Sukcinát dehydrogenáza - Succinate dehydrogenase
| sukcinátdehydrogenáza (sukcinát-ubichinon oxidoreduktáza) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
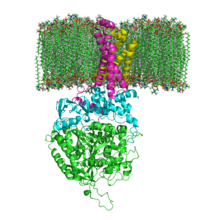 Struktura SQR ve fosfolipidové membráně. SdhA, SdhB, SdhC a SdhD | |||||||||
| Identifikátory | |||||||||
| EC číslo | 1.3.5.1 | ||||||||
| Číslo CAS | 9028-11-9 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
| Sukcinát dehydrogenáza | |
|---|---|
| Identifikátory | |
| Symbol | Respirační komplex II |
| OPM nadčeleď | 3 |
| OPM protein | 1zoy |
| Membranome | 656 |
Sukcinát dehydrogenáza (SDH) nebo sukcinát-koenzym Q reduktáza (SQR) nebo respirační Komplex II je enzym komplex, nalezený v mnoha bakteriální buňky a v vnitřní mitochondriální membrána z eukaryoty. Je to jediný enzym, který se účastní obou cyklus kyseliny citronové a elektronový transportní řetězec.[1] Histochemická analýza ukazující vysokou sukcinátdehydrogenázu ve svalu prokazuje vysoký mitochondriální obsah a vysoký oxidační potenciál.[2]
V kroku 6 cyklus kyseliny citronové, SQR katalyzuje the oxidace z sukcinát na fumarát s snížení z ubichinon na ubichinol. K tomu dochází ve vnitřním mitochondriu membrána podle spojka obě reakce dohromady.
Struktura

Podjednotky
Mitochondriální a mnoho bakteriální SQR se skládají ze čtyř strukturálně odlišných podjednotky: dva hydrofilní a dva hydrofobní. První dvě podjednotky, a flavoprotein (SdhA) a an železo-sirný protein (SdhB), tvoří hydrofilní hlavu, kde probíhá enzymatická aktivita komplexu. SdhA obsahuje a kovalentně připojený flavin adenin dinukleotid (FAD) kofaktor a sukcinát vazebné místo a SdhB obsahuje tři klastry železo-síra: [2Fe-2S], [4Fe-4S] a [3Fe-4S]. Druhé dvě podjednotky jsou hydrofobní membránové kotvící podjednotky, SdhC a SdhD. Lidské mitochondrie obsahují dvě odlišné izoformy SdhA (Fp podjednotky typu I a typu II), tyto izoformy se také nacházejí v Ascaris suum a Caenorhabditis elegans.[3] Podjednotky tvoří membránově vázané cytochrom b komplex se šesti transmembránový šroubovice obsahující jeden heme b skupina a ubichinon -závazné místo. Dva fosfolipid molekuly, jedna kardiolipin a jeden fosfatidylethanolamin, se také nacházejí v podjednotkách SdhC a SdhD (na obrázku nejsou zobrazeny). Slouží k obsazení hydrofobního prostoru pod lemem b. Tyto podjednotky jsou zobrazeny na přiloženém obrázku. SdhA je zelená, SdhB je zelenomodrá, SdhC je fuchsie a SdhD je žlutá. Kolem SdhC a SdhD je a fosfolipidová membrána s mezimembránovým prostorem v horní části obrázku.[4]
Tabulka složení podjednotky[5]
| Ne. | Název podjednotky | Člověk protein | Popis bílkovin z UniProt | Pfam rodina s lidským proteinem |
|---|---|---|---|---|
| 1 | SdhA | SDHA _ČLOVĚK | Sukcinát dehydrogenáza [ubichinon] flavoproteinová podjednotka, mitochondriální | Pfam PF00890, Pfam PF02910 |
| 2 | SdhB | SDHB _ČLOVĚK | Sukcinát dehydrogenáza [ubichinon] podjednotka železo-síra, mitochondriální | Pfam PF13085, Pfam PF13183 |
| 3 | SdhC | C560_HUMAN | Sukcinát dehydrogenáza cytochrom b560 podjednotka, mitochondriální | Pfam PF01127 |
| 4 | SdhD | DHSD_HUMAN | Sukcinát dehydrogenáza [ubichinon] cytochrom b malá podjednotka, mitochondriální | Pfam PF05328 |
Vazebné místo pro ubichinon
Dva výrazné ubichinon vazebná místa lze rozpoznat na savčím SDH - matrix-proximální QP a matice-distální QD. Vazebné místo pro ubichinon Qp, které vykazuje vyšší afinitu k ubichinonu, se nachází v mezeře složené ze SdhB, SdhC a SdhD. Ubichinon je stabilizován boční řetězy His207 podjednotky B, Ser27 a Arg31 podjednotky C a Tyr83 podjednotky D. Chinonový kruh je obklopen Ile28 podjednotky C a Pro160 podjednotky B. Tyto zbytky spolu s Il209, Trp163 a Trp164 podjednotky B a Ser27 (atom C) podjednotky C tvoří hydrofobní prostředí chinon -vázací kapsa Qp.[6] Naproti tomu vazebné místo pro ubichinon QD, který leží blíže k mezimembránovému prostoru, je složen pouze z SdhD a má nižší afinitu k ubichinonu.[7]
Sukcinátové vazebné místo
SdhA poskytuje vazebné místo pro oxidace z sukcinát. The boční řetězy Thr254, His354 a Arg399 podjednotky A stabilizují molekula zatímco FAD oxiduje a nese elektrony na první z klastry železo-síra, [2Fe-2S].[8] To je vidět na obrázku 5.
Redoxní centra
The sukcinát -závazné místo a ubichinon - vazebné místo je spojeno řetězcem redoxních center včetně FAD a žehlička -síra shluky. Tento řetězec sahá přes 40 Å skrz enzym monomer. Všechny vzdálenosti od středu k okraji jsou menší než doporučený limit 14 Å fyziologický elektronový přenos.[4] Tento elektronový přenos je znázorněno na obrázku 8.
Podjednotka E
| SdhE | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Struktura NMR proteinu NMA1147 z Neisseria meningitidis. Konsorcium pro strukturální genomiku na severovýchod se zaměřuje na mr19 | |||||||||
| Identifikátory | |||||||||
| Symbol | SdhE | ||||||||
| Pfam | PF03937 | ||||||||
| InterPro | IPR005631 | ||||||||
| |||||||||
v molekulární biologie, proteinová doména pojmenovaný SDH5 je také pojmenován SdhE, což je zkratka pro protein sukcinát dehydrogenázy E. V minulosti byl také pojmenován YgfY a DUF339.[9] Jiným názvem pro SdhE je sukcinátdehydrogenázový montážní faktor 2 (Sdhaf2).[10] Tento protein patří do skupiny vysoce konzervovaný malý bílkoviny nalezené v obou eukaryoty a prokaryoty, včetně NMA1147 z Neisseria meningitidis [11] a YgfY z Escherichia coli.[12] Protein SdhE se nachází na mitochondriální membrána je to důležité pro vytváření energie prostřednictvím procesu s názvem elektronový transportní řetězec.[9]
Funkce
Funkce SdhE byla popsána jako a příchuť sukcinát dehydrogenázy. SdhE funguje jako kofaktor garde který zahrnuje FAD do SdhA. To má za následek SdhA flavinylaci, která je nutná pro správnou funkci sukcinát dehydrogenázy. Studie ukazují, že bakterie vyžadují SdhE, aby mohly růst sukcinát, s použitím sukcinátu jako svého jediného zdroje uhlík a navíc pro funkci, sukcinátdehydrogenázy, životně důležité složky elektronový transportní řetězec který vyrábí energii.[9]
Struktura
The struktura z nich bílkoviny sestává z a komplex svazek pěti alfa-šroubovic, který se skládá ze svazku se 3 šroubovicemi nahoru a dolů a ortogonálního svazku se 2 šroubovicemi.[12]
Interakce proteinů
SdhE interaguje s katalytickou podjednotkou komplexu sukcinátdehydrogenázy (SDH).[13]
Lidská nemoc
Pojmenovaný lidský gen SDH5, kóduje protein SdhE. Samotný gen je umístěn v chromozomální poloze 11q13.1. Výsledkem jsou mutace ztráty funkce paraganglioma, a neuroendokrinní nádor.[13]
Dějiny
Nedávné studie, které naznačují, že SdhE je nutný pro bakteriální flavinylaci, jsou v rozporu s předchozími myšlenkami o SdhE. Původně bylo navrženo začlenění FAD do bakteriálního flavoproteiny byl autokatalytický proces. Nedávné studie nyní tvrdí, že SdhE je první protein, který je identifikován jako požadovaný pro flavinylaci v bakteriích. Historicky byl protein SdhE kdysi považován za hypotetický protein.[9] YgfY byl také považován za účastníka transkripční regulace.[12]
Sestavení a zrání
Všechny podjednotky lidského mitochondriálního SDH jsou kódovány v jaderné oblasti genom. Po překladu SDHA podjednotka je přemístěna jako apoprotein do mitochondriální matrice. Následně je jedním z prvních kroků kovalentní vazba FAD kofaktor (flavinylace). Zdá se, že tento proces je regulován některými z meziproduktů cyklu trikarboxylových kyselin. Konkrétně sukcinát, isocitrát a citrát stimulují flavinylaci SDHA.[14] V případě eukaryotického Sdh1 (SDHA u savců) je pro proces inkorporace FAD vyžadován jiný protein - jmenovitě Sdh5 v kvasinkách, sukcinátdehydrogenázový montážní faktor 2 (SDHAF2 ) v buňkách savců.
Před vytvořením a heterodimer s podjednotkou SDHB se zdá, že určitá část SDHA s kovalentně vázaným FAD interaguje s jiným montážním faktorem - SDHAF4 (Sdh8 v kvasinkách). Nenavázaný flavinylovaný SDHA dimerizuje s SDHAF4, který slouží jako a garde. Studie naznačují, že tvorba SDHA-SDHB dimeru je narušena v nepřítomnosti SDHAF4, takže chaperonový montážní faktor může usnadnit interakci podjednotek. Navíc se zdá, že SDHAF4 brání ROS generace přijímáním elektronů ze sukcinátu, které mohou být stále oxidovány nevázanou monomerní podjednotkou SDHA.[7]
Fe-S protetické skupiny podjednotky SDHB jsou v mitochondriální matrici preformovány proteinovým komplexem ISU. Komplex je také považován za schopný vložit klastry železa a síry do SDHB během jeho zrání. Studie naznačují, že inzerce klastru Fe-S předchází tvorbě dimeru SDHA-SDHB. Takové začlenění vyžaduje snížení o cystein zbytky v aktivním místě SDHB. Jak na snížené cysteinové zbytky, tak na již začleněné klastry Fe-S jsou vysoce náchylné ROS poškození. Dva další faktory sestavení SDH, SDHAF1 (Sdh6) a SDHAF3 (Sdh7 v kvasinkách) se zdají být zapojeny do zrání SDHB, aby chránily podjednotku nebo dimer SDHA-SDHB před poškozením klastru Fe-S způsobeným ROS.[7]
Sestava hydrofobní kotvy sestávající z podjednotek SDHC a SDHD zůstává nejasný. Zvláště v případě heme b vkládání a dokonce i jeho funkce. Protetická skupina hem B se nejeví jako součást dráhy přenosu elektronů v komplexu II.[15] Kofaktor spíše udržuje stabilitu kotvy.
Mechanismus
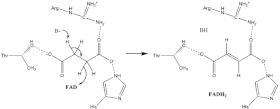
Oxidace jantaru
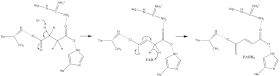
O přesném je známo jen málo sukcinát oxidace mechanismus. Nicméně Krystalická struktura ukázat to FAD, Glu255, Arg286 a His242 podjednotky A (nejsou zobrazeny) jsou vhodnými kandidáty pro počáteční deprotonace krok. Poté existují dva možné eliminační mechanismy: E2 nebo E1cb. Při eliminaci E2 je mechanismus koordinovaný. Základní zbytek nebo kofaktor deprotonuje alfa uhlík a FAD přijímá hydrid z beta uhlík, oxidující svázaný sukcinát na fumarát —Odkaz na obrázek 6. V E1cb, an izolovat je vytvořen meziprodukt, zobrazený na obrázku 7 FAD přijímá hydrid. Je zapotřebí dalšího výzkumu, aby se určilo, kterému eliminačnímu mechanismu sukcinát v sukcinátdehydrogenáze prochází. Oxidovaný fumarát, nyní volně vázán na Aktivní stránky, má možnost opustit protein.
Tunelování elektronů
Po elektrony jsou odvozeny od sukcinát oxidace přes FAD, tunelují podél relé [Fe-S], dokud nedosáhnou klastru [3Fe-4S]. Tyto elektrony jsou následně převedeny na čekající ubichinon molekula v rámci Aktivní stránky. The Žehlička -Síra elektron tunelovací systém je zobrazen na obrázku 9.
Redukce ubichinonu


O1 karbonyl kyslík z ubichinon je orientován na aktivní místo (obrázek 4) uživatelem vodíková vazba interakce s Tyr83 podjednotky D. Přítomnost elektrony v klastru [3Fe-4S] železná síra vyvolává pohyb ubichinon do druhé orientace. To usnadňuje sekundu vodíková vazba interakce mezi O4 karbonylová skupina z ubichinon a Ser27 podjednotky C. Po prvním singlu elektron snížení krok, a semichinon vznikají radikální druhy. Druhý elektron přijíždí z klastru [3Fe-4S], aby poskytl plnou redukci ubichinon na ubichinol. Tento mechanismus ubichinon redukce je zobrazena na obrázku 8.
Protetická skupina hemu
Ačkoli funkčnost heme v sukcinát dehydrogenáze je stále zkoumán, některé studie[kým? ] tvrdili, že první elektron doručeno ubichinon přes [3Fe-4S] může tunelovat tam a zpět mezi heme a ubichinon středně pokročilí. Tímto způsobem heme kofaktor působí jako elektron dřez. Jeho rolí je zabránit interakci meziproduktu s molekulární kyslík k výrobě reaktivní formy kyslíku (ROS). The heme skupina ve vztahu k ubichinon, je zobrazen na obrázku 4.
Rovněž bylo navrženo, aby brána mechanismus může být na místě, aby se zabránilo elektrony z tunelování přímo do heme z klastru [3Fe-4S]. Potenciální kandidát je zbytek His207, který leží přímo mezi shlukem a heme. His207 podjednotky B je v přímé blízkosti klastru [3Fe-4S], vázaného ubichinon a heme; a mohl modulovat elektron tok mezi těmito redox centry.[16]
Přenos protonů
Chcete-li plně snížit chinon v SQR, dva elektrony stejně jako dva protony jsou potřeba. Tvrdilo se, že a molekula vody (HOH39) dorazí na Aktivní stránky a je koordinován His207 podjednotky B, Arg31 podjednotky C a Asp82 podjednotky D. semichinon druh je protonoval podle protony dodáno z HOH39, dokončení ubichinon snížení na ubichinol. His207 a Asp82 s největší pravděpodobností tento proces usnadňují. Další studie tvrdí, že Tyr83 podjednotky D je koordinován s blízkým histidin stejně jako O1 karbonyl kyslík z ubichinon. The histidin zbytek snižuje pKa z tyrosin, takže je vhodnější darovat jeho proton na redukované ubichinon středně pokročilí.
Inhibitory
Existují dvě odlišné třídy inhibitorů komplexu II: ty, které se vážou v jantarové kapse a ty, které se vážou v ubichinonové kapse. Mezi inhibitory ubichinonového typu patří karboxin a thenoyltrifluoraceton. Inhibitory analogu sukcinátu zahrnují syntetickou sloučeninu malonát stejně jako meziprodukty cyklu TCA, malát a oxaloacetát. Oxaloacetát je ve skutečnosti jedním z nejúčinnějších inhibitorů komplexu II. Proč by běžný meziprodukt cyklu TCA inhiboval komplex II, není zcela objasněno, i když může hrát ochrannou roli při minimalizaci produkce superoxidu zprostředkovaného reverzním elektronem v komplexu I.[17] Atpenin 5a jsou vysoce účinné inhibitory komplexu II napodobující vazbu ubichinonu.
Inhibitory ubichinonového typu byly použity jako fungicidy v zemědělství od 60. let. Karboxin se používal hlavně k léčbě nemocí způsobených bazidiomycety jako rezavé stonky a Rhizoctonia nemoci. V poslední době byly vyvinuty další sloučeniny se širším spektrem proti řadě rostlinných patogenů boskalid, penthiopyrad a fluopyram.[18] Některé zemědělsky důležité houby nejsou vůči členům nové generace inhibitorů ubichinonového typu citlivé [19]
Role v nemoci
Základní role sukcinát-koenzym Q reduktázy v elektronový přenosový řetězec z mitochondrie dělá to zásadní ve většině mnohobuňečný organismy, odstranění tohoto enzym z genom Rovněž bylo prokázáno, že je smrtící v embryonálním stadiu u myší.
- Mutace SdhA mohou vést k Leighův syndrom, mitochondriální encefalopatie, a optická atrofie.
- Mutace SdhB mohou vést k tumorogenezi v chromafinové buňky, způsobující třídu nádorů známou jako nedostatek sukcinát dehydrogenázy počítaje v to dědičný paraganglioma a dědičný feochromocytom nedostatek sukcinát dehydrogenázy renální karcinom a nedostatek sukcinát dehydrogenázy gastrointestinální stromální tumor (PODSTATA).[20] Nádory bývají zhoubný. Může také vést ke zkrácení životnosti a zvýšené produkci superoxid ionty.
- Mutace SdhC mohou vést ke zkrácení životnosti, zvýšené produkci superoxid ionty, dědičný paraganglioma a dědičný feochromocytom. Nádory bývají benigní. Tyto mutace jsou neobvyklé.
- Mutace SdhD mohou vést k dědičný paraganglioma a dědičný feochromocytom. Nádory bývají benigní, a vyskytují se často v oblastech hlavy a krku. Tyto mutace mohou také snížit životnost a zvýšit produkci superoxid ionty.
Savčí sukcinátdehydrogenáza funguje nejen v mitochondriální výrobu energie, ale také hraje roli v kyslík snímání a nádor potlačení; a proto je předmětem probíhajícího výzkumu.
Snížené hladiny mitochondriálního enzymu sukcinát dehydrogenázy (SDH), hlavního prvku komplexu II, jsou pozorovány posmrtně v mozcích pacientů s Huntingtonovou chorobou a defekty energetického metabolismu byly identifikovány u presymptomatických i symptomatických pacientů s HD.[21]
Viz také
Reference
- ^ Oyedotun KS, Lemire BD (březen 2004). „Kvartérní struktura Saccharomyces cerevisiae sukcinát dehydrogenázy. Modelování homologie, dokování kofaktorů a simulační studie molekulární dynamiky“. The Journal of Biological Chemistry. 279 (10): 9424–31. doi:10,1074 / jbc.M311876200. PMID 14672929.
- ^ webmaster (04.03.2009). „Použití histochemie ke stanovení vlastností svalu“. Sukcinátdehydrogenáza: Identifikace oxidačního potenciálu. University of California, San Diego. Citováno 2017-12-27.
- ^ Tomitsuka E, Hirawake H, Goto Y, Taniwaki M, Harada S, Kita K (srpen 2003). „Přímý důkaz pro dvě odlišné formy flavoproteinové podjednotky lidského mitochondriálního komplexu II (sukcinát-ubichinon reduktáza)“. Journal of Biochemistry. 134 (2): 191–5. doi:10.1093 / jb / mvg144. PMID 12966066.
- ^ A b Yankovskaya V, Horsefield R, Törnroth S, Luna-Chavez C, Miyoshi H, Léger C a kol. (Leden 2003). "Architektura výroby sukcinát dehydrogenázy a reaktivních forem kyslíku". Věda. 299 (5607): 700–4. Bibcode:2003Sci ... 299..700Y. doi:10.1126 / science.1079605. PMID 12560550.
- ^ Sun F, Huo X, Zhai Y, Wang A, Xu J, Su D a kol. (Červenec 2005). "Krystalová struktura mitochondriálního respiračního membránového proteinového komplexu II". Buňka. 121 (7): 1043–57. doi:10.1016 / j.cell.2005.05.025. PMID 15989954.
- ^ Horsefield R, Yankovskaya V, Sexton G, Whittingham W, Shiomi K, Omura S a kol. (Březen 2006). „Strukturální a výpočetní analýza chinon-vazebného místa komplexu II (sukcinát-ubichinon oxidoreduktáza): mechanismus přenosu elektronů a protonového vedení během redukce ubichinonu“. The Journal of Biological Chemistry. 281 (11): 7309–16. doi:10,1074 / jbc.M508173200. PMID 16407191.
- ^ A b C Van Vranken JG, Na U, Winge DR, Rutter J (prosinec 2014). „Proteinem zprostředkovaná sestava sukcinát dehydrogenázy a jejích kofaktorů“. Kritické recenze v biochemii a molekulární biologii. 50 (2): 168–80. doi:10.3109/10409238.2014.990556. PMC 4653115. PMID 25488574.
- ^ Kenney WC (duben 1975). „Reakce N-ethylmaleimidu v aktivním místě sukcinátdehydrogenázy“. The Journal of Biological Chemistry. 250 (8): 3089–94. PMID 235539.
- ^ A b C d McNeil MB, Clulow JS, Wilf NM, Salmond GP, Fineran PC (2012). „SdhE je konzervovaný protein požadovaný pro flavinylaci sukcinátdehydrogenázy v bakteriích“. J Biol Chem. 287 (22): 18418–28. doi:10,1074 / jbc.M111.293803. PMC 3365757. PMID 22474332.
- ^ https://www.genecards.org/cgi-bin/carddisp.pl?gene=SDHAF2
- ^ Liu G, Sukumaran DK, Xu D, Chiang Y, Acton T, Goldsmith-Fischman S, Honig B, Montelione GT, Szyperski T (květen 2004). „Struktura NMR hypotetického proteinu NMA1147 z Neisseria meningitidis odhaluje zřetelný svazek 5 šroubovice“. Proteiny. 55 (3): 756–8. doi:10,1002 / prot. 20009. PMID 15103637.
- ^ A b C Lim K, Doseeva V, Demirkan ES, Pullalarevu S, Krajewski W, Galkin A, Howard A, Herzberg O (únor 2005). „Krystalová struktura YgfY z Escherichia coli, proteinu, který se může podílet na regulaci transkripce“. Proteiny. 58 (3): 759–63. doi:10,1002 / prot.20337. PMID 15593094.
- ^ A b Hao HX, Khalimonchuk O, Schraders M, Dephoure N, Bayley JP, Kunst H a kol. (Srpen 2009). „SDH5, gen potřebný k ochucení sukcinátdehydrogenázy, je mutován v paragangliomu“. Věda. 325 (5944): 1139–42. Bibcode:2009Sci ... 325.1139H. doi:10.1126 / science.1175689. PMC 3881419. PMID 19628817.
- ^ Brandsch R, Bichler V (červen 1989). "Vazba kovalentního kofaktoru na flavoenzymy vyžaduje specifické efektory". European Journal of Biochemistry. 182 (1): 125–8. doi:10.1111 / j.1432-1033.1989.tb14808.x. PMID 2659351.
- ^ Sun F, Huo X, Zhai Y, Wang A, Xu J, Su D a kol. (Červenec 2005). "Krystalová struktura mitochondriálního respiračního membránového proteinového komplexu II". Buňka. 121 (7): 1043–57. doi:10.1016 / j.cell.2005.05.025. PMID 15989954.
- ^ Tran QM, Rothery RA, Maklashina E, Cecchini G, Weiner JH (říjen 2006). „Chinonové vazebné místo v Escherichia coli sukcinát dehydrogenáze je vyžadováno pro přenos elektronů do hemu b“. The Journal of Biological Chemistry. 281 (43): 32310–7. doi:10,1074 / jbc.M607476200. PMID 16950775.
- ^ Muller FL, Liu Y, Abdul-Ghani MA, Lustgarten MS, Bhattacharya A, Jang YC, Van Remmen H (leden 2008). „Vysoká míra produkce superoxidu v mitochondriích kosterního svalstva dýchajících na komplexních substrátech vázaných na I i na komplex II“. The Biochemical Journal. 409 (2): 491–9. doi:10.1042 / BJ20071162. PMID 17916065.
- ^ Avenot HF, Michailides TJ (2010). „Pokrok v porozumění molekulárním mechanismům a vývoj rezistence na fungicidy inhibující sukcinát dehydrogenázu (SDHI) ve fytopatogenních houbách“. Ochrana plodin. 29 (7): 643. doi:10.1016 / j.cropro.2010.02.019.
- ^ Dubos T, Pasquali M, Pogoda F, Casanova A, Hoffmann L, Beyer M (leden 2013). "Rozdíly mezi sekvencemi sukcinátdehydrogenázy u isopyrazamu citlivých kmenů Zymoseptoria tritici a necitlivých kmenů Fusarium graminearum". Biochemie a fyziologie pesticidů. 105 (1): 28–35. doi:10.1016 / j.pestbp.2012.11.004. PMID 24238287.
- ^ Barletta JA, Hornick JL (červenec 2012). „Nádory s nedostatkem sukcinátu dehydrogenázy: pokrok v diagnostice a klinické důsledky“. Pokroky v anatomické patologii. 19 (4): 193–203. doi:10.1097 / PAP.0b013e31825c6bc6. PMID 22692282.
- ^ Skillings EA, Morton AJ (2016). „Zpožděný nástup a snížené kognitivní deficity předkondicionováním kyselinou 3-nitropropionovou závisí na pohlaví a délce opakování CAG u myšího modelu R6 / 2 Huntingtonovy nemoci.“ Journal of Huntington's Disease. 5 (1): 19–32. doi:10,3233 / JHD-160189. PMID 27031731.