Koenzym Q - cytochrom c reduktáza - Coenzyme Q – cytochrome c reductase
| UCR_TM | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifikátory | |||||||||
| Symbol | UCR_TM | ||||||||
| Pfam | PF02921 | ||||||||
| InterPro | IPR004192 | ||||||||
| SCOP2 | 1be3 / Rozsah / SUPFAM | ||||||||
| TCDB | 3.D.3 | ||||||||
| OPM nadčeleď | 92 | ||||||||
| OPM protein | 3cx5 | ||||||||
| Membranome | 258 | ||||||||
| |||||||||
| ubichinol - cytochrom-c reduktáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||||
| EC číslo | 7.1.1.8 | ||||||||
| Číslo CAS | 9027-03-6 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
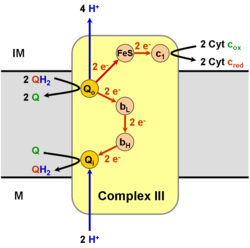
The koenzym Q: cytochrom C - oxidoreduktáza, někdy nazývané cytochrom před naším letopočtem1 komplex, a jindy komplex III, je třetím komplexem v elektronový transportní řetězec (ES 1.10.2.2 ), hrající rozhodující roli v biochemické generaci ATP (oxidační fosforylace ). Komplex III je multisubunitový transmembránový protein kódovaný jak mitochondriálními (cytochrom b ) a jaderné genomy (všechny ostatní podjednotky). Komplex III je přítomen v mitochondrie všech zvířat a všech aerobních eukaryot a vnitřních membrán většiny eubakterie. Mutace v komplexu III nesnášenlivost cvičení stejně jako multisystémové poruchy. BC1 komplex obsahuje 11 podjednotek, 3 respirační podjednotky (cytochrom B, cytochrom C1, Rieskeho protein), 2 jádra bílkoviny a 6 nízkomolekulárních bílkoviny.
Ubichinol - cytochrom-c reduktáza katalyzuje chemickou reakci
- QH2 + 2 ferricytochrom c Q + 2 ferrocytochrom c + 2 H+

Tedy dva substráty tohoto enzymu jsou chinol (QH2) a ferri- (Fe3+) cytochrom c, zatímco jeho 3 produkty jsou chinon (Q), ferro- (Fe2+) cytochrom c a H+.
Tento enzym patří do rodiny oxidoreduktázy, konkrétně ty, které působí na difenoly a příbuzné látky jako dárce s cytochromem jako akceptorem. Tento enzym se účastní oxidační fosforylace. Má čtyři kofaktory: cytochrom c1, cytochrom b-562, cytochrom b-566 a 2-železo ferredoxin z Rieske typ.
Nomenklatura
The systematické jméno této třídy enzymů je ubichinol: ferricytochrom-c oxidoreduktáza. Mezi další běžně používaná jména patří:
|
|
Struktura

Ve srovnání s ostatními hlavními podjednotkami čerpajícími protony elektronový transportní řetězec, počet nalezených podjednotek může být malý, stejně malý jako tři polypeptidové řetězce. Toto číslo se zvyšuje a jedenáct podjednotek se nachází ve vyšších zvířatech.[2] Tři podjednotky mají protetické skupiny. The cytochrom b podjednotka má dva b-typ lemy (bL a bH), cytochrom C podjednotka má jednu C-typ hemu (C1 ) a podjednotka Rieske Iron Sulphur Protein (ISP) má dvě železo, dvě síru klastr železo-síra (2Fe • 2S).
Struktury komplexu III: PDB: 1KYO, PDB: 1L0L
Složení komplexu
U obratlovců př1 komplex nebo komplex III obsahuje 11 podjednotek: 3 respirační podjednotky, 2 jádrové proteiny a 6 nízkomolekulárních proteinů.[3][4] Proteobakteriální komplexy mohou obsahovat až tři podjednotky.[5]
Tabulka podjednotkového složení komplexu III
| Ne. | Název podjednotky | Člověk protein | Popis bílkovin z UniProt | Pfam rodina s lidským proteinem |
|---|---|---|---|---|
| Proteiny respirační podjednotky | ||||
| 1 | MT-CYB / Cyt b | CYB_HUMAN | Cytochrom b | Pfam PF13631 |
| 2 | CYC1 / Cyt c1 | CY1_HUMAN | Cytochrom c1, hemový protein, mitochondriální | Pfam PF02167 |
| 3 | Rieske / UCR1 | UCRI_HUMAN | Cytochrom b-c1 komplexní podjednotka Rieske, mitochondriální ES 1.10.2.2 | Pfam PF02921 , Pfam PF00355 |
| Podjednotky jádrového proteinu | ||||
| 4 | QCR1 / SU1 | QCR1_HUMAN | Cytochrom b-c1 komplexní podjednotka 1, mitochondriální | Pfam PF00675, Pfam PF05193 |
| 5 | QCR2 / SU2 | QCR2_HUMAN | Cytochrom b-c1 komplexní podjednotka 2, mitochondriální | Pfam PF00675, Pfam PF05193 |
| Nízkomolekulární proteinové podjednotky | ||||
| 6 | QCR6 / SU6 | QCR6_HUMAN | Cytochrom b-c1 komplexní podjednotka 6, mitochondriální | Pfam PF02320 |
| 7 | QCR7 / SU7 | QCR7_HUMAN | Cytochrom b-c1 komplexní podjednotka 7 | Pfam PF02271 |
| 8 | QCR8 / SU8 | QCR8_HUMAN | Cytochrom b-C1 komplexní podjednotka 8 | Pfam PF02939 |
| 9 | QCR9 / SU9 / UCRC | QCR9_HUMANA | Cytochrom b-C1 komplexní podjednotka 9 | Pfam PF09165 |
| 10 | QCR10 / SU10 | QCR10_HUMAN | Cytochrom b-c1 komplexní podjednotka 10 | Pfam PF05365 |
| 11 | QCR11 / SU11 | QCR11_HUMAN | Cytochrom b-C1 komplexní podjednotka 11 | Pfam PF08997 |
- A U obratlovců produkt štěpení 8 kDa z N-konce proteinu Rieske (Signální peptid ) je v komplexu zachován jako podjednotka 9. Podjednotky 10 a 11 tedy odpovídají houbovým QCR9p a QCR10p.
Reakce
Katalyzuje redukci cytochrom C oxidací koenzym Q (CoQ) a souběžné čerpání 4 protony z mitochondriální matice do mezimembránového prostoru:
- QH2 + 2 cytochrom C (FeIII) + 2 H+
v → Q + 2 cytochrom C (FeII) + 4 H+
ven
V procesu tzv Q cyklus,[6][7] dva protony jsou spotřebovány z matrice (M), čtyři protony jsou uvolňovány do mezimembránového prostoru (IM) a dva elektrony jsou předávány cytochromu C.
Reakční mechanismus

Mechanismus reakce komplexu III (cytochrom bc1, koenzym Q: cytochrom C oxidoreduktáza) je známý jako ubichinonový („Q“) cyklus. V tomto cyklu se uvolní čtyři protony na pozitivní stranu „P“ (meziprostor membrány), ale pouze dva protony se zachytí z negativní strany „N“ (matice). Jako výsledek, a protonový gradient se tvoří přes membránu. V celkové reakci dva ubichinoly jsou oxidovány na ubichinony a jeden ubichinon je snížena na ubichinol. V úplném mechanismu jsou dva elektrony přeneseny z ubichinolu na ubichinon prostřednictvím dvou meziproduktů cytochromu c.
Celkově:
- 2 x QH2 oxidovaný do Q
- 1 x Q snížena do QH2
- 2 x Cyt c snížena
- 4 x H+ propuštěn do mezimembránového prostoru
- 2 x H+ vyzvednout z matice
Reakce probíhá podle následujících kroků:
1. kolo:
- Cytochrom b váže ubichinol a ubichinon.
- Centrum 2Fe / 2S a BL každý hem vytáhne elektron z vázaného ubichinolu a uvolní dva vodíky do mezimembránového prostoru.
- Jeden elektron se převede na cytochrom c1 ze středu 2Fe / 2S, zatímco další je přenesen z BL hem k B.H Heme.
- Cytochrom c1 převede svůj elektron na cytochrom c (nezaměňovat s cytochromem c1) a B.H Heme přenáší svůj elektron na blízký ubichinon, což vede k tvorbě ubisemichinonu.
- Cytochrom c difunduje. Uvolňuje se první ubichinol (nyní oxidovaný na ubichinon), zatímco semichinon zůstává vázán.
2. kolo:
- Druhý ubichinol je vázán cytochromem b.
- Centrum 2Fe / 2S a BL každý hem vytáhne elektron z vázaného ubichinolu a uvolní dva vodíky do mezimembránového prostoru.
- Jeden elektron se převede na cytochrom c1 ze středu 2Fe / 2S, zatímco další je přenesen z BL hem k B.H Heme.
- Cytocrome c1 poté převede svůj elektron na cytochrom c, zatímco blízký semichinon produkovaný z 1. kola zachytí druhý elektron z BH lem spolu se dvěma protony z matrice.
- Ubiquinol (nyní oxidovaný na ubichinon) se uvolňuje společně s nově vytvořeným ubichinolem.[8]
Inhibitory komplexu III
Existují tři odlišné skupiny inhibitorů komplexu III.
- Antimycin A váže se na Qi místo a inhibuje přenos elektronů v komplexu III z hemu bH na oxidovaný Q (inhibitor místa Qi).
- Myxothiazol a stigmatellin váže se na QÓ místo a inhibuje přenos elektronů ze snížené QH2 na sirný protein Rieske Iron. Myxothiazol a stigmatellin se vážou na odlišné, ale překrývající se kapsy v QÓ stránky.
- Myxothiazol se váže blíže na cytochrom bL (proto se označuje jako „proximální "inhibitor).
- Stigmatellin se váže dále od hemu bL a blíže sírovému proteinu Rieske Iron, se kterým silně interaguje.
Některé z nich byly komerčně dostupné jako fungicidy ( strobilurin deriváty, z nichž nejznámější je azoxystrobin; QoI inhibitory) a jako látky proti malárii (atovakvon ).
Taky propylhexedrin inhibuje cytochrom c reduktázu.[9]
Volné radikály kyslíku
Malý zlomek elektronů opustí elektronový transportní řetězec, než dosáhne komplex IV. Předčasný únik elektronů do kyslík vede k tvorbě superoxid. Relevance této jinak drobné vedlejší reakce je v tom superoxid a další reaktivní formy kyslíku jsou vysoce toxické a předpokládá se, že hrají roli při několika patologických stavech a také při stárnutí ( teorie volných radikálů stárnutí ).[10] K úniku elektronů dochází hlavně na QÓ stránky a je stimulován antimycin A. Antimycin A zamkne b hemes ve sníženém stavu tím, že zabrání jejich reoxidaci na Qi místo, které zase způsobuje ustálené koncentrace QÓ semichinon stoupat, přičemž druhý druh reaguje kyslík tvořit superoxid. Předpokládá se, že účinek vysokého membránového potenciálu má podobný účinek.[11] Superoxid produkované v místě Qo mohou být uvolňovány jak do mitochondriální matrice[12][13] a do mezimembránového prostoru, kde pak může dosáhnout cytosolu.[12][14] To by mohlo být vysvětleno skutečností, že komplex III může produkovat superoxid jako propustná pro membránu Ahoj• spíše než nepropustné pro membránu Ó−.
2.[13]
Názvy lidských genů
MT-CYB: mtDNA kódovaný cytochrom b; mutace spojené s nesnášenlivostí cvičení
CYC1: cytochrom c1
CYCS: cytochrom c
UQCRFS1: Rieskeho železný sirný protein
UQCRB: Protein vázající ubichinon, mutace spojená s nedostatkem mitochondriálního komplexu III jaderného typu 3
UQCRH: pantový protein
UQCRC2: Jádro 2, mutace spojené s nedostatkem mitochondriálního komplexu III, jaderný typ 5
UQCRC1: Jádro 1
UQCR: 6,4 kB podjednotka
UQCR10: 7,2 kB podjednotka
TTC19: Nově identifikovaná podjednotka, mutace spojené s komplexním nedostatkem III jaderného typu 2.
Mutace v komplexních genech III u lidských onemocnění
Mutace v komplexních genech souvisejících s III se obvykle projevují jako intolerance cvičení.[15][16] Bylo hlášeno, že další mutace způsobují septooptickou dysplázii[17] a multisystémové poruchy.[18] Mutace v BCS1L, gen zodpovědný za správné zrání komplexu III, může mít za následek Björnstadův syndrom a GRACILNÍ syndrom, což u novorozenců jsou smrtelné stavy, které mají multisystémové a neurologické projevy typizující závažné mitochondriální poruchy. Patogenita několika mutací byla ověřena na modelových systémech, jako jsou kvasinky.[19]
Rozsah, v jakém jsou tyto různé patologie způsobeny bioenergetickými deficity nebo nadprodukcí superoxid je v současné době neznámý.
Viz také
Další obrázky

ATD
Reference
- ^ PDB: 1ntz; Gao X, Wen X, Esser L, Quinn B, Yu L, Yu CA, Xia D (srpen 2003). „Strukturální základ pro redukci chinonu v komplexu bc1: srovnávací analýza krystalových struktur mitochondriálního cytochromu bc1 s navázaným substrátem a inhibitory v místě Qi“. Biochemie. 42 (30): 9067–80. doi:10.1021 / bi0341814. PMID 12885240.
- ^ Iwata S, Lee JW, Okada K, Lee JK, Iwata M, Rasmussen B, Link TA, Ramaswamy S, Jap BK (červenec 1998). "Kompletní struktura 11-podjednotky bovinního mitochondriálního komplexu cytochromu bc1". Věda. 281 (5373): 64–71. Bibcode:1998Sci ... 281 ... 64I. doi:10.1126 / science.281.5373.64. PMID 9651245.
- ^ Zhang Z, Huang L, Shulmeister VM, Chi YI, Kim KK, Hung LW a kol. (1998). "Přenos elektronů pohybem domény v cytochromu bc1". Příroda. 392 (6677): 677–84. Bibcode:1998 Natur.392..677Z. doi:10.1038/33612. PMID 9565029. S2CID 4380033.
- ^ Hao GF, Wang F, Li H, Zhu XL, Yang WC, Huang LS a kol. (2012). "Výpočetní objev pikomolárních inhibitorů Q (o) místa komplexu cytochromu bc1". J Am Chem Soc. 134 (27): 11168–76. doi:10.1021 / ja3001908. PMID 22690928.
- ^ Yang XH, Trumpower BL (1986). "Čištění komplexu tří podjednotek ubichinol-cytochrom c oxidoreduktáza z Paracoccus denitrificans". J Biol Chem. 261 (26): 12282–9. PMID 3017970.
- ^ Kramer DM, Roberts AG, Muller F, Cape J, Bowman MK (2004). „Reakce bypassu Q-cyklu v místě Qo komplexů cytochromu bc1 (a souvisejících)“. Chinony a chinonové enzymy, část B. Pervitin Enzymol. Metody v enzymologii. 382. 21–45. doi:10.1016 / S0076-6879 (04) 82002-0. ISBN 978-0-12-182786-1. PMID 15047094.
- ^ Crofts AR (2004). "Komplex cytochromu bc1: funkce v kontextu struktury". Annu. Physiol. 66: 689–733. doi:10.1146 / annurev.physiol.66.032102.150251. PMID 14977419.
- ^ Ferguson SJ, Nicholls D, Ferguson S (2002). Bioenergetika (3. vyd.). San Diego: Academic. str. 114–117. ISBN 978-0-12-518121-1.
- ^ Holmes, J. H .; Sapeika, N; Zwarenstein, H (1975). „Inhibiční účinek léků proti obezitě na NADH dehydrogenázu homogenátů srdce srdce“. Výzkum komunikace v chemické patologii a farmakologii. 11 (4): 645–6. PMID 241101.
- ^ Muller, F.L .; Lustgarten, M. S .; Jang, Y .; Richardson, A. & Van Remmen, H. (2007). "Trendy v teoriích oxidačního stárnutí". Volný radikál. Biol. Med. 43 (4): 477–503. doi:10.1016 / j.freeradbiomed.2007.03.034. PMID 17640558.
- ^ Skulachev VP (květen 1996). „Role nespojených a nespojených oxidací při udržování bezpečně nízkých hladin kyslíku a jeho jednoelektronových redukčních činidel“. Q. Rev. Biophys. 29 (2): 169–202. doi:10.1017 / s0033583500005795. PMID 8870073.
- ^ A b Muller F (2000). „Povaha a mechanismus produkce superoxidu v elektronovém transportním řetězci: jeho význam pro stárnutí“. STÁŘÍ. 23 (4): 227–253. doi:10.1007 / s11357-000-0022-9. PMC 3455268. PMID 23604868.
- ^ A b Muller FL, Liu Y, Van Remmen H (listopad 2004). „Komplex III uvolňuje superoxid na obě strany vnitřní mitochondriální membrány“. J. Biol. Chem. 279 (47): 49064–73. doi:10,1074 / jbc.M407715200. PMID 15317809.
- ^ Han D, Williams E, Cadenas E (leden 2001). „Generace superoxidového anionu závislá na mitochondriálním respiračním řetězci a jeho uvolňování do mezimembránového prostoru“. Biochem. J. 353 (Pt 2): 411–6. doi:10.1042/0264-6021:3530411. PMC 1221585. PMID 11139407.
- ^ DiMauro S (listopad 2006). "Mitochondriální myopatie" (PDF). Curr Opin Rheumatol. 18 (6): 636–41. doi:10.1097 / 01.bor.0000245729.17759.f2. PMID 17053512. S2CID 29140366.
- ^ DiMauro S (červen 2007). "Mitochondriální DNA lék". Biosci. Rep. 27 (1–3): 5–9. doi:10.1007 / s10540-007-9032-5. PMID 17484047. S2CID 5849380.
- ^ Schuelke M, Krude H, Finckh B, Mayatepek E, Janssen A, Schmelz M, Trefz F, Trijbels F, Smeitink J (březen 2002). „Septooptická dysplazie spojená s novou mutací mitochondriálního cytochromu b“. Ann. Neurol. 51 (3): 388–92. doi:10.1002 / ana.10151. PMID 11891837. S2CID 12425236.
- ^ Wibrand F, Ravn K, Schwartz M, Rosenberg T, Horn N, Vissing J (říjen 2001). "Multisystémová porucha spojená s missense mutací v mitochondriálním genu cytochromu b". Ann. Neurol. 50 (4): 540–3. doi:10,1002 / analog. 1224. PMID 11601507. S2CID 8944744.
- ^ Fisher N, Castleden CK, Bourges I, Brasseur G, Dujardin G, Meunier B (březen 2004). „Mutace spojené s lidskými chorobami v cytochromu b studovány v kvasinkách. J. Biol. Chem. 279 (13): 12951–8. doi:10,1074 / jbc.M313866200. PMID 14718526.
Další čtení
- Marres CM, Slater EC (1977). „Polypeptidové složení čištěného QH2: cytochrom c oxidoreduktáza z mitochondrií hovězího srdce“. Biochim. Biophys. Acta. 462 (3): 531–548. doi:10.1016/0005-2728(77)90099-8. PMID 597492.
- Rieske JS (1976). "Složení, struktura a funkce komplexu III dýchacího řetězce". Biochim. Biophys. Acta. 456 (2): 195–247. doi:10.1016/0304-4173(76)90012-4. PMID 788795.
- Wikstrom M, Krab K, Saraste M (1981). "Proton-translokační cytochromové komplexy". Annu. Biochem. 50: 623–655. doi:10.1146 / annurev.bi.50.070181.003203. PMID 6267990.
externí odkazy
- cytochrom před naším letopočtem1 komplexní web (Edward A. Berry) na Wayback Machine (archivováno 9. října 2006) na lbl.gov
- cytochrom před naším letopočtem1 komplexní web (Antony R. Crofts) na uiuc.edu
- Databáze PROMISE: cytochrom před naším letopočtem1 komplex na Archiv. Dnes (archivováno 27. srpna 1999) na scripps.edu
- Interaktivní molekulární model komplexu III na Wayback Machine (archivováno 12. ledna 2009) (vyžaduje Zvonkohra MDL )
- UMich Orientace proteinů v membránách rodiny / superrodina-3 - Vypočtené polohy bc1 a souvisejících komplexů v membránách
- Koenzym + Q-cytochrom-c + reduktáza v americké národní lékařské knihovně Lékařské předměty (Pletivo)