Cytochrom c - Cytochrome c





The komplex cytochromůnebo cyt C, je malý hemeprotein nalezen volně spojený s vnitřní membrána z mitochondrie. Patří k rodina cytochromu c proteinů a hraje hlavní roli v buněčné apoptóze. Cytochrom c je vysoce rozpustné ve vodě, na rozdíl od jiných cytochromy, a je podstatnou součástí elektronový transportní řetězec, kde nese jeden elektron. Je schopen podstoupit oxidace a snížení jako jeho žehlička atom konvertuje mezi železný a železitý formy, ale neváže se kyslík. Přenáší elektrony mezi Komplexy III (Koenzym Q - Cyt C reduktáza) a IV (Cyt C oxidáza). U lidí je cytochrom c kódován CYCS gen.[5][6]
Distribuce druhů
Cytochrom c je vysoce konzervovaný protein napříč spektrem druhů, který se nachází v rostlinách, zvířatech a mnoha jednobuněčných organismech. To spolu s jeho malou velikostí (molekulová hmotnost asi 12 000) daltony ),[7] je užitečné při studiích kladistika.[8] Molekula cytochromu c byla studována pro záblesk, který poskytuje do evoluční biologie.
Cytochrom c má primární strukturu skládající se z řetězce přibližně 100 aminokyseliny. Mnoho organismů vyššího řádu vlastní řetězec 104 aminokyselin.[9] Sekvence cytochromu c u lidí jsou identické se sekvencemi šimpanzů (našich nejbližších příbuzných), ale liší se od sekvencí koní.[10]
Cytochrom c má aminokyselinovou sekvenci, která je u eukaryot vysoce konzervovaná a liší se pouze několika zbytky. U více než třiceti druhů testovaných v jedné studii bylo konzervováno 34 ze 104 aminokyselin; identické v jejich charakteristické poloze.[11] Například člověk cytochrom oxidáza reaguje s pšeničným cytochromem C, in vitro; což platilo pro všechny páry testovaných druhů.[11] Redoxní potenciál +0,25 voltů je navíc ve všech cytochromech stejný C studované molekuly.[11]
Struktura

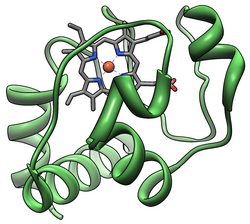
Cytochrom c patří do třídy I Rodina cytochromů typu C.[13] a obsahuje charakteristický motiv aminokyseliny CXXCH (cystein-jakýkoli-jakýkoli-cystein-histidin), který váže hem.[14].Tento motiv je umístěn směrem k N-konec z peptid řetězce a obsahuje histidin jako pátý ligand hemového železa. Šestý ligand je poskytován a methionin zbytky nalezené směrem k C-konec. Páteř bílkovin je složen do pěti α-šroubovice které jsou očíslovány α1-α5 od N-konce k C-konci. Helixy α3, α4 a α5 se označují jako šroubovice 50., 60. a 70. let, když se odkazuje na mitochondriální cytochrom c.[15]
Heme c

Zatímco většina hemových proteinů je připojena k protetické skupině ligací iontů železa a terciárními interakcemi, hemová skupina cytochromu c vytváří thioetherové vazby se dvěma cystein postranní řetězce proteinu.[16] Jednou z hlavních vlastností hemu c, která umožňuje cytochromu c mít různé funkce, je jeho schopnost mít různé redukční potenciály v přírodě. Tato vlastnost určuje kinetiku a termodynamiku reakce přenosu elektronů.[17]
Dipólový moment
Dipólový moment hraje důležitou roli při orientaci proteinů správným směrem a při zvyšování jejich schopnosti vázat se na jiné molekuly.[18][19] Dipólový moment cytochromu c je výsledkem shluku negativně nabitých postranních řetězců aminokyselin v „zadní části“ enzymu.[19] Přes rozdíly v počtu vázaných skupin hemu a změny v sekvenci je dipólový moment obratlovců cytochromů c pozoruhodně zachován. Například cytochromy obratlovců c mají dipólový moment přibližně 320 debye zatímco cytochromy c rostlin a hmyzu mají dipólový moment přibližně 340 debye.[19]
Funkce
Cytochrom c je součástí elektronový transportní řetězec v mitochondriích. The heme skupina cytochromu c přijímá elektrony z před naším letopočtem1 komplex a přenáší elektrony do komplex IV. Cytochrom c se také podílí na iniciaci apoptóza. Po uvolnění cytochromu c do cytoplazmy se protein váže faktor aktivující apoptotickou proteázu-1 (Apaf-1).[5]
Cytochrom c může také katalyzovat několik redoxních reakcí, jako je hydroxylace a aromatický oxidace a ukazuje peroxidáza aktivita oxidací různých donorů elektronů, jako je 2,2-azino-bis(3-ethylbenzthiazolin-6-sulfonová kyselina) (ABTS ), 2-keto-4-thiomethylmaslová kyselina a 4-aminoantipyrin.
Bakteriální cytochrom C funguje jako a nitrit reduktáza.[20]
Role v apoptóze
Cytochrom c objevil v roce 1996 také Dr. Xiaodong Wang mít mezilehlou roli v apoptóza, kontrolovaná forma buněčné smrti používaná k zabíjení buněk v procesu vývoje nebo v reakci na infekci nebo poškození DNA.[21]
Cytochrom c se váže na kardiolipin ve vnitřní mitochondriální membráně, čímž zakotví její přítomnost a zabrání jejímu uvolnění z mitochondrií a zahájení apoptózy. Zatímco počáteční přitažlivost mezi kardiolipinem a cytochromem c je elektrostatická kvůli extrémnímu pozitivnímu náboji na cytochromu c, konečná interakce je hydrofobní, kdy se hydrofobní ocas z kardiolipinu vloží do hydrofobní části cytochromu c.
Během rané fáze apoptózy je stimulována mitochondriální produkce ROS a kardiolipin je oxidován funkcí peroxidázy komplexu kardiolipin-cytochrom c. Hemoprotein je poté oddělen od mitochondriální vnitřní membrány a může být extrudován do rozpustné cytoplazmy póry ve vnější membráně.[22]
Trvalá výška v vápník hladiny předchází cyt C uvolnění z mitochondrií. Uvolňování malého množství cyt C vede k interakci s Receptor IP3 (IP3R) na endoplazmatické retikulum (ER), což způsobuje uvolňování ER vápníku. Celkové zvýšení vápníku spouští masivní uvolňování cyt C, který pak působí ve smyčce pozitivní zpětné vazby k udržení uvolňování ER vápníku prostřednictvím IP3R.[23] To vysvětluje, jak může uvolňování vápníku ER dosáhnout cytotoxických hladin. Toto uvolňování cytochromu c se zase aktivuje kaspáza 9 cystein proteáza. Caspase 9 pak může pokračovat v aktivaci kaspáza 3 a kaspáza 7, kteří jsou zodpovědní za zničení buňky zevnitř.
Inhibice apoptózy
Jedním ze způsobů aktivace buněčné apoptózy je uvolňování cytochromu c z mitochondrií do cytosolu. Studie ukázala, že buňky jsou schopné chránit se před apoptózou blokováním uvolňování cytochromu c pomocí Bcl-xL.[24] Dalším způsobem, jak mohou buňky kontrolovat apoptózu, je fosforylace Tyr48, která by z cytochromu c udělala antiapoptotický přepínač.[25]
Jako antioxidační enzym

Cytochrom c je známo, že hraje roli v elektronový transportní řetězec a buňka apoptóza. Nedávná studie však ukázala, že může také působit jako antioxidační enzym v mitochondriích; a dělá to odstraněním superoxid (Ó2–) a peroxid vodíku (H2Ó2) z mitochondrie.[26] Proto není v mitochondriích pro buněčné dýchání vyžadován pouze cytochrom c, ale v mitochondriích je také nutný k omezení produkce O2– a H2Ó2.[26]
Extramitochondriální lokalizace
Obecně se má za to, že cytochrom c je za normálních fyziologických podmínek lokalizován pouze v mitochondriálním mezimembránovém prostoru.[27] Uvolňování cytochromu-c z mitochondrií do cytosolu, kde aktivuje kaspáza rodina proteázy Předpokládá se, že je primárním spouštěčem vedoucím k nástupu apoptózy.[28] Měření množství unikajícího cytochromu c z mitochondrií do cytosolu a ven z buňky do kultivačního média je citlivou metodou ke sledování stupně apoptózy.[29][30] Podrobné imunoelektronové mikroskopické studie s řezy tkání potkanů využívající protilátky specifické pro cytochrom c však poskytují přesvědčivý důkaz, že cytochrom-c za normálních buněčných podmínek je také přítomen na extramitochondriálních místech.[31] V pankreatických acinárních buňkách a přední hypofýza, byla v. detekována silná a specifická přítomnost cytochromu-c zymogen granule a v růstový hormon granule. V pankreatu byl cytochrom-c také nalezen při kondenzaci vakuoly a na acináru lumen. Ukázalo se, že extramitochondriální lokalizace cytochromu c je specifická, protože byla zcela zrušena adsorpcí primární protilátky s čištěným cytochromem c.[31] Přítomnost cytochromu-c mimo mitochondrie na konkrétním místě za normálních fyziologických podmínek vyvolává důležité otázky týkající se jeho buněčné funkce a translokačního mechanismu.[31] Kromě cytochromu c byla pozorována extramitochondriální lokalizace také u velkého počtu dalších proteinů, včetně těch kódovaných mitochondriální DNA.[32][33][34] To zvyšuje možnost existence dosud neidentifikovaných specifických mechanismů pro translokaci proteinů z mitochondrií do jiných buněčných destinací.[34][35]
Aplikace
Detekce superoxidu

Cytochrom c byl použit k detekci produkce peroxidů v biologických systémech. Při produkci superoxidu se počet oxidovaného cytochromu c3+ zvyšuje a snižuje cytochrom c2+ klesá.[36] Superoxid se však často vyrábí s oxidem dusnatým. V přítomnosti oxidu dusnatého je redukce cytochromu c3+ je potlačen.[37] To vede k oxidaci cytochromu c2+ na cytochrom c3+ podle kyselina peroxynitrous, meziprodukt vyrobený reakcí oxidu dusnatého a superoxidu.[37] Přítomnost peroxynitrit nebo H2Ó2 a oxid dusičitý NE2 v mitochondriích mohou být smrtelné, protože nitrují tyrosin zbytky cytochromu c, což vede k narušení funkce cytochromu c jako nosiče elektronů v řetězci přenosu elektronů.[38]
Viz také
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000172115 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000063694 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b "Entrez Gene: cytochrom c".
- ^ Tafani M, Karpinich NO, Hurster KA, Pastorino JG, Schneider T, Russo MA, Farber JL (březen 2002). „Uvolňování cytochromu c po aktivaci receptoru Fas závisí na translokaci nabídky v plné délce a indukci přechodu mitochondriální permeability“. The Journal of Biological Chemistry. 277 (12): 10073–82. doi:10,1074 / jbc.M111350200. PMID 11790791.
- ^ „Cytochrom c - Homo sapiens (člověk)“. P99999. Konsorcium UniProt.
hmotnost je 11 749 Daltonů
- ^ Margoliash E (říjen 1963). "Primární struktura a vývoj cytochromu c". Sborník Národní akademie věd Spojených států amerických. 50 (4): 672–9. doi:10.1073 / pnas.50.4.672. PMC 221244. PMID 14077496.
- ^ Aminokyselinové sekvence v proteinech cytochromu c různých druhů, převzato z Strahler, Arthur; Věda a historie Země, 1997. strana 348.
- ^ Lurquin PF, Stone L, Cavalli-Sforza LL (2007). Geny, kultura a vývoj člověka: syntéza. Oxford: Blackwell. p. 79. ISBN 978-1-4051-5089-7.
- ^ A b C Stryer L (1975). Biochemie (1. vyd.). San Francisco: W.H. Freeman a společnost. p.362. ISBN 978-0-7167-0174-3.
- ^ McPherson A, DeLucas LJ (2015). "Krystalizace proteinu mikrogravitace". Mikrogravitace NPJ. 1: 15010. doi:10.1038 / npjmgrav.2015.10. PMC 5515504. PMID 28725714.
- ^ Ambler RP (květen 1991). "Variabilita sekvencí v bakteriálních cytochromech c". Biochimica et Biophysica Acta (BBA) - bioenergetika. 1058 (1): 42–7. doi:10.1016 / S0005-2728 (05) 80266-X. PMID 1646017.
- ^ Mavridou DA, Ferguson SJ, Stevens JM (březen 2013). "Sestava cytochromu c". IUBMB Life. 65 (3): 209–16. doi:10.1002 / iub.1123. PMID 23341334. S2CID 32216217.
- ^ Liu J, Chakraborty S, Hosseinzadeh P, Yu Y, Tian S, Petrik I, Bhagi A, Lu Y (2014-04-23). „Metaloproteiny obsahující centra cytochromu, železa a síry nebo mědi redox“. Chemické recenze. 114 (8): 4366–4469. doi:10.1021 / cr400479b. ISSN 0009-2665. PMC 4002152. PMID 24758379.
- ^ Kang X, Carey J (listopad 1999). "Role hemu ve strukturní organizaci cytochromu c sondovaná semisyntézou". Biochemie. 38 (48): 15944–51. doi:10.1021 / bi9919089. PMID 10625461.
- ^ Zhao Y, Wang ZB, Xu JX (leden 2003). „Vliv cytochromu c na tvorbu a eliminaci O2– a H2Ó2 v mitochondriích ". The Journal of Biological Chemistry. 278 (4): 2356–60. doi:10,1074 / jbc.M209681200. PMID 12435729.
- ^ Koppenol WH, Margoliash E (duben 1982). "Asymetrická distribuce nábojů na povrchu koňského cytochromu c. Funkční důsledky". The Journal of Biological Chemistry. 257 (8): 4426–37. PMID 6279635.
- ^ A b C Koppenol WH, Rush JD, Mills JD, Margoliash E (červenec 1991). „Dipólový moment cytochromu c“. Molekulární biologie a evoluce. 8 (4): 545–58. doi:10.1093 / oxfordjournals.molbev.a040659. PMID 1656165.
- ^ Schneider J, Kroneck PM (2014). „Kapitola 9: Produkce amoniaku multihemovými cytochromy c“. V Kroneck PM, Torres ME (eds.). Kovem řízená biogeochemie plynných sloučenin v životním prostředí. Kovové ionty v biologických vědách. 14. Springer. 211–236. doi:10.1007/978-94-017-9269-1_9. ISBN 978-94-017-9268-4. PMID 25416396.
- ^ Liu X, Kim CN, Yang J, Jemmerson R, Wang X (červenec 1996). "Indukce apoptotického programu v bezbuněčných extraktech: požadavek na dATP a cytochrom c". Buňka. 86 (1): 147–57. doi:10.1016 / S0092-8674 (00) 80085-9. PMID 8689682. S2CID 12604356.
- ^ Orrenius S, Životovský B (září 2005). "Oxidace kardiolipinu osvobozuje cytochrom c". Přírodní chemická biologie. 1 (4): 188–9. doi:10.1038 / nchembio0905-188. PMID 16408030. S2CID 45381495.
- ^ Boehning D, Patterson RL, Sedaghat L, Glebova NO, Kurosaki T, Snyder SH (prosinec 2003). „Cytochrom c se váže na inositol (1,4,5) trisfosfátové receptory a zesiluje na vápníku závislou apoptózu.“ Přírodní buněčná biologie. 5 (12): 1051–61. doi:10.1038 / ncb1063. PMID 14608362. S2CID 27761335.
- ^ Kharbanda S, Pandey P, Schofield L, Israels S, Roncinske R, Yoshida K, Bharti A, Yuan ZM, Saxena S, Weichselbaum R, Nalin C, Kufe D (červen 1997). „Role pro Bcl-xL jako inhibitor akumulace cytosolického cytochromu C v apoptóze způsobené poškozením DNA“. Sborník Národní akademie věd Spojených států amerických. 94 (13): 6939–42. doi:10.1073 / pnas.94.13.6939. PMC 21263. PMID 9192670.
- ^ García-Heredia JM, Díaz-Quintana A, Salzano M, Orzáez M, Pérez-Payá E, Teixeira M, De la Rosa MA, Díaz-Moreno I (prosinec 2011). „Fosforylace tyrosinu přeměňuje alkalický přechod na biologicky relevantní proces a způsobuje, že se lidský cytochrom c chová jako antiapoptotický přepínač.“ Journal of Biological Anorganic Chemistry. 16 (8): 1155–68. doi:10.1007 / s00775-011-0804-9. PMID 21706253. S2CID 24156094.
- ^ A b Bowman SE, Bren KL (prosinec 2008). „The chemistry and biochemistry of heme c: functional bases for covalent attachment“. Zprávy o přírodních produktech. 25 (6): 1118–30. doi:10.1039 / b717196j. PMC 2654777. PMID 19030605.
- ^ Neupert W (1997). "Import bílkovin do mitochondrií". Roční přehled biochemie. 66: 863–917. doi:10,1146 / annurev.biochem. 66.1.863. PMID 9242927.
- ^ Kroemer G, Dallaporta B, Resche-Rigon M (1998). "Regulátor mitochondriální smrti / života u apoptózy a nekrózy". Roční přehled fyziologie. 60: 619–42. doi:10.1146 / annurev.physiol.60.1.619. PMID 9558479.
- ^ Loo JF, Lau PM, Ho HP, Kong SK (říjen 2013). „Test bio-čárového kódu na bázi aptameru s amplifikací izotermické rekombinázy polymerázy pro detekci cytochromu-c a screening protinádorových léků“. Talanta. 115: 159–65. doi:10.1016 / j.talanta.2013.04.051. PMID 24054573.
- ^ Waterhouse NJ, Trapani JA (červenec 2003). „Nový kvantitativní test na uvolňování cytochromu c v apoptotických buňkách“. Buněčná smrt a diferenciace. 10 (7): 853–5. doi:10.1038 / sj.cdd.4401263. PMID 12815469.
- ^ A b C Soltys BJ, Andrews DW, Jemmerson R, Gupta RS (2001). „Cytochrom-C se lokalizuje v sekrečních granulích v pankreatu a přední hypofýze“. Cell Biology International. 25 (4): 331–8. doi:10.1006 / cbir.2000.0651. PMID 11319839. S2CID 2106599.
- ^ Gupta RS, Ramachandra NB, Bowes T, Singh B (2008). "Neobvyklá buněčná dispozice mitochondriálních molekulárních chaperonů Hsp60, Hsp70 a Hsp10". Biologie extracelulárních molekulárních chaperonů. Sympózium Nadace Novartis. Symposia Novartis Foundation. 291. s. 59–68, diskuse 69–73, 137–40. doi:10.1002 / 9780470754030.ch5. ISBN 978-0-470-75403-0. PMID 18575266.
- ^ Sadacharan SK, Singh B, Bowes T, Gupta RS (listopad 2005). „Lokalizace mitochondriální DNA kódovaná podjednotkami I a II cytochrom c oxidázy v granulích potkaního pankreatického zymogenu a granulích růstového hormonu hypofýzy“. Histochemie a buněčná biologie. 124 (5): 409–21. doi:10.1007 / s00418-005-0056-2. PMID 16133117. S2CID 24440427.
- ^ A b Soltys BJ, Gupta RS (2000). Mitochondriální proteiny na neočekávaných buněčných místech: export proteinů z mitochondrií z evoluční perspektivy. International Review of Cytology. 194. 133–96. doi:10.1016 / s0074-7696 (08) 62396-7. ISBN 978-0-12-364598-2. PMID 10494626.
- ^ Soltys BJ, Gupta RS (květen 1999). "Proteiny mitochondriální matice na neočekávaných místech: jsou exportovány?". Trendy v biochemických vědách. 24 (5): 174–7. doi:10.1016 / s0968-0004 (99) 01390-0. PMID 10322429.
- ^ McCord JM, Fridovich I (listopad 1969). "Superoxid dismutáza. Enzymatická funkce pro erytrocuprein (hemocuprein)". The Journal of Biological Chemistry. 244 (22): 6049–55. PMID 5389100.
- ^ A b Thomson L, Trujillo M, Telleri R, Radi R (červen 1995). „Kinetika cytochromu c2+ oxidace peroxynitritem: důsledky pro měření superoxidu v biologických systémech produkujících oxid dusnatý “. Archivy biochemie a biofyziky. 319 (2): 491–7. doi:10.1006 / abbi.1995.1321. PMID 7786032.
- ^ Domazou AS, Gebicka L, Didik J, Gebicki JL, van der Meijden B, Koppenol WH (duben 2014). „Kinetika reakce oxidu dusičitého s železem (II) - a železem (III) cytochromem c“. Radikální biologie a medicína zdarma. 69: 172–80. doi:10.1016 / j.freeradbiomed.2014.01.014. PMID 24447894.
Další čtení
- Kumarswamy R, Chandna S (únor 2009). „Předpokládaní partneři v Bax zprostředkovaném uvolňování cytochromu-c: ANT, CypD, VDAC nebo žádný z nich?“. Mitochondrie. 9 (1): 1–8. doi:10.1016 / j.mito.2008.10.003. PMID 18992370.
- VP Skulachev (únor 1998). "Cytochrom c v apoptotických a antioxidačních kaskádách". FEBS Dopisy. 423 (3): 275–80. doi:10.1016 / S0014-5793 (98) 00061-1. PMID 9515723. S2CID 10267410.
- Mannella CA (1998). „Konformační změny v proteinu mitochondriálního kanálu, VDAC a jejich funkční implikace“. Journal of Structural Biology. 121 (2): 207–18. doi:10.1006 / jsbi.1997.3954. PMID 9615439.
- Ferri KF, Jacotot E, Blanco J, Esté JA, Kroemer G (2000). "Mitochondriální kontrola buněčné smrti vyvolané proteiny kódovanými HIV-1". Annals of the New York Academy of Sciences. 926: 149–64. doi:10.1111 / j.1749-6632.2000.tb05609.x. PMID 11193032. S2CID 21997163.
- Britton RS, Leicester KL, Bacon BR (říjen 2002). "Železná toxicita a chelatační terapie". International Journal of Hematology. 76 (3): 219–28. doi:10.1007 / BF02982791. PMID 12416732. S2CID 22572183.
- Haider N, Narula N, Narula J (prosinec 2002). „Apoptóza při srdečním selhání představuje programované přežití buněk, nikoli smrt, kardiomyocytů a pravděpodobnost reverzní remodelace“. Deník srdečního selhání. 8 (6 doplňků): S512–7. doi:10.1054 / jcaf.2002.130034. PMID 12555167.
- Castedo M, Perfettini JL, Andreau K, Roumier T, Piacentini M, Kroemer G (prosinec 2003). "Mitochondriální apoptóza vyvolaná obálkou HIV-1". Annals of the New York Academy of Sciences. 1010: 19–28. doi:10.1196 / anály.1299.004. PMID 15033690. S2CID 37073602.
- Ng S, Smith MB, Smith HT, Millett F (listopad 1977). "Vliv modifikace jednotlivých cytochromu c lysinů na reakci s cytochromem b5". Biochemie. 16 (23): 4975–8. doi:10.1021 / bi00642a006. PMID 199233.
- Lynch SR, Sherman D, Copeland RA (leden 1992). „Vazba cytochromu c ovlivňuje konformaci cytochromu a v cytochrom c oxidáze“. The Journal of Biological Chemistry. 267 (1): 298–302. PMID 1309738.
- Garber EA, Margoliash E (únor 1990). „Interakce cytochromu c s cytochromem c oxidázou: porozumění přechodu z vysoké na nízkou afinitu“. Biochimica et Biophysica Acta (BBA) - bioenergetika. 1015 (2): 279–87. doi:10.1016 / 0005-2728 (90) 90032-Y. PMID 2153405.
- Bedetti CD (květen 1985). „Imunocytochemická demonstrace cytochrom c oxidázy metodou imunoperoxidázy: specifické barvivo pro mitochondrie v lidských tkáních fixovaných ve formalínu a zalitých v parafinu“. The Journal of Histochemistry and Cytochemistry. 33 (5): 446–52. doi:10.1177/33.5.2580882. PMID 2580882.
- Tanaka Y, Ashikari T, Shibano Y, Amachi T, Yoshizumi H, Matsubara H (červen 1988). "Konstrukce lidského genu pro cytochrom c a jeho funkční exprese v Saccharomyces cerevisiae". Journal of Biochemistry. 103 (6): 954–61. doi:10.1093 / oxfordjournals.jbchem.a122393. PMID 2844747.
- Evans MJ, Scarpulla RC (prosinec 1988). „Gen lidského somatického cytochromu c: dvě třídy zpracovaných pseudogenů ohraničují období rychlého molekulárního vývoje“. Sborník Národní akademie věd Spojených států amerických. 85 (24): 9625–9. doi:10.1073 / pnas.85.24.9625. PMC 282819. PMID 2849112.
- Passon PG, Hultquist DE (červenec 1972). "Rozpustná cytochrom b 5 reduktáza z lidských erytrocytů". Biochimica et Biophysica Acta (BBA) - bioenergetika. 275 (1): 62–73. doi:10.1016/0005-2728(72)90024-2. hdl:2027.42/34070. PMID 4403130.
- Dowe RJ, Vitello LB, Erman JE (srpen 1984). "Sedimentační rovnovážné studie interakce mezi cytochromem c a cytochromem c peroxidázou". Archivy biochemie a biofyziky. 232 (2): 566–73. doi:10.1016/0003-9861(84)90574-5. PMID 6087732.
- Michel B, Bosshard HR (srpen 1984). "Spektroskopická analýza interakce mezi cytochromem c a cytochromem c oxidázou". The Journal of Biological Chemistry. 259 (16): 10085–91. PMID 6088481.
- Broger C, Nałecz MJ, Azzi A (říjen 1980). "Interakce cytochromu c s komplexem mitochondriálního dýchacího řetězce cytochromu bc1". Biochimica et Biophysica Acta (BBA) - bioenergetika. 592 (3): 519–27. doi:10.1016/0005-2728(80)90096-1. PMID 6251869.
- Smith HT, Ahmed AJ, Millett F (květen 1981). „Elektrostatická interakce cytochromu c s cytochromem c1 a cytochromoxidázou“. The Journal of Biological Chemistry. 256 (10): 4984–90. PMID 6262312.
- Geren LM, Millett F (říjen 1981). "Studie přenosu fluorescenční energie interakce mezi adrenodoxinem a cytochromem c". The Journal of Biological Chemistry. 256 (20): 10485–9. PMID 6270113.
- Favre B, Zolnierowicz S, Turowski P, Hemmings BA (červen 1994). „Katalytická podjednotka proteinové fosfatázy 2A je karboxyl-methylovaná in vivo.“ The Journal of Biological Chemistry. 269 (23): 16311–7. PMID 8206937.
- Gao B, Eisenberg E, Greene L (červenec 1996). „Vliv konstitutivní polymerace proteinu tepelného šoku 70 kDa na jeho interakci s proteinovým substrátem“. The Journal of Biological Chemistry. 271 (28): 16792–7. doi:10.1074 / jbc.271.28.16792. PMID 8663341.
externí odkazy
- Protein cytochromu c
- Apoptóza a kaspáza 3 - PMAP Mapa proteolýzy -animace
- Cytochrom + c v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- Přehled všech strukturálních informací dostupných v PDB pro UniProt: P99999 (Cytochrom c) na PDBe-KB.
Galerie PDB | |
|---|---|
|

