Ciliate - Ciliate
| Ciliate | |
|---|---|
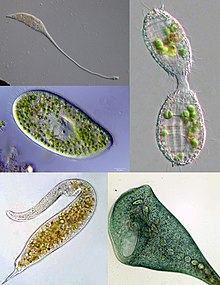 | |
| Některé příklady ciliate rozmanitosti. Ve směru hodinových ručiček zleva nahoře: Lacrymaria, Coleps, Stentor, Dileptus, Paramecium | |
| Vědecká klasifikace | |
| Clade: | SAR |
| Infrakingdom: | Alveolata |
| Kmen: | Ciliophora Doflein, 1901 opravit. |
| Subfyla a třídy[1] | |
Viz text pro podtřídy. | |
| Synonyma | |
| |
The náčelníci jsou skupina prvoky charakterizována přítomností tzv. vlasových organel řasy, které mají stejnou strukturu jako eukaryotické bičíky, ale jsou obecně kratší a jsou přítomny v mnohem větším počtu, s jiným zvlněný vzor než bičíky. Cilia se vyskytují u všech členů skupiny (i když zvláštní Suctoria mít je jen pro část jejich životní cyklus ) a různě se používají při plavání, plazení, připoutání, krmení a senzaci.
Ciliates jsou důležitou skupinou protistů, běžné téměř všude, kde je voda - v jezerech, rybnících, oceánech, řekách a půdách. Bylo popsáno asi 4 500 jedinečných volně žijících druhů a potenciální počet existujících druhů se odhaduje na 27 000–40 000.[2] V tomto počtu je zahrnuto mnoho ektosymbiotikum a endosymbiotikum druhy, stejně jako některé povinná a oportunistický paraziti. Ciliate druhy se pohybují ve velikosti od pouhých 10 µm v některých colpodeans u některých až 4 mm dlouhé geleiidy a zahrnují některé z nejvíce morfologicky složité prvoky.[3][4]
Ve většině systémů taxonomie, "Ciliophora"je zařazen jako kmen[5] pod některým z několika království, počítaje v to Chromista,[6] Protista[7] nebo Prvoci.[8] V některých starších systémech klasifikace, jako jsou vlivné taxonomické práce z Alfred Kahl, řasinkaté prvoky jsou umístěny uvnitř třída "Ciliata"[9][10] (termín, který může také odkazovat na a rod ryb ). V taxonomickém schématu schváleném Mezinárodní společnost protistologů, což vylučuje formální hodnost označení jako „kmen“ a „třída“ je „Ciliophora“ nepřiřazená taxon v rámci Alveolata.[11][12]
Struktura buněk

Nuclei
Na rozdíl od většiny ostatních eukaryoty náčelníci mají dva různé druhy jádra: malý, diploidní mikrojader ("generativní jádro", které nese zárodečná linie buňky) a velký, polyploidní makronukleus ("vegetativní jádro", které se stará o obecnou buněčnou regulaci, vyjadřující fenotyp organismu). Ten je generován z mikrojader zesílením genom a těžké úpravy. Mikrojader předává svůj genetický materiál potomkům, ale nevyjadřuje své geny. Makronukleus poskytuje malá jaderná RNA pro vegetativní růst.
K rozdělení makronukleu dochází amitóza, a segregace chromozomy dochází procesem, jehož mechanismus není znám. Tento proces není dokonalý a po přibližně 200 generacích vykazuje buňka známky stárnutí. Makronukleá musí být pravidelně regenerována z mikrojader. K tomu většinou dochází během časování. Zde se dvě buňky seřadí, mikrojader podstoupí redukční dělení buněk, některé z haploidní dcery jsou vyměněny a poté se spojí, aby vytvořily nová mikrojader a makronukleů.
Cytoplazma
Jídlo vakuoly jsou tvořeny skrz fagocytóza a obvykle následují určitou cestu buňkou, když je jejich obsah tráven a rozdělen podle lysozomy takže látky, které vakuola obsahuje, jsou potom dostatečně malé na to šířit přes membránu potravinové vakuoly do buňky. Všechno, co zbylo v potravinové vakuole v době, kdy dosáhne cytoproctu (anální pór ) je vybíjen exocytóza. Většina náměstků má také jednoho nebo více prominentních kontraktilní vakuoly, které shromažďují vodu a vylučují ji z buňky, aby ji udržovaly osmotický tlak, nebo v nějaké funkci k udržení iontové rovnováhy. V některých rodech, jako např Paramecium, tyto mají výrazný tvar hvězdy, přičemž každý bod je sběrnou trubicí.
Specializované struktury v náměstcích
Cilia jsou uspořádány do řádků s názvem kineties. V některých formách existují například také polykinetidy těla spirotrichs kde obecně tvoří štětiny zvané cirri. Častěji jsou řasinky uspořádány mono- a dikinetidy, které zahrnují jednu a dvě kinetosomy (bazální těla), z nichž každé může podporovat cilium. Jsou uspořádány do řádků s názvem kineties, které probíhají od přední k zadní části buňky. Tělo a orální kinetidy tvoří infraciliature, organizace jedinečná pro nálevníky a důležitá při jejich klasifikaci, a zahrnuje různé fibrily a mikrotubuly podílí se na koordinaci řasinek.
Infračervené záření je jednou z hlavních složek systému buněčná kůra. Jiní jsou alveoly, malé vezikuly pod buněčnou membránou, které jsou zabaleny proti ní a tvoří pelikula zachování tvaru buňky, který se liší od pružného a stahujícího se po tuhý. Četné mitochondrie a extrusomy jsou také obecně přítomny. Přítomnost plicních sklípků, struktura řasinek, forma mitózy a různé další podrobnosti naznačují blízký vztah mezi nálevníky, Apicomplexa, a dinoflageláty. Tyto povrchně odlišné skupiny tvoří alveoláty.
Krmení
Většina náměstků je heterotrofy, krmení menšími organismy, jako je bakterie a řasy a detritus zametl do ústní drážky (úst) modifikovanou ústní řasinkou. To obvykle zahrnuje řadu membrány nalevo od úst a parorální membrána po pravé straně, z nichž obě vycházejí polykinetidy, skupiny mnoha řasinek spolu s přidruženými strukturami. Jídlo se řasinkami pohybuje ústními póry do jícnu, který vytváří potravinové vakuoly.
Techniky krmení se však značně liší. Někteří nálevníci jsou bez úst a krmí se absorpcí (osmotrofie ), zatímco jiné jsou dravé a živí se jinými prvoky a zejména jinými nálevníky. Někteří nálevníci parazitují zvířata, i když pouze jeden druh, Balantidium coli Je známo, že způsobuje onemocnění u lidí.[13]
Reprodukce a sexuální jevy
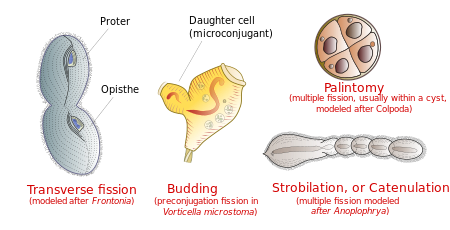
Reprodukce
Ciliates se množí nepohlavně, různými druhy štěpení.[14] Během štěpení prochází mikrojader mitóza a makronukleus se prodlužuje a prochází amitóza (kromě mezi Karyorelictean nálevníky, jejichž makrojádra se nerozdělují). Buňka se poté rozdělí na dvě a každá nová buňka získá kopii mikrojader a makronukleů.

Buňka je obvykle rozdělena příčně na přední polovina nálevníka ( proter) formující jeden nový organismus a zadní polovina ( opisthe) tvořící další. U některých skupin nálevníků se však vyskytují jiné typy štěpení. Tyto zahrnují pučící (vznik malých řasinkatých potomků neboli „rojů“ z těla zralého rodiče); strobilace (vícenásobné dělení podél těla buňky, produkující řetězec nových organismů); a palintomie (vícenásobné štěpení, obvykle v a cysta ).[15]
Štěpení může nastat spontánně, jako součást vegetativního buněčný cyklus. Alternativně může postupovat v důsledku samooplodnění (autogamie ),[16] nebo to může následovat časování, sexuální fenomén, ve kterém jsou nálevníci kompatibilní typy páření vyměňovat genetický materiál. Zatímco konjugace je někdy popisována jako forma reprodukce, není přímo spojena s reprodukčními procesy a nevede přímo ke zvýšení počtu jednotlivých nálevníků nebo jejich potomků.[17]
Časování
- Přehled
Ciliate konjugace je sexuální fenomén, jehož výsledkem je genetická rekombinace a jaderná reorganizace v buňce.[17][15] Během konjugace tvoří dva nálevníci kompatibilního typu páření most mezi nimi cytoplazmy. Mikrojader podstoupí redukční dělení buněk, makronukleá zmizí a haploidní mikrojádra jsou vyměňována po můstku. U některých nálevníků (peritrichů, chonotrichové a nějaký suctorians ), konjugační buňky se trvale spojí a jeden konjugant je absorbován druhým.[13][18] U většiny nálevníkových skupin se však buňky po konjugaci oddělí a obě tvoří nová mikrojader ze svých mikrojader.[19] Po konjugaci a autogamii vždy následuje štěpení.[15]
V mnoha náměstcích, jako např Paramecium, konjugační partneři (gamoti) jsou podobné nebo nerozeznatelné co do velikosti a tvaru. Toto se označuje jako "isogamontická" konjugace. V některých skupinách se partneři liší velikostí a tvarem. Toto se označuje jako "anisogamontická" konjugace. v přisedlé peritrichy například jeden sexuální partner (mikrokonjugant) je malý a mobilní, zatímco druhý (makrokonjugant) je velký a přisedlý.[17]
- Fáze konjugace
 Fáze konjugace v Paramecium caudatum
Fáze konjugace v Paramecium caudatum

v Paramecium caudatum, fáze konjugace jsou následující (viz obrázek vpravo):
- Kompatibilní páření se setkávají a částečně fúzují
- Mikrojader podstoupí meiózu a produkují čtyři haploidní mikrojader na buňku.
- Tři z těchto mikrojader se rozpadají. Čtvrtý prochází mitózou.
- Tyto dvě buňky si vyměňují mikrojader.
- Buňky se poté oddělí.
- Mikrojader v každé buňce fúzují a tvoří diploidní mikrojader.
- Mitóza se vyskytuje třikrát, což vede k osmi mikrojaderům.
- Čtyři z nových mikrojader se transformují do makronukleů a starý makronukle se rozpadá.
- Binární štěpení nastává dvakrát, čímž se získají čtyři identické dceřiné buňky.
Přesmyky DNA (kódování genů)
Ciliates obsahují dva typy jader: somatický "makronukleus "a zárodečná linie "mikrojader ". Během pohlavní reprodukce (konjugace) se předává pouze DNA v mikronukleu. Na druhé straně je aktivně exprimována pouze DNA v makronukleu a vede k fenotypu organismu. Makronukleární DNA je z mikronukleární DNA odvozena úžasně rozsáhlé přeskupení a amplifikace DNA.
Makronukleus začíná jako kopie mikrojader. Mikronukleární chromozomy jsou fragmentovány na mnoho menších kousků a zesíleny, aby poskytly mnoho kopií. Výsledné makronukleární chromozomy často obsahují pouze jeden gen. v Tetrahymena, mikronukle má 10 chromozomů (pět na haploidní genom), zatímco makronukle má více než 20 000 chromozomů.[20]
Kromě toho jsou mikronukleární geny přerušovány četnými „interně eliminovanými sekvencemi“ (IES). Během vývoje makronukleu jsou IES odstraněny a zbývající genové segmenty, makronukleární destinační sekvence (MDS), jsou spojeny dohromady, aby poskytly funkční gen. Tetrahymena má asi 6 000 IES a asi 15% mikronukleární DNA je během tohoto procesu eliminováno. Proces se řídí malé RNA a epigenetický chromatin známky.[20]
v spirotrich nájemci (např Oxytricha ), proces je ještě složitější díky „genovému zakódování“: MDS v mikronukleu jsou často v jiném pořadí a orientaci než v makronukleárním genu, a tak kromě delece DNA inverze a přemístění jsou vyžadovány pro „dešifrování“. Tento proces je veden dlouhými RNA odvozenými z rodičovského makronukleu. Více než 95% mikronukleární DNA je eliminováno během vývoje makronukleárního spirotrichu.[20]
Fosilní záznam
Až donedávna byly známé nejstarší fosilie fosilií tintinnids z Ordovické období. V roce 2007 Li et al. zveřejnil popis fosilních nálevníků z Formace Doushantuo, asi před 580 miliony let, v Ediacaranské období. Patřily mezi ně dva typy tintinnidů a možný rodový suctorian.[21] Fosílie Vorticella byl objeven uvnitř kokonu pijavice z Triasové období, asi před 200 miliony let.[22]
Klasifikace

Pro nálevníky bylo navrženo několik různých klasifikačních schémat. Následující schéma je založeno na molekule fylogenetická analýza až čtyř genů ze 152 druhů představujících 110 rodin:[1]
Subphylum Postciliodesmatophora
- Třída Heterotrichea (např. Stentor )
- Třída Karyorelictea
Subphylum Intramacronucleata

- Třída Armophorea
- Třída Cariacotrichea (pouze jeden druh, Cariacothrix caudata )
- Třída Muranotrichea
- Třída Parablepharismea
- Třída Colpodea (např. Colpoda )
- Třída Litostomatea
- Podtřída Haptoria (např. Didinium )
- Podtřída Rhynchostomatia
- Podtřída Trichostomatia (např. Balantidium )
- Třída Nassophorea
- Třída Phyllopharyngea
- Podtřída Chonotrichia
- Podtřída Cyrtophoria
- Podtřída Rhynchodia
- Podtřída Suctoria (např. Podophyra )
- Podtřída Synhymenie
- Třída Oligohymenophorea
- Podtřída Apostomatia
- Podtřída Astomatia
- Podtřída Hymenostomatia (např. Tetrahymena )
- Podtřída Peniculia (např. Paramecium )
- Podtřída Peritrichia (např. Vorticella )
- Podtřída Scuticociliatia
- Třída Plagiopylea
- Třída Prostomatea (např. Coleps )
- Třída Protocruziea
- Třída Spirotrichea
- Podtřída Choreotrichia
- Podtřída Euplotia
- Podtřída Hypotrichia
- Podtřída Licnophoria
- Podtřída Oligotrichia
- Podtřída Phacodiniidea
- Podtřída Protohypotrichia
jiný
Některé staré klasifikace zahrnuty Opalinidae v náměstcích. Základní rozdíl mezi multiciliate bičíky (např., hemimastigidy, Stephanopogon, Multicilia, opaliny ) a nálevníky je přítomnost makronukleů v nálevnících samotných.[23]
Patogenita
Jediný člen kmene nálevníků, o kterém je známo patogenní pro lidi je Balantidium coli,[24][25] který způsobuje nemoc balantidiáza. Pro domácí prase není patogenní.[26]
Reference
- ^ A b Gao, Feng; Warren, Alan; Zhang, Qianqian; Gong, červen; Miao, Miao; Slunce, Ping; Xu, Dapeng; Huang, Jie; Yi, Zhenzhen (2016-04-29). „Evoluční hypotéza řasených protistů založená na všech datech s revidovanou klasifikací kmene Ciliophora (Eukaryota, Alveolata)“. Vědecké zprávy. 6: 24874. Bibcode:2016NatSR ... 624874G. doi:10.1038 / srep24874. ISSN 2045-2322. PMC 4850378. PMID 27126745.
- ^ Foissner, W .; Hawksworth, David, eds. (2009). Protist rozmanitost a zeměpisné rozdělení. Témata v oblasti biologické rozmanitosti a ochrany. Springer Nizozemsko. str. 111. ISBN 9789048128006.
- ^ Nielsen, Torkel Gissel; Kiørboe, Thomas (1994). „Regulace biomasy zooplanktonu a produkce v mírném pobřežním ekosystému. 2. Ciliates“. Limnologie a oceánografie. 39 (3): 508–519. Bibcode:1994LimOc..39..508N. doi:10.4319 / lo.1994.39.3.0508.
- ^ Lynn, Denis (2008). Ciliated Protozoa 3. vydání. Springer. str.129. ISBN 978-1-4020-8238-2.
- ^ „Zpráva ITIS“. Integrovaný taxonomický informační systém. Citováno 11. prosince 2014.
- ^ Cavalier-Smith, Thomas (01.01.2018). „Kingdom Chromista a jeho osm kmenů: nová syntéza s důrazem na cílení periplastidových proteinů, vývoj cytoskeletů a periplastidů a starověké divergence“. Protoplasma. 255 (1): 297–357. doi:10.1007 / s00709-017-1147-3. ISSN 1615-6102. PMC 5756292. PMID 28875267.
- ^ Yi Z, Song W, Clamp JC, Chen Z, Gao S, Zhang Q (prosinec 2008). „Přehodnocení systematických vztahů v řádu Euplotida (Protista, Ciliophora) s využitím nových sekvencí genu kódujícího rRNA s malou podjednotkou a testování použití kombinovaných datových souborů pro konstrukci fylogenetů komplexu Diophrys“. Mol. Fylogenet. Evol. 50 (3): 599–607. doi:10.1016 / j.ympev.2008.12.006. PMID 19121402.
- ^ Miao M, Song W, Chen Z a kol. (2007). „Unikátní euplotidní nálevník, Gastrocirrhus (Protozoa, Ciliophora): hodnocení jeho fylogenetické polohy odvozené z genové sekvence rRNA malé podjednotky“. J. Eukaryot. Microbiol. 54 (4): 371–8. doi:10.1111 / j.1550-7408.2007.00271.x. PMID 17669163.
- ^ Alfred Kahl (1930). Urtiere oder Protozoa I: Wimpertiere oder Ciliata - Svazek I Obecná část a Prostomata.
- ^ „Lékařská definice CILIATA“. www.merriam-webster.com. Citováno 2017-12-22.
- ^ Adl, Sina M .; Bass, David; Lane, Christopher E .; Lukeš, Julius; Schoch, Conrad L .; Smirnov, Alexey; Agatha, Sabine; Berney, Cedric; Brown, Matthew W .; Burki, Fabien; Cárdenas, Paco (2019). „Revize klasifikace, nomenklatury a rozmanitosti eukaryot“. Journal of Eukaryotic Microbiology. 66 (1): 4–119. doi:10.1111 / jeu.12691. ISSN 1550-7408. PMC 6492006. PMID 30257078.
- ^ Adl, Sina M .; et al. (2005). „Nová vyšší klasifikace eukaryot s důrazem na taxonomii protistů“. Journal of Eukaryotic Microbiology. 52 (5): 399–451. doi:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873.
- ^ A b Lynn, Denis (2008). The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature (3. vyd.). Springer. str.58. ISBN 978-1-4020-8238-2.
1007/978-1-4020-8239-9
- ^ H., Lynn, Denis (2008). Přidružené prvoky: charakteristika, klasifikace a průvodce literaturou. New York: Springer. str. 30. ISBN 9781402082382. OCLC 272311632.
- ^ A b C H., Lynn, Denis (2008). Přidružené prvoky: charakteristika, klasifikace a průvodce literaturou. New York: Springer. str. 23. ISBN 9781402082382. OCLC 272311632.
- ^ Berger JD (říjen 1986). „Autogamie v parameciu. Specifická angažovanost meiózy ve fázi buněčného cyklu“. Exp. Cell Res. 166 (2): 475–85. doi:10.1016/0014-4827(86)90492-1. PMID 3743667.
- ^ A b C Raikov, I.B (1972). „Jaderné jevy během konjugace a autogamie u nálevníků“. Výzkum v protozoologii. 4: 149.
- ^ Finley, Harold E. "Konjugace Microstoma vorticella"Transaction of the American Microscopical Society (1943): 97-121.
- ^ „Úvod do Ciliaty“. Citováno 2009-01-16.
- ^ A b C Mochizuki, Kazufumi (2010). „Přesmyky DNA řízené nekódujícími RNA v nálevnících“. Wiley Interdisciplinary Reviews: RNA. 1 (3): 376–387. doi:10.1002 / wrna.34. PMC 3746294. PMID 21956937.
- ^ Li, C.-W .; et al. (2007). "Ciliated protozoans from the Precambrian Doushantuo Formation, Wengan, South China". Geologická společnost, Londýn, speciální publikace. 286 (1): 151–156. Bibcode:2007GSLSP.286..151L. doi:10.1144 / SP286.11.
- ^ Bomfleur, Benjamin; Kerp, Hans; Taylor, Thomas N .; Moestrup, Øjvind; Taylor, Edith L. (2012-12-18). „Koks pijavice triasové z Antarktidy obsahuje zvíře s fosilním zvonem“. Sborník Národní akademie věd Spojených států amerických. 109 (51): 20971–20974. Bibcode:2012PNAS..10920971B. doi:10.1073 / pnas.1218879109. ISSN 1091-6490. PMC 3529092. PMID 23213234.
- ^ Cavalier-Smith, T. (2000). Flagellate megaevolution: základ pro diverzifikaci eukaryot. In: Leadbeater, B.S.C., Green, J.C. (eds.). Bičíkovci. Jednota, rozmanitost a evoluce. London: Taylor and Francis, str. 361-390, str. 362, [1].
- ^ "Balantidiasis". DPDx - Laboratorní identifikace parazitárních chorob v oblasti veřejného zdraví. Centra pro kontrolu a prevenci nemocí. 2013.
- ^ Ramachandran, Ambili (23. května 2003). "Úvod". Parazit: Balantidium coli Nemoc: Balantidiasis. Parazit. Stanfordská Univerzita.CS1 maint: ref = harv (odkaz)
- ^ Schister, Frederick L. a Lynn Ramirez-Avila (říjen 2008). "Současný světový stav Balantidium coli". Recenze klinické mikrobiologie. 21 (4): 626–638. doi:10.1128 / CMR.00021-08. PMC 2570149. PMID 18854484.
Další čtení
- Lynn, Denis H. (2008). Přidružené prvoky: charakteristika, klasifikace a průvodce literaturou. New York: Springer. ISBN 9781402082382. OCLC 272311632.
- Ciliates: buňky jako organismy. Hausmann, Klaus., Bradbury, Phyllis C. (Phyllis Clarke). Stuttgart: Gustav Fischer Verlag. 1996. ISBN 978-3437250361. OCLC 34782787.CS1 maint: ostatní (odkaz)
- Ilustrovaný průvodce k prvokům: organismy tradičně označované jako prvoky nebo nově objevené skupiny. Lee, John J., Leedale, Gordon F., Bradbury, Phyllis C. (Phyllis Clarke) (2. vyd.). Lawrence, Kan., USA: Společnost protozoologů. 2000. ISBN 9781891276224. OCLC 49191284.CS1 maint: ostatní (odkaz)
| Mikroskopické objevy1 |
|  | ||||
|---|---|---|---|---|---|---|
| Obecná témata | ||||||
| související témata |
| |||||
| Související lidé |
| |||||
| Uznání | ||||||
| ||||||