Protein Wilsonovy choroby - Wilson disease protein






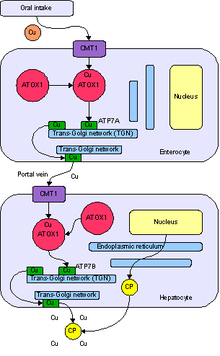

Protein Wilsonovy choroby (WND), také známý jako ATP7B protein, je přeprava mědi ATPáza typu P. který je kódován ATP7B gen. Protein ATP7B se nachází v trans-Golgiho síť jater a mozku a vyrovnává hladinu mědi v těle vylučováním přebytečné mědi do žluči a plazmy. Může způsobit genetickou poruchu genu ATP7B Wilsonova nemoc, onemocnění, při kterém se měď hromadí v tkáních, což vede k neurologický nebo psychiatrické problémy a onemocnění jater.
Gen
Protein Wilsonovy choroby je spojen s ATP7B gen, přibližně 80 kB, nacházející se na člověku chromozom 13 a skládá se z 21 exonů. MRNA přepsaná ATP7B Gen má velikost 7,5 Kb a který kóduje protein 1465 aminokyseliny.[5]
Gen je členem transportu kationtů typu P ATPáza rodina a kóduje protein s několika doménami překlenujícími membránu, ATPázu konsensuální sekvence, pantová doména, a fosforylace alespoň dva domnělé měď -závazné stránky. Tento protein funguje jako monomer a exportuje měď z buněk, jako je například odtok jaterní mědi do žluč. Alternativní transkripce varianty spoje, kódující různé izoformy s odlišnými buněčný lokalizace.[6] Wilsonova nemoc je způsobena různými mutace. Jednou z běžných mutací je mutace jednoho páru bazí, H1069Q.[5]
Struktura
Protein ATP7B transportuje měď ATPáza typu P., syntetizovaný jako membránový protein 165 KDa u člověka hepatom buněčná linie,[5] a což je 57% homologní na Menkesova choroba - asociovaný protein ATP7A.[7]
ATP7B se skládá z několika domén:
- Fosfatázová doména (TGEA motiv Thr-Gly-Glu-Ala)[5]
- Fosforylační doména (motiv DKTGT Asp-Lys-Thr-Gly-Thr)[5]
- ATP vazebná doména (Motiv TGDN)[5]
- Doména vázající kov (šest motivů vázání mědi na N-konec v cytosol )[5]
- Osm transmembránové segmenty[5]
Motiv CPC (Cys-Pro-Cys) v transmembránovém segmentu 6 charakterizuje protein jako transport těžkých kovů ATPáza.[8]
The měď vazebný motiv také vykazuje vysokou afinitu k jiným iontům přechodných kovů, jako je zinek Zn (II), kadmium Cd (II), zlato Au (III) a rtuť Hg (II). Měď je však schopna snižovat vazebnou afinitu k zinku při nízké koncentraci a dramaticky zvyšovat vazebnou afinitu k mědi se zvyšující se koncentrací, aby byla zajištěna silná vazba mezi motivem a mědí.[8]
Jako ATPázy typu P., ATP7B prochází auto-fosforylace konzervovaného klíče kyselina asparagová (D) zbytek v motivu DKTGT. Vazba ATP na protein iniciuje reakci a měď se váže na transmembránovou oblast. Poté na zbytku kyseliny asparagové v motivu DKTGT dochází k fosforylaci s uvolňováním Cu. Pak defosforylace zbytku kyseliny asparagové získá protein do stavu připraveného pro další transport.[9]
Funkce
Většina proteinu ATP7B se nachází v trans-Golgiho síť (TGN) ze dne hepatocyty, který se liší od jeho homologní protein ATP7A.[10] Malé množství ATP7B se nachází v mozek.[11]Jako protein transportující měď je jednou z hlavních funkcí dodávka mědi na enzymy závislé na mědi Golgiho aparát (např. holo-ceruloplazmin (CPN)).[10]
V lidském těle játra hraje důležitou roli v regulaci mědi včetně odstraňování mědi navíc.[10] ATP7B se účastní fyziologické cesty v procesu odstraňování mědi dvěma způsoby: vylučováním mědi plazma a vylučovat měď žluč.[7]
Interakce
ATOX1
ATP7B přijímá měď cytosolický protein antioxidant 1 měděný chaperon (ATOX1).[5] Tento protein cílí na ATP7B přímo v játrech za účelem transportu mědi. ATOX1 přenáší měď z cytosolu na doménu vázající kov ATP7B, která řídí katalytickou aktivitu ATP7B.[12]
Několik mutací v ATOX1 může blokovat dráhy mědi a způsobit ji Wilsonova nemoc.[12]
GLRX
ATP7B interaguje s glutaredoxin-1 (GLRX). Následný transport je podporován redukcí intramolekul disulfidové vazby GLRX katalýzou.[13]
Sdružení s Wilsonovou chorobou
Wilsonova choroba nastává, když dochází k hromadění mědi uvnitř jater mitochondriální poškození a ničení buněk a vykazuje příznaky onemocnění jater. Ztráta vylučování mědi žlučí pak vede ke zvyšující se koncentraci mědi v moči a způsobuje problémy s ledvinami. Proto mohou být příznaky Wilsonovy choroby různé nemoc ledvin a neurologické onemocnění.[12] Hlavní příčinou je porucha ATP7B[12] mutacemi jediného páru bází, delecemi, posuny rámců, chybami spojení ATP7B gen.[5]
Viz také
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000123191 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000006567 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b C d E F G h i j Terada K, Schilsky ML, Miura N, Sugiyama T (říjen 1998). "ATP7B (WND) protein". International Journal of Biochemistry & Cell Biology. 30 (10): 1063–7. doi:10.1016 / S1357-2725 (98) 00073-9. PMID 9785470.
- ^ „Entrez Gene: ATP7B ATPáza, transport Cu ++, beta polypeptid“.
- ^ A b Harris ED (2000). "Buněčný transport mědi a metabolismus". Každoroční přehled výživy. 20: 291–310. doi:10.1146 / annurev.nutr.20.1.291. PMID 10940336.
- ^ A b Bertini I, Gray H, Stiefel E, Valentine J (2006). Biologická anorganická chemie: struktura a reaktivita. Sausalito, CA: University Science Books. ISBN 1-891389-43-2.
- ^ Banci L, Bertini I, Cantini F, Ciofi-Baffoni S (srpen 2010). „Buněčná distribuce mědi: přístup biologie mechanistických systémů“. Buněčné a molekulární biologické vědy. 67 (15): 2563–89. doi:10.1007 / s00018-010-0330-x. PMID 20333435.
- ^ A b C Lutsenko S, LeShane ES, Shinde U (červenec 2007). „Biochemický základ regulace lidských ATPáz transportujících měď“. Archivy biochemie a biofyziky. 463 (2): 134–48. doi:10.1016 / j.abb.2007.04.013. PMC 2025638. PMID 17562324.
- ^ Crisponi G, Nurchi VM, Fanni D, Gerosa C, Nemolato S, Faa G (duben 2010). "Nemoci související s mědí: Od chemie k molekulární patologii". Recenze koordinační chemie. 254 (7–8): 876–889. doi:10.1016 / j.ccr.2009.12.018.
- ^ A b C d Cox DW, Moore SD (říjen 2002). "Měď transportující ATPázy typu P a lidská nemoc". Journal of Bioenergetics and Biomembranes. 34 (5): 333–8. doi:10.1023 / A: 1021293818125. PMID 12539960.
- ^ Lim CM, Cater MA, Mercer JF, La Fontaine S (září 2006). „Interakce glutaredoxinu závislá na mědi s N konci měď-ATPáz (ATP7A a ATP7B) defektních u Menkesových a Wilsonových chorob“ (PDF). Biochem. Biophys. Res. Commun. 348 (2): 428–36. doi:10.1016 / j.bbrc.2006.07.067. PMID 16884690.
Další čtení
- Harris ED (2000). "Buněčný transport mědi a metabolismus". Annu. Rev. Nutr. 20: 291–310. doi:10.1146 / annurev.nutr.20.1.291. PMID 10940336.
- Cox DW, Moore SD (2003). "Měď transportující ATPázy typu P a lidská nemoc". J. Bioenerg. Biomembr. 34 (5): 333–8. doi:10.1023 / A: 1021293818125. PMID 12539960.
- Lutsenko S, Efremov RG, Tsivkovskii R, Walker JM (2003). „Lidská ATPáza transportující měď ATP7B (protein Wilsonovy choroby): biochemické vlastnosti a regulace“. J. Bioenerg. Biomembr. 34 (5): 351–62. doi:10.1023 / A: 1021297919034. PMID 12539962.
- Chappuis P, Bost M, Misrahi M, Duclos-Vallée JC, Woimant F (2006). „[Wilsonova choroba: klinické a biologické aspekty]“. Ann. Biol. Clin. (Paříž). 63 (5): 457–66. PMID 16230279.
- La Fontaine S, Mercer JF (2007). „Obchodování s měď-ATPázami, ATP7A a ATP7B: role v homeostáze mědi“. Oblouk. Biochem. Biophys. 463 (2): 149–67. doi:10.1016 / j.abb.2007.04.021. PMID 17531189.
- Lutsenko S, LeShane ES, Shinde U (2007). „Biochemický základ regulace lidských ATPáz transportujících měď“. Oblouk. Biochem. Biophys. 463 (2): 134–48. doi:10.1016 / j.abb.2007.04.013. PMC 2025638. PMID 17562324.
- Banci L, Bertini I, Cantini F, Ciofi-Baffoni S (srpen 2010). „Buněčná distribuce mědi: přístup biologie mechanistických systémů“. Buněčné a molekulární biologické vědy. 67 (15): 2563–89. doi:10.1007 / s00018-010-0330-x. PMID 20333435.
externí odkazy
- GeneReviews / NIH / NCBI / UW vstup na Wilsonovu chorobu nebo hepatolentikulární degeneraci
- Wilson + nemoc + protein v americké národní lékařské knihovně Lékařské předměty (Pletivo)
Galerie PDB | |
|---|---|
|

