Nikotinový acetylcholinový receptor - Nicotinic acetylcholine receptor


Nikotinové acetylcholinové receptorynebo nAChRs, jsou receptor polypeptidy které reagují na neurotransmiter acetylcholin. Nikotinové receptory také reagují na léky, jako je agonista nikotin. Nacházejí se v centrálním a periferním nervovém systému, svalu a mnoha dalších tkáních mnoha organismů. Na neuromuskulární spojení jsou primárním receptorem ve svalu pro komunikaci motor-nerv-sval, který řídí svalovou kontrakci. V periferním nervovém systému: (1) přenášejí odchozí signály z presynaptických do postsynaptických buněk v sympatickém a parasympatickém nervovém systému a (2) jsou to receptory nacházející se na kosterním svalu, které přijímají acetylcholin uvolněný k signalizaci svalové kontrakce. V imunitním systému regulují nAChR zánětlivé procesy a signalizují odlišnými intracelulárními cestami.[1] v hmyz, cholinergní systém je omezen na centrální nervový systém.[2]
Jsou brány v úvahu nikotinové receptory cholinergní receptory, protože reagují na acetylcholin. Nikotinové receptory dostaly své jméno nikotin který nestimuluje muskarinové acetylcholinové receptory ale místo toho se selektivně váže na nikotinové receptory.[3][4][5] Muskarinový acetylcholinový receptor také dostává svůj název od chemické látky, která se selektivně váže na tento receptor - muskarin.[6] Samotný acetylcholin se váže na muskarinové i nikotinové acetylcholinové receptory.[7]
Tak jako ionotropní receptory, nAChR jsou přímo spojeny s iontovými kanály. Nové důkazy naznačují, že tyto receptory lze také použít druhí poslové (tak jako metabotropní receptory v některých případech.[8] Nikotinové acetylcholinové receptory jsou nejlépe studované z ionotropních receptorů.[3]
Jelikož nikotinové receptory pomáhají přenášet odchozí signály pro sympatické a parasympatické systémy, interferují s přenosem těchto signálů antagonisté nikotinových receptorů, jako je hexamethonium. Například antagonisté nikotinového receptoru interferují s baroreflex který normálně upravuje změny krevního tlaku sympatickou a parasympatickou stimulací srdce.[Citace je zapotřebí ]
Struktura
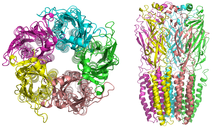
Nikotinové receptory s molekulovou hmotností 290 kDa,[9] jsou tvořeny pěti podjednotkami, uspořádanými symetricky kolem středu pór.[3] Každá podjednotka obsahuje čtyři transmembránové domény s N-a C-koncem umístěným extracelulárně. Mají podobnosti s GABAA receptory, glycinové receptory a typ 3 serotoninové receptory (což jsou všechny ionotropní receptory) nebo podpis Cys-loop proteiny.[10]
U obratlovců jsou nikotinové receptory široce klasifikovány do dvou podtypů na základě jejich primárních míst exprese: svalový typ nikotinové receptory a neuronového typu nikotinové receptory. V receptorech svalového typu, které se nacházejí na neuromuskulárním spojení, jsou receptory buď embryonální forma složená z podjednotek α1, β1, γ a δ v poměru 2: 1: 1: 1, nebo dospělá forma složená z α1, β1, δ a ε podjednotky v poměru 2: 1: 1: 1.[3][4][5][11] Neuronální podtypy jsou různé homomerní (všechny jeden typ podjednotky) nebo heteromerní (alespoň jedna α a jedna β) kombinace dvanácti různých podjednotek nikotinových receptorů: α2 - α10 a β2 - β4. Mezi příklady neuronových podtypů patří: (α4)3(β2)2, (α4)2(β2)3, (α3)2(β4)3, α4α6β3 (β2)2, (α7)5, a mnoho dalších. U receptorů svalového i neuronového typu jsou podjednotky velmi podobné, zejména v hydrofobní regionech.[Citace je zapotřebí ]
Řada studií elektronové mikroskopie a rentgenové krystalografie poskytla strukturní informace s velmi vysokým rozlišením pro svalové a neuronální nAChR a jejich vazebné domény.[9][12][13][14]
Vazba na receptor
Stejně jako u všech iontových kanálů řízených ligandem vyžaduje otevření pórů kanálu nAChR vazbu chemického posla. Několik různých termínů se používá k označení molekul, které vážou receptory, jako je např ligand, agonista nebo vysílač. Stejně jako endogenní agonista acetylcholin, agonisté NAChR zahrnout nikotin, epibatidin, a cholin. Mezi nikotinové antagonisty, které blokují receptor, patří mecamylamin, dihydro-β-erytroidin a hexamethonium.[Citace je zapotřebí ]
Ve svalových nAChR jsou vazebná místa acetylcholinu umístěna na rozhraní podjednotek α a ε nebo δ. V neuronálních nAChR je vazebné místo lokalizováno na rozhraní podjednotky a a p nebo mezi dvěma podjednotkami α v případě a7 receptorů. Vazebné místo se nachází v extracelulární doméně poblíž N konec.[4][15] Když se agonista váže na web, všechny přítomné podjednotky procházejí konformační změnou a kanál je otevřen[16] a otevře se pór o průměru asi 0,65 nm.[4]
Otevření kanálu
Nikotinové AChR mohou existovat v různých vzájemně konvertibilních konformačních stavech. Vazba agonisty stabilizuje otevřené a znecitlivělý státy. Za normálních fyziologických podmínek potřebuje receptor k otevření přesně dvě molekuly ACh.[17] Otevření kanálu umožňuje kladné nabití ionty pohybovat se přes něj; zejména, sodík vstupuje do buňky a draslík východy. Čistý tok kladně nabitých iontů je dovnitř.
NAChR je neselektivní kationtový kanál, což znamená, že může procházet několik různých pozitivně nabitých iontů.[3] Je propustný pro Na+ a K.+, s některými kombinacemi podjednotek, které jsou také propustné pro Ca2+.[4][18][19] Množství sodíku a draslíku kanály umožňují jejich póry (jejich vodivost ) se pohybuje od 50–110 pS, s vodivostí v závislosti na konkrétním složení podjednotky a na propustném iontu.[20]
Mnoho neuronálních nAChR může ovlivnit uvolňování dalších neurotransmiterů.[5] Kanál se obvykle otevírá rychle a má tendenci zůstat otevřený až do agonista šíří pryč, což obvykle trvá asi 1 milisekunda.[4] AChR se však mohou spontánně otevřít bez vázaných ligandů nebo se mohou spontánně uzavřít s vázanými ligandy a mutace v kanálu mohou posunout pravděpodobnost kterékoli události.[21][16] Vazba ACh proto mění pravděpodobnost otevření pórů, která se zvyšuje s tím, jak se více ACh váže.
NAChR není schopen vázat ACh, když je vázán na některý z hadí jed α-neurotoxiny. Tyto a-neurotoxiny se antagonisticky silně a nekovalentně vážou na nAChR kosterních svalů a v neuronech, čímž blokují působení ACh na postsynaptické membráně, inhibují tok iontů a vedou k paralýze a smrti. NAChR obsahuje dvě vazebná místa pro neurotoxiny hadího jedu. Pokrok v objevování dynamiky vazebného působení těchto stránek se ukázal jako obtížný, i když nedávné studie používají normální mód dynamika[22] pomohly předpovědět povahu vazebných mechanismů hadích toxinů a ACh k nAChR. Tyto studie ukázaly, že za otevření pórů je pravděpodobně zodpovědný pohyb podobný kroucení způsobený vazbou ACh a že jedna nebo dvě molekuly α-bungarotoxin (nebo jiný alfa-neurotoxin s dlouhým řetězcem) stačí k zastavení tohoto pohybu. Zdá se, že toxiny spojují dohromady sousední receptorové podjednotky, čímž brání zkroucení a tím i otevíracímu pohybu.[23]
Účinky
Aktivace receptorů nikotinem mění stav neurony prostřednictvím dvou hlavních mechanismů. Na jedné straně pohyb kationty způsobuje a depolarizace plazmatické membrány (což vede k excitační postsynaptický potenciál v neurony ) vedoucí k aktivaci napěťově řízené iontové kanály. Na druhou stranu vstup vápníku působí přímo nebo nepřímo na různé intracelulární kaskády. To vede například k regulaci činnosti některých geny nebo vydání neurotransmitery.[Citace je zapotřebí ]
Regulace receptoru
Desenzibilizace receptoru
Desenzibilizace receptorů vázaná na ligand byla nejprve charakterizována Katzem a Thesleffem na nikotinovém acetylcholinovém receptoru.[24]
Dlouhodobé nebo opakované vystavení stimulu má často za následek sníženou citlivost tohoto receptoru na stimul, který se nazývá desenzibilizace. Funkce nAChR může být modulována fosforylací[25] aktivací proteinových kináz závislých na druhém poslovi. PKA[24] a PKC,[26] stejně jako tyrosinkinázy,[27] Bylo prokázáno, že fosforylují nAChR, což vede k jeho desenzibilizaci. Bylo hlášeno, že po prodloužené expozici receptoru agonistovi agonista sám způsobí agonistou vyvolanou konformační změnu v receptoru, což vede k desenzibilizaci receptoru.[28]
Desenzibilizované receptory se mohou vrátit do prodlouženého otevřeného stavu, když je agonista vázán v přítomnosti pozitivního alosterického modulátoru, například PNU-120596.[29] Existují také důkazy, které naznačují, že specifické chaperonové molekuly mají regulační účinky na tyto receptory.[30]
Role
Podjednotky nikotinových receptorů patří do multigenové rodiny (16 členů u lidí) a sestavení kombinací podjednotek vede k velkému počtu různých receptorů (více informací viz Databáze iontových kanálů s ligandovou bránou ). Tyto receptory jsou vysoce variabilní kinetický, elektrofyziologické a farmakologické vlastnosti, reagovat na nikotin jinak, při velmi odlišných účinných koncentracích. Tato funkční rozmanitost jim umožňuje účastnit se dvou hlavních typů neurotransmise. Klasický synaptický přenos (kabelový přenos) zahrnuje uvolňování vysokých koncentrací neurotransmiteru působících na bezprostředně sousedící receptory. V porovnání, parakrin přenos (objemový přenos) zahrnuje neurotransmitery vydáno uživatelem synaptická tlačítka, které pak difundují extracelulárním médiem, dokud nedosáhnou svých receptorů, které mohou být vzdálené.[31] Nikotinové receptory lze také nalézt na různých synaptických místech; například svalový nikotinový receptor funguje vždy postsynapticky. Neuronální formy receptoru lze nalézt jak postsynapticky (zapojené do klasické neurotransmise), tak předsynapticky[32] kde mohou ovlivnit uvolňování více neurotransmiterů.
Podjednotky
Bylo identifikováno 17 podjednotek nAChR obratlovců, které jsou rozděleny na podjednotky svalového a neuronového typu. Přestože je a8 podjednotka / gen přítomna u ptačích druhů, jako je kuře, není přítomna u lidských nebo savčích druhů.[33]
Podjednotky nAChR byly rozděleny do 4 podrodin (I-IV) na základě podobností v proteinové sekvenci.[34] Kromě toho byla podrodina III dále rozdělena do 3 typů.
| Neuronální typ | Svalový typ | ||||
| Já | II | III | IV | ||
|---|---|---|---|---|---|
| α9, α10 | α7, α8 | 1 | 2 | 3 | α1, β1, δ, y, ε |
| α2, α3, α4, α6 | β2, β4 | β3, α5 | |||
- geny α: CHRNA1 (sval), CHRNA2 (neuronální), CHRNA3, CHRNA4, CHRNA5, CHRNA6, CHRNA7, CHRNA8, CHRNA9, CHRNA10
- β geny: CHRNB1 (sval), CHRNB2 (neuronální), CHRNB3, CHRNB4
- Jiné geny: CHRND (delta), CHRNE (epsilon), CHRNG (gama)
Neuronální nAChR jsou transmembránové proteiny ta forma pentamerické struktury shromážděné z rodiny podjednotek složených z α2-α10 a β2-β4.[35] Tyto podjednotky byly objeveny od poloviny 80. let do počátku 90. let, kdy byly klonovány cDNA pro více podjednotek nAChR z mozků potkanů a kuřat, což vedlo k identifikaci jedenácti různých genů (dvanáct u kuřat), které kódují neuronální podjednotky nAChR; Identifikované geny podjednotky byly pojmenovány α2 – α10 (α8 se vyskytuje pouze u kuřat) a β2 – β4.[36] Bylo také zjištěno, že různé kombinace podjednotek mohou tvořit funkční nAChR, které mohou být aktivovány acetylcholin a nikotin a různé kombinace podjednotek generují podtypy nAChR s různými funkčními a farmakologickými vlastnostmi.[37] Když jsou exprimovány samostatně, jsou α7, a8, a9 a a10 schopné tvořit funkční receptory, ale jiné α podjednotky vyžadují přítomnost p podjednotek, aby se vytvořily funkční receptory.[35] U savců bylo zjištěno, že nAchR podjednotky jsou kódovány 17 geny a z nich je v mozku exprimováno devět genů kódujících a-podjednotky a tři kódující p-podjednotky. β2 podjednotky obsahující nAChR (β2nAChRs) a α7nAChR jsou široce exprimovány v mozku, zatímco jiné podjednotky nAChR mají omezenější expresi.[38]
CHRNA5 / A3 / B4
Důležitý genový klastr nAchR (CHRNA5 / A3 / B4) obsahuje geny kódující podjednotky a5, a3 a p4. Byly identifikovány genetické studie jednonukleotidové polymorfismy (SNP) v chromozomálním lokusu kódujícím tyto tři geny nAChR jako rizikové faktory pro závislost na nikotinu, rakovina plic, chronická obstrukční plicní nemoc, alkoholismus, a onemocnění periferních tepen.[35][39] Geny podjednotky CHRNA5 / A3 / B4 nAChR se nacházejí v těsném klastru v chromozomální oblasti 15q24–25. Podjednotky nAChR kódované tímto lokusem tvoří převládající podtypy nikotinových receptorů exprimované v periferní nervový systém (PNS) a další klíč centrální nervový systém (CNS) weby, například mediální habenula, struktura mezi limbickým předním mozkem a středním mozkem zapojená do hlavních cholinergních obvodových drah.[35] Další výzkum genů CHRNA5 / A3 / B4 ukázal, že „neuronální“ geny nAChR jsou také exprimovány v jiných než neuronálních buňkách, kde se účastní různých základních procesů, jako je zánět.[40] Geny CHRNA5 / A3 / B4 jsou koexprimovány v mnoha buněčných typech a transkripční aktivity promotorových oblastí tří genů jsou regulovány mnoha stejnými transkripčními faktory, což ukazuje, že jejich shlukování může odrážet kontrolu genové exprese.[35]
CHRNA6 / CHRNB3
CHRNB3 a CHRNA6 jsou také seskupeny v genovém klastru umístěném na 8p11.[39] Několik studií ukázalo, že SNPS v CHRNB3 – CHRNA6 byly spojeny se závislostí na nikotinu a kouřením, jako jsou dva SNP v CHRNB3, rs6474413 a rs10958726.[39] Genetické variace v této oblasti také vykazují vliv na náchylnost k užívání návykových látek, včetně konzumace kokainu a alkoholu.[41] Nikotinové receptory obsahující podjednotky α6 nebo p3 exprimované v oblastech mozku, zejména v ventrální tegmentální oblast a substantia nigra, jsou důležité pro drogové chování kvůli jejich roli v dopamin uvolnění.[42] Genetická variace v těchto genech může změnit citlivost na zneužívané léky mnoha způsoby, včetně změny aminokyselinové struktury proteinu nebo způsobit změny v transkripční a translační regulaci.[41]
CHRNA4 / CHRNB2
Další dobře studované geny nAChR zahrnují CHRNA4 a CHRNB2, které byly asociovány jako Autosomálně dominantní noční čelní laloková epilepsie (ADNFLE) geny.[39][43] Obě tyto podjednotky nAChR jsou přítomny v mozku a výskyt mutací v těchto dvou podjednotkách způsobuje generalizovaný typ epilepsie. Mezi příklady patří inzertní mutace CHRNA4 776ins3, která je spojena s nočními záchvaty a psychiatrickými poruchami, a mutace CHRNB2 I312M, která podle všeho způsobuje nejen epilepsii, ale také velmi specifické kognitivní deficity, jako jsou deficity v učení a paměti.[43][44] Mezi těmito dvěma geny se přirozeně vyskytují genetické variace a analýza jednoduchých nukleotidových polymorfismů (SNP) a další modifikace genů ukazují vyšší variace v genu CHRNA4 než v genu CHRNB2, což znamená, že nAChR β2, protein kódovaný CHRNB2, se asociuje s více podjednotek než α4. CHRNA2 byla také hlášena jako třetí kandidát na noční záchvaty čelního laloku.[39][43]
CHRNA7
Několik studií uvádí asociaci mezi CHRNA7 a endofenotypy psychiatrických poruch a závislosti na nikotinu, což přispívá k významnému klinickému významu α7 a je na něm prováděn výzkum.[43] CHRNA7 byl jedním z prvních genů, o kterých se předpokládalo, že se jich účastní schizofrenie. Studie identifikovaly několik polymorfismů promotoru CHRNA7, které snižují transkripční aktivitu genů spojenou se schizofrenií, což odpovídá nálezu snížených hladin a7 nAChR v mozku schizofrenních pacientů.[43] Ve studiích po smrti jedinců se schizofrenií bylo zjištěno, že oba podtypy nAChR, α4β2 a α7, jsou významně sníženy.[45] Míra kouření je navíc významně vyšší u pacientů se schizofrenií, což znamená, že kouření nikotinu může být formou samoléčby.[46]
Pozoruhodné variace
Nikotinové receptory jsou pentamery těchto podjednotek; tj. každý receptor obsahuje pět podjednotek. Existuje tedy obrovský potenciál variací těchto podjednotek. Některé z nich se však vyskytují častěji než jiné. Mezi nejběžněji vyjádřené podtypy patří (α1)2β1δε (dospělý typ svalu), (α3)2(β4)3 (typ ganglion), (α4)2(β2)3 (Typ CNS) a (α7)5 (jiný typ CNS).[47] Následuje srovnání:
| Receptorový typ | Umístění | Účinek; funkce | Nikotinové agonisty | Nikotinové antagonisty |
|---|---|---|---|---|
| Svalový typ: (α1)2β1δε[47] nebo (α1)2β1δγ | Neuromuskulární spojení | EPSP, hlavně zvýšením Na+ a K.+ propustnost | ||
| Ganglionský typ: (α3)2(β4)3 | autonomní ganglia | EPSP, hlavně zvýšením Na+ a K.+ propustnost | ||
| Heteromerní typ CNS: (α4)2(β2)3 | Mozek | Pošta- a presynaptické buzení,[47] hlavně zvýšením Na+ a K.+ propustnost. Hlavní podtyp podílející se na zvyšování pozornosti a obohacující účinky nikotinu a také patofyziologie závislosti na nikotinu.[49][50][51] | ||
| Další typ CNS: (α3)2(β4)3 | Mozek | Pošta- a presynaptické buzení | ||
| Homomerní typ CNS: (α7)5 | Mozek | Pošta- a presynaptické buzení,[47] hlavně zvýšením Na+, K.+ a Ca.2+ propustnost. Hlavní podtyp podílející se na některých kognitivních účincích nikotinu.[52] Navíc aktivace (α7)5 může zlepšit neurovaskulární vazebnou reakci u neurodegenerativního onemocnění[53] a neurogeneze u ischemické cévní mozkové příhody.[54] Podílí se také na proangiogenních účincích nikotinu a urychluje progresi chronického onemocnění ledvin u kuřáků.[55][56][57] |
Viz také
- Muskarinový acetylcholinový receptor
- Muskarinový agonista
- Muskarinový antagonista
- Nikotinový agonista
- Nikotinový antagonista
Reference
- ^ Lu B, Kwan K, Levine YA, Olofsson PS, Yang H, Li J a kol. (Srpen 2014). „Signalizace a7 nikotinového acetylcholinového receptoru inhibuje aktivaci inflammasomu tím, že brání uvolňování mitochondriální DNA“. Molekulární medicína. 20 (1): 350–8. doi:10.2119 / molmed.2013.00117. PMC 4153835. PMID 24849809.
- ^ Yamamoto I (1999). „Nikotin nikotinoidům: 1962 až 1997“. Nikotinoidní insekticidy a nikotinový acetylcholinový receptor. s. 3–27. doi:10.1007/978-4-431-67933-2_1. ISBN 978-4-431-68011-6.
- ^ A b C d E F G h i j k Purves D, Augustine GJ, Fitzpatrick D, Hall WC, LaMantia A, McNamara JO, White LE (2008). Neurovědy (4. vydání). Sinauer Associates. str.122 –6. ISBN 978-0-87893-697-7.
- ^ A b C d E F Siegel GJ, Agranoff BW, Fisher SK, Albers RW, Uhler MD (1999). „Základní neurochemie: molekulární, buněčné a lékařské aspekty“. Fyziologie a farmakologie receptorů GABA (6. vydání). Americká společnost pro neurochemii. Citováno 2008-10-01.
- ^ A b C Itier V, Bertrand D (srpen 2001). "Neuronální nikotinové receptory: od proteinové struktury k funkci". FEBS Dopisy. 504 (3): 118–25. doi:10.1016 / s0014-5793 (01) 02702-8. PMID 11532443.
- ^ Ishii M, Kurachi Y (1. října 2006). "Muskarinové acetylcholinové receptory". Současný farmaceutický design. 12 (28): 3573–81. doi:10.2174/138161206778522056. PMID 17073660.
- ^ Lott EL, Jones EB (2020). "Cholinergní toxicita". StatPearls. Publishing StatPearls. PMID 30969605.
- ^ Kabbani N, Nordman JC, Corgiat BA, Veltri DP, Shehu A, Seymour VA, Adams DJ (prosinec 2013). „Jsou nikotinové acetylcholinové receptory spojeny s G proteiny?“. BioEssays. 35 (12): 1025–34. doi:10.1002 / bies.201300082. PMID 24185813.
- ^ A b Unwin N (březen 2005). "Rafinovaná struktura nikotinového acetylcholinového receptoru při rozlišení 4A". Journal of Molecular Biology. 346 (4): 967–89. doi:10.1016 / j.jmb.2004.12.031. PMID 15701510.
- ^ Cascio M (květen 2004). "Struktura a funkce glycinového receptoru a souvisejících nikotinikoidových receptorů". The Journal of Biological Chemistry. 279 (19): 19383–6. doi:10,1074 / jbc.R300035200. PMID 15023997.
- ^ Giniatullin R, Nistri A, Yakel JL (červenec 2005). „Desenzibilizace nikotinových receptorů ACh: utváření cholinergní signalizace“. Trendy v neurovědách. 28 (7): 371–8. doi:10.1016 / j.tins.2005.04.009. PMID 15979501.
- ^ Brejc K, van Dijk WJ, Klaassen RV, Schuurmans M, van Der Oost J, Smit AB, Sixma TK (květen 2001). „Krystalová struktura proteinu vázajícího ACh odhaluje doménu nikotinových receptorů vázající ligand“. Příroda. 411 (6835): 269–76. Bibcode:2001 Natur.411..269B. doi:10.1038/35077011. PMID 11357122.
- ^ Zouridakis M, Giastas P, Zarkadas E, Chroni-Tzartou D, Bregestovski P, Tzartos SJ (listopad 2014). „Krystalové struktury volných a antagonisticky vázaných stavů lidské extracelulární domény nikotinového receptoru α9“. Přírodní strukturní a molekulární biologie. 21 (11): 976–80. doi:10.1038 / nsmb.2900. PMID 25282151.
- ^ Morales-Perez CL, Noviello CM, Hibbs RE (říjen 2016). „Rentgenová struktura lidského nikotinového receptoru α4β2“. Příroda. 538 (7625): 411–415. Bibcode:2016Natur.538..411M. doi:10.1038 / příroda1985. PMC 5161573. PMID 27698419.
- ^ Squire L (2003). Základní neurověda (2. vyd.). Amsterdam: Acad. Lis. str. 1426. ISBN 978-0-12-660303-3.
- ^ A b Colquhoun D, Sivilotti LG (červen 2004). "Funkce a struktura v glycinových receptorech a některých jejich příbuzných". Trendy v neurovědách. 27 (6): 337–44. CiteSeerX 10.1.1.385.3809. doi:10.1016 / j.tins.2004.04.010. PMID 15165738.
- ^ Aidley DJ (1998). Fyziologie excitabilních buněk (4. vydání). Cambridge, Velká Británie: Cambridge University Press. ISBN 978-0521574150. OCLC 38067558.[stránka potřebná ]
- ^ Beker F, Weber M, Fink RH, Adams DJ (září 2003). „Aktivace muskarinových a nikotinových ACh receptorů odlišně mobilizuje Ca2 + v intrakardiálních gangliových neuronech potkanů“. Journal of Neurophysiology. 90 (3): 1956–64. doi:10.1152 / jn.01079.2002. PMID 12761283.
- ^ Weber M, Motin L, Gaul S, Beker F, Fink RH, Adams DJ (leden 2005). „Intravenózní anestetika inhibují proudy zprostředkované nikotinovým acetylcholinovým receptorem a přechodné Ca2 + v intrakardiálních gangliových neuronech potkanů“. British Journal of Pharmacology. 144 (1): 98–107. doi:10.1038 / sj.bjp.0705942. PMC 1575970. PMID 15644873.
- ^ Mishina M, Takai T, Imoto K, Noda M, Takahashi T, Numa S a kol. (Květen 1986). "Molekulární rozdíl mezi fetální a dospělou formou svalového acetylcholinového receptoru". Příroda. 321 (6068): 406–11. Bibcode:1986 Natur.321..406M. doi:10.1038 / 321406a0. PMID 2423878.
- ^ Einav T, Phillips R (duben 2017). „Monod-Wyman-Changeuxova analýza mutantů iontového kanálu s ligandem“. The Journal of Physical Chemistry B. 121 (15): 3813–3824. arXiv:1701.06122. Bibcode:2017arXiv170106122E. doi:10.1021 / acs.jpcb.6b12672. PMC 5551692. PMID 28134524.
- ^ Levitt M, Sander C, Stern PS (únor 1985). "Dynamika proteinu v normálním režimu: inhibitor trypsinu, crambin, ribonukleáza a lysozym". Journal of Molecular Biology. 181 (3): 423–47. doi:10.1016 / 0022-2836 (85) 90230-x. PMID 2580101.
- ^ Samson AO, Levitt M (duben 2008). „Inhibiční mechanismus acetylcholinového receptoru alfa-neurotoxiny, jak vyplývá z dynamiky normálního režimu“. Biochemie. 47 (13): 4065–70. doi:10.1021 / bi702272j. PMC 2750825. PMID 18327915.
- ^ A b Pitchford S, Day JW, Gordon A, Mochly-Rosen D (listopad 1992). „Desenzitizace nikotinových acetylcholinových receptorů je regulována aktivací vyvolanou akumulací extracelulárního adenosinu“. The Journal of Neuroscience. 12 (11): 4540–4. doi:10.1523 / JNEUROSCI.12-11-04540.1992. PMC 6576003. PMID 1331363.
- ^ Huganir RL, Greengard P (únor 1983). „cAMP-dependentní protein kináza fosforyluje nikotinový acetylcholinový receptor“. Sborník Národní akademie věd Spojených států amerických. 80 (4): 1130–4. Bibcode:1983PNAS ... 80.1130H. doi:10.1073 / pnas.80.4.1130. PMC 393542. PMID 6302672.
- ^ Safran A, Sagi-Eisenberg R, Neumann D, Fuchs S (srpen 1987). „Fosforylace acetylcholinového receptoru proteinkinázou C a identifikace fosforylačního místa v podjednotce delta receptoru“. The Journal of Biological Chemistry. 262 (22): 10506–10. PMID 3038884.
- ^ Hopfield JF, Tank DW, Greengard P, Huganir RL (prosinec 1988). „Funkční modulace nikotinového acetylcholinového receptoru fosforylací tyrosinu“. Příroda. 336 (6200): 677–80. Bibcode:1988Natur.336..677H. doi:10.1038 / 336677a0. PMID 3200319.
- ^ Barrantes FJ (září 1978). "Agonistem zprostředkované změny acetylcholinového receptoru v jeho membránovém prostředí". Journal of Molecular Biology. 124 (1): 1–26. doi:10.1016/0022-2836(78)90144-4. PMID 712829.
- ^ Hurst RS, Hajós M, Raggenbass M, Wall TM, Higdon NR, Lawson JA a kol. (Duben 2005). „Nový pozitivní alosterický modulátor alfa7 neuronového nikotinového acetylcholinového receptoru: charakterizace in vitro a in vivo“. The Journal of Neuroscience. 25 (17): 4396–405. doi:10.1523 / JNEUROSCI.5269-04.2005. PMC 6725110. PMID 15858066.
- ^ Sadigh-Eteghad S, Majdi A, Talebi M, Mahmoudi J, Babri S (květen 2015). „Regulace nikotinových acetylcholinových receptorů u Alzheimerovy choroby: možná role chaperonů“. European Journal of Pharmacology. 755: 34–41. doi:10.1016 / j.ejphar.2015.02.047. PMID 25771456.
- ^ Picciotto MR, Higley MJ, Mineur YS (říjen 2012). „Acetylcholin jako neuromodulátor: cholinergní signalizace formuje funkci a chování nervového systému“. Neuron. 76 (1): 116–29. doi:10.1016 / j.neuron.2012.08.036. PMC 3466476. PMID 23040810.
- ^ Wonnacott S (únor 1997). "Presynaptické nikotinové ACh receptory". Trendy v neurovědách. 20 (2): 92–8. doi:10.1016 / S0166-2236 (96) 10073-4. PMID 9023878.
- ^ Graham A, Court JA, Martin-Ruiz CM, Jaros E, Perry R, Volsen SG a kol. (2002). „Imunohistochemická lokalizace podjednotek nikotinových acetylcholinových receptorů v lidském mozečku“. Neurovědy. 113 (3): 493–507. doi:10.1016 / S0306-4522 (02) 00223-3. PMID 12150770.
- ^ Le Novère N, Changeux JP (únor 1995). „Molekulární vývoj nikotinového acetylcholinového receptoru: příklad multigenové rodiny v excitovatelných buňkách“. Journal of Molecular Evolution. 40 (2): 155–72. Bibcode:1995JMolE..40..155L. doi:10.1007 / BF00167110. PMID 7699721.
- ^ A b C d E Improgo MR, Scofield MD, Tapper AR, Gardner PD (říjen 2010). „Klastr genu CHRNA5 / A3 / B4 na nikotinovém acetylcholinovém receptoru: dvojí role v závislosti na nikotinu a rakovině plic“. Pokrok v neurobiologii. 92 (2): 212–26. doi:10.1016 / j.pneurobio.2010.05.003. PMC 2939268. PMID 20685379.
- ^ Tammimäki A, Horton WJ, Stitzel JA (říjen 2011). „Nedávný pokrok v genové manipulaci a biologii nikotinových acetylcholinových receptorů“. Biochemická farmakologie. 82 (8): 808–19. doi:10.1016 / j.bcp.2011.06.014. PMC 3162071. PMID 21704022.
- ^ Graham A, Court JA, Martin-Ruiz CM, Jaros E, Perry R, Volsen SG a kol. (Září 2002). "Imunohistochemická lokalizace nikotinových podjednotek acetylcholinového receptoru v lidském mozečku". Neurovědy. 113 (3): 493–507. doi:10.1016 / S0306-4522 (02) 00223-3. PMID 12150770.
- ^ Changeux JP (červen 2010). „Závislost na nikotinu a nikotinové receptory: lekce od geneticky modifikovaných myší“. Recenze přírody. Neurovědy. 11 (6): 389–401. doi:10.1038 / nrn2849. PMID 20485364.
- ^ A b C d E Greenbaum L, Lerer B (říjen 2009). „Diferenciální příspěvek genetických variací v mnoha mozkových nikotinových cholinergních receptorech k závislosti na nikotinu: nedávný pokrok a objevující se otevřené otázky“. Molekulární psychiatrie. 14 (10): 912–45. doi:10.1038 / mp.2009.59. PMID 19564872.
- ^ Gahring LC, Rogers SW (leden 2006). "Exprese a funkce neuronového nikotinového acetylcholinového receptoru na nepenonálních buňkách". Deník AAPS. 7 (4): E885-94. doi:10.1208 / aapsj070486. PMC 2750958. PMID 16594641.
- ^ A b Kamens HM, Corley RP, Richmond PA, Darlington TM, Dowell R, Hopfer CJ a kol. (Září 2016). „Důkazy o asociaci mezi nízkofrekvenčními variantami v CHRNA6 / CHRNB3 a antisociální drogovou závislostí“. Genetika chování. 46 (5): 693–704. doi:10.1007 / s10519-016-9792-4. PMC 4975622. PMID 27085880.
- ^ Grady SR, Salminen O, Laverty DC, Whiteaker P, McIntosh JM, Collins AC, Marks MJ (říjen 2007). "Podtypy nikotinových acetylcholinových receptorů na dopaminergních zakončeních myšího striata". Biochemická farmakologie. 74 (8): 1235–46. doi:10.1016 / j.bcp.2007.07.032. PMC 2735219. PMID 17825262.
- ^ A b C d E Steinlein v pořádku, Bertrand D (listopad 2008). „Neuronální nikotinové acetylcholinové receptory: od genetické analýzy po neurologická onemocnění“. Biochemická farmakologie. 76 (10): 1175–83. doi:10.1016 / j.bcp.2008.07.012. PMID 18691557.
- ^ Bertrand D, Elmslie F, Hughes E, Trounce J, Sander T, Bertrand S, Steinlein OK (prosinec 2005). "Mutace CHRNB2 I312M je spojena s epilepsií a zřetelnými deficity paměti". Neurobiologie nemocí. 20 (3): 799–804. doi:10.1016 / j.nbd.2005.05.013. PMID 15964197.
- ^ Breese CR, Lee MJ, Adams CE, Sullivan B, Logel J, Gillen KM a kol. (Říjen 2000). „Abnormální regulace nikotinových receptorů s vysokou afinitou u osob se schizofrenií“. Neuropsychofarmakologie. 23 (4): 351–64. doi:10.1016 / S0893-133X (00) 00121-4. PMID 10989262.
- ^ McLean SL, Grayson B, Idris NF, Lesage AS, Pemberton DJ, Mackie C, Neill JC (duben 2011). „Aktivace nikotinových receptorů α7 zlepšuje deficity kognitivních úkolů u potkanů vyvolané fencyklidinem: důsledky pro léčbu kognitivní dysfunkce u schizofrenie“. Evropská neuropsychofarmakologie. 21 (4): 333–43. doi:10.1016 / j.euroneuro.2010.06.003. hdl:10454/8464. PMID 20630711.
- ^ A b C d Rang HP (2003). Farmakologie (5. vydání). Edinburgh: Churchill Livingstone. ISBN 978-0-443-07145-4.[stránka potřebná ]
- ^ A b Neurosci.pharm - MBC 3320 acetylcholin Archivováno 2007-12-27 na Wayback Machine
- ^ Sarter M (srpen 2015). „Behaviorálně-kognitivní cíle pro cholinergní vylepšení“. Současný názor na behaviorální vědy. 4: 22–26. doi:10.1016 / j.cobeha.2015.01.004. PMC 5466806. PMID 28607947.
- ^ Wu J, Gao M, Shen JX, Shi WX, Oster AM, Gutkin BS (říjen 2013). "Kortikální kontrola funkce VTA a vliv na odměnu za nikotin". Biochemická farmakologie. 86 (8): 1173–80. doi:10.1016 / j.bcp.2013.07.013. PMID 23933294.
- ^ „Nikotin: Biologická aktivita“. Průvodce farmakologií IUPHAR / BPS. Mezinárodní unie základní a klinické farmakologie. Citováno 7. února 2016.
K.is takto; a2p4 = 9900nM [5], a3p2 = 14nM [1], a3p4 = 187nM [1], a4p2 = 1nM [4,6]. Kvůli heterogenitě kanálů nACh jsme neoznačili primární cíl léku pro nikotin, ačkoli se uvádí, že α4β2 je převládajícím subtypem s vysokou afinitou v mozku, který zprostředkovává závislost na nikotinu [2-3].
- ^ Levin ED (květen 2012). "α7-nikotinové receptory a poznání". Aktuální drogové cíle. 13 (5): 602–6. doi:10.2174/138945012800398937. PMID 22300026.
- ^ Sadigh-Eteghad S, Mahmoudi J, Babri S, Talebi M (listopad 2015). „Vliv aktivace alfa-7 nikotinového acetylcholinového receptoru na poškození paměti rozpoznáváním vyvolané beta-amyloidem. Možná role neurovaskulární funkce“. Acta Cirurgica Brasileira. 30 (11): 736–42. doi:10.1590 / S0102-865020150110000003. PMID 26647792.
- ^ Wang J, Lu Z, Fu X, Zhang D, Yu L, Li N a kol. (Květen 2017). „Cesta signalizace nikotinového receptoru alfa-7 se účastní neurogeneze indukované neurony pozitivními na CHAT v subventrikulární zóně“. Výzkum translační mrtvice. 8 (5): 484–493. doi:10.1007 / s12975-017-0541-7. PMC 5704989. PMID 28551702.
- ^ Lee J, Cooke JP (listopad 2012). „Nikotin a patologická angiogeneze“. Humanitní vědy. 91 (21–22): 1058–64. doi:10.1016 / j.lfs.2012.06.032. PMC 3695741. PMID 22796717.
- ^ Jain G, Jaimes EA (říjen 2013). „Signalizace nikotinu a progrese chronického onemocnění ledvin u kuřáků“. Biochemická farmakologie. 86 (8): 1215–23. doi:10.1016 / j.bcp.2013.07.014. PMC 3838879. PMID 23892062.
- ^ Mihalak KB, Carroll FI, Luetje CW (září 2006). „Vareniklin je částečný agonista na alfa4beta2 a plný agonista na alfa7 neuronálních nikotinových receptorech“. Molekulární farmakologie. 70 (3): 801–5. doi:10,1124 / mol. 106,025130. PMID 16766716.
externí odkazy
 Média související s Nikotinové acetylcholinové receptory na Wikimedia Commons
Média související s Nikotinové acetylcholinové receptory na Wikimedia Commons- Vypočítaná prostorová poloha nikotinového acetylcholinového receptoru v lipidové dvojvrstvě