Mitochondriální DNA - Mitochondrial DNA
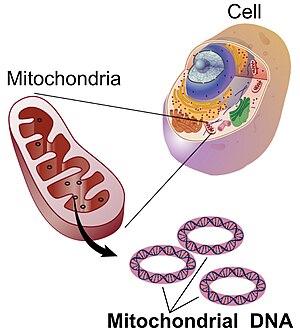
Mitochondriální DNA (mtDNA nebo mDNA)[3] je DNA nacházející se v mitochondrie, buněčný organely v rámci eukaryotický buňky, které přeměňují chemickou energii z potravin na formu, kterou mohou buňky využívat, adenosintrifosfát (ATP). Mitochondriální DNA je pouze malá část DNA v eukaryotické buňce; většina DNA se nachází v buněčné jádro a v rostlinách a řasách také v plastidy jako chloroplasty.
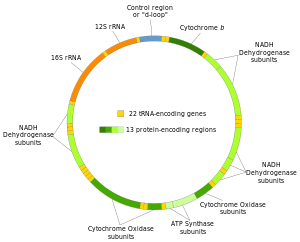
Lidská mitochondriální DNA byla první významnou součástí lidský genom být sekvenován.[4] Toto sekvenování odhalilo, že lidská mtDNA obsahuje 16 569 párů bází a kóduje 13 bílkoviny.
Protože zvířecí mtDNA se vyvíjí rychleji než nukleární genetické markery,[5][6][7] představuje oporu fylogenetika a evoluční biologie. Umožňuje také zkoumání příbuznosti populací, a proto se v roce 2006 stalo důležitým antropologie a biogeografie.
Původ
Nukleární a mitochondriální DNA jsou považovány za oddělené evoluční původu, přičemž mtDNA je odvozena z kruhových genomů bakterie které pohltily první předci dnešních eukaryotických buněk. Tato teorie se nazývá endosymbiotická teorie. V buňkách existujících organismů je drtivá většina proteinů přítomných v mitochondriích (čítající přibližně 1 500 různých typů v savci ) jsou kódovány pro nukleární DNA, ale geny pro některé, ne-li pro většinu z nich se považují za původně bakteriální, od té doby byly přeneseny do eukaryotický jádro během vývoj.[8]
Důvody, proč si mitochondrie zachovaly některé geny, jsou diskutovány. Existence organel odvozených z mitochondrií, u nichž chybí genom, u některých druhů[9] naznačuje, že je možná úplná ztráta genů a přenos mitochondriálních genů do jádra má několik výhod.[10] Obtížnost cílení na dálku produkovaných hydrofobních proteinových produktů na mitochondrii je jednou z hypotéz, proč jsou některé geny zachovány v mtDNA;[11] kolokalizace pro redoxní regulaci je další s odvoláním na vhodnost lokalizované kontroly nad mitochondriálními mechanizmy.[12] Nedávná analýza široké škály genomů mtDNA naznačuje, že oba tyto rysy mohou diktovat retenci mitochondriálních genů.[8]
Struktura a rozmanitost genomu
Ve všech organismech se v mitochondriálních genomech nachází šest hlavních typů genomu, které jsou klasifikovány podle jejich struktury (např. Kruhový versus lineární), velikosti, přítomnosti introny nebo struktury podobné plazmidům a zda je genetickým materiálem singulární molekula nebo soubor homogenní nebo heterogenní molekuly.[13]
V mnoha jednobuněčných organismech (např ciliate Tetrahymena a zelená řasa Chlamydomonas reinhardtii ), a ve vzácných případech také v mnohobuněčných organismech (např. u některých druhů rodu Cnidaria ) je mtDNA shledána lineárně uspořádanou DNA. Většina z těchto lineárních mtDNA má telomeráza -nezávislý telomery (tj. konce lineárního DNA ) s různými způsoby replikace, které z nich učinily zajímavé objekty výzkumu, protože mnoho z těchto jednobuněčných organismů s lineární mtDNA je známo patogeny.[14]
Zvířata
Většina zvířat, konkrétně bilaterálních zvířat, má kruhový mitochondriální genom. Medusozoa a calcarea subtypy však mají druhy s lineárními mitochondriálními chromozomy. [15]
Pokud jde o páry bází, sasanka Isarachnanthus nocturnus má největší mitochondriální genom ze všech zvířat na 80 923 bp.[16]
V únoru 2020 parazit související s medúzy - Henneguya salminicola - bylo objeveno, že postrádá mitochondriální genom, ale zachovává si struktury považované za organely související s mitochondrií. Navíc geny nukleární DNA zapojené do aerobního dýchání a do mitochondriální replikace a transkripce DNA buď chyběly, nebo byly přítomny pouze jako pseudogeny. Toto je první mnohobuněčný organismus, o kterém je známo, že nemá aerobní dýchání a žije zcela bez závislosti na kyslíku.[17][18]
Rostliny a houby
V rostlinách a plísních se vyskytují tři různé typy mitochondriálního genomu. První typ je kruhový genom, který má introny (typ 2) a může mít délku od 19 do 1 000 kbp. Druhým typem genomu je kruhový genom (asi 20–1 000 kbp), který má také strukturu podobnou plazmidu (1 kb) (typ 3). Konečným typem genomu, který lze nalézt v rostlinách a houbách, je lineární genom složený z homogenních molekul DNA (typ 5).
Mezi houbami a rostlinami existuje velká variace v obsahu a velikosti genu mtDNA, i když se zdá, že existuje základní podmnožina genů, které jsou přítomny ve všech eukaryotech (kromě těch několika, které vůbec nemají mitochondrie).[8]Některé druhy rostlin mají obrovské mitochondriální genomy s Silene conica mtDNA obsahující až 11 300 000 párů bází.[19] Překvapivě i ty obrovské mtDNA obsahují stejný počet a druhy genů jako příbuzné rostliny s mnohem menšími mtDNA.[20]Genom mitochondrie okurky (Cucumis sativus ) se skládá ze tří kruhových chromozomů (délky 1556, 84 a 45 kilobází), které jsou zcela nebo z velké části autonomní replikace.[21]
Protisti
Protisti obsahují nejrozmanitější mitochondriální genomy a v tomto království se vyskytuje pět různých typů. Typ 2, typ 3 a typ 5 zmíněné v rostlinných a houbových genomech existují také v některých prvcích, stejně jako dva jedinečné typy genomu. Jedním z těchto jedinečných typů je heterogenní soubor kruhových molekul DNA (typ 4), zatímco druhým je heterogenní soubor lineárních molekul (typ 6). Každý typ genomu 4 a 6 má velikost od 1–200 kbp.
Nejmenší dosud sekvenovaný mitochondriální genom je 5 967 bp mtDNA parazita Plasmodium falciparum.[22][23]
Přenos endosymbiotického genu, proces, při kterém jsou geny kódované v mitochondriálním genomu přenášeny do hlavního genomu buňky, pravděpodobně vysvětluje, proč složitější organismy, jako jsou lidé, mají menší mitochondriální genomy než jednodušší organismy, jako jsou protisté.
| Typ genomu[13] | Království | Introny | Velikost | Tvar | Popis |
|---|---|---|---|---|---|
| 1 | Zvíře | Ne | 11–28 kbp | Oběžník | Jedna molekula |
| 2 | Houby, Rostliny, Protista | Ano | 19–1 000 kbp | Oběžník | Jedna molekula |
| 3 | Houby, Rostliny, Protista | Ne | 20–1 000 kbp | Oběžník | Velká molekula a malé plasmidové struktury |
| 4 | Protista | Ne | 1–200 kbp | Oběžník | Heterogenní skupina molekul |
| 5 | Houby, Rostliny, Protista | Ne | 1–200 kbp | Lineární | Homogenní skupina molekul |
| 6 | Protista | Ne | 1–200 kbp | Lineární | Heterogenní skupina molekul |
Replikace
Mitochondriální DNA je replikována DNA polymeráza gama komplex, který je složen ze 140 kDa katalytické DNA polymerázy kódované POLG gen a dvě doplňkové podjednotky 55 kDa kódované POLG2 gen.[24] Mechanismus replisomu je tvořen DNA polymerázou, BLIKAT a mitochondriální SSB proteiny. BLIKAT je helikáza, který odvíjí krátké úseky dsDNA ve směru 5 'až 3'.[25] Všechny tyto polypeptidy jsou kódovány v jaderném genomu.
V době embryogeneze, replikace mtDNA je přísně down-regulovaná z oplodněného oocytu přes preimplantační embryo.[26] Výsledné snížení počtu kopií mtDNA na buňku hraje roli v úzkém hrdle mitochondrií a využívá variabilita mezi buňkami ke zlepšení dědičnosti škodlivých mutací.[27] Podle Justina St. Johna a kolegů: „At the blastocyst fázi je nástup replikace mtDNA specifický pro buňky trophectoderm.[26] Naproti tomu buňky vnitřní buněčná hmota omezit replikaci mtDNA, dokud neobdrží signály na odlišit na konkrétní typy buněk. “[26]

Geny na mtDNA a jejich transkripce
Dva řetězce lidské mitochondriální DNA se rozlišují jako těžké řetězce a lehké řetězce. Těžký řetězec je bohatý na guanin a kóduje 12 podjednotek systému oxidační fosforylace, dvě ribozomální RNA (12S a 16S) a 14 tRNA. Světelné vlákno kóduje jednu podjednotku a 8 tRNA. Celkově tedy mtDNA kóduje dvě rRNA, 22 tRNA a 13 proteinových podjednotek, které jsou všechny zapojeny do procesu oxidační fosforylace.[28][29]
| Gen | Typ | Produkt | Pozice v mitogenomu | Pramen |
|---|---|---|---|---|
| MT-ATP8 | kódování bílkovin | ATP syntáza, Fo podjednotka 8 (komplex V) | 08 366–0 8 572 (překrývá se s MT-ATP6) | H |
| MT-ATP6 | kódování bílkovin | ATP syntáza, Fo podjednotka 6 (komplex V) | 08 527–09 207 (překrývá se s MT-ATP8) | H |
| MT-CO1 | kódování bílkovin | Cytochrom c oxidáza podjednotka 1 (komplex IV) | 05,904–07,445 | H |
| MT-CO2 | kódování bílkovin | Cytochrom c oxidáza, podjednotka 2 (komplex IV) | 07,586–08,269 | H |
| MT-CO3 | kódování bílkovin | Cytochrom c oxidáza podjednotka 3 (komplex IV) | 09,207–09,990 | H |
| MT-CYB | kódování bílkovin | Cytochrom b (komplex III) | 14,747–15,887 | H |
| MT-ND1 | kódování bílkovin | NADH dehydrogenáza, podjednotka 1 (komplex I) | 03,307–04,262 | H |
| MT-ND2 | kódování bílkovin | NADH dehydrogenáza, podjednotka 2 (komplex I) | 04,470–05,511 | H |
| MT-ND3 | kódování bílkovin | NADH dehydrogenáza, podjednotka 3 (komplex I) | 10,059–10,404 | H |
| MT-ND4L | kódování bílkovin | NADH dehydrogenáza podjednotka 4L (komplex I) | 10 470–10 766 (překrytí s MT-ND4) | H |
| MT-ND4 | kódování bílkovin | NADH dehydrogenáza, podjednotka 4 (komplex I) | 10 760–12 137 (překrytí s MT-ND4L) | H |
| MT-ND5 | kódování bílkovin | NADH dehydrogenáza, podjednotka 5 (komplex I) | 12,337–14,148 | H |
| MT-ND6 | kódování bílkovin | NADH dehydrogenáza, podjednotka 6 (komplex I) | 14,149–14,673 | L |
| MT-RNR2 | kódování bílkovin | Humanin | — | — |
| MT-TA | přenos RNA | tRNA-Alanin (Ala nebo A) | 05,587–05,655 | L |
| MT-TR | přenos RNA | tRNA-Arginin (Arg nebo R) | 10,405–10,469 | H |
| MT-TN | přenos RNA | tRNA-Asparagin (Asn nebo N) | 05,657–05,729 | L |
| MT-TD | přenos RNA | tRNA-Kyselina asparagová (Asp nebo D) | 07,518–07,585 | H |
| MT-TC | přenos RNA | tRNA-Cystein (Cys nebo C) | 05,761–05,826 | L |
| MT-TE | přenos RNA | tRNA-Kyselina glutamová (Glu nebo E) | 14,674–14,742 | L |
| MT-TQ | přenos RNA | tRNA-Glutamin (Gln nebo Q) | 04,329–04,400 | L |
| MT-TG | přenos RNA | tRNA-Glycin (Gly nebo G) | 09,991–10,058 | H |
| MT-TH | přenos RNA | tRNA-Histidin (Jeho nebo H) | 12,138–12,206 | H |
| MT-TI | přenos RNA | tRNA-Isoleucin (Ile nebo já) | 04,263–04,331 | H |
| MT-TL1 | přenos RNA | tRNA-Leucin (Leu-UUR nebo L) | 03,230–03,304 | H |
| MT-TL2 | přenos RNA | tRNA-Leucin (Leu-CUN nebo L) | 12,266–12,336 | H |
| MT-TK | přenos RNA | tRNA-Lysin (Lys nebo K) | 08,295–08,364 | H |
| MT-TM | přenos RNA | tRNA-Methionin (Met nebo M) | 04,402–04,469 | H |
| MT-TF | přenos RNA | tRNA-Fenylalanin (Phe nebo F) | 00,577–00,647 | H |
| MT-TP | přenos RNA | tRNA-Prolin (Pro nebo P) | 15,956–16,023 | L |
| MT-TS1 | přenos RNA | tRNA-Serine (Ser-UCN nebo S) | 07,446–07,514 | L |
| MT-TS2 | přenos RNA | tRNA-Serine (Ser-AGY nebo S) | 12,207–12,265 | H |
| MT-TT | přenos RNA | tRNA-Threonin (Thr nebo T) | 15,888–15,953 | H |
| MT-TW | přenos RNA | tRNA-Tryptofan (Trp nebo W) | 05,512–05,579 | H |
| MT-TY | přenos RNA | tRNA-Tyrosin (Tyr nebo Y) | 05,826–05,891 | L |
| MT-TV | přenos RNA | tRNA-Valine (Val nebo V) | 01,602–01,670 | H |
| MT-RNR1 | ribozomální RNA | Malá podjednotka: SSU (12S) | 00,648–01,601 | H |
| MT-RNR2 | ribozomální RNA | Velká podjednotka: LSU (16S) | 01,671–03,229 | H |
Mezi většinou (ale ne všemi) oblastmi kódujícími proteiny jsou přítomny tRNA (viz mapa lidského mitochondriálního genomu ). Během transkripce získávají tRNA svůj charakteristický tvar L, který je rozpoznáván a štěpen specifickými enzymy. Při zpracování mitochondriální RNA se z primárního transkriptu uvolňují jednotlivé sekvence mRNA, rRNA a tRNA.[31] Složené tRNA proto fungují jako interpunkce sekundární struktury.[32]
Regulace transkripce
Promotory pro zahájení transkripce těžkých a lehkých řetězců jsou umístěny v hlavní nekódující oblasti mtDNA nazývané vytěsňovací smyčka, D-smyčka.[28] Existují důkazy, že transkripce mitochondriálních rRNA je regulována promotorem těžkých řetězců 1 (HSP1) a transkripce polycistronických transkriptů kódujících proteinové podjednotky reguluje HSP2.[28]
Měření hladin RNA kódovaných mtDNA v bovinních tkáních ukázalo, že existují výrazné rozdíly v expresi mitochondriálních RNA ve vztahu k celkové tkáňové RNA.[33] Mezi 12 zkoumanými tkáněmi byla nejvyšší úroveň exprese pozorována v srdci, následované vzorky mozku a steroidogenních tkání.[33]
Jak dokazuje účinek trofického hormonu ACTH na buňkách kůry nadledvin může být exprese mitochondriálních genů silně regulována vnějšími faktory, což zřejmě zvyšuje syntézu mitochondriálních proteinů nezbytných pro produkci energie.[33] Je zajímavé, že zatímco exprese genů kódujících proteiny byla stimulována ACTH, hladiny mitochondriální 16S rRNA nevykazovaly žádnou významnou změnu.[33]
Mitochondriální dědičnost
Ve většině mnohobuněčné organismy, mtDNA je zděděna od matky (zděděna po matce). Mechanismy pro toto zahrnují jednoduché ředění (vejce obsahuje v průměru 200 000 molekul mtDNA, zatímco zdravý člověk spermie údajně obsahuje v průměru 5 molekul),[34][35] degradace mtDNA spermií v mužském genitálním traktu a ve oplodněném vajíčku; a, alespoň u několika organismů, selhání mtDNA spermií vstoupit do vajíčka. Bez ohledu na mechanismus, tento jeden rodič (uniparental dědictví ) vzor dědičnosti mtDNA se vyskytuje u většiny zvířat, většiny rostlin a také u hub.
Ve výjimečných případech lidské děti někdy dědí mtDNA od svých otců i matek, což vede k mtDNA heteroplazmy.[36]
Ženské dědictví
v sexuální reprodukce, mitochondrie se obvykle dědí výhradně od matky; mitochondrie v savčích spermiích jsou po oplodnění obvykle zničeny vaječnou buňkou. Mitochondrie jsou také pouze v ocasu spermií, který se používá k pohonu spermií a někdy se ocas během oplodnění ztratí. V roce 1999 bylo oznámeno, že otcovské mitochondrie spermií (obsahující mtDNA) jsou označeny ubikvitin vybrat je pro pozdější zničení uvnitř embryo.[37] Nějaký in vitro techniky hnojení, zejména vstřikování spermatu do an oocyt, může do toho zasahovat.
Skutečnost, že mitochondriální DNA je zděděna po matce, umožňuje genealogický vědci vysledovat mateřská linie daleko v čase. (Y-chromozomální DNA, otcovsky zděděné, se používá analogickým způsobem k určení patrilineal historie.) Toho se obvykle dosáhne dne lidská mitochondriální DNA sekvenováním hypervariabilní kontrolní oblasti (HVR1 nebo HVR2), a někdy i kompletní molekula mitochondriální DNA, jako genealogický test DNA.[38] Například HVR1 se skládá z přibližně 440 párů bází. Těchto 440 párů bází se porovnává se stejnými oblastmi jiných jedinců (buď konkrétních lidí nebo subjektů v databázi), aby se určila mateřská linie. Nejčastěji se provádí srovnání s revidovaným Cambridge Referenční sekvence. Vilà et al. publikovali studie sledující matrilineální původ domácích psů od vlků.[39]Koncept Mitochondriální Eva je založen na stejném typu analýzy a pokouší se zjistit původ lidstvo sledováním rodu zpět v čase.
Úzké místo mitochondrií
Lze očekávat, že subjekty podléhající uniparentalnímu dědictví a s malou až žádnou rekombinací Mullerova ráčna, akumulace škodlivých mutací, dokud není ztracena funkčnost. Živočišné populace mitochondrií se tomu vyhýbají prostřednictvím vývojového procesu známého jako úzké místo mtDNA. Úzké místo využívá náhodné procesy v buňce zvýšit variabilitu mezi buňkami v mutantní zátěž jak se vyvíjí organismus: jediná vaječná buňka s určitým podílem mutantní mtDNA tak produkuje embryo, ve kterém mají různé buňky různé mutantní zátěže. Výběr na úrovni buněk pak může působit k odstranění těchto buněk s více mutantní mtDNA, což vede ke stabilizaci nebo snížení mutantní zátěže mezi generacemi. O mechanismu, který je základem úzkého místa, se diskutuje,[40][41][42][43] s nedávnou matematickou a experimentální metastatií poskytující důkazy o kombinaci náhodného rozdělení mtDNA při dělení buněk a náhodného obratu molekul mtDNA v buňce.[27]
Mužské dědictví
Dědičnost mužské mitochondriální DNA byla objevena v roce Kuřata Plymouth Rock.[44] Důkazy podporují vzácné případy mužské mitochondriální dědičnosti i u některých savců. Konkrétně existují zdokumentované výskyty pro myši,[45][46] kde byly mužem zděděné mitochondrie následně odmítnuty. Bylo také zjištěno u ovcí,[47] a v klonovaném skotu.[48] U lidí byly dokumentovány vzácné případy mužského mitochondriálního dědictví.[49][50][51][52] Ačkoli mnoho z těchto případů zahrnuje klonovaná embrya nebo následné odmítnutí otcovských mitochondrií, dokumentují další in vivo dědičnost a vytrvalost v laboratorních podmínkách.
U mlžů je pozorována dvojnásobně jednolodní dědičnost mtDNA. U těchto druhů mají ženy pouze jeden typ mtDNA (F), zatímco muži mají mtDNA typu F ve svých somatických buňkách, ale M typ mtDNA (který může být až 30% divergentní) v zárodečná linie buňky.[53] Otcovsky zděděné mitochondrie byly navíc hlášeny u některých druhů hmyzu, jako jsou ovocné mušky,[54][55] včely,[56] a periodické cikády.[57]
Mitochondriální darování
Technika IVF známá jako mitochondriální darování nebo mitochondriální substituční terapie (MRT) vede k potomkům obsahujícím mtDNA od dárcovské ženy a nukleární DNA od matky a otce. Při postupu vřetenového přenosu je jádro vajíčka vloženo do cytoplazmy vajíčka od dárcovské samice, u které bylo odstraněno jádro, ale stále obsahuje mtDNA dárcovské samice. Složené vejce se poté oplodní spermatem muže. Postup se používá, když žena s geneticky vadné mitochondrie si přeje plodit a plodit potomky se zdravými mitochondriemi.[58] Prvním známým dítětem, které se narodilo v důsledku darování mitochondrií, byl chlapec narozený jordánskému páru v Mexiku dne 6. dubna 2016.[59]
Mutace a nemoci
Citlivost
Koncept, na který je mtDNA obzvláště citlivý reaktivní formy kyslíku generované dýchací řetězec vzhledem ke své blízkosti zůstává kontroverzní.[60] mtDNA nehromadí žádné větší poškození oxidační báze než jaderná DNA.[61] Bylo hlášeno, že alespoň některé typy poškození oxidační DNA jsou opravovány efektivněji v mitochondriích než v jádře.[62] mtDNA je balena s proteiny, které se zdají být stejně ochranné jako proteiny nukleárního chromatinu.[63] Navíc mitochondrie vyvinuly jedinečný mechanismus, který udržuje integritu mtDNA degradací nadměrně poškozených genomů a následnou replikací intaktní / opravené mtDNA. Tento mechanismus není v jádře přítomen a je umožněn několika kopiemi mtDNA přítomnými v mitochondriích.[64] Výsledkem mutace v mtDNA může být změna v pokynech pro kódování některých proteinů,[65] které mohou mít vliv na metabolismus a / nebo kondici organismu.
Genetická nemoc
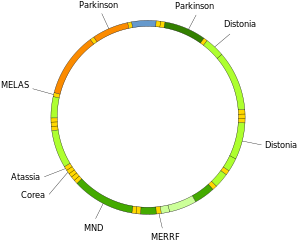
Mutace mitochondriální DNA mohou vést k řadě nemocí včetně nesnášenlivost cvičení a Kearns – Sayreův syndrom (KSS), která způsobí, že člověk ztratí plnou funkci pohybů srdce, očí a svalů. Některé důkazy naznačují, že by mohly významně přispět k procesu stárnutí a patologie spojené s věkem.[66] Zejména v souvislosti s onemocněním se označuje podíl mutantních molekul mtDNA v buňce heteroplazmy. Distribuce heteroplazmy uvnitř buněk a mezi buňkami určuje nástup a závažnost onemocnění[67] a jsou ovlivňovány komplikovanými stochastické procesy uvnitř buňky a během vývoje.[27][68]
Mutace v mitochondriálních tRNA mohou být zodpovědné za závažná onemocnění, jako je MELAS a MERRF syndromy.[69]
Mutace v jaderných genech, které kódují proteiny, které mitochondrie používají, mohou také přispět k mitochondriálním onemocněním. Tato onemocnění nenásledují mitochondriální vzory dědičnosti, ale místo toho se řídí mendelovskými vzory dědičnosti.[70]
Použití při diagnostice onemocnění
V poslední době byla k diagnostice rakoviny prostaty u pacientů s negativním nálezem použita mutace v mtDNA biopsie prostaty.[71][72]Změny mtDNA lze detekovat v bio tekutinách pacientů s rakovinou.[73]
Vztah ke stárnutí
Ačkoli je tato myšlenka kontroverzní, některé důkazy naznačují souvislost mezi stárnutím a dysfunkcí mitochondriálního genomu.[74] Mutace v mtDNA v podstatě narušily pečlivou rovnováhu reaktivní formy kyslíku (ROS) produkce a enzymatické zachycování ROS (enzymy jako superoxiddismutáza, kataláza, glutathionperoxidáza a další). Některé mutace, které zvyšují produkci ROS (např. Snížením antioxidační obrany) u červů, však jejich životnost spíše zvyšují než snižují.[60] Taky, nahé krysy, hlodavci o velikosti myši, žijí asi osmkrát déle než myši, přestože ve srovnání s myší mají sníženou antioxidační obranu a zvýšené oxidační poškození biomolekul.[75] Kdysi se myslelo, že při práci existuje smyčka pozitivní zpětné vazby („začarovaný cyklus“); protože mitochondriální DNA akumuluje genetické poškození způsobené volnými radikály, mitochondrie ztrácejí funkci a prosakují volné radikály do cytosol. Snížení mitochondriální funkce snižuje celkovou metabolickou účinnost.[76] Tento koncept však byl přesvědčivě vyvrácen, když bylo prokázáno, že myši, které byly geneticky pozměněny, aby akumulovaly mutace mtDNA zrychlenou rychlostí, stárnou předčasně, ale jejich tkáně neprodukují více ROS, jak předpovídá hypotéza „začarovaného cyklu“.[77] Některé studie podporující vazbu mezi dlouhověkostí a mitochondriální DNA našly korelace mezi biochemickými vlastnostmi mitochondriální DNA a dlouhověkostí druhů.[78] Probíhá rozsáhlý výzkum s cílem dále zkoumat tento vztah a metody boje proti stárnutí. V současné době genová terapie a nutraceutical suplementace jsou oblíbené oblasti probíhajícího výzkumu.[79][80] Bjelakovic a kol. analyzoval výsledky 78 studií mezi lety 1977 a 2012, kterých se účastnilo celkem 296 707 účastníků, a dospěl k závěru, že antioxidační doplňky nesnižují úmrtnost z jakýchkoli příčin ani prodlužují životnost, zatímco některé z nich, jako je beta karoten, vitamin E a vyšší dávky vitaminu A, může ve skutečnosti zvýšit úmrtnost.[81]
Neurodegenerativní onemocnění
Zvýšená mtPoškození DNA je vlastnost několika neurodegenerativní onemocnění.
Mozek jednotlivců s Alzheimerova choroba mít zvýšené hladiny oxidační poškození DNA v obou nukleární DNA a mtDNA, ale mtDNA má přibližně 10krát vyšší hladiny než nukleární DNA.[82] Bylo navrženo, aby stárli mitochondrie je rozhodujícím faktorem vzniku neurodegenerace u Alzheimerovy choroby.[83]
v Huntingtonova choroba mutant huntingtin protein způsobuje mitochondriální dysfunkci zahrnující inhibici mitochondrií transport elektronů, vyšší úrovně reaktivní formy kyslíku a zvýšil oxidační stres.[84] Mutantní protein huntingtin podporuje oxidační poškození mtDNA i jaderné DNA, které může přispívat k Huntingtonově chorobě patologie.[85]
The Oxidace DNA produkt 8-oxoguanin (8-oxoG) je zavedeným markerem poškození oxidační DNA. U osob s Amyotrofní laterální skleróza (ALS), enzymy, které normálně opravují poškození 8-oxoG DNA v mtDNA páteře motorické neurony jsou narušeny.[86] Oxidační poškození mtDNA motorických neuronů tedy může být významným faktorem v etiologie ALS.
Korelace základního složení mtDNA s délkou života zvířat
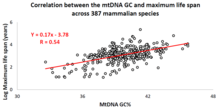
V uplynulém desetiletí to izraelská výzkumná skupina vedená profesorem Vadimem Fraifeldem ukázala jako silnou a významnou korelace mezi základním složením mtDNA a maximální délkou života specifické pro druh zvířete.[87][88][89] Jak bylo prokázáno v jejich práci, vyšší mtDNA guanin + cytosin obsah (GC% ) silně asociuje s delšími maximální životnost napříč druhy zvířat. Dalším pozorováním je, že korelace mtDNA GC% s maximální délkou života je nezávislá na dobře známé korelaci mezi rychlostí metabolismu živočišných druhů a maximální délkou života. MtDNA GC% a klidová rychlost metabolismu vysvětlují rozdíly v maximálním rozpětí života druhů zvířat multiplikativním způsobem (tj. Maximální délka života druhu = jejich rychlost metabolismu mtDNA GC% *).[88] Na podporu vědecké komunity při provádění srovnávacích analýz mezi funkcemi mtDNA a dlouhověkostí u zvířat byla vytvořena specializovaná databáze s názvem MitoAge.[90]
Vztah k jiným než B (nekanonickým) strukturám DNA
Hraniční body pro odstranění se často vyskytují v regionech nebo v jejich blízkosti a vykazují nekanonické (non-B) konformace, jmenovitě vlásenky, kříže a prvky podobné čtyřlístku.[91] Kromě toho existují data podporující zapojení vnitřně zakřivených oblastí a dlouhých G-tetrad, které narušují spirálu, při vyvolávání událostí nestability. Navíc byly konzistentně pozorovány vyšší hustoty zlomu v GC-zkosených oblastech a v těsné blízkosti degenerovaného sekvenčního motivu YMMYMNNMMHM.[92]
Použití ve forenzní oblasti
Na rozdíl od jaderné DNA, která je zděděna od obou rodičů a ve které jsou v procesu přeuspořádány geny rekombinace, v mtDNA z rodiče na potomka obvykle nedochází k žádné změně. Ačkoli mtDNA také rekombinuje, dělá to s vlastními kopiemi ve stejné mitochondrii. Z tohoto důvodu a protože rychlost mutace živočišná mtDNA je vyšší než u nukleární DNA,[93] mtDNA je mocný nástroj pro sledování předků prostřednictvím žen (matrilineage ) a v této roli se používá ke sledování předků mnoha druhů před stovkami generací.
Rychlá rychlost mutací (u zvířat) činí mtDNA užitečnou pro hodnocení genetických vztahů jednotlivců nebo skupin v rámci druhu a také pro identifikaci a kvantifikaci fylogeneze (evoluční vztahy; viz fylogenetika ) mezi různými druhy. Za tímto účelem biologové určují a poté porovnávají sekvence mtDNA od různých jedinců nebo druhů. Data z porovnání se používají ke konstrukci sítě vztahů mezi sekvencemi, která poskytuje odhad vztahů mezi jednotlivci nebo druhy, ze kterých byly mtDNA odebrány. mtDNA lze použít k odhadu vztahu mezi blízce příbuznými i vzdáleně příbuznými druhy. Vzhledem k vysoké rychlosti mutace mtDNA u zvířat se 3. pozice kodonů mění relativně rychle, a poskytují tak informace o genetických vzdálenostech mezi blízce příbuznými jedinci nebo druhy. Na druhou stranu je míra substituce mt-proteinů velmi nízká, proto se změny aminokyselin hromadí pomalu (s odpovídajícími pomalými změnami v pozicích 1. a 2. kodonu), a tak poskytují informace o genetických vzdálenostech vzdáleně příbuzných druhů. Statistické modely, které léčí substituční rychlosti mezi pozicemi kodonů samostatně, lze tedy použít k současnému odhadu fylogenií, které obsahují jak blízce, tak vzdáleně příbuzné druhy[69]
V roce 1996 byla mitochondriální DNA poprvé v soudní síni Spojených států přijata jako důkaz Stát Tennessee v. Paul Ware.[94]
V soudním sporu Spojených států z roku 1998 ve státě Pennsylvania v. Patricia Lynne Rorrer,[95] mitochondriální DNA byla poprvé přijata jako důkaz ve státě Pensylvánie.[96][97] Případ byl uveden v epizodě 55 sezóny 5 seriálu skutečných krimi Soudní spisy (sezóna 5).[98]
Mitochondriální DNA byla poprvé přijata jako důkaz v roce Kalifornie, USA, v úspěšném stíhání Davida Westerfielda za únos a vraždu sedmiletého dítěte z roku 2002 Danielle van Dam v San Diego: bylo použito pro identifikaci člověka i psa.[99] Jednalo se o první pokus v USA, který připustil psí DNA.[100]
Pozůstatky Král Richard III byly identifikovány porovnáním jeho mtDNA se dvěma matrilineálními potomky jeho sestry.[101]
Využití v evoluční biologii a systematické biologii
mtDNA je konzervována napříč eukaryotickým organismem vzhledem k zásadní roli mitochondrií v buněčné dýchání. Kvůli méně účinné opravě DNA (ve srovnání s jadernou DNA) má však relativně vysokou rychlost mutace (ale pomalá ve srovnání s jinými oblastmi DNA, jako je mikrosatelity ) což je užitečné pro studium evolučních vztahů -fylogeneze —Organismů. Biologové mohou určit a poté porovnat sekvence mtDNA mezi různými druhy a použít srovnání k vytvoření evoluční strom pro zkoumaný druh.
Například zatímco většina jaderné geny jsou téměř totožné mezi lidmi a šimpanzi, jejich mitochondriální genomy se liší o 9,8%. Lidské a gorila mitochondriální genomy se liší o 11,8%, což ukazuje, že jsme více příbuzní šimpanzům než gorilám.[102]
Dějiny
Mitochondriální DNA objevili v 60. letech Margit M. K. Nass a Sylvan Nass elektronová mikroskopie jako vlákna citlivá na DNase uvnitř mitochondrií,[103] a Ellen Haslbrunner, Hans Tuppy a Gottfried Schatz biochemickými testy na vysoce purifikovaných mitochondriálních frakcích.[104]
Mitochondriální databáze sekvencí
Ke shromažďování sekvencí mitochondriálního genomu a dalších informací bylo založeno několik specializovaných databází. Ačkoli se většina z nich zaměřuje na sekvenční data, některé z nich obsahují fylogenetické nebo funkční informace.
- AmtDB: databáze starověkých lidských mitochondriálních genomů.[105]
- InterMitoBase: anotovaná databáze a analytická platforma interakcí protein-protein pro lidské mitochondrie.[106] (zřejmě naposledy aktualizováno v roce 2010, ale stále k dispozici)
- MitoBreak: databáze hraničních bodů mitochondriální DNA.[107]
- MitoFish a MitoAnnotator: mitochondriální genomová databáze ryb.[108] Viz také Cawthorn a kol.[109]
- Mitome: databáze srovnávací mitochondriální genomiky u metazoanských zvířat[110] (již není k dispozici)
- MitoRes: zdroj jaderných kódovaných mitochondriálních genů a jejich produktů v metazoa[111] (zjevně se již neaktualizuje)
- MitoSatPlant: Mitochondriální databáze mikrosatelitů viridiplantae.[112]
- MitoZoa 2.0: databáze pro srovnávací a evoluční analýzy mitochondriálních genomů v Metazoa.[113] (již není k dispozici)
Databáze mitochondriálních mutací
Existuje několik specializovaných databází, které uvádějí polymorfismy a mutace v lidské mitochondriální DNA spolu s hodnocením jejich patogenity.
- MitImpact: Sbírka předpočítaných předpovědí patogenity pro všechny změny nukleotidů, které způsobují nesynonymní substituce v genech kódujících lidský mitochondriální protein [3].
- MITOMAPA: Kompendium polymorfismů a mutací v lidské mitochondriální DNA [4].
Viz také
Reference
- ^ Siekevitz P (1957). "Powerhouse buňky". Scientific American. 197 (1): 131–40. Bibcode:1957SciAm.197a.131S. doi:10.1038 / scientificamerican0757-131.
- ^ Iborra FJ, Kimura H, Cook PR (květen 2004). „Funkční organizace mitochondriálních genomů v lidských buňkách“. Biologie BMC. 2: 9. doi:10.1186/1741-7007-2-9. PMC 425603. PMID 15157274.
- ^ Sykes B. (10. září 2003). "Mitochondriální DNA a historie člověka". Lidský genom. Wellcome Trust. Archivovány od originál dne 7. září 2015. Citováno 5. února 2012.
- ^ Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJ, Staden R, Young IG. (1981). "Pořadí a organizace lidského mitochondriálního genomu". Příroda. 290 (5806): 457–65. Bibcode:1981 Natur.290..457A. doi:10.1038 / 290457a0. PMID 7219534. S2CID 4355527.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Boursot P, Bonhomme F (1. ledna 1986). "[Není dostupný]". Génétique, Sélection, Évolution. 18 (1): 73–98. doi:10.1186/1297-9686-18-1-73. PMC 2713894. PMID 22879234.
- ^ Delsuc F, Stanhope MJ, Douzery EJ (srpen 2003). „Molekulární systematika pásovců (Xenarthra, Dasypodidae): příspěvek maximální věrohodnosti a Bayesovské analýzy mitochondriálních a jaderných genů“. Molekulární fylogenetika a evoluce. 28 (2): 261–75. doi:10.1016 / s1055-7903 (03) 00111-8. PMID 12878463.
- ^ Hassanin A, An J, Ropiquet A, Nguyen TT, Couloux A (březen 2013). "Kombinace více autosomálních intronů pro studium mělké fylogeneze a taxonomie laurasiatherských savců: Aplikace na kmen Bovini (Cetartiodactyla, Bovidae)". Molekulární fylogenetika a evoluce. 66 (3): 766–75. doi:10.1016 / j.ympev.2012.11.003. PMID 23159894.
- ^ A b C Johnston IG, Williams BP (únor 2016). „Evoluční závěr napříč eukaryoty identifikuje specifické tlaky upřednostňující retenci mitochondriálních genů“. Buněčné systémy. 2 (2): 101–11. doi:10.1016 / j.cels.2016.01.013. PMID 27135164.
- ^ van der Giezen M, Tovar J, Clark CG (2005). „Organely odvozené od mitochondrií v protistech a houbách“. Průzkum buněčné biologie. International Review of Cytology. 244. 175–225. doi:10.1016 / S0074-7696 (05) 44005-X. ISBN 978-0-12-364648-4. PMID 16157181.
- ^ Adams KL, Palmer JD (prosinec 2003). „Vývoj obsahu mitochondriálních genů: ztráta genů a přenos do jádra“. Molekulární fylogenetika a evoluce. 29 (3): 380–95. doi:10.1016 / S1055-7903 (03) 00194-5. PMID 14615181.
- ^ Björkholm P, Harish A, Hagström E, Ernst AM, Andersson SG (srpen 2015). „Mitochondriální genomy jsou zachovány díky selektivním omezením cílení na proteiny“. Sborník Národní akademie věd Spojených států amerických. 112 (33): 10154–61. Bibcode:2015PNAS..11210154B. doi:10.1073 / pnas.1421372112. PMC 4547212. PMID 26195779.
- ^ Allen JF (srpen 2015). „Proč si chloroplasty a mitochondrie zachovávají své vlastní genomy a genetické systémy: Kolokace pro redoxní regulaci genové exprese“. Sborník Národní akademie věd Spojených států amerických. 112 (33): 10231–8. Bibcode:2015PNAS..11210231A. doi:10.1073 / pnas.1500012112. PMC 4547249. PMID 26286985.
- ^ A b Kolesnikov AA, Gerasimov ES (prosinec 2012). "Rozmanitost organizace mitochondriálního genomu". Biochemie. Biokhimiia. 77 (13): 1424–35. doi:10.1134 / S0006297912130020. PMID 23379519. S2CID 14441187.
- ^ Nosek J, Tomáska L, Fukuhara H, Suyama Y, Kovác L (květen 1998). "Lineární mitochondriální genomy: 30 let po linii". Trendy v genetice. 14 (5): 184–8. doi:10.1016 / S0168-9525 (98) 01443-7. PMID 9613202.
- ^ Lavrov, Dennis V .; Pett, Walker (26. září 2016). „Zvířecí mitochondriální DNA, jak ji neznáme: Organizace mt-genomu a evoluce v nonbilaterálních liniích“. Biologie genomu a evoluce. 8 (9): 2896–2913. doi:10.1093 / gbe / evw195. ISSN 1759-6653. PMC 5633667. PMID 27557826.
- ^ Stampar, Sérgio N .; Broe, Michael B .; Macrander, Jason; Reitzel, Adam M .; Brugler, Mercer R .; Daly, Marymegan (15. dubna 2019). „Lineární mitochondriální genom v Anthozoa (Cnidaria): Případová studie v Ceriantharia“. Vědecké zprávy. 9. doi:10.1038 / s41598-019-42621-z. ISSN 2045-2322. PMC 6465557. PMID 30988357.
- ^ Yahalomi, Dayana; Atkinson, Stephen D .; Neuhof, Moran; Chang, E. Sally; Philippe, Hervé; Cartwright, Paulyn; Bartoloměj, Jerri L .; Huchon, Dorothée (10. března 2020). „Cnidarský parazit z lososa (Myxozoa: Henneguya) postrádá mitochondriální genom “. Sborník Národní akademie věd. 117 (10): 5358–5363. doi:10.1073 / pnas.1909907117. PMC 7071853. PMID 32094163.
- ^ Starr, Michelle. „Vědci najdou vůbec první zvíře, které k přežití nepotřebuje kyslík“. ScienceAlert. Citováno 25. února 2020.
- ^ Sloan DB, Alverson AJ, Chuckalovcak JP, Wu M, McCauley DE, Palmer JD, Taylor DR (leden 2012). „Rychlý vývoj obrovských multichromozomálních genomů v mitochondriích kvetoucích rostlin s výjimečně vysokou mírou mutací“. PLOS Biology. 10 (1): e1001241. doi:10.1371 / journal.pbio.1001241. PMC 3260318. PMID 22272183.
- ^ Ward BL, Anderson RS, Bendich AJ (září 1981). „Mitochondriální genom je velký a proměnlivý v rodině rostlin (cucurbitaceae)“. Buňka. 25 (3): 793–803. doi:10.1016/0092-8674(81)90187-2. PMID 6269758. S2CID 45090891.
- ^ Alverson AJ, Rice DW, Dickinson S, Barry K, Palmer JD (červenec 2011). „Počátky a rekombinace multichromozomálního mitochondriálního genomu okurky o velikosti bakterií“. Rostlinná buňka. 23 (7): 2499–513. doi:10.1105 / tpc.111.087189. JSTOR 41433488. PMC 3226218. PMID 21742987.
- ^ „Mitochondriální DNA (mtDNA)“ (PDF). Integrované technologie DNA. Archivovány od originál (PDF) dne 29. července 2016. Citováno 25. února 2016.
- ^ Tyagi S, Pande V, Das A (únor 2014). "Celá sekvence mitochondriálního genomu indického izolátu pole Plasmodium falciparum". Korejský žurnál parazitologie. 52 (1): 99–103. doi:10.3347 / kjp.2014.52.1.99. PMC 3949004. PMID 24623891.
- ^ Yakubovskaya E, Chen Z, Carrodeguas JA, Kisker C, Bogenhagen DF (leden 2006). „Funkční lidská mitochondriální DNA polymeráza gama tvoří heterotrimer“. The Journal of Biological Chemistry. 281 (1): 374–82. doi:10,1074 / jbc.M509730200. PMID 16263719.
- ^ Jemt E, Farge G, Bäckström S, Holmlund T, Gustafsson CM, Falkenberg M (Listopad 2011). „Mitochondriální DNA helikáza TWINKLE se může shromažďovat na uzavřeném kruhovém templátu a podporovat zahájení syntézy DNA“. Výzkum nukleových kyselin. 39 (21): 9238–49. doi:10.1093 / nar / gkr653. PMC 3241658. PMID 21840902.
- ^ A b C St John JC, Facucho-Oliveira J, Jiang Y, Kelly R, Salah R (2010). „Mitochondriální přenos DNA, replikace a dědičnost: cesta z gamety embryem a do potomků a embryonálních kmenových buněk“. Aktualizace lidské reprodukce. 16 (5): 488–509. doi:10.1093 / humupd / dmq002. PMID 20231166.
- ^ A b C Johnston IG, Burgstaller JP, Havlicek V, Kolbe T, Rülicke T, Brem G, Poulton J, Jones NS (červen 2015). „Stochastické modelování, Bayesiánská inference a nová měření in vivo objasňují diskutovaný mechanismus úzkého místa mtDNA“. eLife. 4: e07464. doi:10,7554 / eLife.07464. PMC 4486817. PMID 26035426.
- ^ A b C Barshad G, Marom S, Cohen T, Mishmar D (2018). "Mitochondrial DNA Transcription and Its Regulation: An Evolutionary Perspective". Trendy Genet. 34 (9): 682–692. doi:10.1016/j.tig.2018.05.009. PMID 29945721.
- ^ Barchiesi A, Vascotto C (2019). "Transcription, Processing, and Decay of Mitochondrial RNA in Health and Disease". Int J Mol Sci. 20 (9): 2221. doi:10.3390/ijms20092221. PMC 6540609. PMID 31064115.
- ^ Homo sapiens mitochondrion, complete genome. "Revised Cambridge Reference Sequence (rCRS): accession NC_012920", Národní centrum pro biotechnologické informace. Citováno dne 20. února 2017.
- ^ Falkenberg M, Larsson NG, Gustafsson CM (19 June 2007). "DNA replication and transcription in mammalian mitochondria". Roční přehled biochemie. 76 (1): 679–99. doi:10.1146/annurev.biochem.76.060305.152028. PMID 17408359.
- ^ Ojala D, Montoya J, Attardi G (April 1981). "tRNA punctuation model of RNA processing in human mitochondria". Příroda. 290 (5806): 470–4. Bibcode:1981Natur.290..470O. doi:10.1038/290470a0. PMID 7219536. S2CID 4323371.
- ^ A b C d Raikhinstein M, Hanukoglu I (listopad 1993). „RNA kódované mitochondriálním genomem: diferenciální regulace kortikotropinem v bovinních adrenokortikálních buňkách“. Sborník Národní akademie věd Spojených států amerických. 90 (22): 10509–13. Bibcode:1993PNAS ... 9010509R. doi:10.1073 / pnas.90.22.10509. PMC 47806. PMID 7504267.
- ^ Wolff JN, Gemmell NJ (November 2008). "Lost in the zygote: the dilution of paternal mtDNA upon fertilization". Dědičnost. 101 (5): 429–34. doi:10.1038/hdy.2008.74. PMID 18685570. S2CID 5065133.
- ^ Gabriel MS, Chan SW, Alhathal N, Chen JZ, Zini A (August 2012). "Influence of microsurgical varicocelectomy on human sperm mitochondrial DNA copy number: a pilot study". Journal of Assisted Reproduction and Genetics. 29 (8): 759–64. doi:10.1007/s10815-012-9785-z. PMC 3430774. PMID 22562241.
- ^ Luo S, Valencia CA, Zhang J, Lee NC, Slone J, Gui B, Wang X, Li Z, Dell S, Brown J, Chen SM, Chien YH, Hwu WL, Fan PC, Wong LJ, Atwal PS, Huang T (December 2018). "Biparental Inheritance of Mitochondrial DNA in Humans". Sborník Národní akademie věd Spojených států amerických. 115 (51): 13039–13044. doi:10.1073/pnas.1810946115. PMC 6304937. PMID 30478036.
- ^ Sutovsky P, Moreno RD, Ramalho-Santos J, Dominko T, Simerly C, Schatten G (November 1999). "Ubiquitin tag for sperm mitochondria". Příroda. 402 (6760): 371–2. Bibcode:1999Natur.402..371S. doi:10.1038/46466. PMID 10586873. S2CID 205054671. Discussed in: Travis, John (2000). "Mom's Eggs Execute Dad's Mitochondria". Vědecké zprávy. 157 (1): 5. doi:10.2307/4012086. JSTOR 4012086. Archivovány od originál dne 19. prosince 2007.
- ^ "Hiring a DNA Testing Company Genealogy". Hledání rodiny. Církev Ježíše Krista Svatých posledních dnů. 2016. Citováno 2. října 2016.
- ^ Vilà C, Savolainen P, Maldonado JE, Amorim IR, Rice JE, Honeycutt RL, Crandall KA, Lundeberg J, Wayne RK (June 1997). "Mnohočetný a starověký původ domácího psa". Věda. 276 (5319): 1687–9. doi:10.1126/science.276.5319.1687. PMID 9180076.
- ^ Wolff JN, White DJ, Woodhams M, White HE, Gemmell NJ (2011). "The strength and timing of the mitochondrial bottleneck in salmon suggests a conserved mechanism in vertebrates". PLOS ONE. 6 (5): e20522. Bibcode:2011PLoSO...620522W. doi:10.1371/journal.pone.0020522. PMC 3105079. PMID 21655224.
- ^ Cree LM, Samuels DC, de Sousa Lopes SC, Rajasimha HK, Wonnapinij P, Mann JR, Dahl HH, Chinnery PF (February 2008). "A reduction of mitochondrial DNA molecules during embryogenesis explains the rapid segregation of genotypes". Genetika přírody. 40 (2): 249–54. doi:10.1038/ng.2007.63. PMID 18223651. S2CID 205344980.
- ^ Cao L, Shitara H, Horii T, Nagao Y, Imai H, Abe K, Hara T, Hayashi J, Yonekawa H (March 2007). "The mitochondrial bottleneck occurs without reduction of mtDNA content in female mouse germ cells". Genetika přírody. 39 (3): 386–90. doi:10.1038/ng1970. PMID 17293866. S2CID 10686347.
- ^ Wai T, Teoli D, Shoubridge EA (December 2008). "The mitochondrial DNA genetic bottleneck results from replication of a subpopulation of genomes". Genetika přírody. 40 (12): 1484–8. doi:10.1038/ng.258. PMID 19029901. S2CID 225349.
- ^ Alexander M, Ho SY, Molak M, Barnett R, Carlborg Ö, Dorshorst B, Honaker C, Besnier F, Wahlberg P, Dobney K, Siegel P, Andersson L, Larson G (October 2015). "Mitogenomic analysis of a 50-generation chicken pedigree reveals a rapid rate of mitochondrial evolution and evidence for paternal mtDNA inheritance". Biologické dopisy. 11 (10): 20150561. doi:10.1098/rsbl.2015.0561. PMC 4650172. PMID 26510672.
- ^ Gyllensten U, Wharton D, Josefsson A, Wilson AC (July 1991). "Paternal inheritance of mitochondrial DNA in mice". Příroda. 352 (6332): 255–7. Bibcode:1991Natur.352..255G. doi:10.1038/352255a0. PMID 1857422. S2CID 4278149.
- ^ Shitara H, Hayashi JI, Takahama S, Kaneda H, Yonekawa H (February 1998). "Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage". Genetika. 148 (2): 851–7. PMC 1459812. PMID 9504930.
- ^ Zhao X, Li N, Guo W, Hu X, Liu Z, Gong G, Wang A, Feng J, Wu C (October 2004). "Further evidence for paternal inheritance of mitochondrial DNA in the sheep (Ovis aries)". Dědičnost. 93 (4): 399–403. doi:10.1038/sj.hdy.6800516. PMID 15266295.
- ^ Steinborn R, Zakhartchenko V, Jelyazkov J, Klein D, Wolf E, Müller M, Brem G (April 1998). "Composition of parental mitochondrial DNA in cloned bovine embryos". FEBS Dopisy. 426 (3): 352–6. doi:10.1016/S0014-5793(98)00350-0. PMID 9600265.
- ^ Singh AN (April 2012). "A105 Family Decoded: Discovery of Genome-Wide Fingerprints for Personalized Genomic Medicine". ScienceMED. 3 (2): 115–126.
- ^ Singh AN (May 2018). "Customized Biomedical Informatics". BMC Big Data Analytics. 3. doi:10.1186/s41044-018-0030-3.
- ^ Schwartz M, Vissing J (August 2002). "Paternal inheritance of mitochondrial DNA". The New England Journal of Medicine. 347 (8): 576–80. doi:10.1056/NEJMoa020350. PMID 12192017.
- ^ Luo S, Valencia CA, Zhang J, Lee NC, Slone J, Gui B, Wang X, Li Z, Dell S, Brown J, Chen SM, Chien YH, Hwu WL, Fan PC, Wong LJ, Atwal PS, Huang T (December 2018). "Biparental Inheritance of Mitochondrial DNA in Humans". Sborník Národní akademie věd Spojených států amerických. 115 (51): 13039–13044. doi:10.1073/pnas.1810946115. PMC 6304937. PMID 30478036.
- ^ Passamonti M, Ghiselli F (February 2009). "Doubly uniparental inheritance: two mitochondrial genomes, one precious model for organelle DNA inheritance and evolution". DNA a buněčná biologie. 28 (2): 79–89. doi:10.1089/dna.2008.0807. PMID 19196051.
- ^ Kondo R, Matsuura ET, Chigusa SI (April 1992). "Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method". Genetický výzkum. 59 (2): 81–4. doi:10.1017/S0016672300030287. PMID 1628820.
- ^ Wolff JN, Nafisinia M, Sutovsky P, Ballard JW (January 2013). "Paternal transmission of mitochondrial DNA as an integral part of mitochondrial inheritance in metapopulations of Drosophila simulans". Dědičnost. 110 (1): 57–62. doi:10.1038/hdy.2012.60. PMC 3522233. PMID 23010820.
- ^ Meusel MS, Moritz RF (December 1993). "Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs". Současná genetika. 24 (6): 539–43. doi:10.1007/BF00351719. PMID 8299176. S2CID 15621371.
- ^ Fontaine KM, Cooley JR, Simon C (September 2007). "Evidence for paternal leakage in hybrid periodical cicadas (Hemiptera: Magicicada spp.)". PLOS ONE. 2 (9): e892. Bibcode:2007PLoSO...2..892F. doi:10.1371/journal.pone.0000892. PMC 1963320. PMID 17849021.
- ^ Frith M (14 October 2003). "Ban on scientists trying to create three-parent baby". Nezávislý.
- ^ Roberts M (27 September 2016). "First 'three person baby' born using new method". BBC novinky. Citováno 28. září 2016.
- ^ A b Alexeyev MF (October 2009). "Is there more to aging than mitochondrial DNA and reactive oxygen species?". FEBS Journal. 276 (20): 5768–87. doi:10.1111/j.1742-4658.2009.07269.x. PMC 3097520. PMID 19796285.
- ^ Anson RM, Hudson E, Bohr VA (February 2000). "Mitochondrial endogenous oxidative damage has been overestimated". FASEB Journal. 14 (2): 355–60. doi:10.1096/fasebj.14.2.355. PMID 10657991.
- ^ Thorslund T, Sunesen M, Bohr VA, Stevnsner T (April 2002). "Repair of 8-oxoG is slower in endogenous nuclear genes than in mitochondrial DNA and is without strand bias". Oprava DNA. 1 (4): 261–73. doi:10.1016/S1568-7864(02)00003-4. PMID 12509245.
- ^ Guliaeva NA, Kuznetsova EA, Gaziev AI (2006). "[Proteins associated with mitochondrial DNA protect it against the action of X-rays and hydrogen peroxide]" [Proteins associated with mitochondrial DNA protect it against the action of X-rays and hydrogen peroxide]. Biofizika (v Rusku). 51 (4): 692–7. PMID 16909848.
- ^ Alexeyev M, Shokolenko I, Wilson G, LeDoux S (květen 2013). „Zachování integrity mitochondriální DNA - kritická analýza a aktualizace“. Perspektivy Cold Spring Harbor v biologii. 5 (5): a012641. doi:10.1101 / cshperspect.a012641. PMC 3632056. PMID 23637283.
- ^ Hogan CM (2010). "Mutace". In Monosson E, Cleveland CJ (eds.). Encyklopedie Země. Washington DC: National Council for Science and the Environment.
- ^ Alexeyev MF, Ledoux SP, Wilson GL (October 2004). "Mitochondrial DNA and aging". Klinická věda. 107 (4): 355–64. doi:10.1042/CS20040148. PMID 15279618. S2CID 5747202.
- ^ Burgstaller JP, Johnston IG, Poulton J (January 2015). "Mitochondrial DNA disease and developmental implications for reproductive strategies". Molekulární lidská reprodukce. 21 (1): 11–22. doi:10.1093/molehr/gau090. PMC 4275042. PMID 25425607.
- ^ Burgstaller JP, Johnston IG, Jones NS, Albrechtová J, Kolbe T, Vogl C, Futschik A, Mayrhofer C, Klein D, Sabitzer S, Blattner M, Gülly C, Poulton J, Rülicke T, Piálek J, Steinborn R, Brem G (June 2014). "MtDNA segregation in heteroplasmic tissues is common in vivo and modulated by haplotype differences and developmental stage". Zprávy buněk. 7 (6): 2031–2041. doi:10.1016/j.celrep.2014.05.020. PMC 4570183. PMID 24910436.
- ^ A b Taylor RW, Turnbull DM (May 2005). "Mitochondriální DNA mutace u lidských nemocí". Genetika hodnocení přírody. 6 (5): 389–402. doi:10.1038 / nrg1606. PMC 1762815. PMID 15861210.
- ^ Angelini C, Bello L, Spinazzi M, Ferrati C (July 2009). "Mitochondrial disorders of the nuclear genome". Acta Myologica. 28 (1): 16–23. PMC 2859630. PMID 19772191.
- ^ Reguly B, Jakupciak JP, Parr RL (October 2010). "3.4 kb mitochondrial genome deletion serves as a surrogate predictive biomarker for prostate cancer in histopathologically benign biopsy cores". Kanadský deník urologické asociace. 4 (5): E118-22. doi:10.5489/cuaj.932. PMC 2950771. PMID 20944788.
- ^ Robinson K, Creed J, Reguly B, Powell C, Wittock R, Klein D, Maggrah A, Klotz L, Parr RL, Dakubo GD (June 2010). "Accurate prediction of repeat prostate biopsy outcomes by a mitochondrial DNA deletion assay". Rakovina prostaty a prostatické nemoci. 13 (2): 126–31. doi:10.1038/pcan.2009.64. PMID 20084081.
- ^ Mair, R (2019). "Measurement of plasma cell-free mitochondrial tumor DNA improves detection of glioblastoma in patient-derived orthotopic xenograft models". Výzkum rakoviny. 79 (1): 220–230. doi:10.1158/0008-5472.CAN-18-0074. PMC 6753020. PMID 30389699.
- ^ de Grey A (1999). Mitochondriální volná radikální teorie stárnutí (PDF). ISBN 978-1-57059-564-6.[stránka potřebná ]
- ^ Lewis KN, Andziak B, Yang T, Buffenstein R (October 2013). "The naked mole-rat response to oxidative stress: just deal with it". Antioxidanty a redoxní signalizace. 19 (12): 1388–99. doi:10.1089/ars.2012.4911. PMC 3791056. PMID 23025341.
- ^ Shigenaga MK, Hagen TM, Ames BN (November 1994). "Oxidative damage and mitochondrial decay in aging". Sborník Národní akademie věd Spojených států amerických. 91 (23): 10771–8. Bibcode:1994PNAS...9110771S. doi:10.1073/pnas.91.23.10771. JSTOR 2365473. PMC 45108. PMID 7971961.
- ^ Trifunovic A, Hansson A, Wredenberg A, Rovio AT, Dufour E, Khvorostov I, Spelbrink JN, Wibom R, Jacobs HT, Larsson NG (December 2005). "Somatic mtDNA mutations cause aging phenotypes without affecting reactive oxygen species production". Sborník Národní akademie věd Spojených států amerických. 102 (50): 17993–8. Bibcode:2005PNAS..10217993T. doi:10.1073/pnas.0508886102. JSTOR 4152716. PMC 1312403. PMID 16332961.
- ^ Aledo JC, Li Y, de Magalhães JP, Ruíz-Camacho M, Pérez-Claros JA (April 2011). "Mitochondrially encoded methionine is inversely related to longevity in mammals". Stárnoucí buňka. 10 (2): 198–207. doi:10.1111/j.1474-9726.2010.00657.x. PMID 21108730.
- ^ Ferrari CK (2004). "Functional foods, herbs and nutraceuticals: towards biochemical mechanisms of healthy aging". Biogerontologie. 5 (5): 275–89. doi:10.1007/s10522-004-2566-z. PMID 15547316. S2CID 11568208.
- ^ Taylor RW (February 2005). "Gene therapy for the treatment of mitochondrial DNA disorders". Znalecký posudek na biologickou terapii. 5 (2): 183–94. doi:10.1517/14712598.5.2.183. PMID 15757380. S2CID 35276183.
- ^ Bjelakovic G, Nikolova D, Gluud C (September 2013). "Antioxidant supplements to prevent mortality". JAMA. 310 (11): 1178–9. doi:10.1001/jama.2013.277028. PMID 24045742.
- ^ Wang J, Xiong S, Xie C, Markesbery WR, Lovell MA (May 2005). "Increased oxidative damage in nuclear and mitochondrial DNA in Alzheimer's disease". Journal of Neurochemistry. 93 (4): 953–62. doi:10.1111/j.1471-4159.2005.03053.x. PMID 15857398.
- ^ Bonda DJ, Wang X, Lee HG, Smith MA, Perry G, Zhu X (April 2014). "Neuronal failure in Alzheimer's disease: a view through the oxidative stress looking-glass". Bulletin neurovědy. 30 (2): 243–52. doi:10.1007/s12264-013-1424-x. PMC 4097013. PMID 24733654.
- ^ Liu Z, Zhou T, Ziegler AC, Dimitrion P, Zuo L (2017). "Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications". Oxidační medicína a buněčná dlouhověkost. 2017: 2525967. doi:10.1155/2017/2525967. PMC 5529664. PMID 28785371.
- ^ Ayala-Peña S (September 2013). "Role of oxidative DNA damage in mitochondrial dysfunction and Huntington's disease pathogenesis". Radikální biologie a medicína zdarma. 62: 102–110. doi:10.1016/j.freeradbiomed.2013.04.017. PMC 3722255. PMID 23602907.
- ^ Kikuchi H, Furuta A, Nishioka K, Suzuki SO, Nakabeppu Y, Iwaki T (April 2002). "Impairment of mitochondrial DNA repair enzymes against accumulation of 8-oxo-guanine in the spinal motor neurons of amyotrophic lateral sclerosis". Acta Neuropathologica. 103 (4): 408–14. doi:10.1007/s00401-001-0480-x. PMID 11904761. S2CID 2102463.
- ^ Lehmann G, Budovsky A, Muradian KK, Fraifeld VE (2006). "Mitochondrial genome anatomy and species-specific lifespan". Omlazovací výzkum. 9 (2): 223–6. doi:10.1089/rej.2006.9.223. PMID 16706648.
- ^ A b Lehmann G, Segal E, Muradian KK, Fraifeld VE (April 2008). "Do mitochondrial DNA and metabolic rate complement each other in determination of the mammalian maximum longevity?". Omlazovací výzkum. 11 (2): 409–17. doi:10.1089/rej.2008.0676. PMID 18442324.
- ^ Lehmann G, Muradian KK, Fraifeld VE (2013). "Telomere length and body temperature-independent determinants of mammalian longevity?". Frontiers in Genetics. 4 (111): 111. doi:10.3389/fgene.2013.00111. PMC 3680702. PMID 23781235.
- ^ Toren D, Barzilay T, Tacutu R, Lehmann G, Muradian KK, Fraifeld VE (January 2016). "MitoAge: a database for comparative analysis of mitochondrial DNA, with a special focus on animal longevity". Výzkum nukleových kyselin. 44 (D1): D1262-5. doi:10.1093/nar/gkv1187. PMC 4702847. PMID 26590258.
- ^ Damas J, Carneiro J, Gonçalves J, Stewart JB, Samuels DC, Amorim A, Pereira F (September 2012). "Mitochondrial DNA deletions are associated with non-B DNA conformations". Výzkum nukleových kyselin. 40 (16): 7606–21. doi:10.1093/nar/gks500. PMC 3439893. PMID 22661583.
- ^ Oliveira PH, da Silva CL, Cabral JM (2013). "An appraisal of human mitochondrial DNA instability: new insights into the role of non-canonical DNA structures and sequence motifs". PLOS ONE. 8 (3): e59907. Bibcode:2013PLoSO...859907O. doi:10.1371/journal.pone.0059907. PMC 3612095. PMID 23555828.
- ^ Brown WM, George M, Wilson AC (April 1979). "Rapid evolution of animal mitochondrial DNA". Sborník Národní akademie věd Spojených států amerických. 76 (4): 1967–71. Bibcode:1979PNAS...76.1967B. doi:10.1073/pnas.76.4.1967. JSTOR 69636. PMC 383514. PMID 109836.
- ^ Davis, C. Leland (1998). "Mitochondrial DNA: State of Tennessee v. Paul Ware" (PDF). Profiles in DNA. 1 (3): 6–7.
- ^ [1] Court case name listed in the appeal.[úplná citace nutná ] Vyvolány 17 April 2015.
- ^ [2][self-publikoval zdroj? ] Defense lawyer. Vyvolány 17 April 2015.
- ^ Garlicki, Debbie (11 March 1998). "DNA Tests Got Rorrer Life in Jail". Ranní volání.
- ^ Pellegrino, Peter; Thomas, Peter; Medstar Television; Courtroom Television Network (2000), Forensic files. a woman scorned Episode 55 Episode 55, [publisher not identified], OCLC 456260156
- ^ "Judge allows DNA in Samantha Runnion case," Associated Press, 18 February 2005. Retrieved 4 April 2007.
- ^ "Canine DNA Admitted In California Murder Case," Archivováno 2. února 2014 v Wayback Machine Pit Bulletin Legal News, 5 December 2013. Retrieved 21 January 2014.
- ^ Kennedy, Maev (4. února 2013). "Richard III: DNA confirms twisted bones belong to king". Opatrovník. Citováno 7. prosince 2014.
- ^ Xu, X .; Arnason, U. (1 May 1996). "A complete sequence of the mitochondrial genome of the western lowland gorilla". Molekulární biologie a evoluce. 13 (5): 691–698. doi:10.1093/oxfordjournals.molbev.a025630. ISSN 0737-4038. PMID 8676744.
- ^ Nass MM, Nass S (December 1963). "INTRAMITOCHONDRIAL FIBERS WITH DNA CHARACTERISTICS: I. Fixation and Electron Staining Reactions". The Journal of Cell Biology. 19 (3): 593–611. doi:10.1083/jcb.19.3.593. PMC 2106331. PMID 14086138.
- ^ Schatz G, Haslbrunner E, Tuppy H (March 1964). "Deoxyribonucleic acid associated with yeast mitochondria". Sdělení o biochemickém a biofyzikálním výzkumu. 15 (2): 127–32. doi:10.1016/0006-291X(64)90311-0. PMID 26410904.
- ^ Ehler E, Novotný J, Juras A, Chylenski M, Moravcík O, Paces J (January 2019). "AmtDB: a database of ancient human mitochondrial genomes". Výzkum nukleových kyselin. 47 (D1): D29–D32. doi:10.1093/nar/gky843. PMC 6324066. PMID 30247677.
- ^ Gu Z, Li J, Gao S, Gong M, Wang J, Xu H, Zhang C, Wang J (June 2011). "InterMitoBase: an annotated database and analysis platform of protein-protein interactions for human mitochondria". BMC Genomics. 12: 335. doi:10.1186/1471-2164-12-335. PMC 3142533. PMID 21718467.
- ^ Damas J, Carneiro J, Amorim A, Pereira F (January 2014). "MitoBreak: the mitochondrial DNA breakpoints database". Výzkum nukleových kyselin. 42 (Database issue): D1261-8. doi:10.1093/nar/gkt982. PMC 3965124. PMID 24170808.
- ^ Iwasaki W, Fukunaga T, Isagozawa R, Yamada K, Maeda Y, Satoh TP, Sado T, Mabuchi K, Takeshima H, Miya M, Nishida M (November 2013). "MitoFish and MitoAnnotator: a mitochondrial genome database of fish with an accurate and automatic annotation pipeline". Molekulární biologie a evoluce. 30 (11): 2531–40. doi:10.1093/molbev/mst141. PMC 3808866. PMID 23955518.
- ^ Cawthorn DM, Steinman HA, Witthuhn RC (November 2011). "Establishment of a mitochondrial DNA sequence database for the identification of fish species commercially available in South Africa". Zdroje molekulární ekologie. 11 (6): 979–91. doi:10.1111/j.1755-0998.2011.03039.x. PMID 21689383.
- ^ Lee YS, Oh J, Kim YU, Kim N, Yang S, Hwang UW (January 2008). "Mitome: dynamic and interactive database for comparative mitochondrial genomics in metazoan animals". Výzkum nukleových kyselin. 36 (Database issue): D938-42. doi:10.1093/nar/gkm763. PMC 2238945. PMID 17940090.
- ^ Catalano D, Licciulli F, Turi A, Grillo G, Saccone C, D'Elia D (January 2006). "MitoRes: a resource of nuclear-encoded mitochondrial genes and their products in Metazoa". BMC bioinformatika. 7: 36. doi:10.1186/1471-2105-7-36. PMC 1395343. PMID 16433928.
- ^ Kumar M, Kapil A, Shanker A (November 2014). "MitoSatPlant: mitochondrial microsatellites database of viridiplantae". Mitochondrie. 19 Pt B: 334–7. doi:10.1016/j.mito.2014.02.002. PMID 24561221.
- ^ D'Onorio de Meo P, D'Antonio M, Griggio F, Lupi R, Borsani M, Pavesi G, Castrignanò T, Pesole G, Gissi C (January 2012). "MitoZoa 2.0: a database resource and search tools for comparative and evolutionary analyses of mitochondrial genomes in Metazoa". Výzkum nukleových kyselin. 40 (Database issue): D1168-72. doi:10.1093/nar/gkr1144. PMC 3245153. PMID 22123747.
externí odkazy
 Média související s Mitochondriální DNA na Wikimedia Commons
Média související s Mitochondriální DNA na Wikimedia Commons
Typy nukleové kyseliny | |||||||
|---|---|---|---|---|---|---|---|
| Složky | |||||||
| Ribonukleové kyseliny (kódování, nekódování ) |
| ||||||
| Deoxyribonukleové kyseliny | |||||||
| Analogy | |||||||
| Klonovací vektory | |||||||
| |||||||