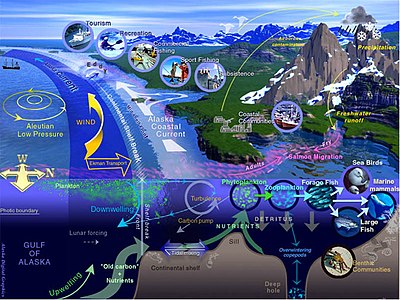
Mořský život - Marine life

| Část série přehledů o |
| mořský život |
|---|
mořský životnebo mořský život nebo oceánský život, je rostliny, zvířata a další organismy kteří žijí v slaná voda moře nebo oceánu, nebo brakická voda pobřežních ústí řek. Na základní úrovni ovlivňuje mořský život povahu planety. Většinou mořské organismy mikroorganismy, produkují kyslík a sekvestrační uhlík. Pobřeží jsou částečně tvarována a chráněna mořským životem a některé mořské organismy dokonce pomáhají vytvářet novou zemi. Termín námořní pochází z latinský kobyla, což znamená moře nebo oceán.
Většina forem života se původně vyvinula mořská stanoviště. Oceány podle objemu poskytují přibližně 90 procent životního prostoru na planetě.[2] Nejdříve obratlovců se objevil v podobě Ryba,[3] kteří žijí výhradně ve vodě. Z některých se vyvinulo obojživelníci kteří tráví části svého života ve vodě a části na souši. Jiné ryby se vyvinuly v suchozemské savce a následně se vrátily do oceánu jako těsnění, delfíni nebo velryby. Rostlinné formy jako např kelp a řasy rostou ve vodě a jsou základem pro některé podvodní ekosystémy. Plankton tvoří obecný základ oceánu potravní řetězec, zejména fytoplankton které jsou klíčové prvovýrobci.
Mořští bezobratlí vykazují širokou škálu modifikací, aby přežili ve špatně okysličených vodách, včetně dýchacích trubic jako v sifony měkkýšů. Ryby mají žábry namísto plíce, i když některé druhy ryb, například lungfish, mít obojí. mořští savci, jako jsou delfíni, velryby, vydry a těsnění musí pravidelně povrchově dýchat vzduch.
Existuje více než 200 000 zdokumentovaných námořních druh s asi dvěma miliony mořských druhů, které ještě nebyly zdokumentovány.[4] Velikost mořských druhů se pohybuje od mikroskopických, včetně fytoplankton který může být tak malý jako 0,02 mikrometru, až obrovský kytovci (velryby, delfíni a sviňuchy), včetně modrá velryba - největší známé zvíře dosahující délky 33 metrů (108 stop).[5][6] Mořské mikroorganismy, počítaje v to protistů, bakterie a viry, byly různě odhadovány jako představující asi 70%[7] nebo asi 90%[8][1] z celkového počtu námořních biomasa.
Voda

Bez vody není život.[9] Byl popsán jako univerzální rozpouštědlo pro svou schopnost rozpustit mnoho látek,[10][11] a jako rozpouštědlo života.[12] Voda je jedinou běžnou látkou, která existuje jako pevný, kapalina a plyn za podmínek normálních pro život na Zemi.[13] The Nobelova cena vítěz Albert Szent-Györgyi odkazoval se na vodu jako matice a matice: matka a lůno života.[14]
Hojnost povrchové vody na Zemi je v oblasti Sluneční Soustava. Země hydrosféra Skládá se převážně z oceánů, ale technicky zahrnuje všechny vodní plochy na světě, včetně vnitrozemských moří, jezer, řek a podzemních vod až do hloubky 2 000 metrů (6 600 ft). Challenger Deep z Mariana příkop v Tichý oceán, s hloubkou 10 900 metrů (6,8 mil).[poznámka 1][15]
Obvykle je planeta rozdělena do pěti samostatných oceánů, ale všechny tyto oceány se spojují do jednoho světový oceán.[16] Hmotnost tohoto světového oceánu je 1,35×1018 metrické tuny, nebo asi 1/4400 z celkové hmotnosti Země. Světový oceán pokrývá oblast 3.618×108 km2 se střední hloubkou 3682 m, což má za následek odhadovaný objem 1.332×109 km3.[17] Pokud by byl celý povrch zemské kůry ve stejné výšce jako hladká koule, hloubka výsledného světového oceánu by byla asi 2,7 kilometru (1,7 mil).[18][19]
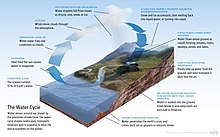
Asi 97,5% vody na Zemi je solný; zbývajících 2,5% je sladká voda. Většina čerstvé vody - asi 69% - je přítomna jako led ledové čepičky a ledovce.[20] Průměrná slanost oceánů Země je asi 35 gramů soli na kilogram mořské vody (3,5% soli).[21] Většina soli v oceánu pochází ze zvětrávání a eroze hornin na souši.[22] Některé soli se uvolňují z sopečná činnost nebo extrahované z chladu vyvřeliny.[23]
Oceány jsou také rezervoárem rozpuštěných atmosférických plynů, které jsou nezbytné pro přežití mnoha vodních forem života.[24] Mořská voda má významný vliv na světové klima, přičemž oceány působí jako velké zásobník tepla.[25] Posuny v rozložení oceánské teploty mohou způsobit významné posuny počasí, například El Niňo-jižní oscilace.[26]

Celkově oceán zabírá 71 procent povrchu světa,[2] v průměru téměř 3,7 km (2,3 mil) do hloubky.[27] Objemem oceán poskytuje asi 90 procent životního prostoru na planetě.[2] Spisovatel sci-fi Arthur C. Clarke poukázal na to, že by bylo vhodnější označovat planetu Zemi jako planetu oceán.[28][29]
Voda se však nachází jinde ve sluneční soustavě. Evropa, jeden z měsíců obíhajících kolem Jupiter, je o něco menší než Zemský měsíc. Existuje velká možnost, že pod jeho ledovou plochou existuje velký mořský oceán.[30] Odhaduje se, že vnější kůra pevného ledu je tlustá asi 10–30 km (6–19 mi) a kapalný oceán pod ním je hluboký asi 100 km.[31] Tím by se oceán Evropy dostal na dvojnásobek objemu oceánu Země. Spekulovalo se o oceánu Evropy mohl podporovat život,[32][33] a může být schopen podporovat mnohobuněčný systém mikroorganismy -li hydrotermální průduchy jsou aktivní na dně oceánu.[34] Enceladus, malý ledový měsíc Saturnu, má také to, co se zdá být podzemní oceán který aktivně odvádí teplou vodu z povrchu měsíce.[35]
Evoluční historie
The Země je stará asi 4,54 miliardy let.[36][37][38] Nejdříve nesporné důkazy o život na Zemi pochází z doby před nejméně 3,5 miliardami let,[39][40] Během Eoarchean Éra po geologii kůra začal tuhnout po dřívějším roztavení Hadean Eon. Mikrobiální rohož fosilie byly nalezeny u 3,48 miliard let staré pískovec v západní Austrálie.[41][42] Další časné fyzické důkazy o a biogenní látka je grafit ve stáří 3,7 miliardy let metasedimentární horniny objeveno v Západní Grónsko[43] stejně jako "pozůstatky biotický život „nalezený v 4,1 miliardách let starých skalách v západní Austrálii.[44][45] Podle jednoho z výzkumníků „Pokud na Zemi vznikl život relativně rychle… pak by to mohlo být běžné v… vesmír."[44]
Všechny organismy na Zemi pocházejí z a společný předek nebo předků genofond.[46][47] Předpokládá se, že vysoce energetická chemie před asi 4 miliardami let vyprodukovala samoreplikující se molekulu a o půl miliardy let později poslední společný předek všeho života existoval.[48] Současná vědecká shoda je, že složitá biochemie, která tvoří život, pochází z jednodušších chemických reakcí.[49] Začátek života mohl zahrnovat samoreplikující se molekuly, jako je RNA[50] a shromáždění jednoduchých buněk.[51] V roce 2016 vědci uvedli soubor 355 geny z poslední univerzální společný předek (LUCA) ze všech život, včetně mikroorganismů, žijících na Země.[52]
Současné druhy jsou stádiem procesu evoluce a svou rozmanitostí jsou produktem dlouhé řady speciačních a zánikových událostí.[53] Společný původ organismů byl nejprve odvozen ze čtyř jednoduchých faktů o organismech: Zaprvé mají geografické distribuce, které nelze vysvětlit lokální adaptací. Zadruhé, rozmanitost života není soubor zcela jedinečných organismů, ale organismy, které sdílejí morfologické podobnosti. Zatřetí, zakrnělé rysy bez jasného účelu připomínají funkční vlastnosti předků a nakonec lze organismy klasifikovat pomocí těchto podobností do hierarchie vnořených skupin - podobně jako v rodokmenu.[54] Moderní výzkum však naznačuje, že kvůli horizontální přenos genů, tento „strom života“ může být komplikovanější než jednoduchý rozvětvený strom, protože některé geny se šířily nezávisle mezi vzdáleně příbuznými druhy.[55][56]
Minulé druhy také zanechaly záznamy o své evoluční historii. Fosílie spolu se srovnávací anatomií dnešních organismů tvoří morfologický nebo anatomický záznam.[57] Porovnáním anatomií moderních i vyhynulých druhů mohou paleontologové odvodit linie těchto druhů. Tento přístup je však nejúspěšnější pro organismy, které měly tvrdé části těla, jako jsou mušle, kosti nebo zuby. Dále, protože prokaryoty, jako jsou bakterie a archea, sdílejí omezený soubor běžných morfologií, jejich fosílie neposkytují informace o jejich předcích.

V poslední době pocházejí důkazy o společném původu ze studia biochemických podobností mezi organismy. Například všechny živé buňky používají stejnou základní sadu nukleotidů a aminokyseliny.[59] Vývoj molekulární genetika odhalil záznam evoluce, která zůstala v genomech organismů: datování, když se druhy rozcházely skrz molekulární hodiny produkovaný mutacemi.[60] Například tato srovnání sekvencí DNA odhalila, že lidé a šimpanzi sdílejí 98% svých genomů a analýza několika oblastí, kde se liší, pomáhá osvětlit, kdy existoval společný předek těchto druhů.[61]
Prokaryoti obývali Zemi před přibližně 3–4 miliardami let.[62][63] Žádné zjevné změny v morfologie nebo v těchto organizmech došlo během příštích několika miliard let k buněčné organizaci.[64] Eukaryotické buňky se objevily před 1,6–2,7 miliardami let. Další velká změna v buněčné struktuře nastala, když byly bakterie pohlceny eukaryotickými buňkami v kooperativní asociaci zvané endosymbióza.[65][66] Pohltené bakterie a hostitelská buňka poté prošly koevolucí, kdy se bakterie vyvinuly buď do mitochondrií nebo hydrogenosomy.[67] Další pohltení sinice podobné organismy vedly k tvorbě chloroplastů v řasách a rostlinách.[68]

Historie života byla ta jednobuněčný eukaryoty, prokaryoty a archea až do doby před asi 610 miliony let, kdy se v oceánech začaly objevovat mnohobuněčné organismy Ediacaran doba.[62][69] The vývoj mnohobuněčnosti se vyskytly v několika nezávislých událostech, v organismech tak rozmanitých jako houby, hnědé řasy, sinice, slizové formy a myxobakterie.[70] V roce 2016 vědci uvedli, že asi před 800 miliony let mohla malá genetická změna v jedné molekule zvané GK-PID umožnit organizmům přejít z jednoho buněčného organismu do jedné z mnoha buněk.[71]
Brzy po vzniku těchto prvních mnohobuněčných organismů se v rozpětí asi 10 milionů let objevilo pozoruhodné množství biologické rozmanitosti, v případě zvaném Kambrijská exploze. Tady většina typy ve fosilních záznamech se objevily moderní zvířata a také jedinečné linie, které následně vyhynuly.[72] Byly navrženy různé spouštěče kambrijské exploze, včetně akumulace kyslík v atmosféra z fotosyntézy.[73]
Asi před 500 miliony let začaly kolonizovat půdu rostliny a houby. Důkazy o výskytu prvních suchozemských rostlin se vyskytují v Ordovik, kolem 450 před miliony let, ve formě fosilních spór.[74] Pozemní rostliny se začaly diverzifikovat v Pozdní silur, z okolí 430 před miliony let.[75] Po kolonizaci půdy rostlinami brzy následovala členovci a další zvířata.[76] Hmyz byly obzvláště úspěšné a dokonce i dnes tvoří většinu živočišných druhů.[77] Obojživelníci se poprvé objevily před 364 miliony let, poté brzy amniotes a ptactvo asi před 155 miliony let (oba z „plaz "podobné linie", savci asi před 129 miliony let, homininae asi před 10 miliony let a moderní lidé asi před 250 000 lety.[78][79][80] Navzdory vývoji těchto velkých zvířat jsou však menší organismy podobné typům, které se vyvinuly na začátku tohoto procesu, stále velmi úspěšné a dominují Zemi, přičemž většina biomasy i druhů jsou prokaryoty.[81]
Odhady počtu zemského proudu druh rozmezí od 10 milionů do 14 milionů,[82] z nichž je zdokumentováno asi 1,2 milionu a více než 86 procent dosud nebylo popsáno.[83]
Mikroorganismy


Mikroorganismy tvoří asi 70% mořská biomasa.[7] A mikroorganismus, nebo mikrob, je mikroskopický organismus příliš malý na to, aby ho bylo možné poznat pouhým okem. To může být jednobuněčný[84] nebo mnohobuňečný. Mikroorganismy jsou rozmanité a zahrnují všechny bakterie a archaea, většina prvoky jako řasy, houby a některá mikroskopická zvířata jako např vířníky.
Mnoho makroskopické zvířata a rostliny mít mikroskopické juvenilní fáze. Někteří mikrobiologové také klasifikují viry (a viroidy ) jako mikroorganismy, ale ostatní je považují za neživé.[85][86]
Mikroorganismy mají zásadní význam pro recyklaci živin ekosystémy jak se chovají rozkladače. Některé mikroorganismy jsou patogenní, způsobující nemoci a dokonce smrt u rostlin a zvířat.[87] Jako obyvatelé největšího prostředí na Zemi řídí mikrobiální námořní systémy změny v každém globálním systému. Mikroby jsou zodpovědné za prakticky všechno fotosyntéza která se vyskytuje v oceánu, stejně jako jízda na kole uhlík, dusík, fosfor, jiný živiny a stopové prvky.[88]

| Mořské mikroorganismy |
| ||||||||||||||||||||||||


Mikroskopický podmořský život je různorodý a stále špatně pochopený, například pokud jde o roli viry v mořských ekosystémech.[89] Většina mořských virů je bakteriofágy, které jsou neškodné pro rostliny a živočichy, ale jsou nezbytné pro regulaci slaných a sladkovodních ekosystémů.[90] Infikují a ničí bakterie ve vodních mikrobiálních komunitách a jsou nejdůležitějším mechanismem recyklace uhlíku v mořském prostředí. Organické molekuly uvolněné z mrtvých bakteriálních buněk stimulují růst čerstvých bakterií a řas.[91] Virová aktivita může také přispět k biologické čerpadlo proces, kterým uhlík je oddělený v hlubokém oceánu.[92]


Proud vzdušných mikroorganismů obíhá planetu nad systémy počasí, ale pod komerčními vzdušnými pruhy.[93] Některé peripatetické mikroorganismy jsou smeteny ze suchozemských prachových bouří, ale většina pochází z mořských mikroorganismů v mořský sprej. V roce 2018 vědci uvedli, že na každý metr čtvereční kolem planety se denně ukládají stovky milionů virů a desítky milionů bakterií.[94][95]
Mikroskopické organismy žijí po celém světě biosféra. Hmotnost prokaryot mikroorganismy - což zahrnuje bakterie a archea, ale nikoliv nukleované eukaryotické mikroorganismy - může to být až 0,8 bilionu tun uhlíku (z celkové biosféry Hmotnost, odhaduje se na 1 až 4 biliony tun).[96] Jednobuněčný barofilní mořské mikroby byly nalezeny v hloubce 10 900 m (35 800 ft) v Mariana příkop, nejhlubší místo v zemských oceánech.[97][98] Mikroorganismy žijí uvnitř skal 580 m (1900 ft) pod mořským dnem pod 2590 m (8,500 ft) oceánu u pobřeží severozápadní Spojené státy,[97][99] stejně jako 2400 m (7 900 ft; 1,5 mi) pod mořským dnem mimo Japonsko.[100] Největší známá teplota, při které může existovat mikrobiální život, je 122 ° C (252 ° F) (Methanopyrus kandleri ).[101] V roce 2014 vědci potvrdili existenci mikroorganismů žijících 800 m (2,600 ft) pod ledem Antarktida.[102][103] Podle jednoho výzkumníka „Mikroby najdete všude - jsou extrémně přizpůsobivé podmínkám a přežijí, ať jsou kdekoli.“[97]
Mořské viry
Viry jsou malé infekční agens které nemají vlastní metabolismus a může replikovat jen uvnitř živých buňky dalších organismy.[104] Viry mohou infikovat všechny typy formy života, z zvířata a rostliny na mikroorganismy, počítaje v to bakterie a archaea.[105] Lineární velikost průměrného viru je přibližně jedna stotina průměru bakterie. Většinu virů nelze vidět pomocí optický mikroskop tak elektronové mikroskopy místo toho se používají.[106]
Viry se vyskytují všude tam, kde je život, a pravděpodobně existovaly od doby, kdy se nejprve vyvinuly živé buňky.[107] Původ virů je nejasný, protože netvoří fosilie, takže molekulární techniky byly použity k porovnání DNA nebo RNA virů a jsou užitečným prostředkem ke zkoumání toho, jak vznikají.[108]
Viry jsou nyní považovány za starodávné a mají původ, který předcházel divergenci života v tři domény.[109] Ale původ virů v evoluční historie života jsou nejasné: některé mohou mít vyvinul z plazmidy - kousky DNA, které se mohou pohybovat mezi buňkami - zatímco jiné se mohly vyvinout z bakterií. V evoluci jsou viry důležitým prostředkem horizontální přenos genů, což se zvyšuje genetická rozmanitost.[110]



Názory na to, zda jsou viry formou, se liší život nebo organické struktury, které interagují s živými organismy.[111] Někteří je považují za formu života, protože nesou genetický materiál, množí se vytvářením více kopií sebe sama prostřednictvím sebezhromáždění a vyvíjejí se skrz přírodní výběr. Chybí jim však klíčové vlastnosti, jako je buněčná struktura, která se obecně považuje za nezbytnou k tomu, aby se mohla považovat za život. Protože mají některé, ale ne všechny takové vlastnosti, byly viry popsány jako replikátory[112] a jako „organismy na pokraji života“.[113]

Bakteriofágy, často právě volal fágy, jsou viry, které parazit bakterie a archaea. Mořské fágy parazity mořských bakterií a archea, jako např sinice.[114] Jsou běžnou a různorodou skupinou virů a jsou nejhojnější biologickou entitou v mořském prostředí, protože jejich hostitelé, bakterie, jsou obvykle numericky dominantním buněčným životem v moři. Obecně je v každém ml mořské vody asi 1 až 10 milionů virů nebo asi desetkrát více dvouvláknových DNA virů, než jsou buněčné organismy,[115][116] ačkoli odhady množství virů v mořské vodě se mohou v širokém rozmezí lišit.[117][118] Ocasní bakteriofágy Zdá se, že dominují mořským ekosystémům v počtu a rozmanitosti organismů.[114] Bakteriofágy patřící do rodin Corticoviridae,[119] Inoviridae[120] a Microviridae[121] je také známo, že infikují různé mořské bakterie.
Mikroorganismy tvoří přibližně 70% mořské biomasy.[7] Odhaduje se, že viry každý den zabijí 20% této biomasy a že v oceánech je 15krát více virů než bakterií a archea. Viry jsou hlavními činiteli odpovědnými za rychlé ničení škodlivých látek květy řas,[122] které často zabíjejí další mořský život.[123]Počet virů v oceánech klesá dále na moři a hlouběji do vody, kde je méně hostitelských organismů.[92]
Existují také archaeanské viry, které se replikují uvnitř archaea: jedná se o dvouvláknové DNA viry neobvyklých a někdy jedinečných tvarů.[124][125] Tyto viry byly podrobně studovány v dokumentu termofilní archaea, zejména objednávky Sulfolobales a Termoprotealy.[126]
Viry jsou důležitým přírodním prostředkem přenos genů mezi různými druhy, což se zvyšuje genetická rozmanitost a řídí evoluci.[110] Předpokládá se, že viry hrály ústřední roli v časném vývoji, před diverzifikací bakterií, archea a eukaryot, v době poslední univerzální společný předek života na Zemi.[127] Viry jsou stále jedním z největších rezervoárů neprozkoumané genetické rozmanitosti na Zemi.[92]
Mořské bakterie


Bakterie tvoří velkou doména z prokaryotický mikroorganismy. Typicky několik mikrometry na délku mají bakterie řadu tvarů, od koulí po tyče a spirály. Bakterie byly mezi prvními formami života, které se objevily Země, a jsou přítomny ve většině z nich stanoviště. Bakterie obývají půdu, vodu, kyselé horké prameny, radioaktivní odpad,[128] a hluboké části zemská kůra. Bakterie také žijí symbiotický a parazitický vztahy s rostlinami a zvířaty.
Kdysi považováno za rostliny tvořící třídu Schizomycetes, bakterie jsou nyní klasifikovány jako prokaryoty. Na rozdíl od buněk zvířat a dalších eukaryoty, bakteriální buňky neobsahují a jádro a zřídka přístav vázané na membránu organely. Ačkoli termín bakterie tradičně zahrnoval všechny prokaryoty, vědecká klasifikace se změnilo po objevu v 90. letech, že prokaryoty sestávají ze dvou velmi odlišných skupin organismů vyvinul od starodávného společného předka. Tyto evoluční domény jsou nazývány Bakterie a Archaea.[129]
Předchůdci moderních bakterií byli jednobuněčné mikroorganismy, které byly první formy života se objeví na Zemi asi před 4 miliardami let. Asi 3 miliardy let byla většina organismů mikroskopická a bakterie a archea byly dominantní formou života.[130][131] Ačkoli bakteriální fosilie existují, jako např stromatolity, jejich nedostatek rozlišovací způsobilosti morfologie brání tomu, aby byly použity k prozkoumání historie bakteriálního vývoje nebo k datu času vzniku konkrétního bakteriálního druhu. Genové sekvence však mohou být použity k rekonstrukci bakterie fylogeneze a tyto studie naznačují, že bakterie se nejprve lišily od linie archaeal / eukaryotic.[132] Bakterie se také účastnily druhé velké evoluční divergence, archaea a eukaryot. Zde eukaryota vyústila do vstupu starodávných bakterií do endosymbiotikum sdružení s předky eukaryotických buněk, které samy o sobě pravděpodobně souvisely s Archaea.[66][133] To zahrnovalo pohlcení proto-eukaryotickými buňkami alfaproteobakteriální symbionty mitochondrie nebo hydrogenosomy, které se stále nacházejí ve všech známých Eukarya. Později některé eukaryoty, které již obsahovaly mitochondrie, také pohltily organismy podobné sinicím. To vedlo k vytvoření chloroplasty v řasách a rostlinách. Existují také některé řasy, které pocházejí z ještě pozdějších endosymbiotických událostí. Zde eukaryota pohltila eukaryotické řasy, které se vyvinuly v plastid „druhé generace“.[134][135] Toto je známé jako sekundární endosymbióza.

Mariňák Thiomargarita namibiensis, největší známá bakterie

Sinice květy může obsahovat smrtící cyanotoxiny.

The chloroplasty z glaukofyty mít peptidoglykan vrstva, důkazy naznačující jejich endosymbiotikum původ z sinice.[136]

Bakterie mohou být prospěšné. Tento Pompejský červ, an extremophile najdete pouze na hydrotermální průduchy, má ochranný kryt bakterií.




Největší známá bakterie, mořská Thiomargarita namibiensis, mohou být viditelné pouhým okem a někdy dosahují 0,75 mm (750 μm).[137][138]
Mořské archaea

The archaea (Řek pro starověký[140]) tvoří a doména a království z jednobuněčný mikroorganismy. Tyto mikroby jsou prokaryoty, což znamená, že nemají buněčné jádro nebo jakýkoli jiný vázaný na membránu organely ve svých celách.
Archaea byly původně klasifikovány jako bakterie, ale tato klasifikace je zastaralá.[141] Buňky archaealu mají jedinečné vlastnosti oddělující je od ostatních dvou domén života, Bakterie a Eukaryota. Archaea se dále dělí na několik uznaných phyla. Klasifikace je obtížná, protože většina nebyla izolována v laboratoři a byla detekována pouze jejich analýzou nukleové kyseliny ve vzorcích z jejich prostředí.
Archea a bakterie mají obecně podobnou velikost a tvar, i když několik archaeí má velmi podivné tvary, například ploché a čtvercové buňky Haloquadratum walsbyi.[142] Přes tuto morfologickou podobnost s bakteriemi mají archaea geny a několik metabolické cesty které jsou více příbuzné těm z eukaryot, zejména enzymy zahrnutý do něčeho, zůčastnit se čeho transkripce a překlad. Jiné aspekty archaeální biochemie jsou jedinečné, například jejich spoléhání se na etherové lipidy v jejich buněčné membrány, jako archaeols. Archaea využívá více zdrojů energie než eukaryota: ty se pohybují od organické sloučeniny, jako jsou cukry, do amoniak, ionty kovů nebo dokonce vodíkový plyn. Sůl-tolerantní archaea (dále jen Haloarchaea ) používají sluneční světlo jako zdroj energie a další druhy archea opravit uhlík; na rozdíl od rostlin a sinice, žádný známý druh archaea nedělá obojí. Archaea rozmnožovat nepohlavně podle binární dělení, fragmentace nebo pučící; na rozdíl od bakterií a eukaryot se netvoří žádné známé druhy výtrusy.
Archaea jsou zvláště četné v oceánech a archaea v plankton může být jednou z nejhojnějších skupin organismů na planetě. Archaea jsou hlavní součástí života Země a mohou hrát roli v obou uhlíkový cyklus a dusíkový cyklus.

Halobakterie, které se nacházejí ve vodě téměř nasycené solí, jsou nyní uznávány jako archaea.

Ploché buňky archaea ve tvaru čtverce Haloquadratum walsbyi

Methanosarcina barkeri, mořská archaea, která produkuje metan

Termofily, jako Pyrolobus fumarii přežít dobře přes 100 ° C.

Kresba jiného mořského termofila, Pyrococcus furiosus




Námořní protisti
Protisti jsou eukaryoty, které nelze klasifikovat jako rostliny, houby nebo zvířata. Obvykle jsou jednobuněčné a mikroskopické. Život vznikl jako jednobuněčné prokaryoty (bakterie a archea) a později se z nich vyvinula složitější eukaryoty. Eukaryoty jsou rozvinutější formy života známé jako rostliny, zvířata, houby a prvoci. Termín protist se historicky začaly používat jako výhodný termín pro eukaryoty, které nelze přísně klasifikovat jako rostliny, zvířata nebo houby. Nejsou součástí moderní kladistiky, protože jsou paraphyletic (chybí společný předek). Protisty lze rozdělit do čtyř skupin podle toho, zda je jejich výživa podobná rostlinám, zvířatům, houbám,[143] nebo jejich směs.[144]
Protisti podle toho, jak dostávají jídlo | |||||||
|---|---|---|---|---|---|---|---|
| Typ protisty | Popis | Příklad | Další příklady | ||||
| Rostlinný | Autotrofní protisti, kteří si vyrábějí vlastní jídlo, aniž by museli konzumovat jiné organismy, obvykle pomocí fotosyntézy |  | Červené řasy, Cyanidium sp. | Zelené řasy, hnědé řasy, rozsivky a nějaký dinoflageláty. Rostlinní protisté jsou důležitými složkami fytoplanktonu diskutováno níže. | |||
| Jako zvíře | Heterotrofní protisti, kteří dostávají potravu konzumující jiné organismy |  | Radiolarian protist, jak je tažen Haeckel | Foraminiferans a některé námořní améby, náčelníci a bičíky. | |||
| Houba | Saprotrofní protisti, kteří získávají potravu ze zbytků organismů, které se rozpadly a rozpadly |  | Marine slizové sítě tvoří labyrintové sítě trubic, ve kterých mohou cestovat améby bez pseudopodů | Mořské lišejníky | |||
| Mixotropy | Rozličný | Mixotrofní a osmotrofický protisti, kteří získávají jídlo kombinací výše uvedeného |  | Euglena mutabilis, fotosyntetický bičíkatý | Mnoho mořských mixotropů se nachází mezi protisty, včetně mezi nálevníky, Rhizaria a dinoflageláty [145] | ||


Protisti jsou velmi různorodé organismy, které jsou v současné době organizovány do 18 kmenů, ale není snadné je zařadit.[147][148] Studie prokázaly, že v oceánech, hlubinných průduchech a říčních sedimentech existuje vysoká protistátní rozmanitost, což naznačuje, že ještě není nutné objevit velké množství eukaryotických mikrobiálních komunit.[149][150] Došlo k malému výzkumu mixotrofní protistům, ale nedávné studie v mořském prostředí zjistily, že mixotrofní protesty přispívají významnou částí protistů biomasa.[145]
- Jednobuněčné a mikroskopické protisty

Diatomy mají sklo podobné buněčným stěnám oxid křemičitý a zavolal frustule.[152]

Fosilní křemelina z 32-40 mya

Jednobuněčná řasa, Gephyrocapsa oceanica

Dva dinoflageláty

Zooxanthellae je fotosyntetická řasa, která žije uvnitř hostitelů korál.

Jednobuněčný ciliate se zelenou zoochlorellae žijící uvnitř endosymbioticky.










Na rozdíl od buněk prokaryot jsou buňky eukaryot vysoce organizované. Rostliny, zvířata a houby jsou obvykle mnohobuněčný a jsou typicky makroskopické. Většina protistů je jednobuněčná a mikroskopická. Existují ale výjimky. Někteří jednobuněční mořští protisti jsou makroskopičtí. Některé mořské slizové formy mají jedinečné životní cykly, které zahrnují přepínání mezi jednobuněčnými, koloniální a mnohobuněčné formy.[153] Jiní mořští protisti nejsou ani jednobuněční, ani mikroskopičtí, jako např Mořská řasa.
- Makroskopické protisty (viz také jednobuněčné makrořasy → )

Jednobuněčný obří améba má až 1000 jádra a dosahuje délky 5 mm.

Gromia sphaerica je velká sférická teste améba což vede k blátě. Jeho průměr je až 3,8 cm.[154]

Spiculosiphon oceana jednobuněčný foraminiferan se vzhledem a životním stylem, který napodobuje a mycí houba, dorůstá do délky 5 cm.

The xenofyor, žije další jednobuněčný foraminiferan hlubinné zóny. Má obrovskou skořápku o průměru až 20 cm.[155]

Obří řasa, a hnědé řasy, není skutečná rostlina, přesto je mnohobuněčná a může dorůst až 50 metrů.





Protisti byli popsáni jako taxonomická brašna, kde se všechno, co se nevejde do jedné z hlavních biologická království Lze umístit.[156] Někteří moderní autoři upřednostňují vyloučit mnohobuněčné organismy z tradiční definice protisty a omezují protisty na jednobuněčné organismy.[157][158] Tato omezenější definice vylučuje mořské řasy a slizové formy.[159]
Mořští živočichové
Jako mladiství se zvířata vyvíjejí z mikroskopických stádií, která mohou zahrnovat výtrusy, vejce a larvy. Alespoň jedna skupina mikroskopických zvířat, parazitický cnidarian Myxozoa, je jednobuněčný ve své dospělé formě a zahrnuje mořské druhy. Ostatní dospělí mořští mikroorganismy jsou mnohobuněčné. Mikroskopický dospělý členovci se běžněji vyskytují ve vnitrozemí ve sladké vodě, ale existují i mořské druhy. Mikroskopický dospělý námořní korýši zahrnout některé copepods, perloočka a tardigrades (vodní medvědi). Nějaké námořní hlístice a vířníky jsou také příliš malé na to, aby je bylo možné rozpoznat pouhým okem, stejně jako mnoho jiných loricifera, včetně nedávno objevených anaerobní druhy, které tráví život v anoxický životní prostředí.[160][161] Copepods více přispívají k sekundární produktivita a uhlíkový dřez světových oceánů než jakákoli jiná skupina organismů.
- Mořští živočichové

Darkfield fotka a gastrotrich, červ podobné zvíře žijící mezi částicemi sedimentu

Obrněný Pliciloricus enigmaticus, asi 0,2 mm dlouhé, žijí v prostorech mezi mořským štěrkem.

Nákres a tardigrade (vodní medvěd) na zrnku písku

Vířníky, obvykle 0,1–0,5 mm dlouhé, mohou vypadat jako protisti, ale mají mnoho buněk a patří do skupiny Animalia.




Houby


Více než 1 500 druhů houby jsou známy z mořského prostředí.[162] Jsou parazitující na mořské řasy nebo zvířata, nebo jsou saproby krmení mrtvými organickými látkami z řas, korálů, cyst prvoků, mořských trav, dřeva a jiných substrátů.[163] Výtrusy mnoha druhů mají speciální přídavky, které usnadňují připojení k substrátu.[164] Mořské houby lze nalézt také v mořská pěna a kolem hydrotermální oblasti oceánu.[165] Různorodá škála neobvyklých sekundárních metabolity je produkován mořskými houbami.[166]
Mykoplankton jsou saprotropní členové plankton společenství námořní a sladkovodní ekosystémy.[167][168] Skládají se z vláknitý volně žijící houby a kvasinky spojené s planktonovými částicemi nebo fytoplankton.[169] Podobný bakterioplankton, tyto vodní houby hrají významnou roli v heterotrofní mineralizace a koloběh živin.[170] Mykoplankton může mít průměr až 20 mm a délku přes 50 mm.[171]
Typický mililiter mořské vody obsahuje asi 103 do 104 buňky hub.[172] Toto číslo je větší v pobřežních ekosystémech a ústí řek kvůli nutričnímu odtoku ze suchozemských komunit. Vyšší rozmanitost mykoplanktonu se vyskytuje kolem pobřeží a v povrchových vodách až do 1000 metrů, s a svislý profil to záleží na tom, jak hojné fytoplankton je.[173][174] Tento profil se mezi sezónami mění kvůli změnám v dostupnosti živin.[175] Mořské houby přežívají v prostředí s konstantním nedostatkem kyslíku, a proto závisí na difúzi kyslíku turbulence a kyslík generovaný fotosyntetické organismy.[176]
Mořské houby lze klasifikovat jako:[176]
- Nižší houby - přizpůsobené mořským stanovištím (zoosporický houby, včetně mastigomycetů: oomycety a chytridiomycetes )
- Vyšší houby - vláknité, upravené na planktonický životní styl (hyphomycetes, ascomycetes, bazidiomycety ). Většina druhů mykoplanktonu jsou vyšší houby.[173]
Lišejníky jsou mutualistic asociace mezi houbou, obvykle an ascomycete a řasa nebo a sinice. Několik lišejníků se vyskytuje v mořském prostředí.[177] Mnoho dalších se vyskytuje v stříkající zóna, kde zaujímají různé vertikální zóny podle toho, jak tolerantní jsou vůči ponoření.[178] Některé lišejníky žijí dlouho; jeden druh byl datován na 8 600 let.[179] Jejich životnost je však obtížně měřitelná, protože to, co definuje stejný lišejník, není přesné.[180] Lišejníky rostou vegetativním odlomením kusu, který může nebo nemusí být definován jako stejný lišejník, a mohou se sloučit dva lišejníky různého věku, což vyvolává otázku, zda se jedná o stejný lišejník.[180]
The mořský šnek Littoraria irrorata poškozuje rostliny Spartina v mořských bažinách, kde žije, což umožňuje kolonizaci rostliny spory přílivových ascomycetous hub. Hlemýžď pak sžírá růst hub přednostně před samotnou trávou.[181]
Podle fosilních záznamů se houby datují pozdě Proterozoikum éra před 900-570 miliony let. V Číně byly objeveny fosilní mořské lišejníky staré 600 milionů let.[182] Předpokládá se, že mykoplankton se vyvinul z suchozemských hub, pravděpodobně v Paleozoikum éry (před 390 miliony let).[183]
Původ zvířat

Nejdříve zvířata byli mořští bezobratlých, to znamená, obratlovců přišel později. Zvířata jsou mnohobuňečný eukaryoty,[poznámka 2] a liší se od rostlin, řas a hub tím, že chybí buněčné stěny.[184] Mořští bezobratlí jsou zvířata, která obývají a námořní prostředí kromě členů obratlovců strunat kmen; bezobratlým chybí a páteř. Některé se vyvinuly a skořápka nebo tvrdý exoskeleton.
Nejstarší fosilie zvířat mohou patřit do rodu Dickinsonia,[185] Před 571 miliony až 541 miliony let.[186] Individuální Dickinsonia typicky připomínají bilaterálně symetrický žebrovaný ovál. Stále rostly, dokud nebyly pokryty sedimenty nebo jinak zabity,[187] a strávili většinu svého života s těly pevně ukotvenými v sedimentu.[188] Jejich taxonomické spříznění jsou v současné době neznámé, ale jejich způsob růstu je shodný s a bilaterální afinita.[189]
Na rozdíl od Dickinsonia, nejdříve široce přijímané zvířecí fosilie jsou poměrně moderní cnidarians (skupina, která zahrnuje korál, Medúza, mořské sasanky a Hydra ), možná z okolí 580 Ma[190] The Ediacara biota, který vzkvétal posledních 40 milionů let před začátkem EU Kambrijský,[191] byli prvními zvířaty o délce více než jen pár centimetrů. Jako Dickinsonia, mnoho z nich bylo plochých s „prošívaným“ vzhledem a vypadalo to tak divně, že existoval návrh klasifikovat je jako samostatné království, Vendozoa.[192] Jiné však byly interpretovány již dříve měkkýši (Kimberella[193][194]), ostnokožci (Arkarua[195]), a členovci (Spriggina,[196] Parvancorina[197]). O klasifikaci těchto vzorků stále existuje debata, zejména proto, že diagnostické vlastnosti, které umožňují taxonomům klasifikovat novější organismy, například podobnosti s živými organismy, v Ediacarans obecně chybí. Zdá se však, že o tom není pochyb Kimberella byl alespoň a triploblastický bilaterální zvíře, jinými slovy, zvíře podstatně složitější než cnidariáni.[198]
Malá šupinatá fauna jsou velmi smíšenou sbírkou fosilií nalezených mezi pozdním Ediacaranem a Střední kambrie období. Nejdříve, Cloudina, vykazuje známky úspěšné obrany proti predaci a může označovat začátek evoluční závod ve zbrojení. Některé drobné rané kambrické mušle téměř jistě patřily měkkýšům, zatímco majitelé některých „pancéřových desek“ Halkieria a Microdictyon, byly nakonec identifikovány, když byly v kambriu nalezeny úplnější vzorky lagerstätten která zachovala zvířata s měkkým tělem.[199]
Plány těla a phyla
Bezobratlí jsou seskupeni do různých phyla. Neformální kmen lze považovat za způsob seskupování organismů podle jejich tělesný plán.[200][201]:33 Plán těla odkazuje na plán, který popisuje tvar nebo morfologie organismu, jako je jeho symetrie, segmentace a jeho dispozice přílohy. Myšlenka tělesných plánů vznikla obratlovců, které byly seskupeny do jednoho kmene. Plán těla obratlovců je však pouze jedním z mnoha a bezobratlí se skládají z mnoha plánů phyly nebo těla. Na historii objevování tělních plánů lze pohlížet jako na pohyb od světonázoru zaměřeného na obratlovce, až po vidění obratlovců jako jednoho tělesného plánu mezi mnoha. Mezi průkopníky zoologové „Linné identifikoval dva plány těla mimo obratlovce; Cuvier identifikoval tři; a Haeckel měl čtyři, stejně jako Protista s osmi dalšími, celkem dvanáct. Pro srovnání, počet kmenů uznaných moderními zoology je stoupl na 35.[201]

Historicky byly plány těla považovány za plány, které se během roku rychle vyvinuly Kambrijská exploze,[205] ale jemnější chápání evoluce zvířat naznačuje postupný vývoj tělesných plánů v raných fázích Paleozoikum a za.[206] Obecněji lze kmen definovat dvěma způsoby: jak je popsáno výše, jako skupina organismů s určitým stupněm morfologické nebo vývojové podobnosti ( fenetický definice), nebo skupina organismů s určitým stupněm evoluční příbuznosti (dále jen fylogenetické definice).[206]
V 70. letech se již vedla debata o tom, zda vznik moderní fyly byl „výbušný“ nebo postupný, ale skrytý nedostatkem Precambrian fosilie zvířat.[199] Opětovná analýza fosilií z Burgess Shale lagerstätte zvýšil zájem o tuto problematiku, když odhalila zvířata, jako např Opabinie, který se nehodil do žádného známého kmen. V té době to byly interpretovány jako důkaz, že moderní phyla se velmi rychle vyvinula v kambrijské explozi a že „divné divy“ Burgess Shale ukázaly, že raná kambrie byla jedinečným experimentálním obdobím vývoje zvířat.[207] Pozdější objevy podobných zvířat a vývoj nových teoretických přístupů vedly k závěru, že mnoho „divných divů“ byly evoluční „tety“ nebo „bratranci“ moderních skupin[208]—Například to Opabinie byl členem lobopody, což je skupina, která zahrnuje předky členovců a která mohla úzce souviset s moderní dobou tardigrades.[209] Přesto stále existuje spousta debat o tom, zda kambrijská exploze byla skutečně výbušná, a pokud ano, jak a proč k ní došlo a proč se jeví jako jedinečná v historii zvířat.[210]
Bazální bezobratlí zvířata
Nejvíc bazální animal phyla, the animals that evolved first, are the Porifera, Ctenophora, Placozoa a Cnidaria. None of these basal body plans exhibit bilaterální symetrie.
| Animalia |
| |||||||||||||||||||||||||||
| 760 mya |


Mořské houby

Houby are animals of the kmen Porifera (from Modern Latin for bearing pores[214]). They are multicellular organisms that have bodies full of pores and channels allowing water to circulate through them, consisting of jelly-like mesohyl vložené mezi dvě tenké vrstvy buňky. They have unspecialized cells that can transformovat do jiných typů a které v tomto procesu často migrují mezi hlavními buněčnými vrstvami a mezohylem. Houby nemají nervový, zažívací nebo oběhové systémy. Místo toho se většina spoléhá na udržování stálého průtoku vody tělem, aby získala jídlo a kyslík a odstranila odpady.
Sponges are similar to other animals in that they are mnohobuňečný, heterotrofní, nedostatek buněčné stěny a vyrábět spermie. Unlike other animals, they lack true papírové kapesníky a orgány, and have no symetrie těla. The shapes of their bodies are adapted for maximal efficiency of water flow through the central cavity, where it deposits nutrients, and leaves through a hole called the osculum. Many sponges have internal skeletons of spongin and/or spicules of uhličitan vápenatý nebo oxid křemičitý. All sponges are sessile aquatic animals. Although there are freshwater species, the great majority are marine (salt water) species, ranging from tidal zones to depths exceeding 8,800 m (5.5 mi). Some sponges live to great ages; there is evidence of the deep-sea glass sponge Monorhaphis chuni living about 11,000 years.[215][216]
While most of the approximately 5,000–10,000 known species feed on bakterie and other food particles in the water, some host fotosyntéza micro-organisms as endosymbionty and these alliances often produce more food and oxygen than they consume. A few species of sponge that live in food-poor environments have become masožravci that prey mainly on small korýši.[217]

Sponge biodiversity. There are four sponge species in this photo.

Venus' flower basket at a depth of 2572 meters

The long-living Monorhaphis chuni





Linné mistakenly identified sponges as plants in the order Řasy.[218] For a long time thereafter sponges were assigned to a separate subkingdom, Parazoa (význam beside the animals).[219] They are now classified as a paraphyletic kmen from which the higher animals have evolved.[220]
Ctenophores

Ctenophores (from Greek for carrying a comb), commonly known as comb jellies, are a phylum that live worldwide in marine waters. They are the largest non-colonial animals to swim with the help of řasy (hairs or combs).[221] Coastal species need to be tough enough to withstand waves and swirling sediment, but some oceanic species are so fragile and transparent that it is very difficult to capture them intact for study.[222] In the past ctenophores were thought to have only a modest presence in the ocean, but it is now known they are often significant and even dominant parts of the planktonic biomass.[223]:269
The phylum has about 150 known species with a wide range of body forms. Sizes range from a few milimetry to 1.5 m (4 ft 11 in). Cydippids are egg-shaped with their cilia arranged in eight radial comb rows, and deploy retractable tentacles for capturing prey. The benthic platyctenidy are generally combless and flat. Pobřežní beroidy have gaping mouths and lack tentacles. Most adult ctenophores prey on microscopic larvae and vířníky a malé korýši but beroids prey on other ctenophores.

Světlo difrakční along the comb rows of a cydippid, left tentacle deployed, right retracted

Deep-sea ctenophore trailing tentacles studded with tentilla (sub-tentacles)



Egg-shaped cydippid ctenofor

Group of small benthic creeping comb jellies streaming tentacles and living symbioticky on a starfish.

Lobata sp. with paired thick lobes

The sea walnut has a transient anus which forms only when it needs to defecate.[224]




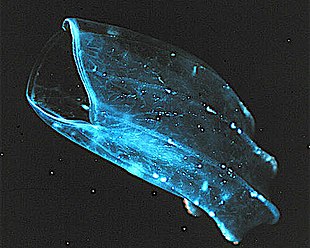
První autoři kombinovali ctenofory s cnidarians. Ctenophores resemble cnidarians in relying on water flow through the body cavity for both digestion and respiration, as well as in having a decentralized nervová síť rather than a brain. Also like cnidarians, the bodies of ctenophores consist of a mass of jelly, with one layer of buňky na vnější straně a další obložení vnitřní dutiny. V ctenoforech jsou však tyto vrstvy hluboké do dvou buněk, zatímco u cnidariánů jsou hluboké pouze do jedné buňky. While cnidarians exhibit radiální symetrie, ctenophores have two anal canals which exhibit biradial symmetry (half-turn rotational symmetry).[225] Postavení ctenoforů v evolučním rodokmenu zvířat je dlouho diskutováno a většinový pohled v současné době, založený na molekulární fylogenetika, je to cnidarians a bilaterians jsou si navzájem bližší, než je tomu u ctenoforů.[223]:222
Placozoa
Placozoa (from Greek for flat animals) have the simplest structure of all animals. Jsou to bazální form of free-living (non-parasitic) mnohobuněčný organismus[226] that do not yet have a common name.[227] They form a phylum containing sofar only three described species, of which the first, the classical Trichoplax adhaerens, was discovered in 1883.[228] Two more species have been discovered since 2017,[229][230] and genetic methods indicate this phylum has a further 100 to 200 nepopsané druhy.[231]

Trichoplax is a small, flattened, animal about one mm across and usually about 25 µm thick. Jako améby they superficially resemble, they continually change their external shape. In addition, spherical phases occasionally form which may facilitate movement. Trichoplax lacks tissues and organs. There is no manifest body symmetry, so it is not possible to distinguish anterior from posterior or left from right. It is made up of a few thousand cells of six types in three distinct layers.[232] The outer layer of simple epitelové buňky medvěd řasy which the animal uses to help it creep along the seafloor.[233] Trichoplax feed by engulfing and absorbing food particles – mainly microbes and organic detritus – with their underside.
Mořští cnidarians

Cnidarians (from Greek for kopřiva) are distinguished by the presence of bodavé buňky, specializované buňky, které používají hlavně k zachycení kořisti. Cnidarians include korály, mořské sasanky, Medúza a hydrozoans. Tvoří a kmen containing over 10,000[234] druh z zvířata found exclusively in aquatic (mainly marine) environments. Jejich těla se skládají z mezoglea, neživá rosolovitá látka, vložená mezi dvě vrstvy epitel které jsou většinou jedno cell thick. They have two basic body forms: swimming medusae a sessile polypy, z nichž oba jsou radiálně symetrické with mouths surrounded by tentacles that bear cnidocytes. Obě formy mají jednu otvor and body cavity that are used for digestion and dýchání.
Fosilní cnidarians byly nalezeny ve skalách vytvořených kolem 580 před miliony let. Fossils of cnidarians that do not build mineralizovaný structures are rare. Scientists currently think cnidarians, ctenofory a bilaterians jsou užší příbuzní vápenaté houby than these are to other houby, a to anthozoans are the evolutionary "aunts" or "sisters" of other cnidarians, and the most closely related to bilaterians.
Cnidarians are the simplest animals in which the cells are organised into tissues.[235] The hvězdice mořská sasanka se používá jako modelový organismus ve výzkumu.[236] It is easy to care for in the laboratory and a protokol has been developed which can yield large numbers of embryos on a daily basis.[237] There is a remarkable degree of similarity in the gene sequence conservation and complexity between the sea anemone and vertebrates.[237] In particular, genes concerned in the formation of the head in vertebrates are also present in the anemone.[238][239]

Sea anemones are common in tidepools.

Jejich chapadla sting and paralyse small fish.

If an island sinks below the sea, coral growth can keep up with rising water and form an atol.

The mantle of the červené papírové lucerny medúzy crumples and expands like a paper lantern.[240]





The Portugalská válka je koloniální sifonofor

Marrus orthocanna another colonial siphonophore, assembled from two types of zooids.

Porpita porpita consists of a colony of hydroidy[241]

Lví hříva medúzy, největší známý Medúza[242]

Turritopsis dohrnii dosahuje biologická nesmrtelnost by transferring its cells back to childhood.[243][244]

The mořská vosa is the most lethal jellyfish in the world.[245]





Bilateral invertebrate animals

Některé z prvních bilaterians were wormlike, and the original bilaterální may have been a bottom dwelling worm with a single body opening.[246] A bilaterian body can be conceptualized as a cylinder with a gut running between two openings, the mouth and the anus. Kolem střeva má vnitřní tělesnou dutinu, a coelom nebo pseudocoelom.[A] Zvířata s touto bilaterálně symetrickou tělesný plán mít hlavu (přední) konec a ocas (zadní) konec, stejně jako zadní (hřbetní) a břicho (ventrální); proto mají také levou a pravou stranu.[247][248]
Mít přední konec znamená, že tato část těla naráží na podněty, jako je jídlo, které upřednostňují cefalizace, vývoj hlavy s smyslové orgány a ústa.[249] Tělo se táhne zpět od hlavy a mnoho bilateriánů má kombinaci kruhových svaly které stahují tělo, prodlužují ho a protilehlá sada podélných svalů, které zkracují tělo;[248] tyto umožňují zvířatům s měkkým tělem a hydrostatický skelet pohnout se peristaltika.[250] Mají také střevo, které prochází v podstatě válcovitým tělem z úst do konečníku. Mnoho bilaterálních kmenů má primární larvy se kterými plavat řasy a mají apikální orgán obsahující smyslové buňky. Pro každou z těchto charakteristik však existují výjimky; například dospělí ostnokožci jsou radiálně symetrickí (na rozdíl od svých larev) a jistí parazitické červy mají extrémně zjednodušené struktury těla.[247][248]

| ← bilaterians |
| |||||||||||||||||||||
Protostomy
Protostomy (z řecký pro first mouth) plocha nadkmen z zvířata. It is a sister clade of the deuterostomy (from Greek for second mouth), with which it forms the Nephrozoa clade. Protostomes are distinguished from deuterostomes by the way their embryos develop. In protostomes the first opening that develops becomes the ústa, while in deuterostomes it becomes the anus.[253][254]
| ← Protostomy |
| ||||||||||||||||||||||||||||||||||||||||||
| (existující) |


Mořští červi
Červi (Old English for hadi) form a number of phyla. Different groups of marine worms are related only distantly, so they are found in several different phyla tak jako Annelida (segmented worms), Chaetognatha (arrow worms), Phoronida (horseshoe worms), and Hemichordata. All worms, apart from the Hemichordata, are protostomes. The Hemichordata are deuterostomes and are discussed in their own section below.
The typical body plan of a worm involves long cylindrical tube-like bodies and no končetiny. Mořští červi vary in size from microscopic to over 1 metre (3.3 ft) in length for some marine polychaete worms (štětinatí červi )[255] and up to 58 metres (190 ft) for the marine nemertean worm (bootlace červ ).[256] Some marine worms occupy a small variety of parazitický niches, living inside the bodies of other animals, while others live more freely in the marine environment or by burrowing underground. Many of these worms have specialized tentacles used for exchanging oxygen and carbon dioxide and also may be used for reproduction. Some marine worms are trubkové červy, tak jako obří trubkový červ which lives in waters near underwater sopky and can withstand temperatures up to 90 degrees Celsia. Platyhelminthes (flatworms) form another worm phylum which includes a class of parasitic tapeworms. The marine tapeworm Polygonoporus giganticus, found in the gut of spermie velryby, can grow to over 30 m (100 ft).[257][258]
Nematody (roundworms) constitute a further worm phylum with tubular trávicí systémy and an opening at both ends.[259][260] Over 25,000 nematode species have been described,[261][262] of which more than half are parasitic. It has been estimated another million remain undescribed.[263] They are ubiquitous in marine, freshwater and terrestrial environments, where they often outnumber other animals in both individual and species counts. They are found in every part of the earth's litosféra, from the top of mountains to the bottom of oceánské příkopy.[264] By count they represent 90% of all animals on the dno oceánu.[265] Their numerical dominance, often exceeding a million individuals per square meter and accounting for about 80% of all individual animals on earth, their diversity of life cycles, and their presence at various trophic levels point at an important role in many ecosystems.[266]

Obří trubkoví červi cluster around hydrotermální průduchy.

Nematody are ubiquitous pseudocoelomates which can parasite marine plants and animals.

Bloodworms are typically found on the bottom of shallow marine waters.



Mořští měkkýši



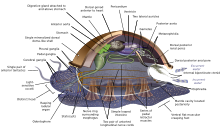
Měkkýši (Latinsky pro měkký) tvoří a kmen with about 85,000 existující uznáno druh.[269] Jsou největší námořní phylum in terms of species count, containing about 23% of all the named marine organismy.[270] Molluscs have more varied forms than other invertebrate phyla. They are highly diverse, not just in size and in anatomický structure, but also in behaviour and in habitat.
The mollusc phylum is divided into 9 or 10 taxonomické třídy. Mezi tyto třídy patří plži, mlži a hlavonožci, as well as other lesser-known but distinctive classes. Plži with protective shells are referred to as šneci, whereas gastropods without protective shells are referred to as slimáci. Plži are by far the most numerous molluscs in terms of species.[271] Mlži zahrnout škeble, ústřice, srdcovky, mušle, lastury a mnoho dalších rodiny. There are about 8,000 marine bivalves species (including brakická voda a ústí druh). A deep sea oceán quahog clam has been reported as having lived 507 years[272] making it the longest recorded life of all animals apart from koloniální animals, or near-colonial animals like houby.[215]
- Plži a mlži

Marine plži jsou mořské šneky nebo mořští slimáci. Tento nudibranch is a sea slug.

Mořský šnek Syrinx aruanus has a shell up to 91 cm long, the largest of any living gastropod.

Molluscs usually have eyes. Bordering the edge of the mantle of a lastura, a škeble mollusc, can be over 100 jednoduché oči.

Common mussel, another bivalve



Hlavonožci zahrnout chobotnice, oliheň a sépie obecná. About 800 living species of marine cephalopods have been identified,[273] and an estimated 11,000 extinct taxony byly popsány.[274] They are found in all oceans, but there are no fully freshwater cephalopods.[275]
- Hlavonožci

The nautilus je žijící fosilie little changed since it evolved 500 million years ago as one of the first hlavonožci.[276][277][278]

Cephalopods, like this sépie obecná, use their dutina pláště pro proudový pohon.

Kolosální chobotnice, the largest of all invertebrates[279]



Molluscs have such diverse shapes that many textbooks base their descriptions of molluscan anatomy on a generalized or hypothetical ancestral mollusc. This generalized mollusc is unsegmented and bilaterálně symetrické with an underside consisting of a single muscular chodidlo. Beyond that it has three further key features. Firstly, it has a muscular cloak called a plášť covering its viscera and containing a significant cavity used for breathing and vylučování. A skořápka secreted by the mantle covers the upper surface. Secondly (apart from bivalves) it has a rasping tongue called a radula used for feeding. Thirdly, it has a nervový systém including a complex digestive system using microscopic, muscle-powered hairs called řasy to exude sliz. The generalized mollusc has two paired nervové šňůry (three in bivalves). The mozek, in species that have one, encircles the jícen. Most molluscs have oči and all have sensors detecting chemicals, vibrations, and touch.[280][281]
Good evidence exists for the appearance of marine gastropods, hlavonožci and bivalves in the Kambrijský doba 541 to 485.4 před miliony let.
Marine arthropods
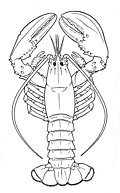

Členovci (Řek pro jointed feet) mít exoskeleton (externí kostra ), a segmentované body, and jointed přílohy (paired appendages). Tvoří a kmen který zahrnuje hmyz, pavoukovci, myriapods, a korýši. Arthropods are characterized by their jointed limbs and pokožka vyroben z chitin, often mineralised with uhličitan vápenatý. The arthropod tělesný plán skládá se z segmenty, each with a pair of přílohy. The rigid cuticle inhibits growth, so arthropods replace it periodically by línání. Their versatility has enabled them to become the most species-rich members of all ecological guilds ve většině prostředí.
The evolutionary ancestry of arthropods dates back to the Kambrijský period and is generally regarded as monofyletický. Nicméně, bazální relationships of arthropods with extinct phyla such as lobopodians have recently been debated.[285][286]
| Panarthropoda |
| ||||||||||||


- Arthropod fossils and living fossils

Fosilní trilobit. Trilobites first appeared about 521 Ma. They were highly successful and were found everywhere in the ocean for 270 Ma.[288]

The Anomalocaris ("abnormal shrimp") was one of the first apex predators and first appeared about 515 Ma.

The largest known arthropod, the mořský štír Jaekelopterus rhenaniae, bylo nalezeno v ústí strata from about 390 Ma. It was up to 2.5 m (8.2 ft) long.[289][290]

Krabi podkovy jsou živé fosilie, essentially unchanged for 450 Ma.


Extant marine arthropods range in size from the microscopic korýš Stygotantulus do Krab japonský. Arthropods' primary internal cavity is a hemocoel, which accommodates their internal orgány, and through which their hemolymfa - analogue of krev - circulates; oni mají open circulatory systems. Like their exteriors, the internal organs of arthropods are generally built of repeated segments. Jejich nervový systém is "ladder-like", with paired břišní nervové šňůry running through all segments and forming paired ganglia in each segment. Their heads are formed by fusion of varying numbers of segments, and their mozky are formed by fusion of the ganglia of these segments and encircle the jícen. The respirační a vyměšovací systems of arthropods vary, depending as much on their environment as on the subphylum ke kterému patří.
- Modern crustaceans

Many crustaceans are very small, like this tiny amphipod, and make up a significant part of the ocean's zooplankton.

The Krab japonský has the longest leg span of any arthropod, reaching 5.5 metres (18 ft) from claw to claw.[291]

The Tasmánský obří krab is long-lived and slow-growing, making it vulnerable to overfishing.[292]

Mantis krevety have the most advanced eyes in the animal kingdom,[293] and smash prey by swinging their club-like raptorial claws.[294]




Arthropod vision relies on various combinations of složené oči and pigment-pit ocelli: in most species the ocelli can only detect the direction from which light is coming, and the compound eyes are the main source of information. Arthropods also have a wide range of chemical and mechanical sensors, mostly based on modifications of the many štítek (bristles) that project through their cuticles. Arthropod methods of reproduction are diverse: terrestrial species use some form of vnitřní hnojení while marine species lay eggs using either internal or vnější hnojení. Arthropod hatchlings vary from miniature adults to grubs that lack jointed limbs and eventually undergo a total metamorfóza to produce the adult form.
Deuterostomy
v deuterostomy the first opening that develops in the growing embryo becomes the řiť, while in protostomes it becomes the mouth. Deuterostomes form a nadkmen z zvířata and are the sister clade of the protostomy.[253][254] The earliest known deuterostomes are Saccorhytus fossils from about 540 million years ago. The Saccorhytus mouth may have functioned also as its anus.[295]

| ← deuterostomy |
| ||||||||||||||||||||||||
| (existující) |

Ostnokožci

Ostnokožci (Řek pro spiny skin) is a phylum which contains only marine invertebrates. The phylum contains about 7000 living druh,[296] making it the second-largest grouping of deuterostomy, po strunatci.
Adult echinoderms are recognizable by their radiální symetrie (usually five-point) and include mořská hvězdice, mořští ježci, pískové dolary, a mořské okurky, stejně jako mořské lilie.[297] Ostnokožci se nacházejí v každé hloubce oceánu, od přílivová zóna do hlubinná zóna. They are unique among animals in having bilateral symmetry at the larval stage, but fivefold symmetry (pentamerismus, a special type of radial symmetry) as adults.[298]
Echinoderms are important both biologically and geologically. Biologically, there are few other groupings so abundant in the biotická poušť z hluboké moře, stejně jako mělčí oceány. Většina ostnokožců je schopna regenerovat tissue, organs, limbs, and rozmnožovat nepohlavně; v některých případech mohou podstoupit úplnou regeneraci z jedné končetiny. Geologicky je hodnota ostnokožců v jejich zkostnatělý kostry, které k mnoha z nich významně přispívají vápenec formace a mohou poskytnout cenné stopy o geologickém prostředí. Byly nejpoužívanějšími druhy v regenerativním výzkumu v 19. a 20. století.

Ostnokožce literally means "spiny skin", as this water melon sea urchin ilustruje.

The okrová mořská hvězda byl první keystone predator být studován. They limit mušle which can overwhelm intertidal communities.[299]

Barvitý mořské lilie in shallow waters




Mořské okurky filter feed on plankton and suspended solids.

The mořské prase, a deep water sea cucumber, is the only echinoderm that uses legged locomotion.

A benthopelagic and bioluminescence plavání mořská okurka, Hluboká 3200 metrů



Někteří vědci se domnívají, že za ozáření ostnokožců bylo odpovědné Mesozoic Marine Revolution. Kromě těžko klasifikovatelného Arkarua (A Precambrian zvíře s ostnokožcovitou pentamerní radiální symetrií), první definitivní členové kmene se objevili blízko začátku Kambrijský.
Hemichordáty

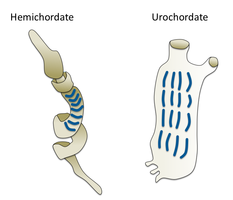
Hemichordáty tvoří sesterský kmen do ostnokožci. Jsou to osamělé červí organizmy, které lidé zřídka vidí kvůli svému životnímu stylu. Zahrnují dvě hlavní skupiny, žalud červi a Pterobranchia. Pterobranchia tvoří třídu obsahující asi 30 druhů malých červovitých zvířat, která žijí ve vylučovaných trubkách na dně oceánu. Žaludové červy tvoří třídu obsahující asi 111 druhů, které obvykle žijí v norách ve tvaru písmene U na mořském dně, od pobřeží až do hloubky 3000 metrů. Červi tam leží a chobot trčí z jednoho otvoru v norě a živí se jako podavače vkladů nebo podavače suspenzí. Předpokládá se, že předkové žaludových červů žili v tubách jako jejich příbuzní, Pterobranchia, ale nakonec začali žít bezpečnější a chráněnější existenci v sedimentárních norách.[303] Některé z těchto červů mohou dorůst až velmi dlouho; jeden konkrétní druh může dosáhnout délky 2,5 metru (8 ft 2 v), ačkoli většina červů žaludů je mnohem menších.
Žaludové červy jsou více vysoce specializované a vyspělejší než jiné organismy podobné červům. Mají oběhový systém se srdcem, který funguje také jako ledvina. Žaludové červy mají žábrovité struktury, které používají k dýchání, podobně jako žábry ryb. Proto se o žaludových červech někdy říká, že jsou spojnicí mezi klasickými bezobratlými a obratlovců. Žaludové červy neustále vytvářejí nové žaberní štěrbiny, jak rostou a někteří starší jedinci mají více než sto na každé straně. Každá štěrbina se skládá z větvové komory, která se otevírá do hltanu rozštěpem ve tvaru písmene U. Cilia tlačila vodu štěrbinami a udržovala stálý tok, stejně jako u ryb.[304] Někteří červi žaludu mají také postanální ocas, který může být homologní s postanálním ocasem obratlovců.
Třídílný tělesný plán žaludového červa již není u obratlovců přítomen, s výjimkou anatomie čelní neurální trubice, která se později vyvinula do mozku rozděleného na tři části. To znamená, že některá z původních anatomií předchůdců raných strunatců je u obratlovců stále přítomna, i když to není vždy vidět. Jednou z teorií je třídílné tělo pocházející z raného společného předka deuterostomů a možná dokonce ze společného bilaterálního předka deuterostomů i protostomů. Studie ukázaly, že genová exprese v embryu sdílí tři stejná signální centra, která formují mozky všech obratlovců, ale místo toho, aby se podílela na tvorbě jejich nervového systému,[305] řídí vývoj různých oblastí těla.[306]
Mořské strunatci
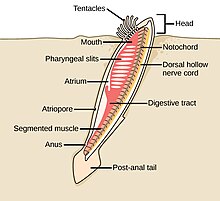
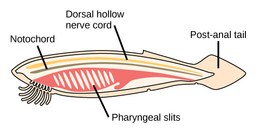
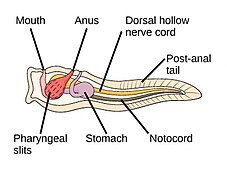
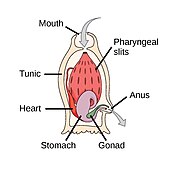
The strunat kmen má tři subfyly, z nichž jeden je obratlovců (viz. níže). Další dva subfyly jsou mořští bezobratlí: pláštěnci (salpy a moře stříká ) a cephalochordates (jako lancelety ). Chordáty bezobratlých jsou blízcí příbuzní obratlovců. Zejména se diskutovalo o tom, jak blízko jsou některé vyhynulé mořské druhy, jako např Pikaiidae, Palaeospondylus, Zhongxiniscus a Vetulicolia, může souviset předky s obratlovci.
- Chordáty bezobratlých jsou blízcí příbuzní obratlovců

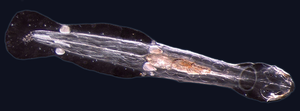
The kopí, malá průsvitná ryba cephalochordate, je nejbližší žijící příbuzný bezobratlých obratlovců.[308][309]

Pláštěnci, jako tito fluoreskující mořské stříkačky, může poskytnout stopy obratlovcům, a tedy i lidským předkům.[310]

Pyrosomy jsou volně plovoucí bioluminiscenční pláštěnky složené ze stovek jedinců.

Salp řetěz




Obratlovců
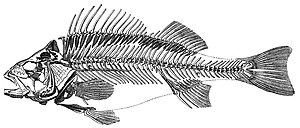
Obratlovci (Latinsky pro klouby páteře) plocha subphylum z strunatci. Jsou to strunatci, kteří mají a páteř (páteř). Páteř poskytuje centrální podpůrnou strukturu pro vnitřní kostra který poskytuje tělu tvar, podporu a ochranu a může poskytnout prostředek k ukotvení ploutví nebo končetin k tělu. Páteř také slouží k uložení a ochraně mícha který leží uvnitř páteře.
Mořské obratlovce lze rozdělit na mořské Ryba a námořní tetrapody.
Mořské ryby
Ryby obvykle dýchají extrakcí kyslíku z vody žábry a mít pokožku chráněnou váhy a sliznice. Používají ploutve pohánět a stabilizovat se ve vodě a obvykle mít a dvoukomorové srdce a oči dobře přizpůsobené pro vidění pod vodou, stejně jako jiné senzorické systémy. Od roku 2017 bylo popsáno více než 33 000 druhů ryb,[311] z toho asi 20 000 mořských ryb.[312]
| ← obratlovců |
| |||||||||||||||||||||||||||
| (existující) |
Ryby bez čelistí

Rané ryby neměly žádné čelisti. Většina z nich vyhynula, když je překonaly čelistní ryby (níže), ale dvě skupiny přežily: hagfish a lampreys. Hagfish tvoří třídu asi 20 druhů úhoř -tvarovaná, sliz -produkce mořských ryb. Jsou jedinými známými živými zvířaty, která mají lebka ale ne páteř. Lampreys tvoří nadtřídu obsahující 38 známých existujících druhů rodu ryby bez čelistí.[313] Dospělý mihule se vyznačuje ozubenými, nálevkovitými sacími ústy. Ačkoli jsou dobře známí tím, že se nudí do masa jiných ryb sát jejich krev,[314] pouze 18 druhů lampreys je ve skutečnosti parazitických.[315] Společně hagfish a lampreys jsou sesterskou skupinou obratlovců. Živé hagfish zůstávají podobné hagfish z doby před asi 300 miliony let.[316] Tyto lampreys jsou velmi starodávná linie obratlovců, ačkoli jejich přesný vztah k hagfishes a čelistní obratlovci je stále předmětem sporu.[317] Molekulární analýza od roku 1992 naznačuje, že hagfish se nejvíce podobá lampreys,[318] a tak jsou i obratlovci v a monofyletický smysl. Jiní je považují za sesterskou skupinu obratlovců v běžném taxonu craniata.[319]
The Tully monstrum je vyhynulý rod měkkých bilateriánů, kteří žili v tropických ústí řek asi před 300 miliony let. Od roku 2016 se vedou spory o to, zda toto zvíře bylo obratlovcem nebo bezobratlým.[320][321] V roce 2020 našli vědci „silné důkazy“, že monstrum v Tully bylo obratlovcem a bylo ryby bez čelistí v linii rodu mihule.[322][323]

Lampreys jsou často parazitické a mají zubatá, nálevkovitá sající ústa.

Zaniklý Pteraspidomorphi, předci čelistní obratlovci


Pteraspidomorphi je zaniklý třída raných ryb bez čelistí předků čelistních obratlovců. Těch několik charakteristik, které s nimi sdílejí, je nyní považováno za primitivní pro všechny obratlovců.
Kolem začátku Devonský, ryby se začaly objevovat s hlubokou přestavbou lebky obratlovců, která vyústila v čelist.[324]Z těchto raných rybích čelistí se vyvinuly všechny čelisti obratlovců, včetně lidské čelisti. Vzhled rané čelisti obratlovců byl popsán jako „možná nejhlubší a nejradikálnější evoluční krok v historii obratlovců“.[325][326] Čelisti umožňují chytit, držet a žvýkat kořist. Ryby bez čelistí měl větší potíže s přežitím než ryby s čelistmi a většina ryb bez čelistí vyhynula během triasu.
Paryby
Čelist ryby spadají do dvou hlavních skupin: ryby s kostnatými vnitřními kostry a ryby s chrupavčitými vnitřními kostry. Chrupavčité ryby, jako např žraloci a paprsky, nechat si vyrobit čelisti a kostry chrupavka spíše než kost. Megalodon je vyhynulý druh žraloka, který žil asi 28 až 1,5 Ma. Vypadalo to hodně jako podsaditá verze velký bílý žralok, ale byl mnohem větší a fosilní délky dosahovaly 20,3 metrů (67 ft).[327] Nalezeno ve všech oceánech[328] byl to jeden z největších a nejmocnějších predátorů v historii obratlovců,[327] a pravděpodobně měl hluboký dopad na mořský život.[329] The Grónský žralok má nejdelší známou životnost ze všech obratlovců, přibližně 400 let.[330] Někteří žraloci, jako je velký bílý, jsou částečně teplokrevní a rodí živě. The rejnok, největší paprsek na světě, byl zaměřen na rybolov a nyní je zranitelný.[331]
- Paryby

Z chrupavčitých ryb se možná vyvinuly ostnaté žraloky.

rejnok, největší paprsek





Zaniklý megalodon připomínal obra velký bílý žralok.

The Grónský žralok žije déle než kterýkoli jiný obratlovec.

Největší existující ryby žralok velrybí, je nyní zranitelné druhy.


Kostnatá ryba



Kostnaté ryby mají čelisti a kostry kost spíše než chrupavka. Kostnaté ryby mají také tvrdé, kostnaté talíře operculum které jim pomáhají dýchat a chránit žábry a často mají a plynový měchýř které používají pro lepší kontrolu nad svým vztlakem. Kostnaté ryby lze dále rozdělit na ryby s lalokové ploutve a ti s paprskové ploutve. Přibližná data ve fylogenetickém stromu pocházejí z Near et al., 2012[333] a Zhu et al., 2009.[334]
| ← kostnatá ryba |
| ||||||||||||||||||||||||||||||
| (existující) |
Lalokové ploutve mají podobu masité laloky podporované kostnatými stopkami, které sahají od těla.[335] Guiyu oneiros, nejdříve známá kostnatá ryba, žila během pozdní doby Silurian Před 419 miliony let. Má kombinaci obou paprskové žebrované a rysy s lalokovými ploutvemi, ačkoli analýza všech jejích rysů ji přibližuje k rybám s lalokovými ploutvemi.[334] Z lalokových ploutví se vyvinuly nohy prvních čtyřnohých suchozemských obratlovců, takže časným předchůdcem člověka byla ryba s laločnatými ploutvemi. Na rozdíl od coelacanths a lungfishes, laločnaté ploutve ryby jsou nyní zaniklé.
Zbývající kostnaté ryby mají paprskové ploutve. Jsou vyrobeny z pásů kůže podepřených kostnatými nebo nadrženými ostny (paprsky), které lze vztyčovat za účelem kontroly tuhosti ploutví.
- Hlavní charakteristický rys chondrosteans (jeseter, vesloň, bichir a rákosník ) je chrupavčitá povaha jejich koster. Předkové chondrostanů jsou považováni za kostnaté ryby, ale charakteristika zkostnatělé kostry se ztratila v pozdějším evolučním vývoji, což vedlo k zesvětlení rámu.[336]
- Neopterygians (z řečtiny pro nové ploutve) se objevil někdy v pozdním permu před dinosaury. Byli velmi úspěšnou skupinou ryb, protože se mohli pohybovat rychleji než jejich předkové. Jejich váhy a kostry se během jejich vývoje začaly zesvětlovat a jejich čelisti se staly silnějšími a efektivnějšími.[337]
Teleosts

Asi 96% všech moderních druhů ryb jsou teleosts,[338] z toho asi 14 000 mořských druhů.[339] Teleosty lze odlišit od jiných kostnatých ryb podle jejich vlastnictví a homocercal ocas, ocas, kde horní polovina zrcadlí dolní polovinu.[340] Další rozdíl spočívá v jejich čelistních kostech - teleosty mají úpravy v čelistním svalstvu, které jim umožňují vyčnívají jejich čelisti. To jim umožňuje chytit kořist a vtáhněte jim to do úst.[340] Obecně platí, že teleosty mají tendenci být rychlejší a pružnější než více bazálních kostnatých ryb. Jejich kosterní struktura se vyvinula směrem k větší lehkosti. Zatímco teleostní kosti jsou v pořádku kalcifikovaný, jsou vyrobeny z lešení vzpěr, spíše než z hustého spongiózní kosti z holostean Ryba.[341]
Teleosty se nacházejí téměř ve všech mořská stanoviště.[342] Mají obrovské rozmanitost a velikost se pohybuje od dospělých blázni 8 mm dlouhý [343] na mořské mušle váha přes 2 000 kg.[344] Následující obrázky ukazují něco z rozmanitosti ve tvaru a barvě moderních mořských teleostů ...








Téměř polovina všech existujících druhů obratlovců jsou teleosts.[345]
Mořští tetrapods

A tetrapod (Řek pro čtyři stopy) je obratlovec s končetiny (chodidla). Tetrapods se vyvinuli z dávných dob laločnaté ryby asi před 400 miliony let během Devonské období když se jejich nejranější předkové vynořili z moře a přizpůsobili se životu na zemi.[346] Tato změna z plánu těla pro dýchání a navigaci v gravitačně neutrální vodě na plán těla s mechanismy umožňujícími zvířeti dýchat vzduch bez dehydratace a pohybovat se po zemi je jednou z nejhlubších známých evolučních změn.[347][348] Tetrapods lze rozdělit do čtyř tříd: obojživelníci, plazi, ptactvo a savci.
| ← tetrapody |
| ||||||||||||||||||
Mořští tetrapods jsou tetrapods, kteří se vrátili ze země zpět do moře. K prvním návratům do oceánu mohlo dojít již v Období karbonu[349] vzhledem k tomu, že k dalším návratům došlo až nedávno Kenozoikum, jako u kytovců, ploutvonožci,[350] a několik moderní obojživelníci.[351] Obojživelníci (z řečtiny pro oba druhy života) žijí část svého života ve vodě a část na souši. K reprodukci většinou vyžadují sladkou vodu. Několik obývá brakickou vodu, ale neexistují žádní skuteční mořští obojživelníci.[352] Existují však zprávy o obojživelnících napadajících mořské vody, například o invazi Černého moře pomocí přírodního hybridu Pelophylax esculentus hlášeno v roce 2010.[353]
Plazi
Plazi (Pozdní latina pro plíživý nebo plazení) nemají vodní larvální stádium, a tímto způsobem jsou na rozdíl od obojživelníků. Většina plazů je oviparous, ačkoli několik druhů squamates jsou viviparous, stejně jako některé zaniklé vodní klady[354] - plod se vyvíjí uvnitř matky, obsažený v a placenta spíše než skořápka. Tak jako amniotes Vajíčka plazů jsou pro ochranu a transport obklopena membránami, které je přizpůsobují reprodukci na suchu. Mnoho z viviparous druhů krmit jejich plody prostřednictvím různých forem placenty analogických k těm z savci, přičemž někteří zajišťují počáteční péči o jejich mláďata.
Někteří plazi mají bližší příbuznost ptactvo než ostatní plazi a mnoho vědců dává přednost tomu, aby se z Reptilie stala monofyletická skupina zahrnující ptáky.[355][356][357][358] Existující patří mezi ně jiné než ptáky mořské želvy, mořští hadi, želvy, mořský leguán a mořský krokodýl. V současné době z přibližně 12 000 existujících plaz druhů a poddruhů, jen asi 100 z nich je klasifikováno jako mořští plazi.[359]
S výjimkou některých mořských hadů je většina existujících mořských plazů oviparous a je třeba se vrátit na pevninu a položit vajíčka. Kromě mořských želv tento druh obvykle tráví většinu svého života na zemi nebo v její blízkosti, spíše než v oceánu. Mořští hadi obecně upřednostňují mělké vody v blízkosti pevniny, kolem ostrovů, zejména vod, které jsou poněkud chráněné, a také v blízkosti ústí řek.[360][361] Na rozdíl od suchozemských hadů se u mořských hadů vyvinuly zploštělé ocasy, které jim pomáhají plavat.[362]

Mořští hadi mají zploštělé ocasy.

Prastarý Ichthyosaurus communis nezávisle vyvinuté ploutve podobné delfínům.





Nějaký vyhynulý mořští plazi, jako např ichtyosaury, se vyvinulo viviparous a neměl požadavek na návrat na pevninu. Ichthyosaurs připomínal delfíny. Poprvé se objevily asi před 245 miliony let a zmizely asi před 90 miliony let. Pozemský předek ichtyosaura neměl na zádech ani na ocase žádné rysy, které by mohly pomáhat evolučnímu procesu. Přesto jejichtyosaurus vyvinul a hřbetní a ocasní ploutev což zlepšilo jeho schopnost plavat.[363] Biolog Stephen Jay Gould řekl, že ichtyosaurus byl jeho oblíbeným příkladem konvergentní evoluce.[364] Nejstarší mořští plazi se objevili v Permu. Během Druhohor mnoho skupin plazů se přizpůsobilo životu v mořích, včetně ichtyosaury, plesiosaurs, mosasaurs, nothosaurs, placodonts, mořské želvy, thalattosaurs a thalattosuchians. Mořští plazi byli poté méně početní masový zánik na konci Křídový.
Ptactvo

Mořští ptáci jsou přizpůsobeno k životu uvnitř námořní životní prostředí. Často se jim říká mořští ptáci. I když se mořští ptáci značně liší v životním stylu, chování a fyziologii, často vykazují nápadné rysy konvergentní evoluce, jako stejné problémy životního prostředí a krmení výklenky vedly k podobným úpravám. Mezi příklady patří albatros, tučňáci, terejové, a auks.
Obecně platí, že mořští ptáci žijí déle, plemeno později a mají méně mláďat než suchozemští ptáci, ale do svých mláďat investují spoustu času. Většina druh vnořit se kolonie, které se mohou lišit velikostí od několika desítek ptáků po miliony. Mnoho druhů je proslulých podnikáním dlouhých ročních migrace, přejezd přes rovník nebo v některých případech obeplout Zemi. Živí se jak na povrchu oceánu, tak pod ním, a dokonce se živí navzájem. Mořští ptáci mohou být velmi pelagický pobřežní, nebo v některých případech strávit část roku úplně od moře. Někteří mořští ptáci klesají z výšek a vrhají se do vody a zanechávají stopy podobné páře, podobné jako u bojových letadel.[365] Terejové ponořte se do vody rychlostí až 100 kilometrů za hodinu (60 mph). Mají vzduchové vaky pod kůží v obličeji a na hrudi, které fungují jako bublinková folie, tlumící náraz vodou.

Racek stříbřitý zaútočte shora na školy sledě.

Gentoo tučňák plavání pod vodou

Gannety „potápějí“ vysokou rychlostí.

Albatrosy se pohybují na obrovských plochách oceánu a pravidelně obíhají po celém světě.




První mořští ptáci se vyvinuli v Křídový doba a na ostrově se objevily moderní rodiny mořských ptáků Paleogen.
Savci

Savci (z latiny pro prsa) jsou charakterizovány přítomností mléčné žlázy v kterém ženy vyrobit mléko za krmení (ošetřování) svých mláďat. Existuje asi 130 živých a nedávno vyhynulých mořských živočichů savec druhy jako těsnění, delfíni, velryby, kapustňáci, mořské vydry a lední medvědi.[366] Nepředstavují samostatný taxon nebo systematické seskupení, ale místo toho je sjednocuje jejich závislost na krmení mořského prostředí. Kytovci i sireniani jsou plně vodní, a proto jsou povinnými obyvateli vody. Tuleni a lachtani jsou semiaquatic; většinu času tráví ve vodě, ale musí se vrátit na pevninu kvůli důležitým činnostem jako např páření, chov a línání. Naproti tomu vydry i lední medvěd jsou mnohem méně přizpůsobeny vodním živobytím. Jejich strava se také značně liší: někteří mohou jíst zooplankton; jiní mohou jíst ryby, chobotnice, korýše a mořské trávy; a několik jich může jíst jiné savce.
V procesu konvergentní evoluce, mořští savci, zejména kytovci, jako jsou delfíni a velryby, přestavěli své tělesný plán paralelně s efektivnější fusiform tělesný plán pelagické ryby. Přední nohy se staly ploutve a zadní nohy zmizely, a hřbetní ploutev znovu se objevil a ocas se proměnil v mocnou horizontálu náhoda. Tento plán těla je adaptací na to, že je aktivním predátorem ve výšce táhnout životní prostředí. S dnes již vyhynulým mořským plazem došlo k paralelní konvergenci ichtyosaurus.[367]

Ohrožený modrá velryba, největší živé zvíře[368]

Delfín skákavý, který má nejvyšší encephisation jakéhokoli zvířete po lidech[369]

Dugong pasoucí se dál mořská tráva






Prvovýrobci

Prvovýrobci jsou autotrofní organismy, které vytvářejí své vlastní jídlo, místo aby jedly jiné organismy. To znamená, že primární producenti se stávají výchozím bodem v EU potravní řetězec pro heterotrofní organismy, které jedí jiné organismy. Někteří mořští primární producenti jsou specializované bakterie a archea, které jsou chemotrofy, připravují si vlastní jídlo shromažďováním hydrotermální průduchy a studené prosakuje a pomocí chemosyntéza. Nicméně většina námořní prvovýroba pochází z organismů, které používají fotosyntéza na oxidu uhličitém rozpuštěném ve vodě. Tento proces využívá energii ze slunečního záření k přeměně vody a oxid uhličitý[370]:186–187 na cukry, které lze použít jak jako zdroj chemické energie, tak i organických molekul, které se používají ve strukturních složkách buněk.[370]:1242 Prvotní producenti moří jsou důležití, protože vytvářejí většinu z nich a podporují téměř celý život mořských živočichů kyslík a jídlo, které poskytuje jiným organismům chemickou energii, kterou potřebují k existenci.
Hlavními mořskými primárními producenty jsou sinice, řasy a mořské rostliny. The kyslík uvolněný jako vedlejší produkt fotosyntézy je zapotřebí téměř všechno živé buněčné dýchání. Primární producenti mají navíc celosvětový vliv uhlík a voda cykly. Stabilizují pobřežní oblasti a mohou poskytovat stanoviště mořským živočichům. Termín divize byl tradičně používán místo kmen když diskutujeme o prvovýrobcích, ale Mezinárodní kód nomenklatury pro řasy, houby a rostliny nyní přijímá oba výrazy jako ekvivalenty.[371]
Sinice


Sinice byly prvními organismy, které vyvinuly schopnost přeměňovat sluneční světlo na chemickou energii. Tvoří kmen (rozdělení) bakterií, které se pohybují od jednobuněčných po vláknitý a zahrnout koloniální druhy. Vyskytují se téměř všude na Zemi: ve vlhké půdě, ve sladkovodním i mořském prostředí a dokonce i na antarktických skalách.[372] Zejména některé druhy se vyskytují jako unášené buňky plovoucí v oceánu a jako takové byly mezi prvními z fytoplankton.
Prvními primárními producenty, kteří používali fotosyntézu, byly oceánské sinice asi před 2,3 miliardami let.[373][374] Uvolňování molekulárních kyslík sinicemi jako vedlejší produkt fotosyntézy vyvolaných globálních změn v prostředí Země. Protože kyslík byl v té době toxický pro většinu života na Zemi, vedlo to k téměř vyhynutí organismy netolerující kyslík, a dramatická změna který přesměroval vývoj hlavních živočišných a rostlinných druhů.[375]
Drobná mořská sinice Prochlorococcus, objevený v roce 1986, dnes tvoří součást základny oceánu potravní řetězec a představuje velkou část fotosyntézy otevřeného oceánu[376] a odhadem 20% kyslíku v zemské atmosféře.[377] Je to pravděpodobně nejhojnější rod na Zemi: jeden mililiter povrchové mořské vody může obsahovat 100 000 buněk nebo více.[378]
Biologové původně klasifikovali sinice jako řasy a označovali je jako „modrozelené řasy“. Novější názor je, že sinice jsou bakterie, a proto ani nejsou stejné Království jako řasy. Většina úřadů dnes vylučuje všechny prokaryoty, a tedy sinice z definice řas.[379][380]
Řasy



Řasy je neformální termín pro rozšířenou a různorodou skupinu fotosyntézy protistů které nemusí nutně úzce souviset a jsou tedy polyfyletický. Mořské řasy lze rozdělit do šesti skupin:
- zelené řasy, neformální skupina obsahující asi 8 000 uznávaných druhů.[381] Mnoho druhů žije většinu svého života jako jednotlivé buňky nebo jsou vláknité, zatímco jiné se tvoří kolonie jsou tvořeny dlouhými řetězci buněk nebo jsou vysoce diferencovanými makroskopickými mořskými řasami.
- červené řasy (sporný) kmen obsahující přibližně 7 000 uznávaných druhů,[382] většinou mnohobuňečný a včetně mnoha pozoruhodných mořské řasy.[382][383]
- hnědé řasy, a třída obsahující asi 2 000 uznávaných druhů,[384] většinou mnohobuňečný a včetně mnoha mořských řas, včetně kelp
- rozsivky, (sporný) kmen obsahující asi 100 000 uznávaných druhů převážně jednobuněčných řas. Diatomy každoročně generují asi 20 procent kyslíku produkovaného na planetě,[151] přijmout více než 6,7 miliardy metrických tun křemík každý rok z vod, ve kterých žijí,[385] a přispívají téměř polovinou organického materiálu nalezeného v oceánech. Mušle (frustule ) mrtvých rozsivek může dosáhnout až půl míle hluboko na dně oceánu.[386]
- dinoflageláty, kmen jednobuněčných bičíkovců s asi 2 000 mořskými druhy.[387] Je známo mnoho dinoflagelátů fotosyntetický, ale velká část z nich je ve skutečnosti mixotrofní kombinující fotosyntézu s požitím kořisti (fagotrofie ).[388] Některé druhy jsou endosymbionty mořských živočichů a hrají důležitou roli v biologii korálové útesy. Jiné předcházejí jiným prvokům a několik forem je parazitických.
- euglenofyty, kmen jednobuněčných bičíkovců pouze s několika členy námořní pěchoty
Na rozdíl od vyšších rostlin postrádají řasy kořeny, stonky nebo listy. Mohou být klasifikovány podle velikosti jako mikrořasy nebo makrořasy.
Mikrořasy jsou mikroskopické druhy řas, které nejsou viditelné pouhým okem. Většinou jsou jednobuněčný druhy, které existují jako jednotlivci nebo v řetězcích nebo skupinách, i když některé jsou mnohobuňečný. Mikrořasy jsou důležitou součástí mořských protistů (diskutováno výše ), stejně jako fytoplankton (diskutováno níže ). Oni jsou velmi různorodý. Odhaduje se, že existuje 200 000–800 000 druhů, z nichž bylo popsáno asi 50 000 druhů.[389] V závislosti na druhu se jejich velikost pohybuje od několika mikrometrů (μm) do několika set mikrometrů. Jsou speciálně přizpůsobeny prostředí, v němž dominují viskózní síly.

Chlamydomonas globosajednobuněčná zelená řasa se dvěma bičíky viditelné vlevo dole

Chlorella vulgaris, společná zelená mikrořasy, v endosymbióza s ciliate[390]

Centrická rozsivka

Dinoflageláty




Makrořasy jsou větší, mnohobuňečný a viditelnější druhy řas, běžně nazývané mořské řasy. Mořské řasy obvykle rostou v mělkých pobřežních vodách, kde jsou ukotveny k mořskému dnu pomocí sevření. Mořské řasy, které se zmítají, se mohou umýt na plážích. Kelp je velká hnědá mořská řasa, která se vytváří pod vodou lesy pokrývající přibližně 25% světového pobřeží.[391] Patří mezi nejproduktivnější a nejdynamičtější ekosystémy na Zemi.[392] Nějaký Sargassum mořské řasy jsou planktonické (volně plovoucí). Stejně jako mikrořasy jsou i technicky řasy (mořské řasy) mořští protisti protože to nejsou skutečné rostliny.

Mořské řasy jsou makroskopickou formou
Červené nebo hnědý nebo zelené řasy.
Sargassum mořské řasy jsou planktonická hnědá řasa se vzduchovými měchýři, které jí pomáhají plavat.

Sargassum ryby jsou maskovaní, aby žili mezi driftováním Sargassum Mořská řasa.



- Jednobuněčné makrořasy (viz také makroskopičtí protisti ← )

Jednobuněčný bublinkové řasy žije v přílivové zóny. Může mít průměr 4 cm.[393]

Jednobuněčný skleničku na mořskou pannu jsou houbovité řasy, které dorůstají až do výšky 10 cm.

Zabijácké řasy jsou jednobuněčné organismy, ale vypadají jako kapradiny a rostou stonky dlouhé až 80 cm.[394]



Jednobuněčné organismy jsou obvykle mikroskopické, méně než jedna desetina milimetru dlouhé. Existují výjimky. Skleničku na mořskou pannu, rod subtropického zelené řasy, je jednobuněčný, ale pozoruhodně velký a složitý ve formě s jediným velkým jádrem, což z něj dělá modelový organismus pro studium buněčná biologie.[395] Další jednobuněčné řasy, Caulerpa taxifolia, má vzhled cévnaté rostliny včetně "listů" uspořádaných úhledně do stopek jako kapradina. Selektivní chov v akváriích za účelem produkce tvrdších kmenů vedl k náhodnému úniku do Středomoří, kde se stalo invazivní druhy hovorově známý jako zabijácké řasy.[396]
Původ rostlin

Zpátky v Silurian, se vyvinul nějaký fytoplankton Červené, hnědý a zelené řasy. Tyto řasy pak napadly zemi a začaly se vyvíjet do suchozemských rostlin, které dnes známe. Později v Křídový, některé z těchto suchozemských rostlin se vrátily do moře jako mořské rostliny, jako např mangrovy a mořské trávy.[397]
Mořské rostliny najdete v přílivové zóny a mělké vody, jako např mořské trávy jako eelgrass a želví tráva, Thalassia. Tyto rostliny se přizpůsobily vysoké slanosti oceánského prostředí. Život rostlin může také vzkvétat v brakických vodách ústí řek, kde mangrovy nebo cordgrass nebo plážová tráva plážová tráva může růst.

Mořští draci maskovaný, aby vypadal jako plovoucí mořské řasy žijící v řasových lesích a na loukách z mořských řas.[398]



Celková světová plocha mangrovových lesů byla v roce 2010 odhadována na 134 257 kilometrů čtverečních (51337 čtverečních mil) (na základě satelitních údajů).[399][400] Celková světová plocha louk z mořských řas je obtížnější určit, ale byla konzervativně odhadnuta v roce 2003 na 177 000 kilometrů čtverečních (68 000 čtverečních mil).[401]
Mangrovy a mořské trávy poskytují důležitá stanoviště mateřských škol pro mořský život, fungují jako úkryty a místa pro hledání potravy pro larvální a juvenilní formy větších ryb a bezobratlých.
- Spalding, M. (2010) Atlas světa mangrovů, Routledge. ISBN 9781849776608. doi:10.4324/9781849776608.
Plankton a trofické interakce

Plankton (z řečtiny pro poutníci) jsou různorodá skupina organismů, které žijí v vodní sloup velkých vodních ploch, ale nedokáže plavat proti proudu. Výsledkem je, že bloudí nebo unášejí proudy.[402] Plankton jsou definovány jejich ekologický výklenek, nikoliv fylogenetické nebo taxonomické klasifikace. Jsou klíčovým zdrojem potravy pro mnoho mořských živočichů píce ryby na velryby. Plankton lze rozdělit na rostlinnou složku a živočišnou složku.
Fytoplankton
Fytoplankton jsou rostlinné složky komunity planktonu („fyto“ pochází z řečtiny pro rostlina). Oni jsou autotrofní (samozkrmování), což znamená, že si vytvářejí vlastní potravu a nepotřebují konzumovat jiné organismy.
Fytoplankton se skládá hlavně z mikroskopické fotosyntézy eukaryoty které obývají horní vrstvu zalitou sluncem ve všech oceánech. Potřebují sluneční světlo, aby mohli fotosyntetizovat. Většina fytoplanktonu jsou jednobuněčné řasy, ale jiný fytoplankton jsou bakterie a některé jsou protistů.[403] Skupiny fytoplanktonu zahrnují sinice (výše), rozsivky, různé další typy řasy (červená, zelená, hnědá a žlutozelená), dinoflageláty, euglenoidy, coccolithophorids, kryptomonády, chryzofyty, chlorofyty, prasinofyty, a silicoflagellates. Tvoří základnu prvovýroba který pohání oceán web s potravinami a představují polovinu současné globální primární produkce, více než suchozemské lesy.[404]


- Fytoplankton

Fytoplankton je základem oceánského potravinového řetězce.

Fytoplankton má mnoho tvarů a velikostí.

Diatomy jsou jedním z nejběžnějších typů fytoplanktonu.

Koloniální fytoplankton





Sinice Prochlorococcus představuje velkou část primární produkce oceánu.

Zelená sinice spodina se vyplavila na skálu v Kalifornii

Gyrodinium, jeden z mála nahých dinoflagelátů, kterým chybí brnění

Zoochlorellae (zelená) žijící uvnitř ciliate Stichotricha secunda







Kvetou řasy z Emiliania huxleyi u jižního pobřeží Anglie




Zooplankton


Zooplankton jsou živočišnou složkou planktonické komunity ("zoo" pochází z řečtiny pro zvíře). Oni jsou heterotrofní (jiné krmení), což znamená, že nemohou vyrábět své vlastní jídlo a musí místo toho konzumovat jiné rostliny nebo zvířata jako potravu. To zejména znamená, že jedí fytoplankton.



úzce replikovat některé radiolarianské skořápkové vzory[407]
Zooplankton je obecně větší než fytoplankton, většinou stále mikroskopický, ale některé lze pozorovat pouhým okem. Mnoho prvoky (jednobuněčné protistů že kořist na jiný mikroskopický život) jsou zooplankton, včetně zooflageláty, foraminiferans, radiolariáni a nějaký dinoflageláty. Jiné dinoflageláty jsou mixotrofní a lze je také klasifikovat jako fytoplankton; rozdíl mezi rostlinami a zvířaty se často rozpadá ve velmi malých organismech. Další zooplankton zahrnuje pelagický cnidarians, ctenofory, měkkýši, členovci a pláštěnci, stejně jako plankton šípové červy a štětinatí červi.
Radiolariáni jsou jednobuněčné protistů s propracovanými křemičitými granáty
Microzooplankton: hlavní pastevci planktonu

Radiolariáni mají mnoho tvarů.

Skupina planktic foraminiferans

Copepods jíst fytoplankton. Tenhle nese vejce.

Dinoflagelát, Protoperidinium vytlačuje velký krmný závoj, aby zachytil kořist.




Větší zooplankton může být dravý na menším zooplanktonu.
Makrozooplankton

Tomopteris, planktonový segmentovaný červ s neobvyklou žlutou barvou bioluminiscence[408]

Marine amphipod

Pelagická mořská okurka







| Externí video | |
|---|---|
Mnoho mořských živočichů začíná život jako zooplankton ve formě vajíček nebo larev, než se z nich vyvinou dospělí. Tyto jsou meroplanktic, to znamená, že jsou planktonní jen po část svého života.
- Larvy a mladiství

Z vajíčka se vylíhla larva lososa

Oceánský měsíček larva

Juvenilní plankton oliheň
 Larva stádium ostnatého humra
Larva stádium ostnatého humra




Mixotrofní plankton


Dinoflageláty jsou často mixotrofní nebo bydlet v symbióza s jinými organismy.
- Mixoplankton

Tintinnid ciliate Favella

Euglena mutabilis, fotosyntetický bičíkatý

Noctiluca scintillans, bioluminiscenční dinoflagelát



Některé dinoflageláty jsou bioluminiscenční. V noci se může oceánská voda vnitřně rozsvítit a jiskra s modrým světlem kvůli těmto dinoflagelátům.[410][411] Bioluminiscenční dinoflageláty mají scintillony, individuální cytoplazmatický těla, která obsahují dinoflagellát luciferáza, hlavní enzym zapojený do luminiscence. Luminiscence, někdy nazývaná fosforescence moře, nastává jako krátké (0,1 s) modré záblesky nebo jiskry, když jsou stimulovány jednotlivé scintillony, obvykle mechanickými poruchami, například od lodi nebo plavce nebo surfování.[412]
Mořské jídlo web

Ve srovnání s pozemským prostředím mají mořská prostředí pyramidy biomasy, které jsou obrácené na základně. Zejména biomasa spotřebitelů (kozonožci, krill, krevety, píce) je větší než biomasa primárních producentů. Stává se to proto, že primárními producenty oceánu jsou drobný fytoplankton, který obvykle bývá r-stratégové které rychle rostou a množí se, takže malá hmota může mít vysokou rychlost primární produkce. Naproti tomu jsou často suchozemští prvovýrobci, jako jsou zralé lesy K-stratégové které rostou a množí se pomalu, takže k dosažení stejné rychlosti primární produkce je zapotřebí mnohem většího množství.
Kvůli této inverzi tvoří většinu mořského živočicha zooplankton biomasa. Tak jako primární spotřebitelé, jsou zásadním spojením mezi primárními producenty (hlavně fytoplanktonem) a zbytkem mořské potravinové sítě (sekundární spotřebitelé ).[413]
Pokud fytoplankton zemře dříve, než se sní, sestupuje skrz eufotická zóna jako součást mořský sníh a usazuje se do hlubin moře. Tímto způsobem fytoplankton každoročně odděluje do oceánu asi 2 miliardy tun oxidu uhličitého, což způsobí, že oceán se stane záchytem oxidu uhličitého, který drží asi 90% veškerého zachyceného uhlíku.[414]
V roce 2010 vědci zjistili, že velryby přenášejí živiny z hlubin oceánu zpět na povrch pomocí procesu, kterému říkali velrybí čerpadlo.[415] Velryby se krmí na hlubších úrovních v oceánu, kde krill je nalezen, ale pravidelně se vracejte na povrch, abyste dýchali. Tam velryby vyprázdněte tekutinu bohaté na dusík a železo. Místo ponoření zůstává kapalina na povrchu, kde fytoplankton konzumovat to. V Mainském zálivu poskytuje velrybí čerpadlo více dusíku než řeky.[416]
Usazeniny a biogenní bláto
Usazeniny na dně oceánu mají dva hlavní původy, terigenní a biogenní. Terénní sedimenty tvoří asi 45% celkového mořského sedimentu a pochází z eroze skály na souši, přepravované řekami a odtokem půdy, polétavým prachem, sopkami nebo broušením ledovci.
Biogenní sedimenty tvoří dalších 55% celkového sedimentu a pocházejí z kosterních zbytků mořští protisti (jednobuněčné planktonové a bentosové organismy). Může být také přítomno mnohem menší množství vysrážených minerálů a meteorického prachu. Sliz, v souvislosti s mořským sedimentem, neodkazuje na konzistenci sedimentu, ale na jeho biologický původ. Termín bláto původně používal John Murray „otec moderní oceánografie“, který tento termín navrhl radiolarianský bláto pro usazeniny oxidu křemičitého z radiolarianských skořápek vynesených na povrch během Expedice Challenger.[417] A biogenní sliz je pelagický sediment obsahující nejméně 30 procent z kosterních pozůstatků mořských organismů.
Hlavní typy biogenního bahna | |||||||
|---|---|---|---|---|---|---|---|
| typ | minerální formuláře | protist odpovědný | jméno kostra | popis | |||
| Křemičitý bláto | SiO2 křemen sklenka opál rohovník | rozsivky |  | frustrace | Velikost jednotlivých rozsivek se pohybuje od 0,002 do 0,2 mm.[418] | ||
| radiolariáni |  | kostra | Radiolarians jsou prvoky s průměry mezi 0,1 a 0,2 mm, které produkují složité minerální kostry, obvykle vyrobené z oxidu křemičitého | ||||
| Vápenatý bláto | CaCO3 kalcit aragonit vápenec křída | foraminiferans |  | test | Existuje asi 10 000 živých druhů foraminiferanů,[419] obvykle menší než 1 mm. | ||
| coccolithophores |  | kokcolith | Coccolithophores jsou sférické buňky obvykle menší než 0,1 mm v průměru, uzavřené vápnitými deskami zvanými kokokolity.[420] Coccolithové jsou důležití mikrofosílie. Jsou největším globálním zdrojem biogenního uhličitanu vápenatého a významně přispívají ke globálnímu uhlíkovému cyklu.[421] Jsou hlavní složkou křídy, jako je bílé útesy Doveru. | ||||

Propracovaná minerální kostra radiolarianu vyrobená z oxidu křemičitého.

Diatomy, hlavní složky mořského planktonu, mají také tzv. Křemíkové kostry frustule.

Coccolithophores nechat si vyrobit talíře nebo váhy vyrobené z uhličitanu vápenatého kokcithové





Rozsivka mikrofosilní před 40 miliony let

Křemelina je měkký, křemičitý, sedimentární hornina složený z mikrofosilií ve formě frustule (skořápky) jedné buňky rozsivky (kliknutím zvětšíte).

Ilustrace a Globigerina sliz

Mušle (testy ), obvykle vyrobený z uhličitanu vápenatého, z a foraminiferal bláto na dně hlubokého oceánu




Biogeochemické cykly

Oceány jako celek tvoří jeden mořský systém, kde voda - „univerzální rozpouštědlo“ [422] - rozpouští živiny a látky obsahující prvky jako kyslík, uhlík, dusík a fosfor. Tyto látky jsou nekonečně cyklovány a recyklovány, chemicky kombinovány a poté znovu rozloženy, rozpuštěny a poté vysráženy nebo odpařeny, dovezeny a vyvezeny zpět do země a atmosféry a oceánského dna. Powered both by the biological activity of marine organisms and by the natural actions of the sun and tides and movements within the Earth's crust, these are the marine biogeochemical cycles.[423][424]

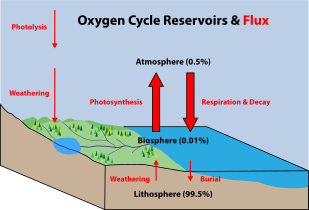
 Marine fosforový cyklus
Marine fosforový cyklus

Interakce pozemků

Land interactions impact marine life in many ways. Coastlines typically have kontinentální police extending some way from the shore. These provide extensive shallows sunlit down to the seafloor, allowing for photosynthesis and enabling habitats for seagrass meadows, coral reefs, kelp forests and other benthic life. Further from shore the continental shelf slopes towards deep water. Vítr blowing at the ocean surface nebo deep ocean currents can result in cold and nutrient rich waters from hlubin hlubin pohybující se nahoru kontinentální svahy. To může mít za následek upwellings along the outer edges of continental shelves, providing conditions for phytoplankton blooms.
Water evaporated by the sun from the surface of the ocean can precipitate on land and eventually return to the ocean as odtok or discharge from rivers, enriched with nutrients as well as znečišťující látky. As rivers discharge into ústí řek, sladkovodní se mísí s slaná voda a stává se brakický. This provides another shallow water habitat where mangrovové lesy a ústí ryb thrive. Overall, life in inland lakes can evolve with greater diversity than happens in the sea, because freshwater habitats are themselves diverse and compartmentalised in a way marine habitats are not. Some aquatic life, such as losos a úhoři, migrovat back and forth between freshwater and marine habitats. These migrations can result in exchanges of pathogens and have impacts on the way life evolves in the ocean.
Antropogenní dopady
Human activities affect marine life and mořská stanoviště přes nadměrný rybolov, znečištění, okyselení a zavedení invazivní druhy. Tyto dopady mořské ekosystémy a potravinářské weby a může mít za následek dosud nerozpoznané následky pro biologická rozmanitost a pokračování mořských forem života.[427]
Události týkající se biologické rozmanitosti a zániku

Biodiverzita is the result of over three billion years of vývoj. Until approximately 600 million years ago, all life consisted of archaea, bakterie, prvoky a podobné jednobuněčné organismy. Dějiny biologické rozmanitosti během EU Phanerozoic (posledních 540 milionů let), začíná rychlým růstem během Kambrijská exploze – a period during which nearly every kmen z mnohobuněčné organismy se poprvé objevil. Over the next 400 million years or so, invertebrate diversity showed little overall trend and vertebrate diversity shows an overall exponential trend.[429]
However, more than 99 percent of all species that ever lived on Earth, amounting to over five billion species,[430] se odhadují na vyhynulý.[431][432] These extinctions occur at an uneven rate. The dramatic rise in diversity has been marked by periodic, massive losses of diversity classified as události hromadného vyhynutí.[429] Mass extinction events occur when life undergoes precipitous global declines. Most diversity and biomasa on earth is found among the mikroorganismy, which are difficult to measure. Recorded extinction events are therefore based on the more easily observed changes in the diversity and abundance of larger mnohobuněčné organismy, rather than the total diversity and abundance of life.[433] Marine fossils are mostly used to measure extinction rates because of their superior fossil record and stratigrafické range compared to land organisms.
Založeno na fosilní záznam, background rate of extinctions on Earth is about two to five taxonomické rodiny of marine animals every million years. The Skvělá událost okysličování was perhaps the first major extinction event. Protože Kambrijská exploze five further major mass extinctions have significantly exceeded the background extinction rate.[434] Nejhorší bylo Událost vyhynutí permu a triasu, Před 251 miliony let. Obratlovcům trvalo 30 milionů let, než se z této události vzpamatovali.[435] In addition to these major mass extinctions there are numerous minor ones, as well as the current ongoing mass-extinction caused by human activity, the Vyhynutí holocénu sometimes called the "sixth extinction".
mořská biologie

During the sixth century BC, the Greek philosopher Xenofany (570-475 BC) recognised that some fosilní shells were remains of shellfish. He used this to argue that what was at the time dry land was once under the sea.[436] This was an important step in advancing from simply stating an idea to backing it with evidence and observation.[437]
Later, during the fourth century BC, another Greek philosopher Aristoteles (384–322 BC) attempted a comprehensive klasifikace of animals which included systematic descriptions of many marine species,[438][439] and particularly species found in the Mediterranean Sea.[440] These pioneering works include Dějiny zvířat, obecná biologie zvířat, Části zvířat, srovnávací anatomie a fyziologie zvířat a Generace zvířat, o vývojové biologii. The most striking passages are about the sea-life visible from observation on Lesbos and available from the catches of fishermen. Jeho pozorování sumec, elektrické ryby (Torpédo ) and angler-fish are detailed, as is his writing on hlavonožci, jmenovitě Chobotnice, Sépie (sépie obecná ) and the paper nautilus (Argonauta argo ). Jeho popis hectocotyl arm, used in sexual reproduction, was widely disbelieved until its rediscovery in the 19th century. He separated aquatic mammals from fish, and knew that sharks and rays were part of a group he called Selachē (selachians ).[441] He gave accurate descriptions of the ovoviviparous embryological development of the hound shark Mustelus mustelus.[442] His classification of living things contains elements which were still in use in the 19th century. What the modern zoologist would call vertebrates and invertebrates, Aristotle called "animals with blood" and "animals without blood" (he did not know that complex invertebrates do make use of hemoglobin, but of a different kind from vertebrates). He divided animals with blood into live-bearing (mammals), and egg-bearing (birds and fish). Invertebrates ("animals without blood") he divided into insects, crustacea (further divided into non-shelled – cephalopods – and shelled) and testacea (molluscs).[443][444]
In contemporary times, marine life is a field of study both in mořská biologie a v biologická oceánografie. In biology many phyla, families and genera have some species that live in the sea and others that live on land. Marine biology classifies species based on the environment rather than on taxonomy. For this reason marine biology encompasses not only organisms that live only in a marine environment, but also other organisms whose lives revolve around the sea. Biological oceanography is the study of how organisms affect and are affected by the fyzika, chemie, a geologie z oceanographic system. Biological oceanography mostly focuses on the mikroorganismy within the ocean; looking at how they are affected by their environment and how that affects larger marine creatures and their ecosystem.[445] Biological oceanography is similar to mořská biologie, but it studies ocean life from a different perspective. Biological oceanography takes a bottom up approach in terms of the food web, while marine biology studies the ocean from a top down perspective. Biological oceanography mainly focuses on the ecosystem of the ocean with an emphasis on plankton: their diversity (morphology, nutritional sources, motility, and metabolism); their productivity and how that plays a role in the global carbon cycle; and their distribution (predation and life cycle).[445][446][447] Biological oceanography also investigates the role of microbes in food webs, and how humans impact the ecosystems in the oceans.[445]
Viz také
- Modrá planeta – British nature documentary series - David Attenborough
- Sčítání mořského života
- Kolonizace půdy
- Taxonomy of invertebrates – System of classification of animals with emphasis on the invertebrates
Poznámky
- ^ This is the measurement taken by the vessel Kaiko in March 1995 and is considered the most accurate measurement to date. Viz Challenger Deep článek pro více informací.
- ^ Myxozoa were thought to be an exception, but are now thought to be heavily modified members of the Cnidaria. Jímenez-Guri, Eva; Philippe, Hervé; Okamura, Beth; Holland, Peter W. H. (6 July 2007). "Buddenbrockia Is a Cnidarian Worm". Věda. 317 (5834): 116–118. Bibcode:2007Sci...317..116J. doi:10.1126/science.1142024. ISSN 0036-8075. PMID 17615357. S2CID 5170702.
Reference
- ^ A b Cavicchioli, Ricardo; Ripple, William J .; Timmis, Kenneth N .; Azam, Farooq; Bakken, Lars R.; Baylis, Matthew; Behrenfeld, Michael J.; Boetius, Antje; Boyd, Philip W .; Classen, Aimée T.; Crowther, Thomas W.; Danovaro, Roberto; Foreman, Christine M.; Huisman, Jef; Hutchins, David A.; Jansson, Janet K .; Karl, David M .; Koskella, Britt; Mark Welch, David B.; Martiny, Jennifer B. H.; Moran, Mary Ann; Orphan, Victoria J .; Reay, David S.; Remais, Justin V.; Rich, Virginie I .; Singh, Brajesh K.; Stein, Lisa Y .; Stewart, Frank J.; Sullivan, Matthew B.; et al. (2019). "Scientists' warning to humanity: Microorganisms and climate change". Příroda Recenze Mikrobiologie. 17 (9): 569–586. doi:10.1038 / s41579-019-0222-5. PMC 7136171. PMID 31213707.
 Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0. - ^ A b C "National Oceanic and Atmospheric Administration – Ocean". NOAA. Citováno 20. února 2019.
- ^ Tiny Fish May Be Ancestor of Nearly All Living Vertebrates Živá věda, 11 June 2014.
- ^ Drogin, Bob (2 August 2009). "Mapping an ocean of species". Los Angeles Times. Citováno 18. srpna 2009.
- ^ Paul, Gregory S. (2010). "The Evolution of Dinosaurs and their World". The Princeton Field Guide to Dinosaurs. Princeton: Princeton University Press. str.19.
- ^ Bortolotti, Dan (2008). Wild Blue: Přirozená historie největšího zvířete na světě. Svatomartinský tisk.
- ^ A b C Bar-On, YM; Phillips, R; Milo, R (2018). „Distribuce biomasy na Zemi“. PNAS. 115 (25): 6506–6511. doi:10.1073 / pnas.1711842115. PMC 6016768. PMID 29784790.
- ^ Census Of Marine Life Přístupné 29. října 2020.
- ^ Xiao-feng, Pang (2014) Water: Molecular Structure And Properties, chapter 5, pp. 390–461, World Scientific. ISBN 9789814440448
- ^ Greenwood, Norman N.; Earnshaw, Alan (1997). Chemie prvků (2. vyd.). Butterworth-Heinemann. str. 620. ISBN 978-0-08-037941-8.
- ^ "Water, the Universal Solvent". USGS. Archivováno z původního dne 9. července 2017. Citováno 27. června 2017.
- ^ Reece, Jane B. (31 October 2013). Campbell Biology (10. vyd.). Pearson. str. 48. ISBN 9780321775658.
- ^ Reece, Jane B. (31 October 2013). Campbell Biology (10. vyd.). Pearson. str. 44. ISBN 9780321775658.
- ^ Collins J. C. (1991) The Matrix of Life: A View of Natural Molecules from the Perspective of Environmental Water Molecular Presentations. ISBN 9780962971907.
- ^ "7,000 m Class Remotely Operated Vehicle KAIKO 7000". Japonská agentura pro vědu a technologii mořské Země (JAMSTEC). Citováno 7. června 2008.
- ^ How many oceans are there? NOAA. Updated: 9 April 2020.
- ^ Charette, Matthew A .; Smith, Walter H. F. (June 2010). "The Volume of Earth's Ocean" (PDF). Oceánografie. 23 (2): 112–14. doi:10.5670/oceanog.2010.51. Archivovány od originál (PDF) dne 2. srpna 2013. Citováno 6. června 2013.
- ^ "sphere depth of the ocean – hydrology". Encyklopedie Britannica. Citováno 12. dubna 2015.
- ^ "Third rock from the Sun – restless Earth". Kosmos NASA. Citováno 12. dubna 2015.
- ^ Perlman, Howard (17 March 2014). "The World's Water". USGS Water-Science School. Citováno 12. dubna 2015.
- ^ Kennish, Michael J. (2001). Praktická příručka námořní vědy. Řada mořských věd (3. vydání). CRC Press. str. 35. ISBN 978-0-8493-2391-1.
- ^ „Proč je oceán slaný?“.
- ^ Mullen, Leslie (11 June 2002). "Salt of the Early Earth". NASA Astrobiology Magazine. Archivovány od originál dne 22. července 2007. Citováno 14. března 2007.
- ^ Morris, Ron M. "Oceanic Processes". NASA Astrobiology Magazine. Archivovány od originál dne 15. dubna 2009. Citováno 14. března 2007.
- ^ Scott, Michon (24 April 2006). "Earth's Big heat Bucket". NASA Earth Observatory. Citováno 14. března 2007.
- ^ Sample, Sharron (21 June 2005). "Sea Surface Temperature". NASA. Archivovány od originál dne 3. dubna 2013. Citováno 21. dubna 2007.
- ^ "Volumes of the World's Oceans from ETOPO1". NOAA. Archived from the original on 11 March 2015. Citováno 20. února 2019.CS1 maint: BOT: stav původní adresy URL neznámý (odkaz)
- ^ Planeta „Země“: Měli jsme ji nazvat „mořskou“ Invertigátor nabídky, 25 January 2017.
- ^ Odhalení Planet Ocean Věda NASA, 14. března 2002.
- ^ Dyches, Preston; Brown, Dwayne (12 May 2015). "NASA Research Reveals Europa's Mystery Dark Material Could Be Sea Salt". NASA. Citováno 12. května 2015.
- ^ "Water near surface of a Jupiter moon only temporary".
- ^ Tritt, Charles S. (2002). "Possibility of Life on Europa". Milwaukee School of Engineering. Archivovány od originál dne 9. června 2007. Citováno 10. srpna 2007.
- ^ Schulze-Makuch, Dirk; Irwin, Louis N. (2001). "Alternative Energy Sources Could Support Life on Europa" (PDF). Departments of Geological and Biological Sciences, University of Texas v El Pasu. Archivovány od originál (PDF) dne 3. července 2006. Citováno 21. prosince 2007.
- ^ Friedman, Louis (14 December 2005). "Projects: Europa Mission Campaign". Planetární společnost. Archivovány od originál dne 11. srpna 2011. Citováno 8. srpna 2011.
- ^ Ocean Within Enceladus May Harbor Hydrothermal Activity, NASA Press Release. 11. března 2015.
- ^ "Age of the Earth". Geologický průzkum Spojených států. 9. července 2007. Citováno 31. května 2015.
- ^ Dalrymple 2001, str. 205–221
- ^ Manhesa, Gérard; Allègre, Claude J.; Dupréa, Bernard; Hamelin, Bruno (May 1980). "Lead isotope study of basic-ultrabasic layered complexes: Speculations about the age of the earth and primitive mantle characteristics". Dopisy o Zemi a planetách. 47 (3): 370–382. Bibcode:1980E&PSL..47..370M. doi:10.1016/0012-821X(80)90024-2. ISSN 0012-821X.
- ^ Schopf, J. William; Kudryavtsev, Anatolij B .; Czaja, Andrew D .; Tripathi, Abhishek B. (5. října 2007). "Důkazy o archeanském životě: stromatolity a mikrofosílie". Prekambrický výzkum. 158 (3–4): 141–155. Bibcode:2007PreR..158..141S. doi:10.1016 / j.precamres.2007.04.009. ISSN 0301-9268.
- ^ Raven & Johnson 2002, str. 68
- ^ Baumgartner, R. J.; et al. (2019). "Nano−porous pyrite and organic matter in 3.5-billion-year-old stromatolites record primordial life" (PDF). Geologie. 47 (11): 1039–1043. Bibcode:2019Geo....47.1039B. doi:10.1130/G46365.1.
- ^ Earliest signs of life: Scientists find microbial remains in ancient rocks Phys.org. 26. září 2019.
- ^ Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; et al. (Leden 2014). „Důkazy o biogenním grafitu v časných archaeanských metasedimentárních horninách Isua“. Nature Geoscience. 7 (1): 25–28. Bibcode:2014NatGe ... 7 ... 25O. doi:10.1038 / ngeo2025. ISSN 1752-0894.
- ^ A b Borenstein, Seth (19. října 2015). „Náznaky života na tom, co bylo považováno za pustou ranou Zemi“. Associated Press. Citováno 9. října 2018.
- ^ Bell, Elizabeth A .; Boehnike, Patrick; Harrison, T. Mark; et al. (24 November 2015). „Potenciálně biogenní uhlík konzervovaný v 4,1 miliard let starém zirkonu“ (PDF). Sborník Národní akademie věd Spojených států amerických. 112 (47): 14518–14521. Bibcode:2015PNAS..11214518B. doi:10.1073 / pnas.1517557112. ISSN 0027-8424. PMC 4664351. PMID 26483481. Citováno 30. prosince 2015.
- ^ Penny, David; Poole, Anthony (December 1999). "The nature of the last universal common ancestor". Aktuální názor na genetiku a vývoj. 9 (6): 672–677. doi:10.1016/S0959-437X(99)00020-9. ISSN 0959-437X. PMID 10607605.
- ^ Theobald, Douglas L. (13 May 2010). "A formal test of the theory of universal common ancestry". Příroda. 465 (7295): 219–222. Bibcode:2010Natur.465..219T. doi:10.1038/nature09014. ISSN 0028-0836. PMID 20463738. S2CID 4422345.
- ^ Doolittle, W. Ford (Únor 2000). "Uprooting the Tree of Life" (PDF). Scientific American. 282 (2): 90–95. Bibcode:2000SciAm.282b..90D. doi:10.1038 / scientificamerican0200-90. ISSN 0036-8733. PMID 10710791. Archivovány od originál (PDF) dne 7. září 2006. Citováno 5. dubna 2015.
- ^ Peretó, Juli (March 2005). "Controversies on the origin of life" (PDF). Mezinárodní mikrobiologie. 8 (1): 23–31. ISSN 1139-6709. PMID 15906258. Archivovány od originál (PDF) dne 24. srpna 2015.
- ^ Joyce, Gerald F. (11 July 2002). "The antiquity of RNA-based evolution". Příroda. 418 (6894): 214–221. Bibcode:2002Natur.418..214J. doi:10.1038/418214a. ISSN 0028-0836. PMID 12110897. S2CID 4331004.
- ^ Trevors, Jack T.; Psenner, Roland (December 2001). "From self-assembly of life to present-day bacteria: a possible role for nanocells". Recenze mikrobiologie FEMS. 25 (5): 573–582. doi:10.1111/j.1574-6976.2001.tb00592.x. ISSN 1574-6976. PMID 11742692.
- ^ Wade, Nicholas (25. července 2016). „Seznamte se s Lucem, předkem všeho živého“. New York Times. Citováno 25. července 2016.
- ^ Bapteste, Eric; Walsh, David A. (June 2005). "Does the 'Ring of Life' ring true?". Trendy v mikrobiologii. 13 (6): 256–261. doi:10.1016/j.tim.2005.03.012. ISSN 0966-842X. PMID 15936656.
- ^ Darwin 1859, str.1
- ^ Doolittle, W. Ford; Bapteste, Eric (13 February 2007). "Pattern pluralism and the Tree of Life hypothesis". Sborník Národní akademie věd Spojených států amerických. 104 (7): 2043–2049. Bibcode:2007PNAS..104.2043D. doi:10.1073/pnas.0610699104. ISSN 0027-8424. PMC 1892968. PMID 17261804.
- ^ Kunin, Victor; Goldovsky, Leon; Darzentas, Nikos; Ouzounis, Christos A. (July 2005). "The net of life: Reconstructing the microbial phylogenetic network". Výzkum genomu. 15 (7): 954–959. doi:10.1101/gr.3666505. ISSN 1088-9051. PMC 1172039. PMID 15965028.
- ^ Jablonski, David (25 June 1999). "The Future of the Fossil Record". Věda. 284 (5423): 2114–2116. doi:10.1126/science.284.5423.2114. ISSN 0036-8075. PMID 10381868. S2CID 43388925.
- ^ Ciccarelli, Francesca D.; Doerks, Tobias; von Mering, Christian; et al. (3. března 2006). „Směrem k automatické rekonstrukci vysoce vyřešeného stromu života“. Věda. 311 (5765): 1283–1287. Bibcode:2006Sci ... 311.1283C. CiteSeerX 10.1.1.381.9514. doi:10.1126 / science.1123061. ISSN 0036-8075. PMID 16513982. S2CID 1615592.
- ^ Mason, Stephen F. (6 September 1984). "Origins of biomolecular handedness". Příroda. 311 (5981): 19–23. Bibcode:1984Natur.311...19M. doi:10.1038/311019a0. ISSN 0028-0836. PMID 6472461. S2CID 103653.
- ^ Vlk, Jurij I.; Rogozin, Igor B.; Grishin, Nick V .; Koonin, Eugene V. (1. září 2002). "Genome trees and the tree of life". Trendy v genetice. 18 (9): 472–479. doi:10.1016/S0168-9525(02)02744-0. ISSN 0168-9525. PMID 12175808.
- ^ Varki, Ajit; Altheide, Tasha K. (December 2005). "Comparing the human and chimpanzee genomes: searching for needles in a haystack" (PDF). Výzkum genomu. 15 (12): 1746–1758. doi:10.1101/gr.3737405. ISSN 1088-9051. PMID 16339373.
- ^ A b Cavalier-Smith, Thomas (29. června 2006). "Cell evolution and Earth history: stasis and revolution". Filozofické transakce Královské společnosti B: Biologické vědy. 361 (1470): 969–1006. doi:10.1098/rstb.2006.1842. ISSN 0962-8436. PMC 1578732. PMID 16754610.
- ^ Schopf, J. William (29. června 2006). „Fosilní důkazy o archaejském životě“. Filozofické transakce Královské společnosti B: Biologické vědy. 361 (1470): 869–885. doi:10.1098 / rstb.2006.1834. ISSN 0962-8436. PMC 1578735. PMID 16754604.
- ^ Schopf, J. William (19 July 1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Sborník Národní akademie věd Spojených států amerických. 91 (15): 6735–6742. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. ISSN 0027-8424. PMC 44277. PMID 8041691.
- ^ Poole, Anthony M .; Penny, David (January 2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. ISSN 0265-9247. PMID 17187354.
- ^ A b Dyall, Sabrina D.; Brown, Mark T.; Johnson, Patricia J. (9. dubna 2004). "Ancient Invasions: From Endosymbionts to Organelles". Věda. 304 (5668): 253–257. Bibcode:2004Sci...304..253D. doi:10.1126/science.1094884. ISSN 0036-8075. PMID 15073369. S2CID 19424594.
- ^ Martin, William (October 2005). "The missing link between hydrogenosomes and mitochondria". Trendy v mikrobiologii. 13 (10): 457–459. doi:10.1016/j.tim.2005.08.005. ISSN 0966-842X. PMID 16109488.
- ^ Lang, B. Franz; Gray, Michael W.; Burger, Gertraud (December 1999). "Mitochondrial genome evolution and the origin of eukaryotes". Výroční přehled genetiky. 33: 351–397. doi:10.1146/annurev.genet.33.1.351. ISSN 0066-4197. PMID 10690412.
- ^ DeLong, Edward F.; Pace, Norman R. (1. srpna 2001). "Environmental Diversity of Bacteria and Archaea". Systematická biologie. 50 (4): 470–478. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. ISSN 1063-5157. PMID 12116647.
- ^ Kaiser, Dale (Prosinec 2001). "Building a multicellular organism". Výroční přehled genetiky. 35: 103–123. doi:10.1146/annurev.genet.35.102401.090145. ISSN 0066-4197. PMID 11700279. S2CID 18276422.
- ^ Zimmer, Carle (7. ledna 2016). „Genetické převrácení pomohlo organizmům přejít z jedné buňky do mnoha“. The New York Times. Citováno 7. ledna 2016.
- ^ Valentine, James W.; Jablonski, David; Erwin, Douglas H. (1 March 1999). "Fossils, molecules and embryos: new perspectives on the Cambrian explosion". Rozvoj. 126 (5): 851–859. ISSN 0950-1991. PMID 9927587. Citováno 30. prosince 2014.
- ^ Ohno, Susumu (January 1997). "The reason for as well as the consequence of the Cambrian explosion in animal evolution". Journal of Molecular Evolution. 44 (Suppl. 1): S23–S27. Bibcode:1997JMolE..44S..23O. doi:10.1007/PL00000055. ISSN 0022-2844. PMID 9071008. S2CID 21879320.
- Valentine, James W .; Jablonski, David (2003). "Morphological and developmental macroevolution: a paleontological perspective". International Journal of Developmental Biology. 47 (7–8): 517–522. ISSN 0214-6282. PMID 14756327. Citováno 30. prosince 2014.
- ^ Wellman, Charles H .; Osterloff, Peter L.; Mohiuddin, Uzma (2003). „Fragmenty prvních suchozemských rostlin“ (PDF). Příroda. 425 (6955): 282–285. Bibcode:2003 Natur.425..282W. doi:10.1038 / nature01884. PMID 13679913. S2CID 4383813.
- ^ Barton, Nicholas (2007). Vývoj. str. 273–274. ISBN 9780199226320. Citováno 30. září 2012.
- ^ Waters, Elizabeth R. (December 2003). "Molecular adaptation and the origin of land plants". Molekulární fylogenetika a evoluce. 29 (3): 456–463. doi:10.1016/j.ympev.2003.07.018. ISSN 1055-7903. PMID 14615186.
- ^ Mayhew, Peter J. (Srpen 2007). „Proč je tolik druhů hmyzu? Perspektivy z fosilií a fylogenezí“. Biologické recenze. 82 (3): 425–454. doi:10.1111 / j.1469-185X.2007.00018.x. ISSN 1464-7931. PMID 17624962. S2CID 9356614.
- ^ Carroll, Robert L. (Květen 2007). "The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians". Zoologický žurnál Linneanské společnosti. 150 (Supplement s1): 1–140. doi:10.1111/j.1096-3642.2007.00246.x. ISSN 1096-3642.
- ^ Wible, John R .; Rougier, Guillermo W .; Novacek, Michael J .; Asher, Robert J. (21 June 2007). "Křídové eutherians a Laurasian původ pro placentární savce v blízkosti hranice K / T". Příroda. 447 (7147): 1003–1006. Bibcode:2007Natur.447.1003W. doi:10.1038 / nature05854. ISSN 0028-0836. PMID 17581585. S2CID 4334424.
- ^ Witmer, Lawrence M. (28. července 2011). "Palaeontology: An icon knocked from its perch". Příroda. 475 (7357): 458–459. doi:10.1038/475458a. ISSN 0028-0836. PMID 21796198. S2CID 205066360.
- ^ Schloss, Patrick D.; Handelsman, Jo (Prosinec 2004). "Status of the Microbial Census". Recenze mikrobiologie a molekulární biologie. 68 (4): 686–691. doi:10.1128/MMBR.68.4.686-691.2004. ISSN 1092-2172. PMC 539005. PMID 15590780.
- ^ Miller & Spoolman 2012, str.62
- ^ Mora, Camilo; Tittensor, Derek P .; Adl, Sina; et al. (23. srpna 2011). „Kolik druhů je na Zemi a v oceánu?“. PLOS Biology. 9 (8): e1001127. doi:10.1371 / journal.pbio.1001127. ISSN 1545-7885. PMC 3160336. PMID 21886479.
- ^ Madigan M; Martinko J, vyd. (2006). Brockova biologie mikroorganismů (13. vydání). Pearson Education. str. 1096. ISBN 978-0-321-73551-5.
- ^ Rybicki EP (1990). „Klasifikace organismů na pokraji života nebo problémy se systematikou virů“. Jihoafrický žurnál vědy. 86: 182–6. ISSN 0038-2353.
- ^ Lwoff A (1956). "The concept of virus". Journal of General Microbiology. 17 (2): 239–53. doi:10.1099/00221287-17-2-239. PMID 13481308.
- ^ 2002 WHO mortality data Accessed 20 January 2007
- ^ "Functions of global ocean microbiome key to understanding environmental changes". www.sciencedaily.com. University of Georgia. 10. prosince 2015. Citováno 11. prosince 2015.
- ^ Suttle, C.A. (2005). "Viruses in the Sea". Příroda. 437 (9): 356–361. Bibcode:2005 Natur.437..356S. doi:10.1038 / nature04160. PMID 16163346. S2CID 4370363.
- ^ Shors p. 5
- ^ Shors p. 593
- ^ A b C Suttle CA. Marine viruses—major players in the global ecosystem. Příroda Recenze Mikrobiologie. 2007;5(10):801–12. doi:10.1038 / nrmicro1750. PMID 17853907.
- ^ Živé bakterie jezdí na vzdušných proudech Země Smithsonian Magazine, 11. ledna 2016.
- ^ Robbins, Jim (13. dubna 2018). „Biliony za biliony virů každý den padají z nebe“. The New York Times. Citováno 14. dubna 2018.
- ^ Reche, Isabel; D’Orta, Gaetano; Mladenov, Natalie; Winget, Danielle M; Suttle, Curtis A (29. ledna 2018). „Míra ukládání virů a bakterií nad mezní vrstvou atmosféry“. ISME Journal. 12 (4): 1154–1162. doi:10.1038 / s41396-017-0042-4. PMC 5864199. PMID 29379178.
- ^ Zaměstnanci (2014). "The Biosphere". Aspen Global Change Institute. Citováno 10. listopadu 2014.
- ^ A b C Choi, Charles Q. (17. března 2013). „Mikrobům se daří v nejhlubším místě na Zemi“. LiveScience. Citováno 17. března 2013.
- ^ Glud, Ronnie; Wenzhöfer, Frank; Middelboe, Mathias; Oguri, Kazumasa; Turnewitsch, Robert; Canfield, Donald E .; Kitazato, Hiroshi (17 March 2013). "High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth". Nature Geoscience. 6 (4): 284–288. Bibcode:2013NatGe...6..284G. doi:10.1038/ngeo1773.
- ^ Oskin, Becky (14. března 2013). „Intraterrestrials: Life Thrives in Ocean Floor“. LiveScience. Citováno 17. března 2013.
- ^ Morelle, Rebecca (15 December 2014). "Microbes discovered by deepest marine drill analysed". BBC novinky. Citováno 15. prosince 2014.
- ^ Takai K; Nakamura K; Toki T; Tsunogai U; et al. (2008). „Proliferace buněk při 122 ° C a izotopově těžká produkce CH4 hypertermofilním methanogenem při vysokotlaké kultivaci“. Sborník Národní akademie věd Spojených států amerických. 105 (31): 10949–54. Bibcode:2008PNAS..10510949T. doi:10.1073 / pnas.0712334105. PMC 2490668. PMID 18664583.
- ^ Fox, Douglas (20. srpna 2014). „Jezera pod ledem: tajná zahrada Antarktidy“. Příroda. 512 (7514): 244–246. Bibcode:2014 Natur.512..244F. doi:10.1038 / 512244a. PMID 25143097.
- ^ Mack, Eric (20. srpna 2014). „Život potvrzen pod antarktickým ledem; je vesmír další?“. Forbes. Citováno 21. srpna 2014.
- ^ Wimmer E, Mueller S, Tumpey TM, Taubenberger JK. Syntetické viry: nová příležitost pochopit a předcházet virovým onemocněním. Přírodní biotechnologie. 2009;27(12):1163–72. doi:10,1038 / nbt.1593. PMID 20010599.
- ^ Koonin EV, Senkevich TG, Dolja VV. Starověký virový svět a vývoj buněk. Biology Direct. 2006;1:29. doi:10.1186/1745-6150-1-29. PMID 16984643.
- ^ Collier, Leslie; Balows, Albert; Sussman, Max (1998) Topley a Wilsonova mikrobiologie a mikrobiální infekce deváté vydání, svazek 1, Virologie, editoři svazku: Mahy, Brian a Collier, Leslie. Arnold. Stránky 33–37. ISBN 0-340-66316-2.
- ^ Iyer LM, Balaji S, Koonin EV, Aravind L. Evoluční genomika nukleo-cytoplazmatických velkých DNA virů. Virový výzkum. 2006;117(1):156–84. doi:10.1016 / j.virusres.2006.01.009. PMID 16494962.
- ^ Sanjuán R, Nebot MR, Chirico N, Mansky LM, Belshaw R. Míry virových mutací. Journal of Virology. 2010;84(19):9733–48. doi:10.1128 / JVI.00694-10. PMID 20660197.
- ^ In: Mahy WJ, Van Regenmortel MHV. Desk Encyclopedia of General Virology. Oxford: Academic Press; 2009. ISBN 0-12-375146-2. str. 28.
- ^ A b Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H. Fág jako agenti laterálního přenosu genů. Současný názor v mikrobiologii. 2003;6(4):417–24. doi:10.1016 / S1369-5274 (03) 00086-9. PMID 12941415.
- ^ Jsou viry naživu? Paradigma replikátoru vrhá rozhodující světlo na starou, ale zavádějící otázku. Studium v historii a filozofii biologických a biomedicínských věd. 2016. doi:10.1016 / j.shpsc.2016.02.016. PMID 26965225.
- ^ Koonin, E. V .; Starokadomskyy, P. (7. března 2016). „Jsou viry naživu? Paradigma replikátoru vrhá rozhodující světlo na starou, ale zavádějící otázku“. Studium v historii a filozofii biologických a biomedicínských věd. 59: 125–34. doi:10.1016 / j.shpsc.2016.02.016. PMC 5406846. PMID 26965225.
- ^ Rybicki, EP. Klasifikace organismů na pokraji života nebo problémy se systematikou virů. Jihoafrický žurnál vědy. 1990;86:182–186.
- ^ A b Mann, NH (17. května 2005). „Třetí věk fága“. PLOS Biology. 3 (5): 753–755. doi:10.1371 / journal.pbio.0030182. PMC 1110918. PMID 15884981.
- ^ Wommack KE, Colwell RR. Virioplankton: viry ve vodních ekosystémech. Recenze mikrobiologie a molekulární biologie. 2000;64(1):69–114. doi:10.1128 / MMBR.64.1.69-114.2000. PMID 10704475.
- ^ Suttle CA. Viry v moři. Příroda. 2005;437:356–361. doi:10.1038 / nature04160. PMID 16163346. Bibcode:2005 Natur.437..356S.
- ^ Bergh O, Børsheim KY, Bratbak G, Heldal M. Vysoký výskyt virů ve vodním prostředí. Příroda. 1989;340(6233):467–8. doi:10.1038 / 340467a0. PMID 2755508. Bibcode:1989 Natur.340..467B.
- ^ Wigington CH, Sonderegger D, Brussaard CPD, Buchan A, Finke JF, Fuhrman JA, Lennon JT, Middelboe M, Suttle CA, Stock C, Wilson WH, Wommack KE, Wilhelm SW, Weitz JS. Znovu přezkoumání vztahu mezi mořským virem a množstvím mikrobiálních buněk. Přírodní mikrobiologie. 2016;1:15024. doi:10.1038 / nmicrobiol.2015.24. PMID 27572161.
- ^ Krupovic M, Bamford DH (2007). „Předpokládané profágy související s lytickým bezocasým mořským dsDNA fágem PM2 jsou rozšířené v genomech vodních bakterií“. BMC Genomics. 8: 236. doi:10.1186/1471-2164-8-236. PMC 1950889. PMID 17634101.
- ^ Xue H, Xu Y, Boucher Y, Polz MF (2012). „Vysoká frekvence nového filamentózního fága, VCY φ, v populaci prostředí Vibrio cholerae“. Aplikovaná a environmentální mikrobiologie. 78 (1): 28–33. doi:10.1128 / AEM.06297-11. PMC 3255608. PMID 22020507.
- ^ Roux S, Krupovic M, Poulet A, Debroas D, Enault F (2012). „Evoluce a rozmanitost virové rodiny Microviridae prostřednictvím sbírky 81 nových kompletních genomů shromážděných z čtení viromu“. PLOS ONE. 7 (7): e40418. Bibcode:2012PLoSO ... 740418R. doi:10.1371 / journal.pone.0040418. PMC 3394797. PMID 22808158.
- ^ Suttle CA. Viry v moři. Příroda. 2005;437(7057):356–61. doi:10.1038 / nature04160. PMID 16163346. Bibcode:2005 Natur.437..356S.
- ^ www.cdc.gov. Škodlivé květy řas: Červený příliv: Domů [Citováno 2014-12-19].
- ^ Lawrence CM, Menon S, Eilers BJ, et al.. Strukturální a funkční studie archaálních virů. Journal of Biological Chemistry. 2009;284(19):12599–603. doi:10,1074 / jbc.R800078200. PMID 19158076.
- ^ Prangishvili D, Forterre P, Garrett RA. Viry Archea: sjednocující pohled. Příroda Recenze Mikrobiologie. 2006;4(11):837–48. doi:10.1038 / nrmicro1527. PMID 17041631.
- ^ Prangishvili D, Garrett RA. Výjimečně rozmanité morfotypy a genomy crenarcheálních hypertermofilních virů. Transakce biochemické společnosti. 2004; 32 (Pt 2): 204–8. doi:10.1042 / BST0320204. PMID 15046572.
- ^ Forterre P, Philippe H. Poslední univerzální společný předek (LUCA), jednoduchý nebo komplexní ?. Biologický bulletin. 1999; 196 (3): 373–5; diskuse 375–7. doi:10.2307/1542973. PMID 11536914.
- ^ Fredrickson JK, Zachara JM, Balkwill DL, Kennedy D, Li SM, Kostandarithes HM, Daly MJ, Romine MF, Brockman FJ (2004). „Geomikrobiologie vysoce vadózních sedimentů kontaminovaných jaderným odpadem v lokalitě Hanford ve státě Washington“. Aplikovaná a environmentální mikrobiologie. 70 (7): 4230–41. doi:10.1128 / AEM.70.7.4230-4241.2004. PMC 444790. PMID 15240306.
- ^ Woese CR, Kandler O, Wheelis ML (1990). „Směrem k přirozenému systému organismů: návrh domén Archaea, Bacteria a Eucarya“. Sborník Národní akademie věd Spojených států amerických. 87 (12): 4576–9. Bibcode:1990PNAS ... 87,4576 W.. doi:10.1073 / pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ Schopf JW (1994). „Nesourodé sazby, různé osudy: tempo a způsob vývoje se změnily z prekambria na phanerozoik“. Sborník Národní akademie věd Spojených států amerických. 91 (15): 6735–42. Bibcode:1994PNAS ... 91.6735S. doi:10.1073 / pnas.91.15.6735. PMC 44277. PMID 8041691.
- ^ DeLong EF, Pace NR (2001). „Environmentální rozmanitost bakterií a archea“. Systematická biologie. 50 (4): 470–8. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Brown JR, Doolittle WF (1997). „Archaea a přechod mezi prokaryotem a eukaryotem“. Recenze mikrobiologie a molekulární biologie. 61 (4): 456–502. doi:10.1128/.61.4.456-502.1997. PMC 232621. PMID 9409149.
- ^ Poole AM, Penny D (2007). "Hodnocení hypotéz o původu eukaryot". BioEssays. 29 (1): 74–84. doi:10.1002 / bies.20516. PMID 17187354.
- ^ Lang BF, Gray MW, Burger G (1999). "Vývoj mitochondriálního genomu a původ eukaryot". Výroční přehled genetiky. 33: 351–97. doi:10.1146 / annurev.genet.33.1.351. PMID 10690412.
- ^ McFadden GI (1999). "Endosymbióza a vývoj rostlinné buňky". Aktuální názor na biologii rostlin. 2 (6): 513–9. doi:10.1016 / S1369-5266 (99) 00025-4. PMID 10607659.
- ^ Patrick J. Keeling (2004). „Rozmanitost a evoluční historie plastidů a jejich hostitelů“. American Journal of Botany. 91 (10): 1481–1493. doi:10.3732 / ajb.91.10.1481. PMID 21652304.
- ^ „Největší bakterie: Vědec objevuje novou formu bakteriálního života u afrického pobřeží“, Max Planck Institute for Marine Microbiology, 8. dubna 1999, archivovány od originál dne 20. ledna 2010
- ^ Seznam prokaryotických jmen se Stálým v nomenklatuře - rod Thiomargarita
- ^ Bang C, Schmitz RA (2015). „Archaea spojená s lidským povrchem: nelze podceňovat“. Recenze mikrobiologie FEMS. 39 (5): 631–48. doi:10.1093 / femsre / fuv010. PMID 25907112.
- ^ Archaea Online slovník etymologie. Vyvolány 17 August je 2016.
- ^ Tempo NR (květen 2006). "Čas na změnu". Příroda. 441 (7091): 289. Bibcode:2006 Natur.441..289P. doi:10.1038 / 441289a. PMID 16710401. S2CID 4431143.
- ^ Stoeckenius W (1. října 1981). „Walsbyho čtvercová bakterie: jemná struktura ortogonálního prokaryota“. Journal of Bacteriology. 148 (1): 352–60. doi:10.1128 / JB.148.1.352-360.1981. PMC 216199. PMID 7287626.
- ^ Whittaker, R.H .; Margulis, L. (1978). "Protist klasifikace a království organismů". Biosystémy. 10 (1–2): 3–18. doi:10.1016/0303-2647(78)90023-0. PMID 418827.
- ^ Faure, E; Ne, F; Benoiston, AS; Labadie, K; Bittner, L; Ayata, SD (2019). „Mixotrofní protisté zobrazují kontrastní biogeografie v globálním oceánu“ (PDF). ISME Journal. 13 (4): 1072–1083. doi:10.1038 / s41396-018-0340-5. PMC 6461780. PMID 30643201.
- ^ A b Leles, S.G .; Mitra, A .; Flynn, K.J .; Stoecker, D.K .; Hansen, P.J .; Calbet, A .; McManus, G.B .; Sanders, R.W .; Caron, D. A.; Ne, F .; Hallegraeff, G.M. (1860). „Oceánští protisté s různými formami získané fototrofie vykazují kontrastní biogeografie a hojnost“. Sborník Královské společnosti B: Biologické vědy. 284 (1860): 20170664. doi:10.1098 / rspb.2017.0664. PMC 5563798. PMID 28768886.
- ^ Budd, Graham E; Jensen, Sören (2017). „Původ zvířat a hypotéza„ Savannah “pro časnou bilaterální evoluci“. Biologické recenze. 92 (1): 446–473. doi:10.1111 / brv.12239. PMID 26588818.
- ^ Cavalier-Smith T (prosinec 1993). „Prvoky království a jejich 18 kmenů“. Mikrobiologické recenze. 57 (4): 953–94. doi:10.1128 / MMBR.57.4.953-994.1993. PMC 372943. PMID 8302218.
- ^ Corliss JO (1992). „Měl by pro protisty existovat samostatný kód nomenklatury?“. BioSystems. 28 (1–3): 1–14. doi:10.1016 / 0303-2647 (92) 90003-H. PMID 1292654.
- ^ Slapeta J, Moreira D, López-García P (2005). „Rozsah protistické rozmanitosti: poznatky z molekulární ekologie sladkovodních eukaryot“. Sborník Královské společnosti B: Biologické vědy. 272 (1576): 2073–81. doi:10.1098 / rspb.2005.3195. PMC 1559898. PMID 16191619.
- ^ Moreira D, López-García P (2002). „Molekulární ekologie mikrobiálních eukaryot odhaluje skrytý svět“ (PDF). Trendy v mikrobiologii. 10 (1): 31–8. doi:10.1016 / S0966-842X (01) 02257-0. PMID 11755083.
- ^ A b Vzduch, který dýcháte? Diatom to udělal
- ^ „Více o Diatoms“. Muzeum paleontologie University of California. Archivovány od originál dne 4. října 2012. Citováno 27. června 2019.
- ^ Devreotes P (1989). „Dictyostelium discoideum: modelový systém pro vývoj buněčných interakcí“. Věda. 245 (4922): 1054–8. Bibcode:1989Sci ... 245.1054D. doi:10.1126 / science.2672337. PMID 2672337.
- ^ Matz, Michail V .; Tamara M. Frank; Justin Marshall; Edith A. Widder; Sonke Johnsen (9. prosince 2008). „Giant Deep-Sea Protist Produces Bilaterian-like Traces“ (PDF). Aktuální biologie. Elsevier Ltd. 18 (23): 1849–1854. doi:10.1016 / j.cub.2008.10.028. PMID 19026540. S2CID 8819675.
- ^ Gooday, A. J .; Aranda da Silva, A .; Pawlowski, J. (1. prosince 2011). „Xenophyophores (Rhizaria, Foraminifera) z kaňonu Nazaré (okraj Portugalska, SV Atlantik)“. Deep-Sea Research Část II: Aktuální studie v oceánografii. Geologie, geochemie a biologie podmořských kaňonů západně od Portugalska. 58 (23–24): 2401–2419. Bibcode:2011DSRII..58.2401G. doi:10.1016 / j.dsr2.2011.04.005.
- ^ Neil A C, Reece J B, Simon E J (2004) Základní biologie s fyziologií Pearson / Benjamin Cummings, strana 291. ISBN 9780805375039
- ^ O'Malley MA, Simpson AG, Roger AJ (2012). „Ostatní eukaryoty ve světle evoluční protistologie“. Biologie a filozofie. 28 (2): 299–330. doi:10.1007 / s10539-012-9354-r. S2CID 85406712.
- ^ Adl SM, Simpson AG, Farmer MA, Andersen RA, Anderson OR, Barta JR, Bowser SS, Brugerolle G, Fensome RA, Fredericq S, James TY, Karpov S, Kugrens P, Krug J, Lane CE, Lewis LA, Lodge J Lynn DH, Mann DG, McCourt RM, Mendoza L, Moestrup O, Mozley-Standridge SE, Nerad TA, Shearer CA, Smirnov AV, Spiegel FW, Taylor MF (2005). „Nová vyšší klasifikace eukaryot s důrazem na taxonomii protistů“. The Journal of Eukaryotic Microbiology. 52 (5): 399–451. doi:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- ^ Margulis L, Chapman MJ (19. března 2009). Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth. Akademický tisk. ISBN 9780080920146.
- ^ Janet Fang (6. dubna 2010). „Zvířatům se na dně moře daří bez kyslíku“. Příroda. 464 (7290): 825. doi:10.1038 / 464825b. PMID 20376121.
- ^ „V slaném nálevu mohou být zvířata, kterým se daří bez kyslíku“. Vědecké zprávy. 23. září 2013.
- ^ Hyde, K.D .; E.B.J. Jones; E. Leaño; S.B. Polohovací; A.D. Poonyth; L.L.P. Vrijmoed (1998). „Role hub v mořských ekosystémech“. Biodiverzita a ochrana. 7 (9): 1147–1161. doi:10.1023 / A: 1008823515157. S2CID 22264931.
- ^ Kirk, P.M., Cannon, P.F., Minter, D.W. a Stalpers, J. „Slovník hub“. Edn 10. CABI, 2008
- ^ Hyde, K.D .; E.B.J. Jones (1989). "Uspoření spor v mořských houbách". Botanica Marina. 32 (3): 205–218. doi:10,1515 / botm.1989.32.3.205. S2CID 84879817.
- ^ Le Calvez T, Burgaud G, Mahé S, Barbier G, Vandenkoornhuyse P (říjen 2009). „Houbová rozmanitost v hlubinných hydrotermálních ekosystémech“. Aplikovaná a environmentální mikrobiologie. 75 (20): 6415–21. doi:10.1128 / AEM.00653-09. PMC 2765129. PMID 19633124.
- ^ San-Martín, A .; S. Orejanera; C. Gallardo; M. Silva; J. Becerra; R. Reinoso; M.C. Chamy; K. Vergara; J. Rovirosa (2008). "Steroidy z mořské houby Geotrichum sp". Journal of the Chilean Chemical Society. 53 (1): 1377–1378. doi:10.4067 / S0717-97072008000100011.
- ^ Jones, E.B.G., Hyde, K.D., & Pang, K.-L., eds. (2014). Sladkovodní houby: a houbovité organismy. Berlín / Boston: De Gruyter.
- ^ Jones, E.B.G .; Pang, K.-L., eds. (2012). Mořské houby a houbové organismy. Mořská a sladkovodní botanika. Berlin, Boston: De Gruyter (zveřejněno v srpnu 2012). doi:10.1515/9783110264067. ISBN 978-3-11-026406-7. Citováno 3. září 2015.
- ^ Wang, Xin; Singh, Purnima; Gao, Zheng; Zhang, Xiaobo; Johnson, Zackary I .; Wang, Guangyi (2014). „Distribuce a rozmanitost planktonových hub v teplém bazénu západního Pacifiku“. PLOS ONE. 9 (7): e101523. Bibcode:2014PLoSO ... 9j1523W. doi:10.1371 / journal.pone.0101523.s001. PMC 4081592. PMID 24992154.
- ^ Wang, G .; Wang, X .; Liu, X .; Li, Q. (2012). „Diverzita a biogeochemická funkce planktonických hub v oceánu“. V Raghukumar, C. (ed.). Biologie mořských hub. Berlin, Heidelberg: Springer-Verlag. 71–88. doi:10.1007/978-3-642-23342-5. ISBN 978-3-642-23341-8. S2CID 39378040. Citováno 3. září 2015.
- ^ Damare, Samir; Raghukumar, Chandralata (11. listopadu 2007). „Houby a makroagregace v hlubinných sedimentech“. Mikrobiální ekologie. 56 (1): 168–177. doi:10.1007 / s00248-007-9334-r. ISSN 0095-3628. PMID 17994287. S2CID 21288251.
- ^ Kubánek, Julia; Jensen, Paul R .; Keifer, Paul A .; Sullards, M. Cameron; Collins, Dwight O .; Fenical, William (10. června 2003). „Odolnost mořských řas proti mikrobiálnímu napadení: cílená chemická ochrana proti mořským houbám“. Sborník Národní akademie věd. 100 (12): 6916–6921. Bibcode:2003PNAS..100,6916 tis. doi:10.1073 / pnas.1131855100. ISSN 0027-8424. PMC 165804. PMID 12756301.
- ^ A b Gao, Zheng; Johnson, Zackary I .; Wang, Guangyi (30. července 2009). „Molekulární charakterizace prostorové rozmanitosti a nových linií mykoplanktonu v havajských pobřežních vodách“. Časopis ISME. 4 (1): 111–120. doi:10.1038 / ismej.2009.87. ISSN 1751-7362. PMID 19641535.
- ^ Panzer, Katrin; Yilmaz, Pelin; Weiß, Michael; Reich, Lothar; Richter, Michael; Wiese, Jutta; Schmaljohann, Rolf; Labes, Antje; Imhoff, Johannes F. (30. července 2015). „Identifikace biomů specifických pro stanoviště vodních plísňových komunit pomocí komplexního souboru rRNA s téměř plnou délkou 18S rRNA obohaceného o kontextová data“. PLOS ONE. 10 (7): e0134377. Bibcode:2015PLoSO..1034377P. doi:10.1371 / journal.pone.0134377. PMC 4520555. PMID 26226014.
- ^ Gutierrez, Marcelo H; Pantoja, Silvio; Quinones, Renato a a Gonzalez, Rodrigo R. První záznam o plesnivých houbách v pobřežním upwellingovém ekosystému mimo centrální Chile. Gayana (Concepc.) [online]. 2010, sv. 74, č. 1, s. 66-73. ISSN 0717-6538.
- ^ A b Sridhar, K.R. (2009). „10. Vodní houby - jsou planktonické?“. Dynamika planktonu indických vod. Jaipur, Indie: Pratiksha Publications. 133–148.
- ^ Druhy vyšších mořských hub Archivováno 22 dubna 2013 na Wayback Machine University of Mississippi. Citováno 2012-02-05.
- ^ Sladkovodní a mořské lišejníkotvorné houby Citováno 2012-02-06.
- ^ "Lišejníky". Služba národního parku, americké ministerstvo vnitra, vláda Spojených států. 22. května 2016. Citováno 4. dubna 2018.
- ^ A b „The Earth Life Web, Growth and Development in Lichens“. earthlife.net.
- ^ Silliman B. R. & S. Y. Newell (2003). „Houbové zemědělství v hlemýždi“. PNAS. 100 (26): 15643–15648. Bibcode:2003PNAS..10015643S. doi:10.1073 / pnas.2535227100. PMC 307621. PMID 14657360.
- ^ Yuan X, Xiao S (2005). „Symbióza podobná lišejníku před 600 miliony let“. Věda. 308 (5724): 1017–1020. Bibcode:2005Sci ... 308.1017Y. doi:10.1126 / science.1111347. PMID 15890881. S2CID 27083645.
- ^ Jones, E. B. Gareth; Pang, Ka-Lai (31. srpna 2012). Mořské houby: a houbové organismy. Walter de Gruyter. ISBN 9783110264067.
- ^ Davidson, Michael W. (26. května 2005). "Struktura zvířecích buněk". Molekulární výrazy. Tallahassee, Florida: Florida State University. Citováno 3. září 2008.
- ^ Vogel, Gretchen (20. září 2018). „Tato fosilie je podle molekul tuku uchovaných po dobu půl miliardy let jedním z prvních zvířat na světě“. Věda. AAAS. Citováno 21. září 2018.
- ^ Bobrovskiy, Ilya (2018). „Starověké steroidy zakládají fosilii Ediacaran Dickinsonia jako jedno z prvních zvířat“. Věda. 361 (6408): 1246–1249. Bibcode:2018Sci ... 361.1246B. doi:10.1126 / science.aat7228. PMID 30237355.
- ^ Retallack, G.J. (2007). „Růst, rozpad a pohřební zhutnění Dickinsonia, ikonické fosílie Ediacaran“ (PDF). Alcheringa: Australasian Journal of paleontologie. 31 (3): 215–240. doi:10.1080/03115510701484705. S2CID 17181699.
- ^ Sperling, Erik; Vinther, Jakob; Pisani, Davide; Peterson, Kevin (2008). „Placozoanská afinita k Dickinsonia a vývoj režimů krmení pozdních prekambrických metazoanů “ (PDF). V Cusack, M; Owen, A; Clark, N (eds.). Program s abstrakty. Výroční zasedání paleontologické asociace. 52. Glasgow, Velká Británie. str. 81.
- ^ Gold, D. A .; Runnegar, B .; Gehling, J. G .; Jacobs, D. K. (2015). „Rekonstrukce ontogeneze předků podporuje bilaterální afinitu k Dickinsonii“. Evoluce a vývoj. 17 (6): 315–397. doi:10.1111 / ede.12168. PMID 26492825. S2CID 26099557.
- ^ Jun-Yuan Chen; Oliveri, Paola; Feng Gao; et al. (1. srpna 2002). „Život prekambrických zvířat: Pravděpodobné vývojové a dospělé formy Cnidarian z jihozápadní Číny“ (PDF). Vývojová biologie. 248 (1): 182–196. doi:10.1006 / dbio.2002.0714. ISSN 0012-1606. PMID 12142030. Archivovány od originál (PDF) dne 26. května 2013. Citováno 4. února 2015.
- ^ Grazhdankin, Dima (červen 2004). „Distribuční vzorce v Ediacaranské biotě: facie versus biogeografie a evoluce“. Paleobiologie. 30 (2): 203–221. doi:10.1666 / 0094-8373 (2004) 030 <0203: PODITE> 2.0.CO; 2. ISSN 0094-8373.
- ^ Seilacher, Adolf (Srpen 1992). „Vendobionta a Psammocorallia: ztracené stavby předkambrijské evoluce“. Časopis geologické společnosti. 149 (4): 607–613. Bibcode:1992JGSoc.149..607S. doi:10.1144 / gsjgs.149.4.0607. ISSN 0016-7649. S2CID 128681462. Citováno 4. února 2015.
- ^ Martin, Mark W .; Grazhdankin, Dmitriy V .; Bowring, Samuel A .; et al. (5. května 2000). „Věk neoproterozoických bilaterálních fosílií těla a stop, Bílé moře, Rusko: důsledky pro vývoj metazoanů“. Věda. 288 (5467): 841–845. Bibcode:2000 sci ... 288..841 mil. doi:10.1126 / science.288.5467.841. ISSN 0036-8075. PMID 10797002. S2CID 1019572.
- ^ Fedonkin, Michail A .; Wagoner, Benjamin M. (28. srpna 1997). „Pozdní prekambrická fosilie Kimberella je měkkýšovitý bilaterální organismus “. Příroda. 388 (6645): 868–871. Bibcode:1997 Natur.388..868F. doi:10.1038/42242. ISSN 0028-0836. S2CID 4395089.
- ^ Mooi, bohatý; David, Bruno (prosinec 1998). „Evoluce v bizarním kmeni: Homologie prvních ostnokožců“ (PDF). Americký zoolog. 38 (6): 965–974. doi:10.1093 / icb / 38.6.965. ISSN 1540-7063. Citováno 5. února 2015.
- ^ McMenamin, Mark A. S. (Září 2003). Spriggina je trilobitoidní ekdyzozoán. Geoscience Horizons Seattle 2003. Abstrakty s programy. 35. Boulder, Colo .: Geologická společnost Ameriky. str. 105. OCLC 249088612. Archivovány od originál dne 12. dubna 2016. Citováno 24. listopadu 2007. Příspěvek č. 40-2 představený na výročním zasedání geologické společnosti v Americe v Seattlu v roce 2003 (2. – 5. Listopadu 2003) dne 2. listopadu 2003 na Washington State Convention Center.
- ^ Jih-Pai Lin; Gon, Samuel M., III; Gehling, James G .; et al. (2006). "A Parvancorina- jako členovec z kambriu v jižní Číně “. Historická biologie: Mezinárodní žurnál paleobiologie. 18 (1): 33–45. doi:10.1080/08912960500508689. ISSN 1029-2381. S2CID 85821717.
- ^ Butterfield, Nicholas J. (prosinec 2006). „Zaháknutí některých„ červů “ze skupiny kmenů: fosilní lophotrochozoany v měděné břidlici.“ BioEssays. 28 (12): 1161–1166. doi:10.1002 / bies.20507. ISSN 0265-9247. PMID 17120226. S2CID 29130876.
- ^ A b Bengtson 2004, s. 67–78
- ^ Valentine, James W. (2004). O původu Phyly. Chicago: University Of Chicago Press. str. 7. ISBN 978-0-226-84548-7.
Klasifikace organismů v hierarchických systémech se používaly v sedmnáctém a osmnáctém století. Organismy byly obvykle seskupeny podle jejich morfologických podobností, jak je vnímají tito raní pracovníci, a tyto skupiny byly poté seskupeny podle jejich podobností atd., Aby vytvořily hierarchii.
- ^ A b Valentine, James W (18. června 2004). O původu Phyly. ISBN 9780226845487.
- ^ Redakční rada WoRMS. Světový registr mořských druhů. Dostupný online: http://www.marinespecies.org ve společnosti VLIZ. Přístup: 18. října 2019.
- ^ Novak, B.J .; Fraser, D .; Maloney, T.H. (2020). „Transformace ochrany oceánů: použití sady nástrojů pro genetickou záchranu“. Geny. 11 (2): 209. doi:10,3390 / geny11020209. PMC 7074136. PMID 32085502.
- ^ Gould, Stephen Jay (1990) Wonderful Life: The Burgess Shale and the Nature of History W. W. Norton. ISBN 9780393307009.
- ^ Erwin, Douglas; Valentine, James; Jablonski, David (1997). „Nedávné fosilní nálezy a nové poznatky o vývoji zvířat poskytují nové pohledy na hádanku exploze zvířat během raného kambriu“. Americký vědec (Březen duben).
- ^ A b Budd, G. E.; Jensen, S. (květen 2000). „Kritické přehodnocení fosilních záznamů bilaterální phyly“. Biologické recenze. 75 (2): 253–295. doi:10.1111 / j.1469-185X.1999.tb00046.x. PMID 10881389. S2CID 39772232.
- ^ Gould 1989
- ^ Budd, Graham E. (Únor 2003). „Kambrijský fosilní záznam a původ kmene“ (PDF). Integrativní a srovnávací biologie. 43 (1): 157–165. doi:10.1093 / icb / 43.1.157. ISSN 1557-7023. PMID 21680420. Citováno 6. února 2015.
- ^ Budd, Graham E. (březen 1996). „Morfologie Opabinia regalis a rekonstrukce skupiny kmenů členovců“. Lethaia. 29 (1): 1–14. doi:10.1111 / j.1502-3931.1996.tb01831.x. ISSN 0024-1164.
- ^ Marshall, Charles R. (květen 2006). „Vysvětlení kambrijské„ exploze “zvířat“. Výroční přehled o Zemi a planetárních vědách. 34: 355–384. Bibcode:2006AREPS..34..355M. doi:10.1146 / annurev.earth.33.031504.103001. ISSN 1545-4495. S2CID 85623607.
- ^ King, N .; Rokas, A. (2017). „Přijetí nejistoty při rekonstrukci rané evoluce zvířat“. Aktuální biologie. 27 (19): R1081 – R1088. doi:10.1016 / j.cub.2017.08.054. PMC 5679448. PMID 29017048.
- ^ Feuda, R .; Dohrmann, M .; Pett, W .; Philippe, H .; Rota-Stabelli, O .; Lartillot, N .; Wörheide, G .; Pisani, D. (2017). „Vylepšené modelování heterogenity kompozice podporuje houby jako sestru všech ostatních zvířat“. Aktuální biologie. 27 (24): 3864–3870. doi:10.1016 / j.cub.2017.11.008. PMID 29199080.
- ^ Nielsen, Claus (2019). ""Časná evoluce zvířat: pohled morfologa ", přečíst článek". Royal Society Open Science. 6 (7): 7. doi:10.1098 / rsos.190638. PMC 6689584. PMID 31417759.
- ^ Porifera (n.) Online slovník etymologie. Vyvolány 18 August je 2016.
- ^ A b Petralia, R.S .; Mattson, M.P .; Yao, P.J. (2014). „Stárnutí a dlouhověkost v nejjednodušších zvířatech a hledání nesmrtelnosti“. Recenze výzkumu stárnutí. 16: 66–82. doi:10.1016 / j.arr.2014.05.003. PMC 4133289. PMID 24910306.
- ^ Jochum, K.P .; Wang, X .; Vennemann, T.W .; Sinha, B .; Müller, W.E. (2012). „Křemičitá hlubinná houba Monorhaphis chuni: Potenciální paleoklimatický archiv ve starověkých zvířatech “. Chemická geologie. 300: 143–151. Bibcode:2012ChGeo.300..143J. doi:10.1016 / j.chemgeo.2012.01.009.
- ^ Vacelet & Duport 2004, str. 179–190.
- ^ "Spongia Linnaeus, 1759". Světový registr mořských druhů. Citováno 18. července 2012.
- ^ Rowland, S. M. & Stephens, T. (2001). "Archaeocyatha: Historie fylogenetické interpretace". Journal of Paleontology. 75 (6): 1065–1078. doi:10.1666 / 0022-3360 (2001) 075 <1065: AAHOPI> 2.0.CO; 2. JSTOR 1307076.
- ^ Sperling, E. A .; Pisani, D .; Peterson, K. J. (1. ledna 2007). „Poriferan parafyly a jeho důsledky pro prekambrickou paleobiologii“ (PDF). Geologická společnost, Londýn, speciální publikace. 286 (1): 355–368. Bibcode:2007GSLSP.286..355S. doi:10.1144 / SP286.25. S2CID 34175521. Archivovány od originál (PDF) dne 20. prosince 2009. Citováno 22. srpna 2012.
- ^ Ruppert, E.E .; Fox, R.S. & Barnes, RD (2004). Zoologie bezobratlých (7 ed.). Brooks / Cole. str.182–195. ISBN 978-0-03-025982-1.
- ^ Mills, C.E. „Ctenophores - několik poznámek od odborníka“. Citováno 5. února 2009.
- ^ A b Brusca R. C. a Brusca G. J. (2003) Bezobratlí, Druhé vydání, Sinauer Associates. ISBN 9780878930975.
- ^ Michael Le Page (březen 2019). „Zvíře s konečníkem, které přichází a odchází, může odhalit, jak se naše vyvinulo“. Nový vědec.
- ^ Martindale, Mark; Finnerty, J.R .; Henry, J.Q. (Září 2002). „Radiata a evoluční původ plánu bilaterálního těla“. Molekulární fylogenetika a evoluce. 24 (3): 358–365. doi:10.1016 / s1055-7903 (02) 00208-7. PMID 12220977.
- ^ Placozoa v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- ^ Rüdiger Wehner & Walter Gehring (červen 2007). Zoologie (v němčině) (24. vydání). Stuttgart: Thieme. str. 696.
- ^ F. E. Schulze "Trichoplax adhaerens n. g., n. s. ", Zoologischer Anzeiger (Elsevier, Amsterdam a Jena) 6 (1883), s. 92.
- ^ Eitel, Michael; Francis, Warren; Osigus, Hans-Jürgen; Krebs, Stefan; Vargas, Sergio; Blum, Helmut; Williams, Gray Argust; Schierwater, Bernd; Wörheide, Gert (13. října 2017). „Taxogenomický přístup odhaluje nový rod v kmeni Placozoa“. bioRxiv: 202119. doi:10.1101/202119. S2CID 89829846.
- ^ Schierwater, Bernd; Kamm, Kai; Herzog, Rebecca; Rolfes, Sarah; Osigus, Hans-Jürgen (4. března 2019). „Polyplacotoma mediterranea je nový rozvětvený druh placozoan“. Aktuální biologie. 29 (5): R148 – R149. doi:10.1016 / j.cub.2019.01.068. ISSN 0960-9822. PMID 30836080.
- ^ Trichoplax adhaerens, WoRMS, 2009.
- ^ Smith, Carolyn L .; Varoqueaux, Frédérique; Kittelmann, Maike; Azzam, Rita N .; Cooper, Benjamin; Winters, Christine A .; Eitel, Michael; Fasshauer, Dirk; Reese, Thomas S. (2014). „Nové typy buněk, neurosekreční buňky a tělesný plán raných divergujících metazoanů Trichoplax adhaerens“. Aktuální biologie. 24 (14): 1565–1572. doi:10.1016 / j.cub.2014.05.046. ISSN 0960-9822. PMC 4128346. PMID 24954051.
- ^ Barnes, Robert D. (1982). Zoologie bezobratlých. Philadelphia: Holt-Saunders International. str. 84–85. ISBN 978-0-03-056747-6.
- ^ Zhang, Z.-Q. (2011). „Biodiverzita zvířat: Úvod do vyšší klasifikace a taxonomického bohatství“ (PDF). Zootaxa. 3148: 7–12. doi:10.11646 / zootaxa.3148.1.3.
- ^ "Nematostella vectensis v1.0 ". Portál genomu. University of California. Citováno 19. ledna 2014.
- ^ "Nematostella". Nematostella.org. Archivovány od originál dne 8. května 2006. Citováno 18. ledna 2014.
- ^ A b Genikhovich, G. & U. Technau (2009). „Hvězdná mořská sasanka Nematostella vectensis: Anthozoanský modelový organismus pro studium srovnávací genomiky a funkční evoluční vývojové biologie ". Cold Spring Harbor Protocols. 2009 (9): pdb.emo129. doi:10.1101 / pdb.emo129. PMID 20147257.
- ^ „Odkud pochází naše hlava? Bezmozká mořská sasanka vrhá nové světlo na evoluční původ hlavy“. Věda denně. 12. února 2013. Citováno 18. ledna 2014.
- ^ Sinigaglia, C .; et al. (2013). „Bilaterální gen pro modelování hlavy six3 / 6 řídí vývoj aborální domény u cnidariana“. PLOS Biology. 11 (2): e1001488. doi:10.1371 / journal.pbio.1001488. PMC 3586664. PMID 23483856.
- ^ "Červené papírové lucerny". Skutečné obludnosti. Citováno 25. října 2015.
- ^ „Modrá tlačítka na Floridě.“
- ^ Karleskint G, Richard Turner R a, James Small J (2012) Úvod do mořské biologie Cengage Learning, vydání 4, strana 445. ISBN 9781133364467.
- ^ Bavestrello, Giorgio; Christian Sommer; Michele Sarà (1992). "Obousměrná přeměna v Turritopsis nutricula (Hydrozoa)". Scientia Marina. 56 (2–3): 137–140.
- ^ Piraino, Stefano; F. Boero; B. Aeschbach; V. Schmid (1996). "Zvrácení životního cyklu: medúzy transformující se na polypy a buněčná transdiferenciace v Turritopsis nutricula (Cnidaria, Hydrozoa)". Biologický bulletin. 190 (3): 302–312. doi:10.2307/1543022. JSTOR 1543022. PMID 29227703. S2CID 3956265.
- ^ Fenner PJ, Williamson JA (1996). "Celosvětová úmrtí a těžké envenomation od medúzy bodnutí". Medical Journal of Australia. 165 (11–12): 658–61. doi:10.5694 / j.1326-5377.1996.tb138679.x. PMID 8985452. S2CID 45032896.
- ^ A b Cannon, Johanna Taylor; Vellutini, Bruno Cossermelli; Smith, Julian; Ronquist, Fredrik; Jondelius, Ulf; Hejnol, Andreas (2016). „Xenacoelomorpha je sesterská skupina Nephrozoa“. Příroda. 530 (7588): 89–93. Bibcode:2016Natur.530 ... 89C. doi:10.1038 / příroda16520. PMID 26842059. S2CID 205247296.
- ^ A b Minelli, Alessandro (2009). Perspektivy ve živočišné fylogenezi a evoluci. Oxford University Press. str. 53. ISBN 978-0-19-856620-5.
- ^ A b C Brusca, Richard C. (2016). Úvod do Bilateria a kmene Xenacoelomorpha Triploblasty a bilaterální symetrie poskytují nové cesty pro radiaci zvířat (PDF). Bezobratlí. Sinauer Associates. str. 345–372. ISBN 978-1605353753.
- ^ Finnerty, John R. (listopad 2005). „Upřednostňoval vnitřní transport, spíše než řízený pohyb, vývoj bilaterální symetrie u zvířat?“ (PDF). BioEssays. 27 (11): 1174–1180. doi:10.1002 / bies.20299. PMID 16237677. Archivovány od originál (PDF) dne 10. srpna 2014. Citováno 27. srpna 2019.
- ^ Quillin, K. J. (květen 1998). „Ontogenetické škálování hydrostatických skeletů: škálování geometrického, statického a dynamického napětí žížaly lumbricus terrestris“. The Journal of Experimental Biology. 201 (12): 1871–83. PMID 9600869.
- ^ Tento pravěký červ může být předkem všech zvířat Živá věda, 26. března 2020.
- ^ Cannon, J.T .; Vellutini, B.C .; Smith, J .; Ronquist, F .; Jondelius, U .; Hejnol, A. (2016). „Xenacoelomorpha je sesterská skupina Nephrozoa“. Příroda. 530 (7588): 89–93. Bibcode:2016Natur.530 ... 89C. doi:10.1038 / příroda16520. PMID 26842059. S2CID 205247296.
- ^ A b Wade, Nicholas (30. ledna 2017). „Tento prehistorický předek člověka měl všechno na ústa“. The New York Times. Citováno 31. ledna 2017.
- ^ A b Han, Jian; Morris, Simon Conway; Ou, Qiang; Shu, Degan; Huang, Hai (2017). „Meiofaunal deuterostomes from basal Cambrian of Shaanxi (China)“. Příroda. 542 (7640): 228–231. Bibcode:2017Natur.542..228H. doi:10.1038 / nature21072. ISSN 0028-0836. PMID 28135722. S2CID 353780.
- ^ „Cornwall - Příroda - Červ Superstar“. BBC.
- ^ Mark Carwardine (1995) Guinnessova kniha záznamů o zvířatech. Guinness Publishing. str. 232.
- ^ „Trvalí paraziti“. Čas. 8. dubna 1957.
- ^ Hargis, William J. (1985). „Parazitologie a patologie mořských organismů světového oceánu“. Národní úřad pro oceán a atmosféru. Citovat deník vyžaduje
| deník =(Pomoc) - ^ http://plpnemweb.ucdavis.edu/nemaplex/General/animpara.htm
- ^ Lynne S.Garcia (1999). „Klasifikace lidských parazitů, vektorů a podobných organismů“ (PDF). Clin Infect Dis. 29 (4): 734–6. doi:10.1086/520425. PMID 10589879.
- ^ Hodda, M (2011). „Phylum Nematoda Cobb, 1932. In: Zhang, Z.-Q. Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness“. Zootaxa. 3148: 63–95. doi:10.11646 / zootaxa.3148.1.11.
- ^ Zhang, Z (2013). „Biodiverzita zvířat: Aktualizace klasifikace a rozmanitosti v roce 2013. In: Zhang, Z.-Q. (Ed.) Animal Biodiversity: An Outline of High-level Classification and Survey of Taxonomic Richness (Addenda 2013)“. Zootaxa. 3703 (1): 5–11. doi:10.11646 / zootaxa.3703.1.3. S2CID 85252974.
- ^ Lambshead PJD (1993). „Poslední vývoj ve výzkumu bentické mořské biodiverzity“. Oceanis. 19 (6): 5–24.
- ^ Borgonie G, García-Moyano A, Litthauer D, Bert W, Bester A, van Heerden E, Möller C, Erasmus M, Onstott TC (červen 2011). „Nematoda z pozemského hlubokého podzemí Jižní Afriky“. Příroda. 474 (7349): 79–82. Bibcode:2011Natur.474 ... 79B. doi:10.1038 / nature09974. hdl:1854 / LU-1269676. PMID 21637257. S2CID 4399763.
- ^ Danovaro R, Gambi C, Dell'Anno A, Corinaldesi C, Fraschetti S, Vanreusel A, Vincx M, Gooday AJ (leden 2008). „Exponenciální pokles fungování hlubinného ekosystému spojený se ztrátou bentické biologické rozmanitosti“. Aktuální biologie. 18 (1): 1–8. doi:10.1016 / j.cub.2007.11.056. PMID 18164201. S2CID 15272791. Shrnutí ležel – EurekAlert!.
- ^ Platt HM (1994). "úvodní slovo". In Lorenzen S, Lorenzen SA (eds.). Fylogenetická systematika volně žijících hlístic. London: The Ray Society. ISBN 978-0-903874-22-9.
- ^ Barnes, R.S.K .; Calow, P .; Olive, P.J.W .; Golding, D.W. & Spicer, J.I. (2001). Bezobratlí, syntéza (3. vyd.). UK: Blackwell Science.
- ^ Tyrian Purple Zelený lev, 28. února 2014.
- ^ Chapman, A.D. (2009). Numbers of Living Species in Australia and the World, 2. vydání. Australská studie biologických zdrojů, Canberra. Citováno 12. ledna 2010. ISBN 978-0-642-56860-1 (tištěné); ISBN 978-0-642-56861-8 (online).
- ^ Hancock, Rebecca (2008). „Uznáváme výzkum měkkýšů“. Australské muzeum. Archivovány od originál dne 30. května 2009. Citováno 9. března 2009.
- ^ Ponder, W.F .; Lindberg, D.R., eds. (2008). Fylogeneze a evoluce měkkýšů. Berkeley: University of California Press. str. 481. ISBN 978-0-520-25092-5.
- ^ Munro, D .; Blier, P.U. (2012). "Extrémní dlouhověkost Arctica islandica je spojena se zvýšenou odolností proti peroxidaci v mitochondriálních membránách “. Stárnoucí buňka. 11 (5): 845–55. doi:10.1111 / j.1474-9726.2012.00847.x. PMID 22708840. S2CID 205634828.
- ^ „Vítejte na CephBase“. CephBase. Citováno 29. ledna 2016.
- ^ Wilbur, Karl M .; Clarke, M.R .; Trueman, E.R., eds. (1985), Měkkýši, 12. Paleontologie a neontologie hlavonožců, New York: Academic Press, ISBN 0-12-728702-7
- ^ „Existují nějaké sladkovodní hlavonožci?“. 16. ledna 2013.
- ^ Ewen Callaway (2. června 2008). „Simple-Minded Nautilus Show Flash of Memory“. Nový vědec. Citováno 7. března 2012.
- ^ Kathryn Phillips (15. června 2008). „Živé fosilní vzpomínky“. Journal of Experimental Biology. 211 (12): iii. doi:10.1242 / jeb.020370. S2CID 84279320.
- ^ Robyn Crook & Jennifer Basil (2008). „Dvoufázová paměťová křivka v komorovém nautilu, Nautilus pompilius L. (Cephalopoda: Nautiloidea) ". Journal of Experimental Biology. 211 (12): 1992–1998. doi:10.1242 / jeb.018531. PMID 18515730. S2CID 6305526.
- ^ Black, Richard (26. dubna 2008). „Kolosální chobotnice z mrazáku“. BBC novinky.
- ^ Ruppert, Edward E .; Fox, Richard, S .; Barnes, Robert D. (2004). Zoologie bezobratlých (7. vydání). Cengage Learning. ISBN 978-81-315-0104-7.
- ^ Hayward, PJ (1996). Příručka pro mořské fauny severozápadní Evropy. Oxford University Press. ISBN 978-0-19-854055-7.
- ^ Ruppert, Fox & Barnes (2004), str. 518–522
- ^ Wilson, Heather M .; Anderson, Lyall I. (leden 2004). "Morfologie a taxonomie paleozoických mnohonožek (Diplopoda: Chilognatha: Archipolypoda) ze Skotska". Journal of Paleontology. 78 (1): 169–184. doi:10.1666 / 0022-3360 (2004) 078 <0169: MATOPM> 2.0.CO; 2.
- ^ Stephanie E. Suarez; Michael E. Brookfield; Elizabeth J. Catlos; Daniel F. Stöckli (2017). „Omezení stáří zirkonu U-Pb na nejstarší zaznamenané pozemské zvíře dýchající vzduch“. PLOS ONE. 12 (6): e0179262. Bibcode:2017PLoSO..1279262S. doi:10.1371 / journal.pone.0179262. PMC 5489152. PMID 28658320.
- ^ Campbell, L.I .; Rota-Stabelli, O .; Edgecombe, G.D .; Marchioro, T .; Longhorn, S.J .; Telford, M.J .; Philippe, H .; Rebecchi, L .; Peterson, K.J .; Pisani, D. (2011). „MicroRNA a fylogenomika řeší vztahy Tardigrady a naznačují, že sametové červy jsou sesterskou skupinou Arthropoda“. Sborník Národní akademie věd. 108 (38): 15920–15924. Bibcode:2011PNAS..10815920C. doi:10.1073 / pnas.1105499108. PMC 3179045. PMID 21896763.
- ^ Smith, F.W .; Goldstein, B. (2017). "Segmentace v Tardigrada a diverzifikace segmentových vzorů v Panarthropoda". Struktura a vývoj členovců. 46 (3): 328–340. doi:10.1016 / j.asd.2016.10.005. PMID 27725256.
- ^ Budd, G. E. (2001). „Proč jsou členovci segmentováni?“. Evoluce a vývoj. 3 (5): 332–42. doi:10.1046 / j.1525-142X.2001.01041.x. PMID 11710765. S2CID 37935884.
- ^ „Archivovaná kopie“. Archivovány od originál dne 26. ledna 2011. Citováno 10. března 2011.CS1 maint: archivovaná kopie jako titul (odkaz)
- ^ Braddy, Simon J .; Poschmann, Markus; Tetlie, O. Erik (2007). „Obří dráp odhaluje dosud největšího členovce“. Biologické dopisy. 4 (1): 106–109. doi:10.1098 / rsbl.2007.0491. PMC 2412931. PMID 18029297.
- ^ Cressey, Daniel (21. listopadu 2007). „Obří mořský štír objeven“. Příroda. doi:10.1038 / novinky.2007.272. Citováno 10. června 2013.
- ^ „Ošklivý obří krab Japonska“. Populární věda. 96 (6): 42. 1920.
- ^ D. R. Currie a T. M. Ward (2009). Krab jižní australský (Pseudocarcinus gigas) Rybářství (PDF). Jižní australský institut pro výzkum a vývoj. Zpráva o hodnocení rybolovu pro PIRSA. Archivovány od originál (PDF) dne 28. března 2012. Citováno 9. prosince 2013.
- ^ Patrick Kilday (28. září 2005). „Krevety kudlanky se pyšní nejpokročilejšími očima“. Denní kalifornský.
- ^ S. N. Patek a R. L. Caldwell (2005). "Extrémní nárazové a kavitační síly biologického kladiva: úderné síly krevetky páví". Journal of Experimental Biology. 208 (19): 3655–3664. doi:10.1242 / jeb.01831. PMID 16169943. S2CID 312009.
- ^ Ghosh, Pallab (30. ledna 2017). „Vědci najdou„ nejstaršího předka člověka “'". BBC. Citováno 30. ledna 2017.
- ^ „Web pro rozmanitost zvířat - Echinodermata“. Muzeum zoologie University of Michigan. Citováno 26. srpna 2012.
- ^ "Sea Lily". Science Encyclopedia. Citováno 5. září 2014.
- ^ Fox, Richarde. "Asterias forbesi". Anatomie bezobratlých online. Lander University. Citováno 14. června 2014.
- ^ Holsinger, K. (2005). Keystone druhy. Citováno 10. května 2010, z „Archivovaná kopie“. Archivovány od originál dne 30. června 2010. Citováno 12. května 2010.CS1 maint: archivovaná kopie jako titul (odkaz)
- ^ Simakov, O .; Kawashima, T .; Marlétaz, F .; Jenkins, J .; Koyanagi, R .; Mitros, T .; Hisata, K .; Bredeson, J .; Shoguchi, E .; Gyoja, F .; Yue, J.X. (2015). „Hemichordátové genomy a původ deuterostomu“. Příroda. 527 (7579): 459–65. Bibcode:2015 Natur.527..459S. doi:10.1038 / příroda16150. PMC 4729200. PMID 26580012.
- ^ Jak lidé dostali hltan z tohoto „ošklivého zvířete“, Budoucnost, 23. listopadu 2015.
- ^ A b C d Clark, M.A., Choi, J. a Douglas, M. (2018) Strunatci Biologie 2e. OpenStax. ISBN 9781947172951. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0
- ^ Tajemství života Oesia: Prehistorický červ postavil na mořském dně trubkovité „domy“
- ^ Barnes, Robert D. (1982). Zoologie bezobratlých. Philadelphia, PA: Holt-Saunders International. str. 1018–1026. ISBN 978-0-03-056747-6.
- ^ Sekundární organizátoři raného mozku a umístění mezo-diencefalických dopaminergních prekurzorových buněk Archivováno 10. března 2014 v Wayback Machine Citováno 10. března 2014
- ^ Rob Mitchum (15. března 2012). „Tajný původ mozku obratlovců“. Příroda. ScienceLife. 483 (7389): 289–94. doi:10.1038 / příroda 10838. PMC 3719855. PMID 22422262. Citováno 18. února 2014.
- ^ Strunatci OpenStax, 9. května 2019.
- ^ Gewin, V (2005). „Funkční genomika zesiluje biologickou zápletku“. PLOS Biology. 3 (6): e219. doi:10.1371 / journal.pbio.0030219. PMC 1149496. PMID 15941356.
- ^ Lanceletový (amphioxus) genom a původ obratlovců Ars Technica, 19. června 2008.
- ^ Lemaire, P (2011). "Evoluční křižovatka ve vývojové biologii: pláštěnci". Rozvoj. 138 (11): 2143–2152. doi:10,1242 / dev.048975. PMID 21558365. S2CID 40452112.
- ^ „FishBase: Globální informační systém o rybách“. FishBase. Citováno 17. ledna 2017.
- ^ "Kolik ryb v moři? Sčítání mořského života zahajuje první zprávu". Věda denně. Citováno 17. ledna 2017.
- ^ Docker, Margaret F. (2006) „Příspěvky Billa Beamisha k výzkumu Lamprey a nedávné pokroky v oboru“, Recenze guelph ichtyologie, 7.
- ^ Hardisty, M. W .; Potter, I. C. (1971). Hardisty, M. W .; Potter, I. C. (eds.). The Biology of Lampreys (1. vyd.). Akademický tisk. ISBN 978-0-123-24801-5.
- ^ Gill, Howard S .; Renaud, Claude B .; Chapleau, François; Mayden, Richard L .; Potter, Ian C .; Douglas, M. E. (2003). "Fylogeneze živých parazitických lamp (Petromyzontiformes) na základě morfologických údajů". Copeia. 2003 (4): 687–703. doi:10.1643 / IA02-085.1. S2CID 85969032.
- ^ "Myxini". Muzeum paleontologie University of California. Archivovány od originál dne 15. prosince 2017. Citováno 17. ledna 2017.
- ^ Greena, Stephen A .; Bronner, Marianne E. (2014). „The Lamprey: A jawless vertebels model system for zkoumání původu neurální lišty a dalších vlastností obratlovců“. Diferenciace. 87 (1–2): 44–51. doi:10.1016 / j.diff.2014.02.001. PMC 3995830. PMID 24560767.
- ^ Stock, D .; Whitt, GS (7. srpna 1992). "Důkazy z 18S ribozomálních RNA sekvencí, které lampreys a hagfish tvoří přirozenou skupinu". Věda. 257 (5071): 787–9. Bibcode:1992Sci ... 257..787S. doi:10.1126 / science.1496398. PMID 1496398.
- ^ Nicholls, H. (10. září 2009). "Z úst do úst". Příroda. 461 (7261): 164–166. doi:10.1038 / 461164a. PMID 19741680.
- ^ McCoy, Victoria E .; Saupe, Erin E .; Lamsdell, James C .; et al. (28. dubna 2016). „„ Tully monster “je obratlovec“. Příroda. 532 (7600): 496–499. Bibcode:2016Natur.532..496M. doi:10.1038 / příroda16992. PMID 26982721. S2CID 205247805.
- ^ Sallan, L .; Giles, S .; Sansom, R. S .; et al. (20. února 2017). „„ Tully Monster “není obratlovcem: znaky, konvergence a taphonomie u paleozoických problematických zvířat“ (PDF). Paleontologie. 60 (2): 149–157. doi:10.1111 / pala.12282.
- ^ Prastaré „tulí monstrum“ bylo obratlovcem, nikoli bezpáteřním blobem, tvrdí studie Živá věda, 4. května 2020.
- ^ McCoy, V.E .; Wiemann, J .; Lamsdell, J.C .; Whalen, C.D .; Lidgard, S .; Mayer, P .; Petermann, H .; Briggs, D.E. (2020). "Chemické podpisy měkkých tkání rozlišují mezi obratlovci a bezobratlými z karbonu Mazon Creek Lagerstätte v Illinois". Geobiologie. 18 (5): 560–565. doi:10,1111 / gbi.12397. PMID 32347003.
- ^ Kimmel, C. B .; Miller, C. T .; Keynes, R. J. (2001). „Vzorování neurální lišty a vývoj čelisti“. Anatomy Journal. 199 (1&2): 105–119. doi:10.1017 / S0021878201008068. PMC 1594948. PMID 11523812.
- ^ Gai, Z .; Zhu, M. (2012). „Původ čelisti obratlovců: Křižovatka mezi modelem založeným na vývojové biologii a fosilními důkazy“. Bulletin čínské vědy. 57 (30): 3819–3828. Bibcode:2012ChSBu..57,3819G. doi:10.1007 / s11434-012-5372-z.
- ^ Maisey, J. G. (2000). Objevování fosilních ryb. Westview Press. s. 1–223. ISBN 978-0-8133-3807-1.
- ^ A b Wroe, S .; Huber, D. R .; Lowry, M .; McHenry, C .; Moreno, K .; Clausen, P .; Ferrara, T. L .; Cunningham, E .; Dean, M. N .; Summers, A. P. (2008). „Trojrozměrná počítačová analýza mechaniky čelistí bílých žraloků: jak těžké může velké bílé sousto?“ (PDF). Journal of Zoology. 276 (4): 336–342. doi:10.1111 / j.1469-7998.2008.00494.x.
- ^ Pimiento, Catalina; Dana J. Ehret; Bruce J. MacFadden; Gordon Hubbell (10. května 2010). Stepanova, Anna (ed.). „Starověká školka pro vyhynulého velkého žraloka Megalodona z panamského miocénu“. PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO ... 510552P. doi:10.1371 / journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ Lambert, Olivier; Bianucci, Giovanni; Post, Klaas; de Muizon, Christian; Salas-Gismondi, Rodolfo; Urbina, Mario; Reumer, Jelle (1. července 2010). „Obří sousto nové raptoriální velryby spermií z miocénní epochy Peru“. Příroda. 466 (7302): 105–108. Bibcode:2010Natur.466..105L. doi:10.1038 / nature09067. PMID 20596020. S2CID 4369352.
- ^ Nielsen, Julius; Hedeholm, Rasmus B .; Heinemeier, Jan; Bushnell, Peter G .; Christiansen, Jørgen S .; Olsen, Jesper; Ramsey, Christopher Bronk; Brill, Richard W .; Simon, Malene; Steffensen, Kirstine F .; Steffensen, John F. (2016). „Radiokarbon oční čočky odhaluje staletí dlouhověkosti u žraloka grónského (Somniosus microcephalus)". Věda. 353 (6300): 702–4. Bibcode:2016Sci ... 353..702N. doi:10.1126 / science.aaf1703. PMID 27516602. S2CID 206647043. Shrnutí ležel – Sci novinky (12. srpna 2016).
- ^ Marshall, A .; Bennett, M.B .; Kodja, G .; Hinojosa-Alvarez, S .; Galvan-Magana, F .; Harding, M .; Stevens, G. & Kashiwagi, T. (2011). "Manta birostris". Červený seznam ohrožených druhů IUCN. 2011: e.T198921A9108067. doi:10.2305 / IUCN.UK.2011-2.RLTS.T198921A9108067.en.
- ^ Black, Richard (11. června 2007). „Ochrana Sawfish získává zuby“. BBC novinky.
- ^ Thomas J. Near; et al. (2012). „Řešení fylogeneze ryb s paprskovými ploutvemi a načasování diverzifikace“. PNAS. 109 (34): 13698–13703. Bibcode:2012PNAS..10913698N. doi:10.1073 / pnas.1206625109. PMC 3427055. PMID 22869754.
- ^ A b Zhu, M; Zhao, W; Jia, L; Lu, J; Qiao, T; Qu, Q (2009). "Nejstarší kloubový osteichthyan odhaluje mozaikové postavy gnathostome". Příroda. 458 (7237): 469–474. Bibcode:2009 Natur.458..469Z. doi:10.1038 / nature07855. PMID 19325627. S2CID 669711.
- ^ Clack, J. A. (2002) Získání půdy. Indiana University
- ^ „Chondrosteans: Jeseter Příbuzní“. paleos.com. Archivovány od originál dne 25. prosince 2010.
- ^ López-Arbarello, A (2012). „Fylogenetické vzájemné vztahy ryb Ginglymodian (Actinopterygii: Neopterygii)“. PLOS ONE. 7 (7): e39370. Bibcode:2012PLoSO ... 739370L. doi:10.1371 / journal.pone.0039370. PMC 3394768. PMID 22808031.
- ^ Berra, Tim M. (2008). Distribuce sladkovodních ryb. University of Chicago Press. str. 55. ISBN 978-0-226-04443-9.
- ^ Lackmann, Alec R .; Andrews, Allen H .; Butler, Malcolm G .; Bielak-Lackmann, Ewelina S .; Clark, Mark E. (23. května 2019). „Bigmouth Buffalo Ictiobus cyprinellus nastavuje sladkovodní teleostní rekord, protože vylepšená věková analýza odhaluje stoletou dlouhověkost“. Komunikační biologie. 2 (1): 197. doi:10.1038 / s42003-019-0452-0. ISSN 2399-3642. PMC 6533251. PMID 31149641.
- ^ A b Benton, Michael (2005). „Vývoj ryb po devonu“. Paleontologie obratlovců (3. vyd.). John Wiley & Sons. 175–84. ISBN 978-1-4051-4449-0.
- ^ Bone, Q .; Moore, R. (2008). Biologie ryb. Věnec věnec. str. 29. ISBN 978-0-415-37562-7.
- ^ Dorit, R. L .; Walker, W. F .; Barnes, R. D. (1991). Zoologie. Nakladatelství Saunders College Publishing. str.67–69. ISBN 978-0-03-030504-7.
- ^ „Vědci popisují nejmenší a nejlehčí rybu na světě“. Scripps Institution of Oceanography. 20. července 2004. Citováno 9. dubna 2016.
- ^ Roach, John (13. května 2003). „Objevena nejtěžší kostnatá ryba na světě?“. Zprávy z National Geographic. Citováno 9. ledna 2016.
- ^ Světová unie ochrany přírody. 2014. Červený seznam ohrožených druhů IUCN, 2014.3. Souhrnná statistika pro globálně ohrožené druhy. Tabulka 1: Počty ohrožených druhů podle hlavních skupin organismů (1996–2014).
- ^ Narkiewicz, Katarzyna; Narkiewicz, Marek (leden 2015). "Věk nejstarších tetrapodských stop z polského Zachełmie". Lethaia. 48 (1): 10–12. doi:10.1111 / let.12083. ISSN 0024-1164.
- ^ Long JA, Gordon MS (září – říjen 2004). „Největší krok v historii obratlovců: paleobiologický přehled přechodu ryby a tetrapoda“. Physiol. Biochem. Zool. 77 (5): 700–19. doi:10.1086/425183. PMID 15547790. S2CID 1260442. jako PDF
- ^ Shubin, N. (2008). Your Inner Fish: A Journey Into the 3,5-Billion-Year History of the Human Body. New York: Pantheon Books. ISBN 978-0-375-42447-2.
- ^ Laurin, M. (2010). Jak stavovci opustili vodu. Berkeley, Kalifornie, USA .: University of California Press. ISBN 978-0-520-26647-6.
- ^ Canoville, Aurore; Laurin, Michel (2010). „Vývoj humerální mikroanatomie a životního stylu u amniotů a některé komentáře k paleobiologickým závěrům“. Biologický žurnál společnosti Linnean. 100 (2): 384–406. doi:10.1111 / j.1095-8312.2010.01431.x.
- ^ Laurin, Michel; Canoville, Aurore; Quilhac, Alexandra (2009). „Využití paleontologických a molekulárních dat v nadstromech pro srovnávací studie: příklad lissamphibian femorální mikroanatomie“. Anatomy Journal. 215 (2): 110–123. doi:10.1111 / j.1469-7580.2009.01104.x. PMC 2740958. PMID 19508493.
- ^ Hopkins Gareth R .; Brodie Edmund D. Jr (2015). „Výskyt obojživelníků ve slaných stanovištích: recenze a evoluční perspektiva“. Herpetologické monografie. 29 (1): 1–27. doi:10.1655 / HERPMONOGRAPHS-D-14-00006. S2CID 83659304.
- ^ Natchev, Nikolay; Tzankov, Nikolay; Geme, Richard (2011). "Invaze zelených žab v Černém moři: ekologie biotopů Pelophylax esculentus komplex (Anura, Amphibia) populace v oblasti laguny Shablenska Тuzla v Bulharsku " (PDF). Herpetologie Poznámky. 4: 347–351.
- ^ Sander, P. Martin (2012). "Reprodukce v časných amniotech". Věda. 337 (6096): 806–808. Bibcode:2012Sci ... 337..806S. doi:10.1126 / science.1224301. PMID 22904001. S2CID 7041966.
- ^ Modesto, S.P .; Anderson, J.S. (2004). „Fylogenetická definice Reptilie“. Systematická biologie. 53 (5): 815–821. doi:10.1080/10635150490503026. PMID 15545258.

- ^ Gauthier, J. A.; Kluge, A.G .; Rowe, T. (1988). „Počáteční vývoj Amnioty“. V Benton, M.J. (ed.). Fylogeneze a klasifikace tetrapodů. 1. Oxford: Clarendon Press. 103–155. ISBN 978-0-19-857705-8.
- ^ Laurin, M .; Reisz, R. R. (1995). „Přehodnocení časné fylogeneze amniotu“ (PDF). Zoologický žurnál Linneanské společnosti. 113 (2): 165–223. doi:10.1111 / j.1096-3642.1995.tb00932.x. Archivovány od originál (PDF) dne 8. června 2019. Citováno 14. srpna 2016.
- ^ Modesto, S.P. (1999). „Pozorování struktury raně permského plazů Stereosternum tumidum Zvládnout". Palaeontologia Africana. 35: 7–19.
- ^ Rasmussen, Arne Redsted; Murphy, John C .; Ompi, Medy; Gibbons, J. Whitfield; Uetz, Peter (8. listopadu 2011). „Mořští plazi“. PLOS ONE. 6 (11): e27373. Bibcode:2011PLoSO ... 627373R. doi:10.1371 / journal.pone.0027373. PMC 3210815. PMID 22087300.
- ^ Stidworthy J. 1974. Hadi světa. Grosset & Dunlap Inc. 160 stran ISBN 0-448-11856-4.
- ^ Mořští hadi na Organizace OSN pro výživu a zemědělství. Zpřístupněno 22. srpna 2020.
- ^ Rasmussen, A.R .; Murphy, J.C .; Ompi, M .; Gibbons, J.W .; Uetz, P. (2011). „Mořští plazi“. PLOS ONE. 6 (11): e27373. Bibcode:2011PLoSO ... 627373R. doi:10.1371 / journal.pone.0027373. PMC 3210815. PMID 22087300.
- ^ Martill D.M. (1993). "Soupy Substrates: Médium pro výjimečné uchování ichthyosaurů z posidonské břidlice (dolní jury) v Německu". Kaupia - Darmstädter Beiträge zur Naturgeschichte, 2 : 77-97.
- ^ Gould, Stephen Jay (1993) „Bent Out of Shape“ v Osm malých prasátek: Úvahy o přírodní historii. Norton, 179–94. ISBN 9780393311396.
- ^ „Sardinka, běh žraloka, krmení fenoménu šílenství v Africe“. Archivovány od originál dne 2. prosince 2008.
- ^ „Seznam druhů a poddruhů Výboru pro taxonomii Společnosti pro mořskou mammalogii“. Společnost pro námořní mammalogii. Říjen 2015. Archivovány od originál dne 6. ledna 2015. Citováno 23. listopadu 2015.
- ^ Romer A. S. a Parsons T. S. (1986) Tělo obratlovců, strana 96, Sanders College Publishing. ISBN 0030584469.
- ^ "Modrá velryba". Světový fond na ochranu přírody. Citováno 15. srpna 2016.
- ^ Marino, Lori (2004). „Evoluce mozku kytovců: Násobení vytváří složitost“ (PDF). Mezinárodní společnost pro srovnávací psychologii (17): 1–16. Archivovány od originál (PDF) dne 16. září 2018. Citováno 15. srpna 2016.
- ^ A b Campbell, Neil A .; Reece, Jane B .; Urry, Lisa Andrea; Cain, Michael L .; Wasserman, Steven Alexander; Minorsky, Peter V .; Jackson, Robert Bradley (2008). Biologie (8 ed.). San Francisco: Pearson - Benjamin Cummings. ISBN 978-0-321-54325-7.
- ^ McNeill, J .; et al., eds. (2012). Mezinárodní kód nomenklatury pro řasy, houby a rostliny (Melbourne Code), přijatý osmnáctým mezinárodním botanickým kongresem Melbourne, Austrálie, červenec 2011 (elektronické vydání). Mezinárodní asociace pro taxonomii rostlin. Citováno 14. května 2017.
- ^ Walsh PJ, Smith S, Fleming L, Solo-Gabriele H, Gerwick WH, eds. (2. září 2011). „Sinice a sinice toxiny“. Oceány a lidské zdraví: Rizika a prostředky z moří. Akademický tisk. str. 271–296. ISBN 978-0-08-087782-2.
- ^ „The Rise of Oxygen - Astrobiology Magazine“. Astrobiologický časopis. 30. července 2003. Citováno 6. dubna 2016.
- ^ Flannery, D. T .; R.M. Walter (2012). „Archeanské všívané mikrobiální rohože a událost Velké oxidace: nové poznatky o starodávném problému“. Australian Journal of Earth Sciences. 59 (1): 1–11. Bibcode:2012AuJES..59 .... 1F. doi:10.1080/08120099.2011.607849. S2CID 53618061.
- ^ Rothschild, Lynn (září 2003). „Pochopte evoluční mechanismy a environmentální limity života“. NASA. Archivovány od originál dne 11. března 2012. Citováno 13. července 2009.
- ^ Nadis S (prosinec 2003). "Buňky, které vládnou mořím" (PDF). Scientific American. 289 (6): 52–3. Bibcode:2003SciAm.289f..52N. doi:10.1038 / scientificamerican1203-52. PMID 14631732. Archivovány od originál (PDF) dne 19. dubna 2014. Citováno 2. června 2019.
- ^ „Nejdůležitější mikrob, o kterém jste nikdy neslyšeli“. npr.org.
- ^ Flombaum, P .; Gallegos, J.L .; Gordillo, R. A .; Rincon, J .; Zabala, L. L .; Jiao, N .; Karl, D. M .; Li, W. K. W .; Lomas, M. W .; Veneziano, D .; Vera, C. S .; Vrugt, J. A .; Martiny, A. C. (2013). „Současná a budoucí globální distribuce mořských sinic Prochlorococcus a Synechococcus“. Sborník Národní akademie věd. 110 (24): 9824–9829. Bibcode:2013PNAS..110.9824F. doi:10.1073 / pnas.1307701110. PMC 3683724. PMID 23703908.
- ^ Nabors, Murray W. (2004). Úvod do botaniky. San Francisco, Kalifornie: Pearson Education, Inc. ISBN 978-0-8053-4416-5.
- ^ Allaby, M., ed. (1992). "Řasy". Stručný slovník botaniky. Oxford: Oxford University Press.
- ^ Guiry MD (říjen 2012). „Kolik druhů řas existuje?“. Journal of Phycology. 48 (5): 1057–63. doi:10.1111 / j.1529-8817.2012.01222.x. PMID 27011267. S2CID 30911529.
- ^ A b Guiry, M.D .; Guiry, G.M. (2016). "Algaebase". www.algaebase.org. Citováno 20. listopadu 2016.
- ^ D. Thomas (2002). Mořské řasy. Životní série. Muzeum přírodní historie, Londýn. ISBN 978-0-565-09175-0.
- ^ Hoek, Christiaan; den Hoeck, Hoeck Van; Mann, David; Jahns, H.M. (1995). Algae: an Introduction to Phycology. Cambridge University Press. str. 166. ISBN 9780521316873. OCLC 443576944.
- ^ Treguer, P .; Nelson, D. M .; Van Bennekom, A. J .; Demaster, D. J .; Leynaert, A .; Queguiner, B. (1995). „Bilance oxidu křemičitého ve světovém oceánu: Reestimate“. Věda. 268 (5209): 375–9. Bibcode:1995Sci ... 268..375T. doi:10.1126 / science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ „King's College London - Lake Megachad“. www.kcl.ac.uk. Citováno 5. května 2018.
- ^ Gómez F (2012). „Kontrolní seznam a klasifikace živých dinoflagelátů (Dinoflagellata, Alveolata)“ (PDF). CICIMAR Océanides. 27 (1): 65–140. Archivovány od originál (PDF) dne 27. listopadu 2013.
- ^ Stoecker DK (1999). "Mixotrofie mezi dinoflageláty". The Journal of Eukaryotic Microbiology. 46 (4): 397–401. doi:10.1111 / j.1550-7408.1999.tb04619.x. S2CID 83885629.
- ^ Starckx, Senne (31. října 2012) Místo na slunci - řasy jsou plodinou budoucnosti, tvrdí vědci z Geelu Flanders Today, Citováno 8. prosince 2012
- ^ Duval, B .; Margulis, L. (1995). "Mikrobiální komunita Ophrydium univerzální kolonie: endosymbionti, obyvatelé a nájemci ". Symbióza. 18: 181–210. PMID 11539474.
- ^ Wernberg, T., Krumhansl, K., Filbee-Dexter, K. a Pedersen, M.F. (2019) „Stav a trendy světových řas“. V: Světová moře: hodnocení životního prostředí, strany 57–78). Akademický tisk. doi:10.1016 / B978-0-12-805052-1.00003-6.
- ^ Mann, K.H. (1973). „Mořské řasy: jejich produktivita a strategie růstu“. Věda. 182 (4116): 975–981. Bibcode:1973Sci ... 182..975M. doi:10.1126 / science.182.4116.975. PMID 17833778. S2CID 26764207.
- ^ Tunnell, John Wesley; Chávez, Ernesto A .; Withers, Kim (2007). Korálové útesy jižního Mexického zálivu. Texas A&M University Press. str. 91. ISBN 978-1-58544-617-9.
- ^ Kompendium invazivních druhů: Caulerpa taxifolia (zabijácké řasy) Centrum pro zemědělství a biologickou vědu. Aktualizováno: 6. listopadu 2018.
- ^ Mandoli, DF (1998). „Vypracování tělesného plánu a fázové změny během vývoje Acetabularia: Jak je postavena složitá architektura obrovské Unicell?“. Roční přehled fyziologie rostlin a molekulární biologie rostlin. 49: 173–198. doi:10.1146 / annurev.arplant.49.1.173. PMID 15012232. S2CID 6241264.
- ^ Pierre Madl; Maricela Yip (2004). „Literature Review of Caulerpa taxifolia“. BUFUS-Info. 19 (31).
- ^ Orth, R.J .; Carruthers, T.J .; Dennison, W.C .; Duarte, C.M .; Fourqurean, J.W .; Heck, K.L .; Hughes, A.R .; Kendrick, GA; Kenworthy, W. J.; Olyarnik, S .; Krátký, F.T. (2006). „Globální krize pro ekosystémy mořských řas“. BioScience. 56 (12): 987–996. doi:10.1641 / 0006-3568 (2006) 56 [987: AGCFSE] 2.0.CO; 2. hdl:10261/88476.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2009). "Phycodurus eques" v FishBase. Verze z července 2009.
- ^ Giri, C; Ochieng, E; Tieszen, LL; Zhu, Z; Singh, A; Loveland, T; et al. (2011). "Stav a distribuce mangrovových lesů na světě s využitím satelitních dat pozorování Země". Globální ekologie a biogeografie. 20 (1): 154–159. doi:10.1111 / j.1466-8238.2010.00584.x.
- ^ Thomas, N .; Lucas, R .; Bunting, P .; Hardy, A .; Rosenqvist, A .; Simard, M. (2017). „Distribuce a hybné síly globálních změn mangrovových lesů, 1996–2010“. PLOS ONE. 12 (6): e0179302. Bibcode:2017PLoSO..1279302T. doi:10.1371 / journal.pone.0179302. PMC 5464653. PMID 28594908.
- ^ Krátký, F.T. a Frederick, T. (2003) Atlas světa mořských řas, University of California Press, strana 24. ISBN 9780520240476
- ^ Lalli, C .; Parsons, T. (1993). Biologická oceánografie: Úvod. Butterworth-Heinemann. ISBN 0-7506-3384-0.
- ^ Lindsey, R., Scott, M. a Simmon, R. (2010) „Co jsou to fytoplankton“. NASA Earth Observatory.
- ^ Field, C. B .; Behrenfeld, M. J .; Randerson, J. T .; Falwoski, P. G. (1998). „Primární produkce biosféry: integrace suchozemských a oceánských složek“. Věda. 281 (5374): 237–240. Bibcode:1998Sci ... 281..237F. doi:10.1126 / science.281.5374.237. PMID 9657713.
- ^ Rost, B. a Riebesell, U. (2004) „Coccolithophores and the biologic pump: responses to environment changes“. V: Coccolithophores: Od molekulárních procesů ke globálním dopadům, strany 99–125, Springer. ISBN 9783662062784.
- ^ Arsenieff, L .; Simon, N .; Rigaut-Jalabert, F .; Le Gall, F .; Chaffron, S .; Corre, E .; Přijít.; Bigeard, E .; Baudoux, A.C. (2018). „První viry infikující mořský diatom Guinardia delicatula“. Hranice v mikrobiologii. 9: 3235. doi:10.3389 / fmicb.2018.03235. PMC 6334475. PMID 30687251.
- ^ Varea, C .; Aragon, J.L .; Barrio, R.A. (1999). "Turingovy vzory na kouli". Fyzický přehled E. 60 (4): 4588–92. Bibcode:1999PhRvE..60,4588V. doi:10.1103 / PhysRevE.60.4588. PMID 11970318.
- ^ Harvey, Edmund Newton (1952). Bioluminiscence. Akademický tisk.
- ^ Navrhované vysvětlení pro zářící moře - včetně aktuálně zářícího kalifornského moře Národní vědecká nadace, 18. října 2011.
- ^ Castro P, Huber ME (2010). Mořská biologie (8. vydání). McGraw Hill. str.95. ISBN 978-0071113021.
- ^ Hastings JW (1996). "Chemické složení a barvy bioluminiscenčních reakcí: recenze". Gen. 173 (1 Spec No): 5–11. doi:10.1016/0378-1119(95)00676-1. PMID 8707056.
- ^ Haddock SH, Moline MA, Case JF (2009). „Bioluminiscence v moři“. Výroční zpráva o námořní vědě. 2: 443–93. Bibcode:2010 ARMS .... 2..443H. doi:10.1146 / annurev-marine-120308-081028. PMID 21141672. S2CID 3872860.
- ^ US Department of Energy (2008) Cyklus uhlíku a biosekvestrace strana 81, Workshop report DOE / SC-108, US Department of Energy Office of Science.
- ^ Campbell, Mike (22. června 2011). „Role mořského planktonu při sekvestraci uhlíku“. EarthTimes. Citováno 22. srpna 2014.
- ^ Roman, J. & McCarthy, J.J. (2010). „Velrybí pumpa: mořští savci zvyšují primární produktivitu v pobřežní pánvi“. PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO ... 513255R. doi:10.1371 / journal.pone.0013255. PMC 2952594. PMID 20949007. e13255.CS1 maint: používá parametr autoři (odkaz)
- ^ Brown, Joshua E. (12. října 2010). „Velrybí hovínko pumpuje zdraví oceánu“. Věda denně. Citováno 18. srpna 2014.
- ^ Thomson, Charles Wyville (2014) Voyage of the Challenger: The Atlantic Cambridge University Press, strana 235. ISBN 9781108074759.
- ^ Grethe R. Hasle; Erik E. Syvertsen; Karen A. Steidingerová; Karl Tangen (25. ledna 1996). „Marine Diatoms“. In Carmelo R. Tomas (ed.). Identifikace mořských rozsivek a dinoflagelátů. Akademický tisk. str. 5–385. ISBN 978-0-08-053441-1. Citováno 13. listopadu 2013.
- ^ Ald, S.M .; et al. (2007). „Rozmanitost, nomenklatura a taxonomie protistů“ (PDF). Syst. Biol. 56 (4): 684–689. doi:10.1080/10635150701494127. PMID 17661235. Archivovány od originál (PDF) dne 31. března 2011. Citováno 1. října 2019.
- ^ Moheimani, N.R .; Webb, J.P .; Borowitzka, M.A. (2012), „Bioremediace a další potenciální aplikace řas kokokolithophoridů: přehled. Výzkum řas, 1 (2): 120–133, doi:10.1016 / j.algal.2012.06.002
- ^ Taylor, A.R .; Chrachri, A .; Wheeler, G .; Goddard, H .; Brownlee, C. (2011). „Napěťově řízený kanál H +, který je základem homeostázy pH v kalcifikujících kokokolithophorech“. PLOS Biology. 9 (6): e1001085. doi:10.1371 / journal.pbio.1001085. PMC 3119654. PMID 21713028.
- ^ „Voda, univerzální rozpouštědlo“. USGS. Archivováno z původního dne 9. července 2017. Citováno 27. června 2017.
- ^ Brum JR, Morris JJ, Décima M a Stukel MR (2014) „Mortalita v oceánech: příčiny a důsledky“. Sborník Eco-DAS IX Symposium, Kapitola 2, strany 16-48. Sdružení pro vědu o limnologii a oceánografii. ISBN 978-0-9845591-3-8.
- ^ Reece, Jane B. (2013). Campbell Biology (10. vyd.). Pearson. ISBN 9780321775658.
- ^ Prentice, I.C. (2001). „Cyklus uhlíku a atmosférický oxid uhličitý“. Změna klimatu 2001: vědecký základ: příspěvek pracovní skupiny I ke třetí hodnotící zprávě Mezivládního panelu pro změnu klimatu / Houghton, J.T. [Upravit.] Citováno 31. května 2012.
- ^ Halpern, BS; Frazier, M .; Afflerbach, J .; et al. (2019). „Nedávné tempo změn v dopadu člověka na světový oceán“. Vědecké zprávy. 9 (1): 11609. Bibcode:2019NatSR ... 911609H. doi:10.1038 / s41598-019-47201-9. PMC 6691109. PMID 31406130.
- ^ Dopady člověka na mořské ekosystémy GEOMAR Helmholtz Center for Ocean Research. Vyvolány 22 October 2019.
- ^ Rosing, M .; Bird, D .; Spánek, N .; Bjerrum, C. (2010). "Žádný klimatický paradox pod slabým ranním Sluncem". Příroda. 464 (7289): 744–747. Bibcode:2010Natur.464..744R. doi:10.1038 / nature08955. PMID 20360739. S2CID 205220182.
- ^ A b Sahney, S .; Benton, M. J.; Ferry, Paul (2010). „Vazby mezi globální taxonomickou rozmanitostí, ekologickou rozmanitostí a expanzí obratlovců na souši“. Biologické dopisy. 6 (4): 544–7. doi:10.1098 / rsbl.2009.1024. PMC 2936204. PMID 20106856.
- ^ McKinney 1997, str.110
- ^ Stearns & Stearns 1999, str.X
- ^ Nováček, Michael J. (8. listopadu 2014). „Brilantní budoucnost pravěku“. The New York Times. New York: Společnost New York Times. ISSN 0362-4331. Citováno 25. prosince 2014.
- ^ Nee, S. (2004). „Extinction, slime, and bottoms“. PLOS Biology. 2 (8): E272. doi:10.1371 / journal.pbio.0020272. PMC 509315. PMID 15314670.
- ^ Ward, Peter D (2006). "Dopad z hlubin". Scientific American. 295 (4): 64–71. Bibcode:2006SciAm.295d..64W. doi:10.1038 / scientificamerican1006-64. PMID 16989482.
- ^ Sahney, S. & Benton, M.J. (2008). „Zotavení z nejhlubšího masového vyhynutí všech dob“ (PDF). Sborník Královské společnosti B: Biologické vědy. 275 (1636): 759–65. doi:10.1098 / rspb.2007.1370. PMC 2596898. PMID 18198148.
- ^ Desmond, Adrian (1975). „Objev mořských přestupků a vysvětlení fosilií ve starověku“. American Journal of Science. 275 (6): 692–697. Bibcode:1975AmJS..275..692D. doi:10,2475 / ajs.275.6.692.
- ^ McKirahan, Richard D. „Xenophanes of Colophon. Filozofie před Sokratem. Indianapolis: Hackett Publishing Company, 1994. 66. Tisk.
- ^ Grene MG, Grene M a Depew D (2004) Filozofie biologie: Epizodická historie strany 1–34, Cambridge University Press. ISBN 9780521643801.
- ^ Boylan, Michael "Aristoteles: biologie" Internetová encyklopedie filozofie. Vyvolány 5 August je 2016.
- ^ Lee, H. D. P. (1948) „Místní jména a datum Aristotelových biologických prací“. Klasická čtvrtletní, 42 (3/4) :61–7
- ^ Zpěvák, Charles. Krátká historie biologie. Oxford 1931.
- ^ Emily Kearns, „Zvířata, znalosti o“, v Oxfordský klasický slovník, 3. vydání, 1996, s. 92.
- ^ Carl T. Bergstrom; Lee Alan Dugatkin (2012). Vývoj. Norton. str. 35. ISBN 978-0-393-92592-0.
- ^ Rhodes, Frank Harold Trevor (1. ledna 1974). Vývoj. Golden Press. str. 7. ISBN 978-0-307-64360-5.
- ^ A b C Lalli, Carol M. a Timothy R. Parsons. "Úvod." Biologická oceánografie: Úvod. První vydání ed. Tarrytown, New York: Pergamon, 1993. 7-21. Tisk.
- ^ Menden-Deuer, Susanne. „Informace o kurzu, biologická oceánografie OCG 561“.
- ^ Miller, Charles B .; Patricia A. Wheeler (2012). Biologická oceánografie (Druhé vydání.). Chinchester, West Sussex: John Wiley & Sons.
Poznámky
Další reference
- Halpern, BS; Walbridge, S .; Selkoe, K.A .; Kappel, C.V .; Micheli, F .; D'Agrosa, C .; Bruno, J.F .; Casey, K. S.; Ebert, C .; Fox, HE; Fujita, R. (2008). „Globální mapa dopadu člověka na mořské ekosystémy“. Věda. 319 (5865): 948–952. Bibcode:2008Sci ... 319..948H. doi:10.1126 / science.1149345. PMID 18276889. S2CID 26206024.
- Paleczny, M .; Hammill, E .; Karpouzi, V .; Pauly, D. (2015). „Populační trend sledovaných mořských ptáků na světě, 1950–2010“. PLOS ONE. 10 (6): e0129342. Bibcode:2015PLoSO..1029342P. doi:10.1371 / journal.pone.0129342. PMC 4461279. PMID 26058068.
- Po 60 milionech let extrémního života havarují mořští ptáci Opatrovník, 22. září 2015.
- Ruppert, E.E .; Fox, R.S. & Barnes, RD (2004). Zoologie bezobratlých (7. vydání). Brooks / Cole. ISBN 978-0-03-025982-1.

