kooperativní asociaci endosymbióza.[307][308] Pohltené bakterie a hostitelská buňka poté prošly koevolucí, kdy se bakterie vyvinuly buď do mitochondrií nebo hydrogenosomy.[309] Další pohltení cyanobacterial podobné organismy vedly k tvorbě chloroplastů v řasách a rostlinách.[310]
Historie života byla ta jednobuněčný eukaryoty, prokaryoty a archea až do doby před asi 610 miliony let, kdy se v oceánech začaly objevovat mnohobuněčné organismy Ediacaran doba.[304][311] The vývoj mnohobuněčnosti se vyskytly v několika nezávislých událostech, v organismech tak rozmanitých jako houby, hnědé řasy, sinice, slizové formy a myxobakterie.[312] In January 2016, scientists reported that, about 800 million years ago, a minor genetic change in a single molecule called GK-PID may have allowed organisms to go from a single cell organism to one of many cells.[313]
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over approximately 10 million years, in an event called the Kambrijská exploze. Tady většina typy ve fosilních záznamech se objevily moderní zvířata a také jedinečné linie, které následně vyhynuly.[314] Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere from photosynthesis.[315]
About 500 million years ago, plants and fungi colonised the land and were soon followed by arthropods and other animals.[316] Insects were particularly successful and even today make up the majority of animal species.[317] Obojživelníci first appeared around 364 million years ago, followed by early amniotes and birds around 155 million years ago (both from "plaz "-like lineages), savci around 129 million years ago, homininae around 10 million years ago and moderní lidé around 250,000 years ago.[318][319][320] However, despite the evolution of these large animals, smaller organisms similar to the types that evolved early in this process continue to be highly successful and dominate the Earth, with the majority of both biomass and species being prokaryotes.[183]
Aplikace
Concepts and models used in evolutionary biology, such as natural selection, have many applications.[321]
Artificial selection is the intentional selection of traits in a population of organisms. This has been used for thousands of years in the domestikace rostlin a živočichů.[322] V poslední době se takový výběr stal důležitou součástí genetické inženýrství, s volitelné značky such as antibiotic resistance genes being used to manipulate DNA. Proteins with valuable properties have evolved by repeated rounds of mutation and selection (for example modified enzymes and new protilátky ) v procesu zvaném řízená evoluce.[323]
Pochopení změn, ke kterým došlo během evoluce organismu, může odhalit geny potřebné ke konstrukci částí těla, geny, které se mohou podílet na člověku genetické poruchy.[324] Například Mexická tetra je albín cavefish, který během evoluce ztratil zrak. Společný chov různých populací této slepé ryby produkoval některé potomky s funkčními očima, protože v izolovaných populacích, které se vyvinuly v různých jeskyních, došlo k různým mutacím.[325] To pomohlo identifikovat geny potřebné pro vidění a pigmentaci.[326]
Evoluční teorie má v medicíně mnoho aplikací. Mnoho lidských nemocí není statickým jevem, ale je schopno evoluce. Viry, bakterie, houby a rakoviny vyvíjet tak, aby byl odolný vůči hostiteli imunitní obrana, stejně jako farmaceutické léky.[327][328][329] Stejné problémy se vyskytují v zemědělství s pesticid[330] a herbicid[331] odpor. Je možné, že čelíme konci efektivní životnosti většiny dostupných antibiotik[332] a předpovídat vývoj a vývoj[333] našich patogenů a navrhování strategií, jak to zpomalit nebo obejít, vyžaduje hlubší znalost komplexních sil, které řídí vývoj na molekulární úrovni.[334]
v počítačová věda, simulace evoluce pomocí evoluční algoritmy a umělý život byla zahájena v 60. letech a byla rozšířena o simulaci umělého výběru.[335] Umělá evoluce se stala široce uznávanou optimalizační metodou jako výsledek práce Ingo Rechenberg v šedesátých letech. Použil evoluční strategie řešit složité technické problémy.[336] Genetické algoritmy zvláště se stal populárním psaním John Henry Holland.[337] Mezi praktické aplikace patří také automatický vývoj počítačových programů.[338] Evoluční algoritmy se nyní používají k efektivnějšímu řešení vícerozměrných problémů než software vytvořený lidskými designéry a také k optimalizaci návrhu systémů.[339]
Sociální a kulturní reakce

V 19. století, zejména po vydání O původu druhů v roce 1859 byla myšlenka, že se život vyvinul, aktivním zdrojem akademické debaty zaměřené na filozofické, sociální a náboženské důsledky evoluce. Dnes je moderní evoluční syntéza přijímána velkou většinou vědců.[61] Evoluce však pro některé zůstává sporným konceptem teisté.[341]
Zatímco různá náboženství a označení spojili své víry s evolucí prostřednictvím konceptů jako teistická evoluce, existují kreacionisté kteří věří, že evoluce je v rozporu s mýty o stvoření nalezené v jejich náboženství a kteří vychovávají různé námitky proti evoluci.[172][342][343] Jak prokázaly reakce na zveřejnění Pozůstatky přirozené historie stvoření v roce 1844 je nejkontroverznějším aspektem evoluční biologie implikace lidská evoluce že lidé mají společný původ s lidoopy a že duševní a morální schopnosti lidstva mají stejné typy přirozených příčin jako jiné zděděné rysy u zvířat.[344] V některých zemích, zejména ve Spojených státech, toto napětí mezi vědou a náboženstvím podpořilo současnou kontroverzi stvoření - evoluce, náboženský konflikt zaměřený na politika a veřejné vzdělávání.[345] Zatímco jiné vědecké obory jako např kosmologie[346] a Věda o Zemi[347] také v rozporu s doslovnými interpretacemi mnoha náboženské texty, evoluční biologie zažívá podstatně větší odpor náboženských literalistů.
Výuka evoluce na hodinách biologie na středních školách v Americe byla ve většině první poloviny 20. století neobvyklá. The Scope Trial Rozhodnutí z roku 1925 způsobilo, že se předmět stal v amerických učebnicích sekundární biologie velmi vzácným po celou generaci, ale později byl postupně znovu zaveden a stal se právně chráněn v roce 1968. Epperson v. Arkansas rozhodnutí. Od té doby konkurenční náboženská víra kreacionismus byl v 70. a 80. letech právně zakázán v učebních osnovách středních škol při různých rozhodnutích, ale vrátil se dovnitř pseudovědecký formulář jako inteligentní design (ID), který bude v roce 2005 opět vyloučen Kitzmiller v. Dover Area School District případ.[348] Debata o Darwinových myšlenkách nevyvolala v Číně významnou polemiku.[349]
Viz také
Reference
- ^ Hall & Hallgrímsson 2008, str.4–6
- ^ „Evoluční zdroje“. Washington DC: Národní akademie věd, inženýrství a medicíny. 2016. Archivováno z původního dne 2016-06-03.
- ^ Futuyma & Kirkpatrick 2017, str. 79–102, kapitola 4: Mutace a variace
- ^ Scott-Phillips, Thomas C .; Laland, Kevin N .; Shuker, David M .; Dickins, Thomas E .; West, Stuart A. (Květen 2014). „Perspektiva specializované konstrukce: kritické hodnocení“. Vývoj. 68 (5): 1231–1243. doi:10.1111 / evo.12332. ISSN 0014-3820. PMC 4261998. PMID 24325256.
Evoluční procesy jsou obecně považovány za procesy, při nichž k těmto změnám dochází. Čtyři takové procesy jsou široce uznávány: přirozený výběr (v širším smyslu zahrnuje sexuální výběr), genetický drift, mutace a migrace (Fisher 1930; Haldane 1932). Poslední dva generují variace; první dva to třídí.
- ^ Hall & Hallgrímsson 2008, s. 3–5
- ^ Voet, Voet & Pratt 2016, s. 1–22, Kapitola 1: Úvod do chemie života
- ^ Darwin 1859
- ^ A b Lewontin, Richard C. (Listopad 1970). „Jednotky výběru“ (PDF). Výroční přehled ekologie a systematiky. 1: 1–18. doi:10.1146 / annurev.es.01.110170.000245. ISSN 1545-2069. JSTOR 2096764. Archivováno (PDF) z původního dne 2015-02-06.
- ^ Futuyma & Kirkpatrick 2017, s. 3–26, Kapitola 1: Evoluční biologie
- ^ Kampourakis 2014, str.127–129
- ^ A b Doolittle, W. Ford (Únor 2000). „Vykořenění stromu života“ (PDF). Scientific American. 282 (2): 90–95. Bibcode:2000SciAm.282b..90D. doi:10.1038 / scientificamerican0200-90. ISSN 0036-8733. PMID 10710791. Archivovány od originál (PDF) dne 2006-09-07. Citováno 2015-04-05.
- ^ Glansdorff, Nicolas; Ying Xu; Labedan, Bernard (9. července 2008). „Poslední univerzální společný předek: vznik, složení a genetické dědictví nepolapitelného předchůdce“. Biology Direct. 3: 29. doi:10.1186/1745-6150-3-29. ISSN 1745-6150. PMC 2478661. PMID 18613974.
- ^ A b Schopf, J. William; Kudryavtsev, Anatolij B .; Czaja, Andrew D .; Tripathi, Abhishek B. (5. října 2007). "Důkazy o archeanském životě: stromatolity a mikrofosílie". Prekambrický výzkum. 158 (3–4): 141–155. Bibcode:2007PreR..158..141S. doi:10.1016 / j.precamres.2007.04.009. ISSN 0301-9268.
- ^ A b Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; et al. (Leden 2014). „Důkazy o biogenním grafitu v časných archaeanských metasedimentárních horninách Isua“. Nature Geoscience. 7 (1): 25–28. Bibcode:2014NatGe ... 7 ... 25O. doi:10.1038 / ngeo2025. ISSN 1752-0894.
- ^ A b Borenstein, Seth (13. listopadu 2013). „Nalezena nejstarší fosílie: Seznamte se se svou mikrobiální matkou“. Rozrušit. Yonkers, New York: Interaktivní síť Mindspark. Associated Press. Archivováno z původního 29. června 2015. Citováno 2015-05-31.
- ^ A b Pearlman, Jonathan (13. listopadu 2013). "Nalezeny nejstarší známky života na Zemi". The Daily Telegraph. Londýn: Telegraph Media Group. Archivováno od originálu dne 2014-12-16. Citováno 2014-12-15.
- ^ A b Noffke, Nora; Christian, Daniel; Wacey, David; Hazen, Robert M. (16. listopadu 2013). "Mikrobiálně indukované sedimentární struktury zaznamenávající starověký ekosystém v ca. 3,48 miliardy let stará komoda, Pilbara, Západní Austrálie ". Astrobiologie. 13 (12): 1103–1124. Bibcode:2013AsBio..13.1103N. doi:10.1089 / ast.2013.1030. ISSN 1531-1074. PMC 3870916. PMID 24205812.
- ^ Futuyma 2004, str. 33
- ^ Panno 2005, str. xv-16
- ^ NAS 2008, p. 17 Archivováno 2015-06-30 na Wayback Machine
- ^ Futuyma, Douglas J., vyd. (1999). „Evoluce, věda a společnost: Evoluční biologie a národní program výzkumu“ (PDF) (Shrnutí). New Brunswick, New Jersey: Office of University Publications, Rutgers, Státní univerzita v New Jersey. OCLC 43422991. Archivovány od originál (PDF) dne 31.01.2012. Citováno 2014-11-24.
- ^ Darwin 1909, str. 53
- ^ Kirk, Raven & Schofield 1983, s. 100–142, 280–321
- ^ Lucretius
- ^ Sedley, David (2003). „Lucretius and the New Empedocles“ (PDF). Leeds International Classical Studies. 2 (4). ISSN 1477-3643. Archivovány od originál (PDF) dne 2014-08-23. Citováno 2014-11-25.
- ^ Torrey, Harry Beal; Felin, Frances (březen 1937). „Byl Aristoteles evolucionista?“. Čtvrtletní přehled biologie. 12 (1): 1–18. doi:10.1086/394520. ISSN 0033-5770. JSTOR 2808399. S2CID 170831302.
- ^ Hull, David L. (Prosinec 1967). „Metafyzika evoluce“. Britský časopis pro dějiny vědy. Cambridge: Cambridge University Press jménem Britská společnost pro dějiny vědy. 3 (4): 309–337. doi:10.1017 / S0007087400002892. JSTOR 4024958.
- ^ Mason 1962, str. 43–44
- ^ Mayr 1982, str. 256–257
- ^ Wagoner, Ben (7. července 2000). „Carl Linnaeus (1707-1778)“. Vývoj (Online výstava). Berkeley, Kalifornie: Muzeum paleontologie University of California. Archivováno z původního dne 2011-04-30. Citováno 2012-02-11.
- ^ Bowler 2003, str. 73–75
- ^ „Erasmus Darwin (1731-1802)“. Vývoj (Online výstava). Berkeley, Kalifornie: Muzeum paleontologie University of California. 4. října 1995. Archivováno od originálu dne 2012-01-19. Citováno 2012-02-11.
- ^ Lamarck 1809
- ^ A b Nardon & Grenier 1991, str. 162
- ^ A b C Gould 2002[stránka potřebná ]
- ^ Ghiselin, Michael T. (Září – říjen 1994). „The Imaginary Lamarck: A Look at Bogus 'History' in Schoolbooks“. Dopis učebnice. OCLC 23228649. Archivovány od originál dne 12. 2. 2008. Citováno 2008-01-23.
- ^ Magner 2002[stránka potřebná ]
- ^ Jablonka, Eva; Jehněčí, Marion J. (Srpen 2007). „Précis of Evolution in Four Dimensions“. Behaviorální a mozkové vědy. 30 (4): 353–365. doi:10.1017 / S0140525X07002221. ISSN 0140-525X. PMID 18081952. S2CID 15879804.
- ^ Burkhardt & Smith 1991
- „Darwin, C. R. Lubbockovi, Johne“. Darwinův korespondenční projekt. Cambridge: Univerzita v Cambridge. Archivováno od původního dne 2014-12-15. Citováno 2014-12-01. Dopis 2532, 22. listopadu 1859.
- ^ Sulloway, Frank J. (Červen 2009). "Proč Darwin odmítl inteligentní design". Journal of Biosciences. 34 (2): 173–183. doi:10.1007 / s12038-009-0020-8. ISSN 0250-5991. PMID 19550032. S2CID 12289290.
- ^ Dawkins 1990[stránka potřebná ]
- ^ „Výsledky hledání pro„ sestup s úpravou “- Kompletní dílo Charlese Darwina online“.
- ^ Střízlivý, Elliott (16. června 2009). „Napsal Darwin Původ zpět?". Proc. Natl. Acad. Sci. USA 106 (Příloha 1): 10048–10055. Bibcode:2009PNAS..10610048S. doi:10.1073 / pnas.0901109106. ISSN 0027-8424. PMC 2702806. PMID 19528655.
- ^ Mayr 2002, str. 165
- ^ Bowler 2003, s. 145–146
- ^ Sokal, Robert R.; Crovello, Theodore J. (březen – duben 1970). „Koncept biologických druhů: kritické hodnocení“. Americký přírodovědec. 104 (936): 127–153. doi:10.1086/282646. ISSN 0003-0147. JSTOR 2459191. S2CID 83528114.
- ^ Darwin, Charles; Wallace, Alfred (20. srpna 1858). „O tendenci druhů vytvářet odrůdy a o zachování odrůd a druhů přirozenými způsoby výběru“. Journal of Proceedings of the Linnean Society of London. Zoologie. 3 (9): 45–62. doi:10.1111 / j.1096-3642.1858.tb02500.x. ISSN 1096-3642. Archivováno z původního dne 2007-07-14. Citováno 2007-05-13.
- ^ Desmond, Adrian J. (17. července 2014). „Thomas Henry Huxley“. Encyklopedie Britannica online. Chicago, Illinois: Encyklopedie Britannica, Inc. Archivováno z původního dne 19. ledna 2015. Citováno 2014-12-02.
- ^ Y. -S. Liu; X. M. Zhou; M. X. Zhi; X. J. Li; Q. L. Wang (září 2009). „Darwinovy příspěvky ke genetice“. Journal of Applied Genetics. 50 (3): 177–184. doi:10.1007 / BF03195671. ISSN 1234-1983. PMID 19638672. S2CID 19919317.
- ^ Weiling, Franz (červenec 1991). „Historická studie: Johann Gregor Mendel 1822–1884“. American Journal of Medical Genetics. 40 (1): 1–25, diskuse 26. doi:10,1002 / ajmg.1320400103. PMID 1887835.
- ^ Wright 1984, str. 480
- ^ Provine 1971
- ^ Stamhuis, Ida H .; Meijer, Onno G .; Zevenhuizen, Erik J. A. (červen 1999). „Hugo de Vries on Heredity, 1889-1903: Statistics, Mendelian Laws, Pangenes, Mutations“. Isis. 90 (2): 238–267. doi:10.1086/384323. JSTOR 237050. PMID 10439561. S2CID 20200394.
- ^ Quammen 2006[stránka potřebná ]
- ^ Bowler 1989[stránka potřebná ]
- ^ Watson, J. D.; Crick, F. H. C. (25. dubna 1953). „Molekulární struktura nukleových kyselin: Struktura nukleové kyseliny deoxyribózy“ (PDF). Příroda. 171 (4356): 737–738. Bibcode:1953Natur.171..737W. doi:10.1038 / 171737a0. ISSN 0028-0836. PMID 13054692. S2CID 4253007. Archivováno (PDF) z původního dne 2014-08-23. Citováno 2014-12-04.
Neuniklo nám naše vědomí, že konkrétní párování, které jsme okamžitě postulovali, naznačuje možný mechanismus kopírování genetického materiálu.
- ^ Hennig 1999, str. 280
- ^ Wiley & Lieberman 2011[stránka potřebná ]
- ^ Dobžanský, Theodosius (Březen 1973). „Nic v biologii nedává smysl, kromě světla evoluce“ (PDF). Americký učitel biologie. 35 (3): 125–129. CiteSeerX 10.1.1.324.2891. doi:10.2307/4444260. JSTOR 4444260. S2CID 207358177. Archivováno (PDF) od originálu 2015-10-23.
- ^ Levinson 2019.
- ^ A b C Kutschera, Ulrich; Niklas, Karl J. (Červen 2004). „Moderní teorie biologické evoluce: rozšířená syntéza“. Naturwissenschaften. 91 (6): 255–276. Bibcode:2004NW ..... 91..255K. doi:10.1007 / s00114-004-0515-r. ISSN 1432-1904. PMID 15241603. S2CID 10731711.
- ^ Cracraft & Bybee 2005[stránka potřebná ]
- ^ Avise, John C.; Ayala, Francisco J. (11. května 2010). „Ve světle evoluce IV: Stav člověka“ (PDF). Proc. Natl. Acad. Sci. USA. 107 (Příloha 2): 8897–8901. doi:10.1073 / pnas.1003214107. ISSN 0027-8424. PMC 3024015. PMID 20460311. Archivováno (PDF) z původního dne 2014-08-23. Citováno 2014-12-29.
- ^ Danchin, Étienne; Charmantier, Anne; Šampaňské, Frances A.; Mesoudi, Alex; Pujol, Benoit; Blanchet, Simon (červen 2011). „Beyond DNA: integrating inclusive inheritance into an extended theory of evolution“. Genetika hodnocení přírody. 12 (7): 475–486. doi:10.1038 / nrg3028. ISSN 1471-0056. PMID 21681209. S2CID 8837202.
- ^ Pigliucci & Müller 2010
- ^ Sturm, Richard A .; Frudakis, Tony N. (srpen 2004). "Barva očí: portály do pigmentačních genů a předků". Trendy v genetice. 20 (8): 327–332. doi:10.1016 / j.tig.2004.06.010. ISSN 0168-9525. PMID 15262401.
- ^ A b Pearson, Helen (25. května 2006). „Genetika: Co je to gen?“. Příroda. 441 (7092): 398–401. Bibcode:2006 Natur.441..398P. doi:10.1038 / 441398a. ISSN 0028-0836. PMID 16724031. S2CID 4420674.
- ^ Visscher, Peter M .; Hill, William G.; Wray, Naomi R. (Duben 2008). „Dědičnost v éře genomiky - koncepty a mylné představy“. Genetika hodnocení přírody. 9 (4): 255–266. doi:10.1038 / nrg2322. ISSN 1471-0056. PMID 18319743. S2CID 690431.
- ^ Oetting, William S .; Brilantní, Murray H .; King, Richard A. (srpen 1996). „Klinické spektrum albinismu u lidí“. Molekulární medicína dnes. 2 (8): 330–335. doi:10.1016/1357-4310(96)81798-9. ISSN 1357-4310. PMID 8796918.
- ^ A b Futuyma 2005[stránka potřebná ]
- ^ Phillips, Patrick C. (listopad 2008). „Epistáza - základní role genových interakcí ve struktuře a vývoji genetických systémů“. Genetika hodnocení přírody. 9 (11): 855–867. doi:10.1038 / nrg2452. ISSN 1471-0056. PMC 2689140. PMID 18852697.
- ^ A b Rongling Wu; Min Lin (březen 2006). "Funkční mapování - jak mapovat a studovat genetickou architekturu dynamických komplexních znaků". Genetika hodnocení přírody. 7 (3): 229–237. doi:10.1038 / nrg1804. ISSN 1471-0056. PMID 16485021. S2CID 24301815.
- ^ Jablonka, Eva; Raz, Gal (červen 2009). „Transgenerační epigenetická dědičnost: prevalence, mechanismy a důsledky pro studium dědičnosti a evoluce“ (PDF). Čtvrtletní přehled biologie. 84 (2): 131–176. CiteSeerX 10.1.1.617.6333. doi:10.1086/598822. ISSN 0033-5770. PMID 19606595. S2CID 7233550.
- ^ Bossdorf, Oliver; Arcuri, Davide; Richards, Christina L .; Pigliucci, Massimo (Květen 2010). "Experimentální změna methylace DNA ovlivňuje fenotypovou plasticitu ekologicky relevantních znaků v Arabidopsis thaliana" (PDF). Evoluční ekologie. 24 (3): 541–553. doi:10.1007 / s10682-010-9372-7. ISSN 0269-7653. S2CID 15763479.
- ^ Jablonka & Lamb 2005[stránka potřebná ]
- ^ Jablonka, Eva; Lamb, Marion J. (prosinec 2002). „Měnící se koncept epigenetiky“. Annals of the New York Academy of Sciences. 981 (1): 82–96. Bibcode:2002NYASA.981 ... 82J. doi:10.1111 / j.1749-6632.2002.tb04913.x. ISSN 0077-8923. PMID 12547675. S2CID 12561900.
- ^ Laland, Kevin N .; Sterelny, Kim (Září 2006). „Perspektiva: Sedm důvodů (ne) pro zanedbání specializované konstrukce“. Vývoj. 60 (9): 1751–1762. doi:10.1111 / j.0014-3820.2006.tb00520.x. ISSN 0014-3820. PMID 17089961. S2CID 22997236.
- ^ Chapman, Michael J .; Margulis, Lynn (Prosinec 1998). „Morfogeneze symbiogenezí“ (PDF). Mezinárodní mikrobiologie. 1 (4): 319–326. ISSN 1139-6709. PMID 10943381. Archivovány od originál (PDF) dne 2014-08-23. Citováno 2014-12-09.
- ^ Wilson, David Sloan; Wilson, Edward O. (Prosinec 2007). „Přehodnocení teoretických základů sociobiologie“ (PDF). Čtvrtletní přehled biologie. 82 (4): 327–348. doi:10.1086/522809. ISSN 0033-5770. PMID 18217526. S2CID 37774648. Archivovány od originál (PDF) dne 2011-05-11.
- ^ Butlin, Roger K .; Tregenza, Tom (28. února 1998). „Úrovně genetického polymorfismu: lokusy markerů versus kvantitativní znaky“. Filozofické transakce královské společnosti B. 353 (1366): 187–198. doi:10.1098 / rstb.1998.0201. ISSN 0962-8436. PMC 1692210. PMID 9533123.
- Butlin, Roger K .; Tregenza, Tom (29. prosince 2000). „Oprava pro Butlina a Tregenzu, Úrovně genetického polymorfismu: markerové lokusy versus kvantitativní znaky“. Filozofické transakce královské společnosti B. 355 (1404): 1865. doi:10.1098 / rstb.2000.2000. ISSN 0962-8436.
Některé z hodnot v tabulce 1 na str. 193 bylo dáno nesprávně. Chyby nemají vliv na závěry vyvozené v článku. Opravená tabulka je uvedena níže.
- Butlin, Roger K .; Tregenza, Tom (29. prosince 2000). „Oprava pro Butlina a Tregenzu, Úrovně genetického polymorfismu: markerové lokusy versus kvantitativní znaky“. Filozofické transakce královské společnosti B. 355 (1404): 1865. doi:10.1098 / rstb.2000.2000. ISSN 0962-8436.
- ^ Wetterbom, Anna; Sevov, Marie; Cavelier, Lucia; Bergström, Tomáš F. (listopad 2006). „Srovnávací genomická analýza člověka a šimpanze naznačuje klíčovou roli pro indely ve vývoji primátů“. Journal of Molecular Evolution. 63 (5): 682–690. Bibcode:2006JMolE..63..682W. doi:10.1007 / s00239-006-0045-7. ISSN 0022-2844. PMID 17075697. S2CID 7257193.
- ^ Amos, William; Harwood, John (28. února 1998). „Faktory ovlivňující úrovně genetické rozmanitosti v přírodních populacích“. Filozofické transakce královské společnosti B. 353 (1366): 177–186. doi:10.1098 / rstb.1998.0200. ISSN 0962-8436. PMC 1692205. PMID 9533122.
- ^ A b Ewens 2004[stránka potřebná ]
- ^ Sawyer, Stanley A .; Parsch, John; Zhang Zhi; Hartl, Daniel L. (17. dubna 2007). "Prevalence pozitivní selekce mezi téměř neutrálními aminokyselinovými náhradami v Drosophila". Proc. Natl. Acad. Sci. USA. 104 (16): 6504–6510. Bibcode:2007PNAS..104,6504S. doi:10.1073 / pnas.0701572104. ISSN 0027-8424. PMC 1871816. PMID 17409186.
- ^ Hastings, P. J .; Lupski, James R.; Rosenberg, Susan M .; Ira, Grzegorz (srpen 2009). "Mechanismy změny počtu kopií genů". Genetika hodnocení přírody. 10 (8): 551–564. doi:10.1038 / nrg2593. ISSN 1471-0056. PMC 2864001. PMID 19597530.
- ^ Carroll, Grenier & Weatherbee 2005[stránka potřebná ]
- ^ Harrison, Paul M .; Gerstein, Mark (17. května 2002). „Studium genů v průběhu věků: rodiny proteinů, pseudogeny a evoluce proteinů“. Journal of Molecular Biology. 318 (5): 1155–1174. doi:10.1016 / S0022-2836 (02) 00109-2. ISSN 0022-2836. PMID 12083509.
- ^ Bowmaker, James K. (květen 1998). "Vývoj barevného vidění u obratlovců". Oko. 12 (3b): 541–547. doi:10.1038 / oko.1998.143. ISSN 0950-222X. PMID 9775215. S2CID 12851209.
- ^ Gregory, T. Ryan; Hebert, Paul D. N. (Duben 1999). „Modulace obsahu DNA: přibližné příčiny a konečné důsledky“. Výzkum genomu. 9 (4): 317–324. doi:10,1101 / gr. 9.4.317 (neaktivní 2020-11-18). ISSN 1088-9051. PMID 10207154. Archivováno z původního dne 2014-08-23. Citováno 2014-12-11.CS1 maint: DOI neaktivní od listopadu 2020 (odkaz)
- ^ Hurles, Matthew (13. července 2004). „Duplikace genů: Genomický obchod s náhradními díly“. PLOS Biology. 2 (7): e206. doi:10.1371 / journal.pbio.0020206. ISSN 1545-7885. PMC 449868. PMID 15252449.
- ^ Liu, Na; Okamura, Katsutomo; Tyler, David M .; et al. (Říjen 2008). „Vývoj a funkční diverzifikace genů zvířecích mikroRNA“. Cell Research. 18 (10): 985–996. doi:10.1038 / cr.2008.278. ISSN 1001-0602. PMC 2712117. PMID 18711447.
- ^ Siepel, Adam (Říjen 2009). „Darwinova alchymie: lidské geny z nekódující DNA“. Výzkum genomu. 19 (10): 1693–1695. doi:10.1101 / gr.098376.109. ISSN 1088-9051. PMC 2765273. PMID 19797681.
- ^ Orengo, Christine A .; Thornton, Janet M. (Červenec 2005). „Proteinové rodiny a jejich vývoj - strukturální perspektiva“. Roční přehled biochemie. Výroční recenze. 74: 867–900. doi:10,1146 / annurev.biochem.74.082803.133029. ISSN 0066-4154. PMID 15954844. S2CID 7483470.
- ^ Long, Manyuan; Betrán, Esther; Thornton, Kevin; Wang, Wen (listopad 2003). "Původ nových genů: záblesky od mladých i starých". Genetika hodnocení přírody. 4 (11): 865–875. doi:10.1038 / nrg1204. ISSN 1471-0056. PMID 14634634. S2CID 33999892.
- ^ Wang, Minglei; Caetano-Anollés, Gustavo (14. ledna 2009). „Evoluční mechanika organizace domény v proteinech a vzestup modularity v proteinovém světě“. Struktura. 17 (1): 66–78. doi:10.1016 / j.str.2008.11.008. ISSN 1357-4310. PMID 19141283.
- ^ Weissman, Kira J .; Müller, Rolf (14. dubna 2008). „Interakce protein – protein v multienzymových megasyntetázách“. ChemBioChem. 9 (6): 826–848. doi:10.1002 / cbic.200700751. ISSN 1439-4227. PMID 18357594. S2CID 205552778.
- ^ Radding, Charles M. (prosinec 1982). "Homologní párování a výměna pramenů v genetické rekombinaci". Výroční přehled genetiky. 16: 405–437. doi:10.1146 / annurev.ge.16.120182.002201. ISSN 0066-4197. PMID 6297377.
- ^ Agrawal, Aneil F. (5. září 2006). „Evoluce sexu: Proč organizmy zaměňují své genotypy?“. Aktuální biologie. 16 (17): R696 – R704. CiteSeerX 10.1.1.475.9645. doi:10.1016 / j.cub.2006.07.063. ISSN 0960-9822. PMID 16950096. S2CID 14739487.
- ^ Peters, Andrew D .; Otto, Sarah P. (červen 2003). "Osvobození genetické odchylky pohlavím" BioEssays. 25 (6): 533–537. doi:10.1002 / bies.10291. ISSN 0265-9247. PMID 12766942.
- ^ Goddard, Matthew R .; Godfray, H. Charles J.; Burt, Austin (31. března 2005). "Pohlaví zvyšuje účinnost přirozeného výběru v experimentálních populacích kvasinek". Příroda. 434 (7033): 636–640. Bibcode:2005 Natur.434..636G. doi:10.1038 / nature03405. ISSN 0028-0836. PMID 15800622. S2CID 4397491.
- ^ Maynard Smith 1978[stránka potřebná ]
- ^ A b Ridley 1993[stránka potřebná ]
- ^ Van Valen, Leigh (1973). „Nový evoluční zákon“ (PDF). Evoluční teorie. 1: 1–30. ISSN 0093-4755. Archivovány od originál (PDF) dne 2014-12-22. Citováno 2014-12-24.
- ^ Hamilton, W. D.; Axelrod, Robert; Tanese, Reiko (1. května 1990). „Sexuální reprodukce jako adaptace na odolnost vůči parazitům (recenze)“. Proc. Natl. Acad. Sci. USA. 87 (9): 3566–3573. Bibcode:1990PNAS ... 87.3566H. doi:10.1073 / pnas.87.9.3566. ISSN 0027-8424. PMC 53943. PMID 2185476.
- ^ Birdsell & Wills 2003, str. 113–117
- ^ A b Morjan, Carrie L .; Rieseberg, Loren H. (Červen 2004). „Jak se druhy vyvíjejí kolektivně: důsledky toku genů a selekce pro šíření výhodných alel“. Molekulární ekologie. 13 (6): 1341–1356. doi:10.1111 / j.1365-294X.2004.02164.x. ISSN 0962-1083. PMC 2600545. PMID 15140081.
- ^ Boucher, Yan; Douady, Christophe J .; Papke, R. Thane; et al. (Prosinec 2003). "Laterální přenos genů a počátky prokaryotických skupin". Výroční přehled genetiky. 37: 283–328. doi:10.1146 / annurev.genet.37.050503.084247. ISSN 0066-4197. PMID 14616063.
- ^ Walsh, Timothy R. (říjen 2006). "Kombinatorická genetická evoluce multirezistence". Současný názor v mikrobiologii. 9 (5): 476–482. doi:10.1016 / j.mib.2006.08.009. ISSN 1369-5274. PMID 16942901.
- ^ Kondo, Natsuko; Nikoh, Naruo; Ijichi, Nobuyuki; et al. (29. října 2002). "Fragment genomu z Wolbachia endosymbiont přenesen do X chromozomu hostitelského hmyzu ". Proc. Natl. Acad. Sci. USA. 99 (22): 14280–14285. Bibcode:2002PNAS ... 9914280K. doi:10.1073 / pnas.222228199. ISSN 0027-8424. PMC 137875. PMID 12386340.
- ^ Sprague, George F., Jr. (prosinec 1991). "Genetická výměna mezi královstvími". Aktuální názor na genetiku a vývoj. 1 (4): 530–533. doi:10.1016 / S0959-437X (05) 80203-5. ISSN 0959-437X. PMID 1822285.
- ^ Gladyshev, Eugene A .; Meselson, Matthew; Arkhipova, Irina R. (30. května 2008). "Masivní horizontální přenos genů v bdelloidních vířičích". Věda. 320 (5880): 1210–1213. Bibcode:2008Sci ... 320.1210G. doi:10.1126 / science.1156407. ISSN 0036-8075. PMID 18511688. S2CID 11862013.
- ^ Baldo, Angela M .; McClure, Marcella A. (září 1999). „Evoluce a horizontální přenos genů kódujících dUTPázu ve virech a jejich hostitelích“. Journal of Virology. 73 (9): 7710–7721. doi:10.1128 / JVI.73.9.7710-7721.1999. ISSN 0022-538X. PMC 104298. PMID 10438861.
- ^ Rivera, Maria C .; Lake, James A. (9. září 2004). „Kruh života poskytuje důkazy o původu genomové fúze eukaryot“. Příroda. 431 (7005): 152–155. Bibcode:2004 Natur.431..152R. doi:10.1038 / nature02848. ISSN 0028-0836. PMID 15356622. S2CID 4349149.
- ^ A b C Hurst, Laurence D. (Únor 2009). „Základní pojmy v genetice: genetika a porozumění výběru“. Genetika hodnocení přírody. 10 (2): 83–93. doi:10.1038 / nrg2506. ISSN 1471-0056. PMID 19119264. S2CID 1670587.
- ^ Darwin 1859, Kapitola XIV
- ^ Otto, Sarah P.; Servedio, Maria R.; Nuismer, Scott L. (srpen 2008). „Výběr závislý na frekvenci a vývoj střídavého páření“. Genetika. 179 (4): 2091–2112. doi:10.1534 / genetika.107.084418. ISSN 0016-6731. PMC 2516082. PMID 18660541.
- ^ A b C Orr, H. Allen (Srpen 2009). „Fitness a její role v evoluční genetice“. Genetika hodnocení přírody. 10 (8): 531–539. doi:10.1038 / nrg2603. ISSN 1471-0056. PMC 2753274. PMID 19546856.
- ^ Haldane, J.B.S. (14. března 1959). „Teorie přirozeného výběru dodnes“. Příroda. 183 (4663): 710–713. Bibcode:1959 Natur.183..710H. doi:10.1038 / 183710a0. ISSN 0028-0836. PMID 13644170. S2CID 4185793.
- ^ Lande, Russell; Arnold, Stevan J. (listopad 1983). "Měření výběru u korelovaných znaků". Vývoj. 37 (6): 1210–1226. doi:10.1111 / j.1558-5646.1983.tb00236.x. ISSN 0014-3820. JSTOR 2408842. PMID 28556011. S2CID 36544045.
- ^ Goldberg, Emma E .; Igić, Boris (listopad 2008). "Na fylogenetické testy nevratného vývoje". Vývoj. 62 (11): 2727–2741. doi:10.1111 / j.1558-5646.2008.00505.x. ISSN 0014-3820. PMID 18764918. S2CID 30703407.
- ^ Collin, Rachel; Miglietta, Maria Pia (listopad 2008). „Obrácení názorů na Dollov zákon“. Trendy v ekologii a evoluci. 23 (11): 602–609. doi:10.1016 / j.tree.2008.06.013. ISSN 0169-5347. PMID 18814933.
- ^ Tomić, Nenad; Meyer-Rochow, Victor Benno (2011). "Atavismy: Lékařské, genetické a evoluční důsledky". Perspektivy v biologii a medicíně. 54 (3): 332–353. doi:10.1353 / pbm.2011.0034. ISSN 0031-5982. PMID 21857125. S2CID 40851098.
- ^ Hoekstra, Hopi E .; Hoekstra, Jonathan M .; Berrigan, David; et al. (31. července 2001). „Síla a tempo směrového výběru ve volné přírodě“. Proc. Natl. Acad. Sci. USA. 98 (16): 9157–9160. Bibcode:2001PNAS ... 98,9157H. doi:10.1073 / pnas.161281098. ISSN 0027-8424. PMC 55389. PMID 11470913.
- ^ Felsenstein, Joseph (Listopad 1979). "Výlety podél rozhraní mezi rušivým a stabilizujícím výběrem". Genetika. 93 (3): 773–795. ISSN 0016-6731. PMC 1214112. PMID 17248980.
- ^ Odum 1971, str. 8
- ^ Okasha 2006
- ^ A b Gould, Stephen Jay (28. února 1998). „Gulliverovy další cesty: nutnost a obtížnost hierarchické teorie výběru“. Filozofické transakce královské společnosti B. 353 (1366): 307–314. doi:10.1098 / rstb.1998.0211. ISSN 0962-8436. PMC 1692213. PMID 9533127.
- ^ Mayr, Ernst (18. března 1997). "Objekty výběru". Proc. Natl. Acad. Sci. USA. 94 (6): 2091–2094. Bibcode:1997PNAS ... 94.2091M. doi:10.1073 / pnas.94.6.2091. ISSN 0027-8424. PMC 33654. PMID 9122151.
- ^ Maynard Smith 1998, str. 203–211; diskuse 211–217
- ^ Hickey, Donal A. (1992). "Evoluční dynamika transponovatelných prvků v prokaryotech a eukaryotech". Genetica. 86 (1–3): 269–274. doi:10.1007 / BF00133725. ISSN 0016-6707. PMID 1334911. S2CID 6583945.
- ^ Gould, Stephen Jay; Lloyd, Elisabeth A. (12. října 1999). „Individualita a adaptace na různých úrovních výběru: jak bychom mohli pojmenovat a zobecnit jednotku darwinismu?“. Proc. Natl. Acad. Sci. USA. 96 (21): 11904–11909. Bibcode:1999PNAS ... 9611904G. doi:10.1073 / pnas.96.21.11904. ISSN 0027-8424. PMC 18385. PMID 10518549.
- ^ Lien, Sigbjørn; Szyda, Joanna; Schechinger, Birgit; et al. (Únor 2000). „Důkaz heterogenity v rekombinaci v lidské pseudoautosomální oblasti: analýza s vysokým rozlišením pomocí psaní spermií a radiačně hybridního mapování“. American Journal of Human Genetics. 66 (2): 557–566. doi:10.1086/302754. ISSN 0002-9297. PMC 1288109. PMID 10677316.
- ^ Barton, Nicholas H. (29. listopadu 2000). „Genetické stopování“. Filozofické transakce královské společnosti B. 355 (1403): 1553–1562. doi:10.1098 / rstb.2000.0716. ISSN 0962-8436. PMC 1692896. PMID 11127900.
- ^ A b Gillespie, John H. (Listopad 2001). „Je velikost populace druhu relevantní pro jeho vývoj?“. Vývoj. 55 (11): 2161–2169. doi:10.1111 / j.0014-3820.2001.tb00732.x. ISSN 0014-3820. PMID 11794777. S2CID 221735887.
- ^ Andersson, Malte; Simmons, Leigh W. (červen 2006). „Sexuální výběr a volba partnera“ (PDF). Trendy v ekologii a evoluci. 21 (6): 296–302. CiteSeerX 10.1.1.595.4050. doi:10.1016 / j.tree.2006.03.015. ISSN 0169-5347. PMID 16769428. Archivováno (PDF) z původního dne 2013-03-09.
- ^ Kokko, Hanna; Brooks, Robert; McNamara, John M .; Houston, Alasdair I. (7. července 2002). „Kontinuum sexuálního výběru“. Sborník královské společnosti B. 269 (1498): 1331–1340. doi:10.1098 / rspb.2002.2020. ISSN 0962-8452. PMC 1691039. PMID 12079655.
- ^ Quinn, Thomas P .; Hendry, Andrew P .; Buck, Gregory B. (2001). „Vyrovnávání přirozené a sexuální selekce u lososa sockeye: interakce mezi velikostí těla, reprodukční příležitostí a náchylností k predaci medvědy“ (PDF). Evoluční ekologický výzkum. 3: 917–937. ISSN 1522-0613. Archivováno (PDF) z původního dne 2016-03-05. Citováno 2014-12-15.
- ^ Hunt, John; Brooks, Robert; Jennions, Michael D .; et al. (23. prosince 2004). „Vysoce kvalitní cvrčci mužského terénu hodně investují do sexuálního projevu, ale umírají mladí“. Příroda. 432 (7020): 1024–1027. Bibcode:2004 Natur.432.1024H. doi:10.1038 / nature03084. ISSN 0028-0836. PMID 15616562. S2CID 4417867.
- ^ Futuyma & Kirkpatrick 2017, str. 55–66, kapitola 3: Přirozený výběr a přizpůsobení
- ^ A b Masel, Joanna (25. října 2011). "Genetický drift". Aktuální biologie. 21 (20): R837 – R838. doi:10.1016 / j.cub.2011.08.007. ISSN 0960-9822. PMID 22032182. S2CID 17619958.
- ^ Lande, Russell (1989). „Rybářské a Wrightovské teorie speciace“. Genom. 31 (1): 221–227. doi:10,1139 / g89-037. ISSN 0831-2796. PMID 2687093.
- ^ Kimura, Motoo (1991). „Neutrální teorie molekulární evoluce: přehled nedávných důkazů“. Japonský žurnál lidské genetiky. 66 (4): 367–386. doi:10,1266 / jjg.66.367. ISSN 0021-504X. PMID 1954033.
- ^ Kimura, Motoo (1989). „Neutrální teorie molekulární evoluce a světový názor neutralistů“. Genom. 31 (1): 24–31. doi:10.1139 / g89-009. ISSN 0831-2796. PMID 2687096.
- ^ Kreitman, Martin (Srpen 1996). „Neutrální teorie je mrtvá. Ať žije neutrální teorie.“ BioEssays. 18 (8): 678–683, diskuse 683. doi:10,1002 / bies. 950180812. ISSN 0265-9247. PMID 8760341.
- ^ Leigh, E.G., Jr. (listopad 2007). „Neutrální teorie: historická perspektiva“. Journal of Evolutionary Biology. 20 (6): 2075–2091. doi:10.1111 / j.1420-9101.2007.01410.x. ISSN 1010-061X. PMID 17956380. S2CID 2081042.
- ^ Neher, Richard A .; Shraiman, Boris I. (srpen 2011). „Genetický koncept a kvazi-neutralita u velkých fakultativně sexuálních populací“. Genetika. 188 (4): 975–996. arXiv:1108.1635. Bibcode:2011arXiv1108.1635N. doi:10.1534 / genetika.111.128876. ISSN 0016-6731. PMC 3176096. PMID 21625002.
- ^ Otto, Sarah P .; Whitlock, Michael C. (červen 1997). „Pravděpodobnost fixace u populací se změnou velikosti“ (PDF). Genetika. 146 (2): 723–733. ISSN 0016-6731. PMC 1208011. PMID 9178020. Archivováno (PDF) z původního dne 2015-03-19. Citováno 2014-12-18.
- ^ A b Charlesworth, Brian (Březen 2009). „Základní pojmy v genetice: efektivní velikost populace a vzorce molekulárního vývoje a variací“. Genetika hodnocení přírody. 10 (3): 195–205. doi:10.1038 / nrg2526. ISSN 1471-0056. PMID 19204717. S2CID 205484393.
- ^ Cutter, Asher D .; Choi, Jae Young (srpen 2010). „Přirozený výběr formuje polymorfismus nukleotidů napříč genomem hlístice Caenorhabditis briggsae". Výzkum genomu. 20 (8): 1103–1111. doi:10.1101 / gr. 104331.109. ISSN 1088-9051. PMC 2909573. PMID 20508143.
- ^ Mitchell-Olds, Thomas; Willis, John H .; Goldstein, David B. (Listopad 2007). „Které evoluční procesy ovlivňují přirozenou genetickou variabilitu fenotypových vlastností?“. Genetika hodnocení přírody. 8 (11): 845–856. doi:10.1038 / nrg2207. ISSN 1471-0056. PMID 17943192. S2CID 14914998.
- ^ Nei, Masatoshi (Prosinec 2005). „Selekcionismus a neutralismus v molekulární evoluci“. Molekulární biologie a evoluce. 22 (12): 2318–2342. doi:10.1093 / molbev / msi242. ISSN 0737-4038. PMC 1513187. PMID 16120807.
- Nei, Masatoshi (květen 2006). „Selekcionismus a neutralismus v molekulární evoluci“. Molekulární biologie a evoluce (Tisková chyba). 23 (5): 2318–42. doi:10,1093 / molbev / msk009. ISSN 0737-4038. PMC 1513187. PMID 16120807.
- ^ Wright, Sewall (1932). „Role mutace, křížení, křížení a selekce v evoluci“. Sborník z VI. Mezinárodního kongresu genetiky. 1: 356–366. Archivováno z původního dne 2014-08-23. Citováno 2014-12-18.
- ^ Coyne, Jerry A.; Barton, Nicholas H .; Turelli, Michael (červen 1997). „Perspective: A Critique of Sewall Wright's Shifting Balance Theory of Evolution“. Vývoj. 51 (3): 643–671. doi:10.2307/2411143. ISSN 0014-3820. JSTOR 2411143. PMID 28568586.
- ^ Haldane, J.B.S. (Červenec 1927). „Matematická teorie přirozeného a umělého výběru, část V: výběr a mutace“. Sborník Cambridge Philosophical Society. 26 (7): 838–844. Bibcode:1927PCPS ... 23..838H. doi:10.1017 / S0305004100015644.
- ^ Fisher 1930
- ^ A b Yampolsky, Lev Y .; Stoltzfus, Arlin (20. prosince 2001). "Předpojatost při zavádění variací jako orientačního faktoru v evoluci". Evoluce a vývoj. 3 (2): 73–83. doi:10.1046 / j.1525-142x.2001.003002073.x. PMID 11341676. S2CID 26956345.
- ^ Sueoka, Noboru (1. dubna 1962). „Na genetickém základě variability a heterogenity složení DNA báze“. Proc. Natl. Acad. Sci. USA. 48 (4): 582–592. Bibcode:1962PNAS ... 48..582S. doi:10.1073 / pnas.48.4.582. PMC 220819. PMID 13918161.
- ^ Freese, Ernst (Červenec 1962). „O vývoji základního složení DNA“. Journal of Theoretical Biology. 3 (1): 82–101. doi:10.1016 / S0022-5193 (62) 80005-8.
- ^ Cox, Edward C .; Yanofsky, Charles (1. listopadu 1967). "Změněné základní poměry v DNA kmene mutátoru Escherichia coli". Proc. Natl. Acad. Sci. USA. 58 (5): 1895–1902. Bibcode:1967PNAS ... 58.1895C. doi:10.1073 / pnas.58.5.1895. PMC 223881. PMID 4866980.
- ^ Shah, Premal; Gilchrist, Michael A. (21. června 2011). „Vysvětlení složitých vzorců využití kodonů s výběrem translační účinnosti, zkreslení mutací a genetického driftu“. Proc. Natl. Acad. Sci. USA. 108 (25): 10231–10236. Bibcode:2011PNAS..10810231S. doi:10.1073 / pnas.1016719108. PMC 3121864. PMID 21646514.
- ^ Bulmer, Michael G. (Listopad 1991). „The selection-mutation-drift theory of synonymous codon usage“. Genetika. 129 (3): 897–907. PMC 1204756. PMID 1752426.
- ^ Fryxell, Karl J .; Zuckerkandl, Emile (Září 2000). „Deaminace cytosinem hraje primární roli ve vývoji izoforů savců“. Molekulární biologie a evoluce. 17 (9): 1371–1383. doi:10.1093 / oxfordjournals.molbev.a026420. PMID 10958853.
- ^ Petrov, Dmitrij A .; Sangster, Todd A .; Johnston, J. Spencer; et al. (11. února 2000). „Důkazy o ztrátě DNA jako determinantu velikosti genomu“. Věda. 287 (5455): 1060–1062. Bibcode:2000Sci ... 287.1060P. doi:10.1126 / science.287.5455.1060. ISSN 0036-8075. PMID 10669421. S2CID 12021662.
- ^ Petrov, Dmitri A. (květen 2002). "Ztráta DNA a vývoj velikosti genomu ve Windows Drosophila". Genetica. 115 (1): 81–91. doi:10.1023 / A: 1016076215168. ISSN 0016-6707. PMID 12188050. S2CID 5314242.
- ^ Duret, Laurent; Galtier, Nicolas (září 2009). „Předpjatá genová konverze a vývoj savčích genomických krajin“. Roční přehled genomiky a lidské genetiky. Výroční recenze. 10: 285–311. doi:10.1146 / annurev-genom-082908-150001. ISSN 1527-8204. PMID 19630562. S2CID 9126286.
- ^ Hershberg, Ruth; Petrov, Dmitrij A. (9. září 2010). „Důkazy, že mutace je u bakterií univerzálně předpojatá vůči AT“. Genetika PLOS. 6 (9): e1001115. doi:10.1371 / journal.pgen.1001115. PMC 2936535. PMID 20838599.
- ^ Stoltzfus, Arlin; McCandlish, David M. (září 2017). „Mutační předsudky ovlivňují paralelní adaptaci“. Molekulární biologie a evoluce. 34 (9): 2163–2172. doi:10,1093 / molbev / msx180. PMC 5850294. PMID 28645195.
- ^ Payne, Joshua L .; Menardo, Fabrizio; Trauner, Andrej; et al. (13. května 2019). "Přechodová odchylka ovlivňuje vývoj rezistence na antibiotika v Mycobacterium tuberculosis". PLOS Biology. 17 (5): e3000265. doi:10.1371 / journal.pbio.3000265. PMC 6532934. PMID 31083647.
- ^ Storz, Jay F .; Natarajan, Chandrasekhar; Signore, Anthony V .; et al. (22. července 2019). „Úloha zkreslení mutací v adaptivní molekulární evoluci: poznatky z konvergentních změn ve funkci proteinu“. Filozofické transakce královské společnosti B. 374 (1777): 20180238. doi:10.1098 / rstb.2018.0238. PMC 6560279. PMID 31154983.
- ^ Svensson, Erik I .; Berger, David (1. května 2019). „Úloha zkreslení mutací v adaptivní evoluci“. Trendy v ekologii a evoluci. 34 (5): 422–434. doi:10.1016 / j.strom.2019.01.015. PMID 31003616.
- ^ Baym, Michael; Lieberman, Tami D .; Kelsic, Eric D .; et al. (9. září 2016). „Spatiotemporální mikrobiální evoluce na antibiotické krajině“. Věda. 353 (6304): 1147–1151. Bibcode:2016Sci ... 353.1147B. doi:10.1126 / science.aag0822. ISSN 0036-8075. PMC 5534434. PMID 27609891.
- ^ A b C Scott, Eugenie C.; Matzke, Nicholas J. (15. května 2007). „Biologický design ve vědeckých učebnách“. Proc. Natl. Acad. Sci. USA. 104 (Příloha 1): 8669–8676. Bibcode:2007PNAS..104 8669S. doi:10.1073 / pnas.0701505104. ISSN 0027-8424. PMC 1876445. PMID 17494747.
- ^ Hendry, Andrew Paul; Kinnison, Michael T. (listopad 2001). "Úvod do mikroevoluce: rychlost, vzor, proces". Genetica. 112–113 (1): 1–8. doi:10.1023 / A: 1013368628607. ISSN 0016-6707. PMID 11838760. S2CID 24485535.
- ^ Leroi, Armand M. (Březen – duben 2000). "Rozsahová nezávislost evoluce". Evoluce a vývoj. 2 (2): 67–77. CiteSeerX 10.1.1.120.1020. doi:10.1046 / j.1525-142x.2000.00044.x. ISSN 1520-541X. PMID 11258392. S2CID 17289010.
- ^ Gould 2002, str. 657–658.
- ^ A b Gould, Stephen Jay (19. července 1994). „Tempo a režim v makroevoluční rekonstrukci darwinismu“. Proc. Natl. Acad. Sci. USA. 91 (15): 6764–6771. Bibcode:1994PNAS ... 91.6764G. doi:10.1073 / pnas.91.15.6764. ISSN 0027-8424. PMC 44281. PMID 8041695.
- ^ Jablonski, David (2000). „Mikroevoluce a makroevoluce: měřítko a hierarchie v evoluční biologii a paleobiologii“. Paleobiologie. 26 (sp4): 15–52. doi:10.1666 / 0094-8373 (2000) 26 [15: MAMSAH] 2.0.CO; 2. ISSN 0094-8373.
- ^ Dougherty, Michael J. (20. července 1998). „Vyvíjí se nebo se mění lidská rasa?“. Scientific American. ISSN 0036-8733. Archivováno z původního dne 2014-05-06. Citováno 2015-09-11.
- ^ Isaak, Mark, ed. (22. července 2003). „Claim CB932: Evolution of degenerate forms“. Archiv TalkOrigins. Houston, Texas: The TalkOrigins Foundation, Inc. Archivováno z původního dne 2014-08-23. Citováno 2014-12-19.
- ^ Lane 1996, str. 61
- ^ Carroll, Sean B. (22. února 2001). „Šance a nutnost: vývoj morfologické složitosti a rozmanitosti“. Příroda. 409 (6823): 1102–1109. Bibcode:2001 Natur.409.1102C. doi:10.1038/35059227. ISSN 0028-0836. PMID 11234024. S2CID 4319886.
- ^ Whitman, William B .; Coleman, David C .; Wiebe, William J. (9. června 1998). „Prokaryotes: neviditelná většina“. Proc. Natl. Acad. Sci. USA. 95 (12): 6578–6583. Bibcode:1998PNAS ... 95,6578W. doi:10.1073 / pnas.95.12.6578. ISSN 0027-8424. PMC 33863. PMID 9618454.
- ^ A b Schloss, Patrick D .; Handelsman, Jo (Prosinec 2004). „Stav mikrobiálního sčítání“. Recenze mikrobiologie a molekulární biologie. 68 (4): 686–691. doi:10.1128 / MMBR.68.4.686-691.2004. ISSN 1092-2172. PMC 539005. PMID 15590780.
- ^ Nealson, Kenneth H. (leden 1999). „Postvikingská mikrobiologie: nové přístupy, nová data, nové poznatky“. Počátky života a vývoj biosfér. 29 (1): 73–93. Bibcode:1999OLEB ... 29 ... 73N. doi:10.1023 / A: 1006515817767. ISSN 0169-6149. PMID 11536899. S2CID 12289639.
- ^ Vzpěr, Angus; MacLean, R. Craig; Brockhurst, Michael A .; Colegrave, Nick (12. února 2009). „Bígl v láhvi“. Příroda. 457 (7231): 824–829. Bibcode:2009 Natur.457..824B. doi:10.1038 / nature07892. ISSN 0028-0836. PMID 19212400. S2CID 205216404.
- ^ Elena, Santiago F .; Lenski, Richard E. (Červen 2003). "Evoluční experimenty s mikroorganismy: dynamika a genetické základy adaptace". Genetika hodnocení přírody. 4 (6): 457–469. doi:10.1038 / nrg1088. ISSN 1471-0056. PMID 12776215. S2CID 209727.
- ^ Mayr 1982, str. 483: „Adaptace ... již nemohla být považována za statický stav, produkt tvůrčí minulosti a místo toho se stala pokračujícím dynamickým procesem.“
- ^ Šesté vydání Oxfordský slovník vědy (2010) definuje přizpůsobování jako „Jakákoli změna ve struktuře nebo fungování po sobě jdoucích generací populace, díky nimž se lépe hodí pro své prostředí.“
- ^ Orr, H. Allen (únor 2005). "Genetická teorie adaptace: krátká historie". Genetika hodnocení přírody. 6 (2): 119–127. doi:10.1038 / nrg1523. ISSN 1471-0056. PMID 15716908. S2CID 17772950.
- ^ Dobžanský 1968, s. 1–34
- ^ Dobžanský 1970, s. 4–6, 79–82, 84–87
- ^ Dobžanský, Theodosius (březen 1956). "Genetika přírodních populací. XXV. Genetické změny v populacích Drosophila pseudoobscura a Drosophila persimilis v některých lokalitách v Kalifornii “. Vývoj. 10 (1): 82–92. doi:10.2307/2406099. ISSN 0014-3820. JSTOR 2406099.
- ^ Nakajima, Akira; Sugimoto, Yohko; Yoneyama, Hiroši; Nakae, Taiji (červen 2002). „Rezistence na fluorochinolony na vysoké úrovni v Pseudomonas aeruginosa Kvůli souhře efluxní pumpy MexAB-OprM a mutaci DNA gyrázy “. Mikrobiologie a imunologie. 46 (6): 391–395. doi:10.1111 / j.1348-0421.2002.tb02711.x. ISSN 1348-0421. PMID 12153116. S2CID 22593331.
- ^ Blount, Zachary D .; Borland, Christina Z .; Lenski, Richard E. (10. června 2008). „Zahajovací článek: Historická událost a vývoj klíčové inovace v experimentální populaci Escherichia coli". Proc. Natl. Acad. Sci. USA. 105 (23): 7899–7906. Bibcode:2008PNAS..105,7899B. doi:10.1073 / pnas.0803151105. ISSN 0027-8424. PMC 2430337. PMID 18524956.
- ^ Okada, Hirosuke; Negoro, Seiji; Kimura, Hiroyuki; Nakamura, Shunichi (10. listopadu 1983). "Evoluční adaptace plazmidem kódovaných enzymů pro degradaci nylonových oligomerů". Příroda. 306 (5939): 203–206. Bibcode:1983 Natur.306..203O. doi:10.1038 / 306203a0. ISSN 0028-0836. PMID 6646204. S2CID 4364682.
- ^ Ach, Susumu (Duben 1984). „Zrození jedinečného enzymu z alternativního čtecího rámce již existující interně se opakující kódovací sekvence“. Proc. Natl. Acad. Sci. USA. 81 (8): 2421–2425. Bibcode:1984PNAS ... 81.2421O. doi:10.1073 / pnas.81.8.2421. ISSN 0027-8424. PMC 345072. PMID 6585807.
- ^ Copley, Shelley D. (červen 2000). „Vývoj metabolické cesty pro degradaci toxického xenobiotika: patchworkový přístup“. Trendy v biochemických vědách. 25 (6): 261–265. doi:10.1016 / S0968-0004 (00) 01562-0. ISSN 0968-0004. PMID 10838562.
- ^ Crawford, Ronald L .; Jung, Carina M .; Popruh, Janice L. (říjen 2007). „Nedávný vývoj pentachlorfenolu (PCP) -4-monooxygenázy (PcpB) a související cesty bakteriální degradace PCP“. Biologický rozklad. 18 (5): 525–539. doi:10.1007 / s10532-006-9090-6. ISSN 0923-9820. PMID 17123025. S2CID 8174462.
- ^ Eshel, Ilan (prosinec 1973). „Výběr klonů a optimální rychlost mutace“. Journal of Applied Probability. 10 (4): 728–738. doi:10.2307/3212376. ISSN 1475-6072. JSTOR 3212376.
- ^ Altenberg 1995, str. 205–259
- ^ Masel, Joanna; Bergman, Aviv (červenec 2003). "Vývoj vlastností evoluční schopnosti prionu kvasinek [PSI +]". Vývoj. 57 (7): 1498–1512. doi:10.1111 / j.0014-3820.2003.tb00358.x. ISSN 0014-3820. PMID 12940355. S2CID 30954684.
- ^ Lancaster, Alex K .; Bardill, J. Patrick; Pravda, Heather L .; Masel, Joanna (únor 2010). „Míra spontánního vzhledu kvasinkového prionu [PSI+] a jeho důsledky pro vývoj vlastností evolučnosti [PSI+] Systém ". Genetika. 184 (2): 393–400. doi:10.1534 / genetika.109.110213. ISSN 0016-6731. PMC 2828720. PMID 19917766.
- ^ Draghi, Jeremy; Wagner, Günter P. (Únor 2008). "Vývoj evolučnosti ve vývojovém modelu". Vývoj. 62 (2): 301–315. doi:10.1111 / j.1558-5646.2007.00303.x. ISSN 0014-3820. PMID 18031304. S2CID 11560256.
- ^ A b Bejder, Lars; Hall, Brian K. (Listopad 2002). "Končetiny u velryb a bezbrannost u jiných obratlovců: mechanismy evoluční a vývojové transformace a ztráty". Evoluce a vývoj. 4 (6): 445–458. doi:10.1046 / j.1525-142X.2002.02033.x. ISSN 1520-541X. PMID 12492145. S2CID 8448387.
- ^ Young, Nathan M .; HallgrÍmsson, Benedikt (prosinec 2005). "Sériová homologie a vývoj kovarianční struktury končetin savců". Vývoj. 59 (12): 2691–2704. doi:10.1554/05-233.1. ISSN 0014-3820. PMID 16526515. S2CID 198156135.
- ^ A b Penny, David; Poole, Anthony (prosinec 1999). "Povaha posledního univerzálního společného předka". Aktuální názor na genetiku a vývoj. 9 (6): 672–677. doi:10.1016 / S0959-437X (99) 00020-9. ISSN 0959-437X. PMID 10607605.
- ^ Hall, Brian K. (srpen 2003). "Sestup s modifikací: jednota, která je základem homologie a homoplasie, jak je vidět prostřednictvím analýzy vývoje a vývoje". Biologické recenze. 78 (3): 409–433. doi:10.1017 / S1464793102006097. ISSN 1464-7931. PMID 14558591. S2CID 22142786.
- ^ Shubin, Neil; Tabin, Clifford J.; Carroll, Sean B. (12. února 2009). „Hluboká homologie a počátky evoluční novinky“. Příroda. 457 (7231): 818–823. Bibcode:2009Natur.457..818S. doi:10.1038 / nature07891. ISSN 0028-0836. PMID 19212399. S2CID 205216390.
- ^ A b C Fong, Daniel F .; Kane, Thomas C .; Culver, David C. (listopad 1995). "Zbytky a ztráta nefunkčních znaků". Výroční přehled ekologie a systematiky. 26: 249–268. doi:10.1146 / annurev.es.26.110195.001341. ISSN 1545-2069.
- ^ ZhaoLei Zhang; Gerstein, Mark (srpen 2004). "Rozsáhlá analýza pseudogenů v lidském genomu". Aktuální názor na genetiku a vývoj. 14 (4): 328–335. doi:10.1016 / j.gde.2004.06.003. ISSN 0959-437X. PMID 15261647.
- ^ Jeffery, William R. (květen – červen 2005). „Adaptivní vývoj degenerace očí u mexických slepých jeskyní“. Journal of Heredity. 96 (3): 185–196. CiteSeerX 10.1.1.572.6605. doi:10.1093 / jhered / esi028. ISSN 0022-1503. PMID 15653557.
- ^ Maxwell, Erin E .; Larsson, Hans C.E. (květen 2007). „Osteologie a myologie křídla Emu (Dromaius novaehollandiae) a jeho vliv na vývoj zakrnělých struktur “. Journal of Morphology. 268 (5): 423–441. doi:10,1002 / jmor.10527. ISSN 0362-2525. PMID 17390336. S2CID 12494187.
- ^ van der Kooi, Casper J .; Schwander, Tanja (listopad 2014). „O osudu sexuálních rysů pod asexualitou“ (PDF). Biologické recenze. 89 (4): 805–819. doi:10.1111 / brv.12078. ISSN 1464-7931. PMID 24443922. S2CID 33644494. Archivováno (PDF) z původního dne 2015-07-23. Citováno 2015-08-05.
- ^ Silvestri, Anthony R., Jr.; Singh, Iqbal (duben 2003). „Nevyřešený problém třetího stoličky: Bylo by bez něj lidem lépe?“. Journal of the American Dental Association. 134 (4): 450–455. doi:10.14219 / jada.archive.2003.0194. ISSN 0002-8177. PMID 12733778. Archivovány od originál dne 2014-08-23.
- ^ Coyne 2009, str. 62
- ^ Darwin 1872 101, 103
- ^ Gray 2007, str. 66
- ^ Coyne 2009, str. 85–86
- ^ Stevens 1982, str. 87
- ^ A b Gould 2002, str. 1235–1236.
- ^ Pallen, Mark J .; Matzke, Nicholas J. (říjen 2006). "Z Původ druhů k původu bakteriálních bičíků " (PDF). Příroda Recenze Mikrobiologie (PDF). 4 (10): 784–790. doi:10.1038 / nrmicro1493. ISSN 1740-1526. PMID 16953248. S2CID 24057949. Archivovány od originál (PDF) dne 2014-12-26. Citováno 2014-12-25.
- ^ Clements, Abigail; Bursac, Dejan; Gatsos, Xenia; et al. (15. září 2009). „Redukovatelná složitost mitochondriálního molekulárního stroje“. Proc. Natl. Acad. Sci. USA. 106 (37): 15791–15795. Bibcode:2009PNAS..10615791C. doi:10.1073 / pnas.0908264106. ISSN 0027-8424. PMC 2747197. PMID 19717453.
- ^ Piatigorsky a kol. 1994, str. 241–250
- ^ Wistow, Graeme (srpen 1993). "Čočky krystalinů: nábor genů a evoluční dynamika". Trendy v biochemických vědách. 18 (8): 301–306. doi:10.1016 / 0968-0004 (93) 90041-K. ISSN 0968-0004. PMID 8236445.
- ^ Johnson, Norman A .; Porter, Adam H. (listopad 2001). „Směrem k nové syntéze: populační genetika a evoluční vývojová biologie“. Genetica. 112–113 (1): 45–58. doi:10.1023 / A: 1013371201773. ISSN 0016-6707. PMID 11838782. S2CID 1651351.
- ^ Baguñà, Jaume; Garcia-Fernàndez, Jordi (2003). „Evo-Devo: dlouhá a klikatá cesta“. International Journal of Developmental Biology. 47 (7–8): 705–713. ISSN 0214-6282. PMID 14756346. Archivováno od originálu 2014-11-28.
- ^ Allin, Edgar F. (prosinec 1975). "Vývoj středního ucha savců". Journal of Morphology. 147 (4): 403–437. doi:10.1002 / jmor.1051470404. ISSN 0362-2525. PMID 1202224. S2CID 25886311.
- ^ Harris, Matthew P .; Hasso, Sean M .; Ferguson, Mark W.J .; Fallon, John F. (21. února 2006). „Vývoj zubů první generace archosauriánů u kuřecího mutanta“. Aktuální biologie. 16 (4): 371–377. doi:10.1016 / j.cub.2005.12.047. ISSN 0960-9822. PMID 16488870. S2CID 15733491.
- ^ Carroll, Sean B. (11. července 2008). „Evo-Devo a rozšiřující se evoluční syntéza: genetická teorie morfologické evoluce“. Buňka. 134 (1): 25–36. doi:10.1016 / j.cell.2008.06.030. ISSN 0092-8674. PMID 18614008. S2CID 2513041.
- ^ Wade, Michael J. (Březen 2007). „Koevoluční genetika ekologických komunit“. Genetika hodnocení přírody. 8 (3): 185–195. doi:10.1038 / nrg2031. ISSN 1471-0056. PMID 17279094. S2CID 36705246.
- ^ Geffeney, Shana; Brodie, Edmund D., Jr.; Ruben, Peter C .; Brodie, Edmund D., III (23. srpna 2002). „Mechanismy adaptace v závodě zbraní Predator-Prey Arms: sodíkové kanály odolné vůči TTX“. Věda. 297 (5585): 1336–1339. Bibcode:2002Sci ... 297.1336G. doi:10.1126 / science.1074310. ISSN 0036-8075. PMID 12193784. S2CID 8816337.
- Brodie, Edmund D., Jr.; Ridenhour, Benjamin J .; Brodie, Edmund D., III (říjen 2002). „Evoluční reakce predátorů na nebezpečnou kořist: hotspoty a studená místa v geografické mozaice koevoluce mezi podvazkovými hady a mloky“. Vývoj. 56 (10): 2067–2082. doi:10.1554 / 0014-3820 (2002) 056 [2067: teropt] 2.0.co; 2. ISSN 0014-3820. PMID 12449493.
- Carroll, Sean B. (21. prosince 2009). „Cokoliv nezabije některá zvířata, může je učinit smrtícími“. The New York Times. New York: The New York Times Company. ISSN 0362-4331. Archivováno od originálu 2015-04-23. Citováno 2014-12-26.
- ^ Sachs, Joel L. (září 2006). "Spolupráce uvnitř a mezi druhy". Journal of Evolutionary Biology. 19 (5): 1415–1418, diskuse 1426–1436. doi:10.1111 / j.1420-9101.2006.01152.x. ISSN 1010-061X. PMID 16910971. S2CID 4828678.
- Nowak, Martin A. (8. prosince 2006). „Pět pravidel pro vývoj spolupráce“. Věda. 314 (5805): 1560–1563. Bibcode:2006Sci ... 314.1560N. doi:10.1126 / science.1133755. ISSN 0036-8075. PMC 3279745. PMID 17158317.
- ^ Paszkowski, Uta (srpen 2006). „Mutualismus a parazitismus: jin a jang rostlinných symbióz“. Aktuální názor na biologii rostlin. 9 (4): 364–370. doi:10.1016 / j.pbi.2006.05.008. ISSN 1369-5266. PMID 16713732.
- ^ Hause, Bettina; Fester, Thomas (květen 2005). "Molekulární a buněčná biologie arbuskulární mykorhizní symbiózy". Planta. 221 (2): 184–196. doi:10.1007 / s00425-004-1436-x. ISSN 0032-0935. PMID 15871030. S2CID 20082902.
- ^ Bertram, John S. (prosinec 2000). "Molekulární biologie rakoviny". Molekulární aspekty medicíny. 21 (6): 167–223. doi:10.1016 / S0098-2997 (00) 00007-8. ISSN 0098-2997. PMID 11173079.
- ^ Reeve, H. Kern; Hölldobler, Bert (5. června 2007). „Vznik superorganismu prostřednictvím meziskupinové soutěže“. Proc. Natl. Acad. Sci. USA. 104 (23): 9736–9740. Bibcode:2007PNAS..104,9736R. doi:10.1073 / pnas.0703466104. ISSN 0027-8424. PMC 1887545. PMID 17517608.
- ^ Axelrod, Robert; Hamilton, W. D. (27. března 1981). "Vývoj spolupráce". Věda. 211 (4489): 1390–1396. Bibcode:1981Sci ... 211.1390A. doi:10.1126 / science.7466396. ISSN 0036-8075. PMID 7466396.
- ^ Wilson, Edward O .; Hölldobler, Bert (20. září 2005). „Eusocialita: původ a důsledky“. Proc. Natl. Acad. Sci. USA. 102 (38): 13367–1371. Bibcode:2005PNAS..10213367W. doi:10.1073 / pnas.0505858102. ISSN 0027-8424. PMC 1224642. PMID 16157878.
- ^ A b Gavrilets, Sergey (říjen 2003). „Perspektiva: modely speciace: co jsme se naučili za 40 let?“. Vývoj. 57 (10): 2197–2215. doi:10.1554/02-727. ISSN 0014-3820. PMID 14628909. S2CID 198158082.
- ^ A b C de Queiroz, Kevin (3. května 2005). „Ernst Mayr a moderní pojetí druhů“. Proc. Natl. Acad. Sci. USA. 102 (Příloha 1): 6600–6607. Bibcode:2005PNAS..102,6600D. doi:10.1073 / pnas.0502030102. ISSN 0027-8424. PMC 1131873. PMID 15851674.
- ^ A b Ereshefsky, Marc (Prosinec 1992). „Eliminativní pluralismus“. Filozofie vědy. 59 (4): 671–690. doi:10.1086/289701. ISSN 0031-8248. JSTOR 188136. S2CID 224829314.
- ^ Mayr 1942, str. 120
- ^ Fraser, Christophe; Alm, Eric J .; Polz, Martin F .; et al. (6. února 2009). „Výzva bakteriálních druhů: smysl pro genetickou a ekologickou rozmanitost“. Věda. 323 (5915): 741–746. Bibcode:2009Sci ... 323..741F. doi:10.1126 / science.1159388. ISSN 0036-8075. PMID 19197054. S2CID 15763831.
- ^ Krátký, Roger Valentine (říjen 1975). "Příspěvek mezka k vědeckému myšlení". Journal of Reproduction and Fertility. Doplněk (23): 359–364. ISSN 0449-3087. OCLC 1639439. PMID 1107543.
- ^ Gross, Briana L .; Rieseberg, Loren H. (květen – červen 2005). „Ekologická genetika homoploidní hybridní speciace“. Journal of Heredity. 96 (3): 241–252. doi:10.1093 / jhered / esi026. ISSN 0022-1503. PMC 2517139. PMID 15618301.
- ^ Burke, John M .; Arnold, Michael L. (prosinec 2001). "Genetika a zdatnost hybridů". Výroční přehled genetiky. 35: 31–52. doi:10.1146 / annurev.genet.35.102401.085719. ISSN 0066-4197. PMID 11700276. S2CID 26683922.
- ^ Vrijenhoek, Robert C. (4. dubna 2006). „Polyploidní hybridy: Vícenásobný původ druhu rosničky“. Aktuální biologie. 16 (7): R245 – R247. doi:10.1016 / j.cub.2006.03.005. ISSN 0960-9822. PMID 16581499. S2CID 11657663.
- ^ Rice, William R .; Hostert, Ellen E. (prosinec 1993). „Laboratorní experimenty se speciací: Co jsme se naučili za 40 let?“. Vývoj. 47 (6): 1637–1653. doi:10.1111 / j.1558-5646.1993.tb01257.x. ISSN 0014-3820. JSTOR 2410209. PMID 28568007. S2CID 42100751.
- Jiggins, Chris D .; Bridle, Jon R. (březen 2004). "Speciace v jablečném červovém mouchu: směs ročníků?". Trendy v ekologii a evoluci. 19 (3): 111–114. doi:10.1016 / j.tree.2003.12.008. ISSN 0169-5347. PMID 16701238.
- Boxhorn, Joseph (1. září 1995). „Pozorované instance speciace“. Archiv TalkOrigins. Houston, Texas: The TalkOrigins Foundation, Inc. Archivováno od původního dne 2009-01-22. Citováno 2008-12-26.
- Weinberg, James R .; Starczak, Victoria R .; Jörg, Daniele (srpen 1992). "Důkazy pro rychlou speciaci po události zakladatele v laboratoři". Vývoj. 46 (4): 1214–1220. doi:10.2307/2409766. ISSN 0014-3820. JSTOR 2409766. PMID 28564398.
- ^ Herrel, Anthony; Huyghe, Katleen; Vanhooydonck, Bieke; et al. (25. března 2008). „Rychlá rozsáhlá evoluční divergence morfologie a výkonu spojená s využíváním jiného zdroje potravy“. Proc. Natl. Acad. Sci. USA. 105 (12): 4792–4795. Bibcode:2008PNAS..105,4792H. doi:10.1073 / pnas.0711998105. ISSN 0027-8424. PMC 2290806. PMID 18344323.
- ^ Losos, Jonathan B .; Warhelt, Kenneth I .; Schoener, Thomas W. (1. května 1997). "Adaptivní diferenciace po experimentální kolonizaci ostrovů v Anolis ještěrky “. Příroda. 387 (6628): 70–73. Bibcode:1997 Natur.387 ... 70L. doi:10.1038 / 387070a0. ISSN 0028-0836. S2CID 4242248.
- ^ Hoskin, Conrad J .; Higgle, Megan; McDonald, Keith R .; Moritz, Craig (27. října 2005). Msgstr "Výztuž řídí rychlou alopatrickou speciaci". Příroda. 437 (7063): 1353–1356. Bibcode:2005 Natur.437.1353H. doi:10.1038 / nature04004. ISSN 0028-0836. PMID 16251964. S2CID 4417281.
- ^ Templeton, Alan R. (Duben 1980). „Teorie Speciace PŘES princip zakladatele “. Genetika. 94 (4): 1011–1038. ISSN 0016-6731. PMC 1214177. PMID 6777243. Archivováno z původního dne 2014-08-23. Citováno 2014-12-29.
- ^ Antonovics, Janis (Červenec 2006). „Vývoj v těsně sousedících populacích rostlin X: dlouhodobé přetrvávání předproduktivní izolace na hranici dolu“. Dědičnost. 97 (1): 33–37. doi:10.1038 / sj.hdy.6800835. ISSN 0018-067X. PMID 16639420. S2CID 12291411.
- ^ Nosil, Patrik; Crespi, Bernard J .; Gries, Regine; Gries, Gerhard (březen 2007). "Přirozený výběr a divergence v preferenci partnera během speciace". Genetica. 129 (3): 309–327. doi:10.1007 / s10709-006-0013-6. ISSN 0016-6707. PMID 16900317. S2CID 10808041.
- ^ Savolainen, Vincent; Anstett, Marie-Charlotte; Lexer, Christian; et al. (11. května 2006). "Sympatická speciace v dlaních na oceánském ostrově". Příroda. 441 (7090): 210–213. Bibcode:2006 Natur.441..210S. doi:10.1038 / nature04566. ISSN 0028-0836. PMID 16467788. S2CID 867216.
- Barluenga, Marta; Stölting, Kai N .; Salzburger, Walter; et al. (9. února 2006). „Sympatická speciace v cichlidách v nikaragujském kráterovém jezeře“. Příroda. 439 (7077): 719–23. Bibcode:2006 Natur.439..719B. doi:10.1038 / nature04325. ISSN 0028-0836. PMID 16467837. S2CID 3165729.
- ^ Gavrilets, Sergey (21. března 2006). "Maynard Smith model sympatrické speciace". Journal of Theoretical Biology. 239 (2): 172–182. doi:10.1016 / j.jtbi.2005.08.041. ISSN 0022-5193. PMID 16242727.
- ^ Wood, Troy E .; Takebayashi, Naoki; Barker, Michael S .; et al. (18. srpna 2009). „Frekvence specie polyploidů u cévnatých rostlin“. Proc. Natl. Acad. Sci. USA. 106 (33): 13875–13879. Bibcode:2009PNAS..10613875W. doi:10.1073 / pnas.0811575106. ISSN 0027-8424. PMC 2728988. PMID 19667210.
- ^ Hegarty, Matthew J .; Hiscock, Simon J. (20. května 2008). „Genomické stopy evolučního úspěchu polyploidních rostlin“. Aktuální biologie. 18 (10): R435 – R444. doi:10.1016 / j.cub.2008.03.043. ISSN 0960-9822. PMID 18492478. S2CID 1584282.
- ^ Jakobsson, Mattias; Hagenblad, Jenny; Tavaré, Simon; et al. (Červen 2006). „Unikátní nedávný původ alotetraploidních druhů Arabidopsis suecica: Důkazy z markerů nukleární DNA “. Molekulární biologie a evoluce. 23 (6): 1217–1231. doi:10.1093 / molbev / msk006. ISSN 0737-4038. PMID 16549398.
- ^ Säll, Torbjörn; Jakobsson, Mattias; Lind-Halldén, Christina; Halldén, Christer (září 2003). „Chloroplastová DNA označuje jediný původ alotetraploidu Arabidopsis suecica". Journal of Evolutionary Biology. 16 (5): 1019–1029. doi:10.1046 / j.1420-9101.2003.00554.x. ISSN 1010-061X. PMID 14635917. S2CID 29281998.
- ^ Bomblies, Kirsten; Weigel, Detlef (Prosinec 2007). "Arabidopsis—Model pro speciaci ". Aktuální názor na genetiku a vývoj. 17 (6): 500–504. doi:10.1016 / j.gde.2007.09.006. ISSN 0959-437X. PMID 18006296.
- ^ Sémon, Marie; Wolfe, Kenneth H. (prosinec 2007). "Důsledky duplikace genomu". Aktuální názor na genetiku a vývoj. 17 (6): 505–512. doi:10.1016 / j.gde.2007.09.007. ISSN 0959-437X. PMID 18006297.
- ^ Eldredge & Gould 1972, s. 82–115
- ^ Benton, Michael J. (7. dubna 1995). „Diverzifikace a zánik v historii života“. Věda. 268 (5207): 52–58. Bibcode:1995Sci ... 268 ... 52B. doi:10.1126 / science.7701342. ISSN 0036-8075. PMID 7701342.
- ^ Raup, David M. (28. března 1986). „Biologický zánik v historii Země“. Věda. 231 (4745): 1528–1533. Bibcode:1986Sci...231.1528R. doi:10.1126 / science.11542058. ISSN 0036-8075. PMID 11542058. S2CID 23012011.
- ^ Avise, John C .; Hubbell, Stephen P.; Ayala, Francisco J. (August 12, 2008). "In the light of evolution II: Biodiversity and extinction". Proc. Natl. Acad. Sci. USA. 105 (Suppl. 1): 11453–11457. Bibcode:2008PNAS..10511453A. doi:10.1073/pnas.0802504105. ISSN 0027-8424. PMC 2556414. PMID 18695213.
- ^ A b C Raup, David M. (July 19, 1994). "The role of extinction in evolution". Proc. Natl. Acad. Sci. USA. 91 (15): 6758–6763. Bibcode:1994PNAS...91.6758R. doi:10.1073/pnas.91.15.6758. ISSN 0027-8424. PMC 44280. PMID 8041694.
- ^ Novacek, Michael J .; Cleland, Elsa E. (May 8, 2001). "The current biodiversity extinction event: scenarios for mitigation and recovery". Proc. Natl. Acad. Sci. USA. 98 (10): 5466–5470. Bibcode:2001PNAS...98.5466N. doi:10.1073/pnas.091093698. ISSN 0027-8424. PMC 33235. PMID 11344295.
- ^ Pimm, Stuart; Raven, Peter; Peterson, Alan; et al. (18. července 2006). "Human impacts on the rates of recent, present and future bird extinctions". Proc. Natl. Acad. Sci. USA. 103 (29): 10941–10946. Bibcode:2006PNAS..10310941P. doi:10.1073/pnas.0604181103. ISSN 0027-8424. PMC 1544153. PMID 16829570.
- ^ Barnosky, Anthony D.; Koch, Paul L.; Feranec, Robert S.; et al. (October 1, 2004). "Assessing the Causes of Late Pleistocene Extinctions on the Continents". Věda. 306 (5693): 70–75. Bibcode:2004Sci...306...70B. CiteSeerX 10.1.1.574.332. doi:10.1126/science.1101476. ISSN 0036-8075. PMID 15459379. S2CID 36156087.
- ^ Lewis, Owen T. (January 29, 2006). "Climate change, species–area curves and the extinction crisis". Filozofické transakce královské společnosti B. 361 (1465): 163–171. doi:10.1098/rstb.2005.1712. ISSN 0962-8436. PMC 1831839. PMID 16553315.
- ^ A b Stearns & Stearns 1999, str.X
- ^ A b Novacek, Michael J. (November 8, 2014). „Brilantní budoucnost pravěku“. The New York Times. New York: The New York Times Company. ISSN 0362-4331. Archivováno z původního dne 2014-12-29. Citováno 2014-12-25.
- ^ „Vědci zjistili, že Země může být domovem 1 bilionu druhů“. Národní vědecká nadace. Arlington County, Virginia. 2. května 2016. Archivováno z původního dne 2016-05-04. Citováno 2016-05-06.
- ^ Jablonski, David (May 8, 2001). "Lessons from the past: Evolutionary impacts of mass extinctions". Proc. Natl. Acad. Sci. USA. 98 (10): 5393–5398. Bibcode:2001PNAS...98.5393J. doi:10.1073/pnas.101092598. ISSN 0027-8424. PMC 33224. PMID 11344284.
- ^ "Věk Země". Geologický průzkum Spojených států. 9. července 2007. Archivováno from the original on 2005-12-23. Citováno 2015-05-31.
- ^ Dalrymple 2001, pp. 205–221
- ^ Manhesa, Gérard; Allègre, Claude J.; Dupréa, Bernard; Hamelin, Bruno (May 1980). „Olovo izotopová studie základních ultrabazických vrstvených komplexů: spekulace o stáří Země a charakteristikách primitivního pláště“. Dopisy o Zemi a planetách. 47 (3): 370–382. Bibcode:1980E & PSL..47..370M. doi:10.1016 / 0012-821X (80) 90024-2. ISSN 0012-821X.
- ^ Raven & Johnson 2002, str. 68
- ^ A b Borenstein, Seth (19. října 2015). „Náznaky života na tom, co bylo považováno za pustou ranou Zemi“. Rozrušit. Yonkers, NY: Interaktivní síť Mindspark. Associated Press. Archivovány od originál dne 23. října 2015. Citováno 8. října 2018.
- ^ Bell, Elizabeth A .; Boehnike, Patrick; Harrison, T. Mark; Mao, Wendy L. (November 24, 2015). „Potenciálně biogenní uhlík konzervovaný v 4,1 miliard let starém zirkonu“ (PDF). Proc. Natl. Acad. Sci. USA. 112 (47): 14518–14521. Bibcode:2015PNAS..11214518B. doi:10.1073 / pnas.1517557112. ISSN 0027-8424. PMC 4664351. PMID 26483481. Archivováno (PDF) od originálu dne 06.11.2015. Citováno 2015-12-30.
- ^ Schouten, Lucy (October 20, 2015). "When did life first emerge on Earth? Maybe a lot earlier than we thought". Christian Science Monitor. Boston, Massachusetts: Christian Science Publishing Society. ISSN 0882-7729. Archivováno od původního dne 2016-03-22. Citováno 2018-07-11.
- ^ Wade, Nicholas (25. července 2016). „Seznamte se s Lucem, předkem všeho živého“. The New York Times. New York: Společnost New York Times. ISSN 0362-4331. Archivováno z původního dne 28. července 2016. Citováno 2016-07-25.
- ^ McKinney 1997, str.110
- ^ Mora, Camilo; Tittensor, Derek P .; Adl, Sina; et al. (August 23, 2011). „Kolik druhů je na Zemi a v oceánu?“. PLOS Biology. 9 (8): e1001127. doi:10.1371 / journal.pbio.1001127. ISSN 1545-7885. PMC 3160336. PMID 21886479.
- ^ Miller & Spoolman 2012, str.62
- ^ Chapman 2009
- ^ Roskov, Y.; Abucay, L.; Orrell, T.; Nicolson, D.; et al., eds. (2016). "Species 2000 & ITIS Catalogue of Life, 2016 Annual Checklist". Species 2000. Leiden, Nizozemsko: Centrum biologické rozmanitosti Naturalis. ISSN 2405-884X. Archivováno od originálu 2016-11-12. Citováno 2016-11-06.
- ^ Peretó, Juli (March 2005). „Spory o původu života“ (PDF). Mezinárodní mikrobiologie. 8 (1): 23–31. ISSN 1139-6709. PMID 15906258. Archivovány od originál (PDF) on 2015-08-24.
- ^ Marshall, Michael (11. listopadu 2020). „Tušení Charlese Darwina o časném životě bylo pravděpodobně správné - v několika načmáraných poznámkách příteli biolog Charles Darwin teoretizoval, jak život začal. Nejen, že to bylo pravděpodobně správné, jeho teorie byla o století napřed před jeho dobou.“. BBC novinky. Citováno 11. listopadu 2020.
- ^ Joyce, Gerald F. (July 11, 2002). „Starověk evoluce založené na RNA“. Příroda. 418 (6894): 214–221. Bibcode:2002 Natur.418..214J. doi:10.1038 / 418214a. ISSN 0028-0836. PMID 12110897. S2CID 4331004.
- ^ Trevors, Jack T .; Psenner, Roland (prosinec 2001). "From self-assembly of life to present-day bacteria: a possible role for nanocells". Recenze mikrobiologie FEMS. 25 (5): 573–582. doi:10.1111 / j.1574-6976.2001.tb00592.x. ISSN 1574-6976. PMID 11742692.
- ^ Theobald, Douglas L. (May 13, 2010). "Formální test teorie univerzálního společného původu". Příroda. 465 (7295): 219–222. Bibcode:2010Natur.465..219T. doi:10.1038 / nature09014. ISSN 0028-0836. PMID 20463738. S2CID 4422345.
- ^ Bapteste, Eric; Walsh, David A. (June 2005). "Does the 'Ring of Life' ring true?". Trendy v mikrobiologii. 13 (6): 256–261. doi:10.1016/j.tim.2005.03.012. ISSN 0966-842X. PMID 15936656.
- ^ Darwin 1859, str.1
- ^ Doolittle, W. Ford; Bapteste, Eric (February 13, 2007). "Pattern pluralism and the Tree of Life hypothesis". Proc. Natl. Acad. Sci. USA. 104 (7): 2043–2049. Bibcode:2007PNAS..104.2043D. doi:10.1073/pnas.0610699104. ISSN 0027-8424. PMC 1892968. PMID 17261804.
- ^ Kunin, Victor; Goldovsky, Leon; Darzentas, Nikos; Ouzounis, Christos A. (July 2005). "The net of life: Reconstructing the microbial phylogenetic network". Výzkum genomu. 15 (7): 954–959. doi:10.1101/gr.3666505. ISSN 1088-9051. PMC 1172039. PMID 15965028.
- ^ Darwin 1837, str.25
- ^ Jablonski, David (June 25, 1999). "The Future of the Fossil Record". Věda. 284 (5423): 2114–2116. doi:10.1126 / science.284.5423.2114. ISSN 0036-8075. PMID 10381868. S2CID 43388925.
- ^ Mason, Stephen F. (September 6, 1984). "Počátky biomolekulární slabosti". Příroda. 311 (5981): 19–23. Bibcode:1984 Natur.311 ... 19M. doi:10.1038 / 311019a0. ISSN 0028-0836. PMID 6472461. S2CID 103653.
- ^ Wolf, Yuri I.; Rogozin, Igor B.; Grishin, Nick V.; Koonin, Eugene V. (1. září 2002). "Genome trees and the tree of life". Trendy v genetice. 18 (9): 472–479. doi:10.1016/S0168-9525(02)02744-0. ISSN 0168-9525. PMID 12175808.
- ^ Varki, Ajit; Altheide, Tasha K. (December 2005). "Comparing the human and chimpanzee genomes: searching for needles in a haystack". Výzkum genomu. 15 (12): 1746–1758. CiteSeerX 10.1.1.673.9212. doi:10.1101/gr.3737405. ISSN 1088-9051. PMID 16339373.
- ^ Ciccarelli, Francesca D .; Doerks, Tobias; von Mering, Christian; et al. (3. března 2006). „Směrem k automatické rekonstrukci vysoce vyřešeného stromu života“ (PDF). Věda. 311 (5765): 1283–1287. Bibcode:2006Sci ... 311.1283C. CiteSeerX 10.1.1.381.9514. doi:10.1126 / science.1123061. ISSN 0036-8075. PMID 16513982. S2CID 1615592. Archivováno (PDF) from the original on 2016-03-04.
- ^ A b Cavalier-Smith, Thomas (29. června 2006). „Evoluce buněk a historie Země: stagnace a revoluce“. Filozofické transakce královské společnosti B. 361 (1470): 969–1006. doi:10.1098 / rstb.2006.1842. ISSN 0962-8436. PMC 1578732. PMID 16754610.
- ^ Schopf, J. William (29. června 2006). „Fosilní důkazy o archaejském životě“. Filozofické transakce královské společnosti B. 361 (1470): 869–885. doi:10.1098 / rstb.2006.1834. ISSN 0962-8436. PMC 1578735. PMID 16754604.
- ^ Schopf, J. William (July 19, 1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proc. Natl. Acad. Sci. USA. 91 (15): 6735–6742. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. ISSN 0027-8424. PMC 44277. PMID 8041691.
- ^ Poole, Anthony M.; Penny, David (January 2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. ISSN 0265-9247. PMID 17187354.
- ^ Dyall, Sabrina D .; Brown, Mark T .; Johnson, Patricia J. (9. dubna 2004). "Starověké invaze: Od endosymbiontů po organely". Věda. 304 (5668): 253–257. Bibcode:2004Sci ... 304..253D. doi:10.1126 / science.1094884. ISSN 0036-8075. PMID 15073369. S2CID 19424594.
- ^ Martin, William (October 2005). "The missing link between hydrogenosomes and mitochondria". Trendy v mikrobiologii. 13 (10): 457–459. doi:10.1016/j.tim.2005.08.005. ISSN 0966-842X. PMID 16109488.
- ^ Lang, B. Franz; Gray, Michael W .; Burger, Gertraud (December 1999). "Mitochondrial genome evolution and the origin of eukaryotes". Výroční přehled genetiky. 33: 351–397. doi:10.1146/annurev.genet.33.1.351. ISSN 0066-4197. PMID 10690412.
- ^ DeLong, Edward F.; Pace, Norman R. (1. srpna 2001). "Environmental Diversity of Bacteria and Archaea" (PDF). Systematická biologie. 50 (4): 470–478. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. ISSN 1063-5157. PMID 12116647. Archivováno (PDF) from the original on 2016-02-22.
- ^ Kaiser, Dale (Prosinec 2001). "Budování mnohobuněčného organismu". Výroční přehled genetiky. 35: 103–123. doi:10.1146 / annurev.genet.35.102401.090145. ISSN 0066-4197. PMID 11700279. S2CID 18276422.
- ^ Zimmer, Carle (7. ledna 2016). "Genetic Flip Helped Organisms Go From One Cell to Many". The New York Times. New York: The New York Times Company. ISSN 0362-4331. Archivováno z původního dne 2016-01-07. Citováno 2016-01-07.
- ^ Valentine, James W.; Jablonski, David; Erwin, Douglas H. (March 1, 1999). "Fossils, molecules and embryos: new perspectives on the Cambrian explosion". Rozvoj. 126 (5): 851–859. ISSN 0950-1991. PMID 9927587. Archivováno from the original on 2015-03-01. Citováno 2014-12-30.
- ^ Ohno, Susumu (January 1997). "The reason for as well as the consequence of the Cambrian explosion in animal evolution". Journal of Molecular Evolution. 44 (Suppl. 1): S23–S27. Bibcode:1997JMolE..44S..23O. doi:10.1007/PL00000055. ISSN 0022-2844. PMID 9071008. S2CID 21879320.
- Valentine, James W .; Jablonski, David (2003). "Morphological and developmental macroevolution: a paleontological perspective". International Journal of Developmental Biology. 47 (7–8): 517–522. ISSN 0214-6282. PMID 14756327. Archivováno od originálu 2014-10-24. Citováno 2014-12-30.
- ^ Waters, Elizabeth R. (December 2003). "Molecular adaptation and the origin of land plants". Molekulární fylogenetika a evoluce. 29 (3): 456–463. doi:10.1016/j.ympev.2003.07.018. ISSN 1055-7903. PMID 14615186.
- ^ Mayhew, Peter J. (Srpen 2007). "Why are there so many insect species? Perspectives from fossils and phylogenies". Biologické recenze. 82 (3): 425–454. doi:10.1111/j.1469-185X.2007.00018.x. ISSN 1464-7931. PMID 17624962. S2CID 9356614.
- ^ Carroll, Robert L. (Květen 2007). "The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians". Zoologický žurnál Linneanské společnosti. 150 (Supplement s1): 1–140. doi:10.1111/j.1096-3642.2007.00246.x. ISSN 1096-3642.
- ^ Wible, John R .; Rougier, Guillermo W .; Novacek, Michael J .; Asher, Robert J. (June 21, 2007)."Křídové eutherians a Laurasian původ pro placentární savce poblíž hranice K / T". Příroda. 447 (7147): 1003–1006. Bibcode:2007Natur.447.1003W. doi:10.1038 / nature05854. ISSN 0028-0836. PMID 17581585. S2CID 4334424.
- ^ Witmer, Lawrence M. (28. července 2011). "Paleontologie: Ikona klepaná z okouna". Příroda. 475 (7357): 458–459. doi:10.1038 / 475458a. ISSN 0028-0836. PMID 21796198. S2CID 205066360.
- ^ Bull, James J.; Wichman, Holly A. (listopad 2001). "Aplikovaná evoluce". Výroční přehled ekologie a systematiky. 32: 183–217. doi:10.1146 / annurev.ecolsys.32.081501.114020. ISSN 1545-2069.
- ^ Doebley, John F .; Gaut, Brandon S .; Smith, Bruce D. (29. prosince 2006). „Molekulární genetika domestikace plodin“. Buňka. 127 (7): 1309–1321. doi:10.1016 / j.cell.2006.12.006. ISSN 0092-8674. PMID 17190597. S2CID 278993.
- ^ Jäckel, Christian; Kast, Peter; Hilvert, Donald (červen 2008). "Protein Design by Directed Evolution". Roční přehled biofyziky. 37: 153–173. doi:10.1146 / annurev.biophys.37.032807.125832. ISSN 1936-122X. PMID 18573077.
- ^ Maher, Brendan (8. dubna 2009). „Evolution: Biology's next top model?“. Příroda. 458 (7239): 695–698. doi:10.1038 / 458695a. ISSN 0028-0836. PMID 19360058. S2CID 41648315.
- ^ Borowsky, Richard (8. ledna 2008). "Obnovení zraku u slepých jeskynních ryb". Aktuální biologie. 18 (1): R23 – R24. doi:10.1016 / j.cub.2007.11.023. ISSN 0960-9822. PMID 18177707. S2CID 16967690.
- ^ Gross, Joshua B .; Borowsky, Richard; Tabin, Clifford J. (2. ledna 2009). Barsh, Gregory S. (ed.). "Nová role pro Mc1r v paralelním vývoji depigmentace v nezávislých populacích jeskynních ryb Astyanax mexicanus". Genetika PLOS. 5 (1): e1000326. doi:10.1371 / journal.pgen.1000326. ISSN 1553-7390. PMC 2603666. PMID 19119422.
- ^ Merlo, Lauren M.F .; Pepper, John W .; Reid, Brian J .; Maley, Carlo C. (Prosinec 2006). „Rakovina jako evoluční a ekologický proces“. Nature Reviews Cancer. 6 (12): 924–935. doi:10.1038 / nrc2013. ISSN 1474-175X. PMID 17109012. S2CID 8040576.
- ^ Pan, Dabo; Weiwei Xue; Wenqi Zhang; et al. (Říjen 2012). „Pochopení mechanismu rezistence vůči viru hepatitidy C NS3 / 4A na ITMN-191 v důsledku mutací R155K, A156V, D168A / E: výpočetní studie“. Biochimica et Biophysica Acta (BBA) - Obecné předměty. 1820 (10): 1526–1534. doi:10.1016 / j.bbagen.2012.06.001. ISSN 0304-4165. PMID 22698669.
- ^ Woodford, Neil; Ellington, Matthew J. (leden 2007). "Vznik rezistence na antibiotika mutací". Klinická mikrobiologie a infekce. 13 (1): 5–18. doi:10.1111 / j.1469-0691.2006.01492.x. ISSN 1198-743X. PMID 17184282.
- ^ Labbé, Pierrick; Berticat, Claire; Berthomieu, Arnaud; et al. (16. listopadu 2007). „Čtyřicet let vývoje nepravidelné insekticidní rezistence u komárů Culex pipiens". Genetika PLOS. 3 (11): e205. doi:10.1371 / journal.pgen.0030205. ISSN 1553-7390. PMC 2077897. PMID 18020711.
- ^ Neve, Paul (říjen 2007). „Výzvy pro vývoj a řízení rezistence vůči herbicidům: 50 let po Harperovi“. Výzkum plevelů. 47 (5): 365–369. doi:10.1111 / j.1365-3180.2007.00581.x. ISSN 0043-1737.
- ^ Rodríguez-Rojas, Alexandro; Rodríguez-Beltrán, Jerónimo; Couce, Alejandro; Blázquez, Jesús (srpen 2013). „Antibiotika a rezistence na antibiotika: hořký boj proti evoluci“. International Journal of Medical Microbiology. 303 (6–7): 293–297. doi:10.1016 / j.ijmm.2013.02.004. ISSN 1438-4221. PMID 23517688.
- ^ Schenk, Martijn F .; Szendro, Ivan G .; Krug, Joachim; de Visser, J. Arjan G.M. (28. června 2012). „Kvantifikace adaptivního potenciálu enzymu rezistence na antibiotika“. Genetika PLOS. 8 (6): e1002783. doi:10.1371 / journal.pgen.1002783. ISSN 1553-7390. PMC 3386231. PMID 22761587.
- ^ Přečtěte si, Andrew F .; Lynch, Penelope A .; Thomas, Matthew B. (7. dubna 2009). „Jak vyrobit insekticidy odolné proti evoluci pro kontrolu malárie“. PLOS Biology. 7 (4): e1000058. doi:10.1371 / journal.pbio.1000058. ISSN 1545-7885. PMC 3279047. PMID 19355786.
- ^ Fraser, Alex S. (18. ledna 1958). „Monte Carlo Analýzy genetických modelů“. Příroda. 181 (4603): 208–209. Bibcode:1958Natur.181..208F. doi:10.1038 / 181208a0. ISSN 0028-0836. PMID 13504138. S2CID 4211563.
- ^ Rechenberg 1973
- ^ Holland 1975
- ^ Koza 1992
- ^ Jamshidi, Mo (15. srpna 2003). "Nástroje pro inteligentní řízení: fuzzy regulátory, neuronové sítě a genetické algoritmy". Filozofické transakce královské společnosti A. 361 (1809): 1781–1808. Bibcode:2003RSPTA.361.1781J. doi:10.1098 / rsta.2003.1225. ISSN 1364-503X. PMID 12952685. S2CID 34259612.
- ^ Browne 2003, str. 376–379
- ^ Přehled filosofických, náboženských a kosmologických kontroverzí najdete na:Vědecké a sociální recepce evoluce v 19. a na počátku 20. století viz:
- Johnston, Ian C. (1999). „Část třetí: Počátky evoluční teorie“. ... A přesto se vyvíjíme: Příručka pro rané dějiny moderní vědy (3. přepracované vydání). Nanaimo, BC: Oddělení liberálních studií, Malaspina University-College. Archivováno od originálu 2016-04-16. Citováno 2015-01-01.
- Bowler 2003
- Zuckerkandl, Emile (30. prosince 2006). "Inteligentní design a biologická složitost". Gen. 385: 2–18. doi:10.1016 / j.gene.2006.03.025. ISSN 0378-1119. PMID 17011142.
- ^ Ross, Marcus R. (Květen 2005). „Kdo čemu věří? Odstranění zmatku nad inteligentním designem a kreacionismem mladé Země“ (PDF). Journal of Geoscience Education. 53 (3): 319–323. Bibcode:2005JGeEd..53..319R. CiteSeerX 10.1.1.404.1340. doi:10.5408/1089-9995-53.3.319. ISSN 1089-9995. S2CID 14208021. Archivováno (PDF) od původního dne 2008-05-11. Citováno 2008-04-28.
- ^ Hameed, Salman (12. prosince 2008). „Podpora pro islámský kreacionismus“ (PDF). Věda. 322 (5908): 1637–1638. doi:10.1126 / science.1163672. ISSN 0036-8075. PMID 19074331. S2CID 206515329. Archivovány od originál (PDF) dne 10.11.2014.
- ^ Bowler 2003
- ^ Miller, Jon D .; Scott, Eugenie C .; Okamoto, Shinji (11. srpna 2006). „Veřejné přijetí evoluce“. Věda. 313 (5788): 765–766. doi:10.1126 / science.1126746. ISSN 0036-8075. PMID 16902112. S2CID 152990938.
- ^ Spergel, David Nathaniel; Verde, Licia; Peiris, Hiranya V .; et al. (2003). „Pozorování mikrovlnné anizotropie sondy Wilkinson (WMAP) prvního roku: Stanovení kosmologických parametrů“. Astrophysical Journal Supplement Series. 148 (1): 175–194. arXiv:astro-ph / 0302209. Bibcode:2003ApJS..148..175S. doi:10.1086/377226. S2CID 10794058.
- ^ Wilde, Simon A .; Valley, John W .; Peck, William H .; Graham, Colin M. (11. ledna 2001). "Důkazy z detritických zirkonů o existenci kontinentální kůry a oceánů na Zemi před 4,4 Gyr". Příroda. 409 (6817): 175–178. Bibcode:2001 Natur.409..175 W.. doi:10.1038/35051550. ISSN 0028-0836. PMID 11196637. S2CID 4319774.
- ^ Pobočka, Glenn (Březen 2007). "Pochopení kreacionismu po Kitzmiller". BioScience. 57 (3): 278–284. doi:10.1641 / B570313. ISSN 0006-3568. S2CID 86665329.
- ^ Xiaoxing Jin (březen 2019). "Překlad a transmutace: Původ druhů v Číně". Britský časopis pro dějiny vědy. Cambridge: Cambridge University Press jménem Britské společnosti pro dějiny vědy. 52 (1): 117–141. doi:10.1017 / S0007087418000808. PMID 30587253.
Bibliografie
- Altenberg, Lee (1995). "Růst genomu a vývoj mapy genotyp-fenotyp". V Banzhaf, Wolfgang; Eeckman, Frank H. (eds.). Evoluce a biopočítání: výpočetní modely evoluce. Přednášky z informatiky. 899. Berlín; New York: Springer-Verlag Berlin Heidelberg. str. 205–259. CiteSeerX 10.1.1.493.6534. doi:10.1007/3-540-59046-3_11. ISBN 978-3-540-59046-0. ISSN 0302-9743. LCCN 95005970. OCLC 32049812.
- Ayala, Francisco J.; Avise, John C., eds. (2014). Základní čtení v evoluční biologii. Baltimore, Maryland: Johns Hopkins University Press. ISBN 978-1-4214-1305-1. LCCN 2013027718. OCLC 854285705.
- Birdsell, John A .; Wills, Christopher (2003). „Evoluční původ a udržování sexuální rekombinace: Přehled současných modelů“. V MacIntyre, Ross J .; Clegg, Michael T. (eds.). Evoluční biologie. Evoluční biologie. 33. New York: Springer Science + Business Media. ISBN 978-1-4419-3385-0. ISSN 0071-3260. OCLC 751583918.
- Bowler, Peter J. (1989). Mendelovská revoluce: Vznik dědičných konceptů v moderní vědě a společnosti. Baltimore, Maryland: Johns Hopkins University Press. ISBN 978-0-8018-3888-0. LCCN 89030914. OCLC 19322402.
- Bowler, Peter J. (2003). Evolution: The History of an Idea (3. zcela rev. A rozšířené vydání). Berkeley, Kalifornie: University of California Press. ISBN 978-0-520-23693-6. LCCN 2002007569. OCLC 49824702.
- Browne, Janet (2003). Charles Darwin: The Power of Place. 2. Londýn: Pimlico. ISBN 978-0-7126-6837-8. LCCN 94006598. OCLC 52327000.
- Burkhardt, Frederick; Smith, Sydney, eds. (1991). Korespondence Charlese Darwina. Korespondence Charlese Darwina. 7: 1858–1859. Cambridge: Cambridge University Press. ISBN 978-0-521-38564-0. LCCN 84045347. OCLC 185662993.
- Carroll, Sean B.; Grenier, Jennifer K .; Weatherbee, Scott D. (2005). Od DNA k rozmanitosti: Molekulární genetika a vývoj zvířecího designu (2. vyd.). Malden, Massachusetts: Blackwell Publishing. ISBN 978-1-4051-1950-4. LCCN 2003027991. OCLC 53972564.
- Chapman, Arthur D. (2009). Počet žijících druhů v Austrálii a ve světě (2. vyd.). Canberra: Oddělení pro životní prostředí, vodu, dědictví a umění: Australská studie biologických zdrojů. ISBN 978-0-642-56860-1. OCLC 780539206. Archivováno od původního dne 2016-12-25. Citováno 2016-11-06.
- Coyne, Jerry A. (2009). Proč je evoluce pravdivá. New York: Viking. ISBN 978-0-670-02053-9. LCCN 2008033973. OCLC 233549529.
- Cracraft, Joel; Bybee, Rodger W., eds. (2005). Evoluční věda a společnost: Vzdělávání nové generace (PDF). Colorado Springs, Colorado: Studium kurikula biologických věd. ISBN 978-1-929614-23-3. OCLC 64228003. Citováno 2014-12-06. „Revised Proceedings of the BSCS, AIBS Symposium November 2004, Chicago, Illinois“
- Dalrymple, G. Brent (2001). „Věk Země ve dvacátém století: problém (většinou) vyřešený“. In Lewis, C.L.E .; Knell, S.J. (eds.). The Age of the Earth: from 4004 BC to AD 2002. Geologická společnost, Londýn, speciální publikace. Zvláštní publikace geologické společnosti. 190. str. 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144 / gsl.sp.2001.190.01.14. ISBN 978-1-86239-093-5. LCCN 2003464816. OCLC 48570033. S2CID 130092094.
- Darwin, Charles (1837). Notebook „B“. Notebook je k dispozici na webu Kompletní dílo Charlese Darwina online. Citováno 2019-10-09.
- Darwin, Charles (1859). O původu druhů pomocí přirozeného výběru nebo zachování příznivých ras v boji o život (1. vyd.). Londýn: John Murray. LCCN 06017473. OCLC 741260650. Kniha je k dispozici od Kompletní dílo Charlese Darwina online. Citováno 2014-11-21.
- Darwin, Charles (1872). Vyjádření emocí u člověka a zvířat. Londýn: John Murray. LCCN 04002793. OCLC 1102785.
- Darwin, Francis, vyd. (1909). Základy původu druhů, náčrt napsaný v roce 1842 (PDF). Cambridge: Vytištěno na University Press. LCCN 61057537. OCLC 1184581. Citováno 2014-11-27.
- Dawkins, Richarde (1990). Slepý hodinář. Věda o tučňácích. Londýn: Knihy tučňáků. ISBN 978-0-14-014481-9. OCLC 60143870.
- Dennett, Daniel (1995). Darwinova nebezpečná myšlenka: Evoluce a významy života. New York: Simon & Schuster. ISBN 978-0-684-80290-9. LCCN 94049158. OCLC 31867409.
- Dobžanský, Theodosius (1968). „K některým základním koncepcím darwinovské biologie“. V Dobžanském, Theodosius; Hecht, Max K .; Steere, William C. (eds.). Evoluční biologie. Svazek 2 (1. vyd.). New York: Appleton-Century-Crofts. s. 1–34. doi:10.1007/978-1-4684-8094-8_1. ISBN 978-1-4684-8096-2. OCLC 24875357.
- Dobžanský, Theodosius (1970). Genetika evolučního procesu. New York: Columbia University Press. ISBN 978-0-231-02837-0. LCCN 72127363. OCLC 97663.
- Eldredge, Niles; Gould, Stephen Jay (1972). "Interpunkční rovnováha: alternativa k fyletickému gradualismu". V Schopf, Thomas J.M. (ed.). Modely v paleobiologii. San Francisco, Kalifornie: Freeman, Cooper. ISBN 978-0-87735-325-6. LCCN 72078387. OCLC 572084.
- Eldredge, Niles (1985). Časové rámce: Přehodnocení darwinovské evoluce a teorie přerušované rovnováhy. New York: Simon & Schuster. ISBN 978-0-671-49555-8. LCCN 84023632. OCLC 11443805.
- Ewens, Warren J. (2004). Genetika matematické populace. Interdisciplinární aplikovaná matematika. Já. Teoretický úvod (2. vyd.). New York: Springer-Verlag New York. ISBN 978-0-387-20191-7. LCCN 2003065728. OCLC 53231891.
- Fisher, Ronald A. (1930). Genetická teorie přirozeného výběru. Oxford: Clarendon Press. ISBN 978-0-19-850440-5. LCCN 30029177. OCLC 18500548.
- Futuyma, Douglas J. (2004). „Ovoce stromu života: Pohled na evoluci a ekologii“. V Cracraft, Joel; Donoghue, Michael J. (eds.). Sestavení stromu života. Oxford; New York: Oxford University Press. ISBN 978-0-19-517234-8. LCCN 2003058012. OCLC 61342697. „Sborník ze sympozia konaného v Americkém muzeu přírodní historie v New Yorku, 2002.“
- Futuyma, Douglas J. (2005). Vývoj. Sunderland, Massachusetts: Sinauer Associates. ISBN 978-0-87893-187-3. LCCN 2004029808. OCLC 57311264.
- Futuyma, Douglas J .; Kirkpatrick, Mark (2017). Vývoj (Čtvrté vydání). Sunderland, Massachusetts: Sinauer Associates. ISBN 978-1-60535-605-1. LCCN 2017000562. OCLC 969439375.
- Gould, Stephen Jay (2002). Struktura evoluční teorie. Cambridge, Massachusetts: Belknap Press z Harvard University Press. ISBN 978-0-674-00613-3. LCCN 2001043556. OCLC 47869352.
- Gray, Peter (2007). Psychologie (5. vydání). New York: Worth Publishers. ISBN 978-0-7167-0617-5. LCCN 2006921149. OCLC 76872504.
- Hall, Brian K.; Hallgrímsson, Benedikt (2008). Strickbergerova evoluce (4. vydání). Sudbury, Massachusetts: vydavatelé Jones a Bartlett. ISBN 978-0-7637-0066-9. LCCN 2007008981. OCLC 85814089.
- Hennig, Willi (1999) [Původně publikováno 1966 (dotisk 1979); přeloženo z autorovy nepublikované revize Grundzüge einer Theorie der phylogenetischen Systematik, publikováno v roce 1950]. Fylogenetická systematika. Překlad D. Dwighta Davise a Rainera Zangerla; předmluva Donna E. Rosena, Garetha Nelsona a Colin Patterson (Reedice ed.). Urbana, Illinois: University of Illinois Press. ISBN 978-0-252-06814-0. LCCN 78031969. OCLC 722701473.
- Holland, John H. (1975). Adaptace v přírodních a umělých systémech: Úvodní analýza s aplikacemi v biologii, řízení a umělé inteligenci. Ann Arbor, Michigan: University of Michigan Press. ISBN 978-0-472-08460-9. LCCN 74078988. OCLC 1531617.
- Jablonka, Eva; Jehněčí, Marion J. (2005). Evoluce ve čtyřech dimenzích: genetická, epigenetická, behaviorální a symbolická variace v historii života. Ilustrace Anny Zeligowski. Cambridge, Massachusetts: MIT Stiskněte. ISBN 978-0-262-10107-3. LCCN 2004058193. OCLC 61896061.
- Kampourakis, Kostas (2014). Pochopení evoluce. Cambridge; New York: Cambridge University Press. ISBN 978-1-107-03491-4. LCCN 2013034917. OCLC 855585457.
- Kirk, Geoffrey; Raven, Johne; Schofield, Malcolm (1983). Presokratičtí filozofové: Kritická historie s výběrem textů (2. vyd.). Cambridge; New York: Cambridge University Press. ISBN 978-0-521-27455-5. LCCN 82023505. OCLC 9081712.
- Koza, John R. (1992). Genetické programování: O programování počítačů pomocí přirozeného výběru. Složité adaptivní systémy. Cambridge, Massachusetts: MIT Press. ISBN 978-0-262-11170-6. LCCN 92025785. OCLC 26263956.
- Lamarck, Jean-Baptiste (1809). Philosophie Zoologique. Paříž: Dentu et L'Auteur. OCLC 2210044. Philosophie zoologique (1809) na Internetový archiv. Citováno 2014-11-29.
- Lane, David H. (1996). Fenomén Teilharda: Prorok pro nový věk (1. vyd.). Macon, Gruzie: Mercer University Press. ISBN 978-0-86554-498-7. LCCN 96008777. OCLC 34710780.
- Levinson, Gene (2019). Přehodnocení evoluce: Revoluce, která se skrývá v očích. Hackensack, New Jersey: World Scientific. ISBN 978-1-78634-726-8. LCCN 2019013762. OCLC 1138095098.
- Lucretius. „Kniha V, řádky 855–877“. De Rerum Natura. Digitální knihovna Perseus. Upravil a přeložil William Ellery Leonard (1916). Medford / Somerville, Massachusetts: Tufts University. OCLC 33233743. Archivováno z původního dne 2014-09-04. Citováno 2014-11-25.
- Magner, Lois N. (2002). Historie věd o živé přírodě (3. rev. A rozšířené vydání). New York: Marcel Dekker. ISBN 978-0-8247-0824-5. LCCN 2002031313. OCLC 50410202.
- Mason, Stephen F. (1962). Historie věd. Collier Books. Science Library, CS9 (New rev. Ed.). New York: Collier Books. LCCN 62003378. OCLC 568032626.
- Maynard Smith, John (1978). Evoluce sexu. Cambridge; New York: Cambridge University Press. ISBN 978-0-521-29302-0. LCCN 77085689. OCLC 3413793.
- Maynard Smith, John (1998). "Jednotky výběru". In Bock, Gregory R .; Sbohem, Jamie A. (eds.). Meze redukcionismu v biologii. Sympózium Nadace Novartis. Symposia Novartis Foundation. 213. Chichester; New York: John Wiley & Sons. 203–221. doi:10.1002 / 9780470515488.ch15. ISBN 978-0-471-97770-4. LCCN 98002779. OCLC 38311600. PMID 9653725. „Příspěvky ze sympozia o mezích redukcionismu v biologii, které se konalo v Nadaci Novartis v Londýně ve dnech 13. – 15. Května 1997.“
- Mayr, Ernst (1942). Systematika a původ druhů z pohledu zoologa. Columbia Biological Series. 13. New York: Columbia University Press. LCCN 43001098. OCLC 766053.
- Mayr, Ernst (1982). Růst biologického myšlení: rozmanitost, evoluce a dědičnost. Překlad John Ray E. Silk. Cambridge, Massachusetts: Belknap Press. ISBN 978-0-674-36445-5. LCCN 81013204. OCLC 7875904.
- Mayr, Ernst (2002) [Původně publikováno 2001; New York: Základní knihy ]. Co je to evoluce. Mistři vědy. Londýn: Weidenfeld a Nicolson. ISBN 978-0-297-60741-0. LCCN 2001036562. OCLC 248107061.
- McKinney, Michael L. (1997). „Jak se vzácné druhy vyhnou vyhynutí? Paleontologický pohled“. V Kunin, William E .; Gaston, Kevin J. (eds.). Biologie vzácnosti: Příčiny a důsledky vzácných - běžných rozdílů (1. vyd.). Londýn; New York: Chapman & Hall. ISBN 978-0-412-63380-5. LCCN 96071014. OCLC 36442106.
- Miller, G. Tyler; Spoolman, Scott E. (2012). Věda o životním prostředí (14. vydání). Belmont, Kalifornie: Brooks / Cole. ISBN 978-1-111-98893-7. LCCN 2011934330. OCLC 741539226. Citováno 2014-12-27.
- Moore, Randy; Decker, Mark; Cotner, Sehoya (2010). Chronologie diskuse evoluce-kreacionismu. Santa Barbara, Kalifornie: Greenwood Press /ABC-CLIO. ISBN 978-0-313-36287-3. LCCN 2009039784. OCLC 422757410.
- Nardon, Paul; Grenier, Anne-Marie (1991). „Serial Endosymbiosis Theory and Weevil Evolution: The Role of Symbiosis“. V Margulis, Lynn; Fester, René (eds.). Symbióza jako zdroj evolučních inovací: Speciace a morfogeneze. Cambridge, Massachusetts: MIT Press. ISBN 978-0-262-13269-5. LCCN 90020439. OCLC 22597587. „Na základě konference konané v Bellagiu v Itálii ve dnech 25. – 30. Června 1989“
- Národní akademie věd; Lékařský ústav (2008). Věda, evoluce a kreacionismus. Washington, DC: National Academy Press. ISBN 978-0-309-10586-6. LCCN 2007015904. OCLC 123539346. Citováno 2014-11-22.
- Odum, Eugene P. (1971). Základy ekologie (3. vyd.). Philadelphia, Pensylvánie: Saunders. ISBN 978-0-7216-6941-0. LCCN 76081826. OCLC 154846.
- Okasha, Samir (2006). Evoluce a úrovně výběru. Oxford; New York: Oxford University Press. ISBN 978-0-19-926797-2. LCCN 2006039679. OCLC 70985413.
- Panno, Joseph (2005). The Cell: Evolution of the First Organism. Fakta o knihovně vědeckých souborů. New York: Fakta o spisu. ISBN 978-0-8160-4946-2. LCCN 2003025841. OCLC 53901436.
- Piatigorsky, Joram; Kantorow, Marc; Gopal-Srivastava, Rashmi; Tomarev, Stanislav I. (1994). "Nábor enzymů a stresových proteinů jako krystalů čočky" V Jansson, Bengt; Jörnvall, Hans; Rydberg, Ulf; et al. (eds.). Směrem k molekulárnímu základu užívání a zneužívání alkoholu. Exs. Experientia. 71. Basilej; Boston: Birkhäuser Verlag. str. 241–50. doi:10.1007/978-3-0348-7330-7_24. ISBN 978-3-7643-2940-2. LCCN 94010167. OCLC 30030941. PMID 8032155.
- Pigliucci, Massimo; Müller, Gerd B., eds. (2010). Evoluce, rozšířená syntéza. Cambridge, Massachusetts: MIT Press. ISBN 978-0-262-51367-8. LCCN 2009024587. OCLC 804875316. Archivováno z původního dne 2015-09-18.
- Pocheville, Arnaud; Danchin, Étienne (2017). „Kapitola 3: Genetická asimilace a paradox slepých variací“. V Hunemanovi, Philippe; Walsh, Denis M. (eds.). Výzva pro moderní syntézu. New York: Oxford University Press. ISBN 978-0-19-937717-6. LCCN 2017448929. OCLC 1001337947.
- Provine, William B. (1971). Počátky teoretické populační genetiky. Chicago History of Science and Medicine (2. vyd.). Chicago, Illinois: University of Chicago Press. ISBN 978-0-226-68464-2. LCCN 2001027561. OCLC 46660910.
- Provine, William B. (1988). „Pokrok v evoluci a smysl života“. V Nitecki, Matthew H. (ed.). Evoluční pokrok. Chicago, Illinois: University of Chicago Press. ISBN 978-0-226-58693-9. LCCN 88020835. OCLC 18380658. „Tato kniha je výsledkem jarního systematického sympozia konaného v květnu 1987 v Field Museum v Chicagu.“
- Quammen, David (2006). Neochotný pan Darwin: Intimní portrét Charlese Darwina a tvorba jeho teorie evoluce. Velké objevy (1. vyd.). New York: Atlas Books /W.W. Norton & Company. ISBN 978-0-393-05981-6. LCCN 2006009864. OCLC 65400177.
- Raven, Peter H.; Johnson, George B. (2002). Biologie (6. vydání). Boston, Massachusetts: McGraw-Hill. ISBN 978-0-07-112261-0. LCCN 2001030052. OCLC 45806501.
- Ray, John (1686). Historia Plantarum [Historie rostlin]. Já. Londini: Typis Mariæ Clark. LCCN agr11000774. OCLC 2126030.
- Rechenberg, Ingo (1973). Evoluční strategie; Optimierung technischer Systeme nach Prinzipien der biologischen Evolution (Disertační práce). Problemata (v němčině). 15. Doslov od Manfred Eigen. Stuttgart-Bad Cannstatt: Frommann-Holzboog. ISBN 978-3-7728-0373-4. LCCN 74320689. OCLC 9020616.
- Ridley, Matt (1993). The Red Queen: Sex and the Evolution of Human Nature. New York: Viking. ISBN 978-0-670-84357-2. OCLC 636657988.
- Stearns, Beverly Peterson; Stearns, Stephen C. (1999). Sledování, na pokraji vyhynutí. New Haven, Connecticut: Yale University Press. ISBN 978-0-300-08469-6. LCCN 98034087. OCLC 803522914.
- Stevens, Anthony (1982). Archetyp: Přirozená historie já. Londýn: Routledge a Kegan Paul. ISBN 978-0-7100-0980-7. LCCN 84672250. OCLC 10458367.
- Voet, Donalde; Voet, Judith G.; Pratt, Charlotte W. (2016). Základy biochemie: Život na molekulární úrovni (Páté vydání). Hoboken, New Jersey: John Wiley & Sons. ISBN 978-1-11-891840-1. LCCN 2016002847. OCLC 939245154.
- West-Eberhard, Mary Jane (2003). Vývojová plasticita a evoluce. Oxford; New York: Oxford University Press. ISBN 978-0-19-512235-0. LCCN 2001055164. OCLC 48398911.
- Wiley, E. O.; Lieberman, Bruce S. (2011). Fylogenetika: Teorie a praxe fylogenetické systematiky (2. vyd.). Hoboken, New Jersey: Wiley-Blackwell. doi:10.1002/9781118017883. ISBN 978-0-470-90596-8. LCCN 2010044283. OCLC 741259265.
- Wright, Sewall (1984). Genetické a biometrické základy. Evoluce a genetika populací. 1. Chicago, Illinois: University of Chicago Press. ISBN 978-0-226-91038-3. LCCN 67025533. OCLC 246124737.
Další čtení
| Prostředky knihovny o Vývoj |
Úvodní čtení
- Barrett, Paul H .; Weinshank, Donald J .; Gottleber, Timothy T., eds. (1981). Shoda s Darwinovým původem druhů, první vydání. Ithaca, New York: Cornell University Press. ISBN 978-0-8014-1319-3. LCCN 80066893. OCLC 610057960.
- Carroll, Sean B. (2005). Endless Forms Most Beautiful: The New Science of Evo Devo and Making of the Animal Kingdom. ilustrace Jamie W. Carrolla, Josha P. Klaissa, Leanne M. Olds (1. vyd.). New York: W.W. Norton & Company. ISBN 978-0-393-06016-4. LCCN 2004029388. OCLC 57316841.
- Charlesworth, Brian; Charlesworth, Deborah (2003). Evoluce: Velmi krátký úvod. Velmi krátké představení. Oxford; New York: Oxford University Press. ISBN 978-0-19-280251-4. LCCN 2003272247. OCLC 51668497.
- Gould, Stephen Jay (1989). Wonderful Life: The Burgess Shale and the Nature of History (1. vyd.). New York: W.W. Norton & Company. ISBN 978-0-393-02705-1. LCCN 88037469. OCLC 18983518.
- Jones, Steve (1999). Téměř jako velryba: Původ druhů Aktualizováno. Londýn; New York: Doubleday. ISBN 978-0-385-40985-8. LCCN 2002391059. OCLC 41420544.
- —— (2000). Darwinův duch: Původ druhů Aktualizováno (1. vyd.). New York: Random House. ISBN 978-0-375-50103-6. LCCN 99053246. OCLC 42690131. Americká verze.
- Mader, Sylvia S. (2007). Biologie. Významné příspěvky Murray P. Pendarvisa (9. vyd.). Boston, Massachusetts: McGraw-Hill Higher Education. ISBN 978-0-07-246463-4. LCCN 2005027781. OCLC 61748307.
- Maynard Smith, John (1993). Teorie evoluce (Zpěv Canto.). Cambridge; New York: Cambridge University Press. ISBN 978-0-521-45128-4. LCCN 93020358. OCLC 27676642.
- Pallen, Mark J. (2009). Drsný průvodce evolucí. Hrubá vodítka Referenční vodítka. Londýn; New York: Drsní průvodci. ISBN 978-1-85828-946-5. LCCN 2009288090. OCLC 233547316.
Pokročilé čtení
- Barton, Nicholas H.; Briggs, Derek E.G.; Eisen, Jonathan A.; et al. (2007). Vývoj. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press. ISBN 978-0-87969-684-9. LCCN 2007010767. OCLC 86090399.
- Coyne, Jerry A .; Orr, H. Allen (2004). Speciace. Sunderland, Massachusetts: Sinauer Associates. ISBN 978-0-87893-089-0. LCCN 2004009505. OCLC 55078441.
- Bergstrom, Carl T.; Dugatkin, Lee Alan (2012). Vývoj (1. vyd.). New York: W.W. Norton & Company. ISBN 978-0-393-91341-5. LCCN 2011036572. OCLC 729341924.
- Hall, Brian K .; Olson, Wendy, eds. (2003). Klíčová slova a koncepty v evoluční vývojové biologii. Cambridge, Massachusetts: Harvard University Press. ISBN 978-0-674-00904-2. LCCN 2002192201. OCLC 50761342.
- Kauffman, Stuart A. (1993). The Origins of Order: Self-organisation and Selection in Evolution. New York; Oxford: Oxford University Press. ISBN 978-0-19-507951-7. LCCN 91011148. OCLC 895048122.
- Maynard Smith, John; Szathmáry, Eörs (1995). Hlavní přechody v evoluci. Oxford; New York: W.H. Freeman Spektrum. ISBN 978-0-7167-4525-9. LCCN 94026965. OCLC 30894392.
- Mayr, Ernst (2001). Co je to evoluce. New York: Základní knihy. ISBN 978-0-465-04426-9. LCCN 2001036562. OCLC 47443814.
- Minelli, Alessandro (2009). Formy stávání: Evoluční biologie vývoje. Překlad Mark Epstein. Princeton, New Jersey; Oxford: Princeton University Press. ISBN 978-0-691-13568-7. LCCN 2008028825. OCLC 233030259.
externí odkazy
- Zvuková pomoc
- Více mluvených článků
Obecná informace
- "Vývoj" na V naší době na BBC
- „Evoluční zdroje z národních akademií“. Washington DC: Národní akademie věd. Citováno 2011-05-30.
- „Understanding Evolution: your one-stop resource for information on evolution“. Berkeley, Kalifornie: University of California, Berkeley. Citováno 2011-05-30.
- „Evoluce evoluce - 150 let Darwinova„ O původu druhů “'". Arlington County, Virginia: Národní vědecká nadace. Citováno 2011-05-30.
- „Časová osa lidské evoluce interaktivní“. Smithsonian Institution, Národní muzeum přírodní historie. 2010-01-28. Citováno 2018-07-14. Je vyžadován Adobe Flash.
Experimenty týkající se procesu biologické evoluce
- Lenski, Richard E.. „Experimentální evoluce“. East Lansing, Michigan: Michiganská státní univerzita. Citováno 2013-07-31.
- Chastain, Erick; Livnat, Adi; Papadimitriou, Christos; Vazirani, Umesh (22. července 2014). „Algoritmy, hry a vývoj“. Proc. Natl. Acad. Sci. USA 111 (29): 10620–10623. Bibcode:2014PNAS..11110620C. doi:10.1073 / pnas.1406556111. ISSN 0027-8424. PMC 4115542. PMID 24979793.
Online přednášky
- „Přednáškový seriál Evolution Matters“. Harvardský online vzdělávací portál. Cambridge, Massachusetts: Harvardská Univerzita. Archivovány od originál dne 18. 12. 2017. Citováno 2018-07-15.
- Stearns, Stephen C.. „EEB 122: Principy evoluce, ekologie a chování“. Otevřete kurzy Yale. New Haven, Connecticut: univerzita Yale. Archivováno od původního dne 2017-12-01. Citováno 2018-07-14.
| Vývoj |  | |
|---|---|---|
| Populace genetika | ||
| Rozvoj | ||
| Z taxony | ||
| Z orgány | ||
| Z procesy | ||
| Tempo a režimy | ||
| Speciace | ||
| Dějiny | ||
| Filozofie | ||
| Příbuzný | ||
| ||