Hlubinné ryby - Deep sea fish

Hlubinné ryby jsou Ryba kteří žijí ve tmě pod sluncem zalitými povrchovými vodami, tj. pod epipelagický nebo fotonická zóna z moře. The lucerna je zdaleka nejběžnější hlubinná ryba. jiný hluboké moře ryby zahrnují baterka na ryby, kuchařský žralok, bristlemouths, ďas mořský, zmije a některé druhy úhoř.
Pouze asi 2% známých mořských druhů obývají pelagický životní prostředí. To znamená, že žijí v vodní sloup na rozdíl od bentický organismy, které žijí na mořském dně nebo na něm.[1] Hlubinné organismy obecně žijí bathypelagic (Hloubka 1 000–4 000 m) a abyssopelagic (4000–6000 m hluboké) zóny. Avšak charakteristiky hlubinných organismů, jako jsou bioluminiscence lze vidět v mezopelagický (200–1 000 m hluboká) zóna. Mezopelagická zóna je zóna fotografická, což znamená, že světlo je minimální, ale stále měřitelné. Minimální vrstva kyslíku existuje někde mezi hloubkou 700 m a 1000 m hluboko v závislosti na místě v oceánu. V této oblasti se také nachází nejvíce živin. The bathypelagic a abyssopelagic zóny jsou aphotický, což znamená, že do této oblasti oceánu neproniká žádné světlo. Tyto zóny tvoří asi 75% obyvatelného oceánského prostoru.[2]
Epipelagická zóna (0–200 m) je oblast, kde světlo proniká do vody a dochází k fotosyntéze. Toto je také známé jako fotonická zóna. Protože se obvykle rozprostírá jen několik set metrů pod vodou, je hluboké moře, přibližně 90% objemu oceánu, ve tmě. Hluboké moře je také extrémně nepřátelským prostředím s teplotami, které zřídka přesahují 3 ° C (37,4 ° F) a klesají až na -1,8 ° C (28,76 ° F) (s výjimkou hydrotermálních větracích ekosystémů, které mohou překročit 350 ° C nebo 662 ° F), nízké hladiny kyslíku a tlaky mezi 20 a 1 000 atmosféry (mezi 2 a 100 megapascalů ).[3]
životní prostředí

V hlubokém oceánu se vody táhnou hluboko pod epipelagickou zónu a podporují velmi odlišné druhy pelagických ryb přizpůsobených pro život v těchto hlubších zónách.[4]
V hluboké vodě, mořský sníh je nepřetržitá sprcha převážně organická detritus padající z horních vrstev vodního sloupce. Jeho původ spočívá v činnostech v rámci produktivního fotonická zóna. Mořský sníh zahrnuje mrtvé nebo umírající plankton, protistů (rozsivky ), fekální látky, písek, saze a jiný anorganický prach. "Sněhové vločky" rostou v průběhu času a mohou dosáhnout průměru několika centimetrů, cestovat několik týdnů, než se dostanou na dno oceánu. Většina organických složek mořského sněhu je však spotřebována mikroby, zooplankton a další zvířata živící se filtrem během prvních 1 000 metrů jejich cesty, tj. v epipelagické zóně. Tímto způsobem lze mořský sníh považovat za základ hlubinného moře mezopelagický a bentický ekosystémy: Vzhledem k tomu, že se k nim sluneční světlo nemůže dostat, hlubinné organismy se do značné míry spoléhají na mořský sníh jako zdroj energie. Vzhledem k tomu, že v hlubokém moři není žádné světlo (aphotické), chybí zde primární producenti. Proto se většina organismů v bathypelagice spoléhá na mořský sníh z oblastí výše ve vertikálním sloupci.
Některé hlubinné pelagické skupiny, například lucerna, Ridgehead, mořské líhně, a lightfish rodiny se někdy nazývají pseudoceanic protože spíše než aby měli rovnoměrnou distribuci v otevřené vodě, vyskytují se ve výrazně vyšších množstvích kolem strukturních oáz, zejména podmořské hory a znovu kontinentální svahy. Tento jev je vysvětlen obdobným množstvím druhů kořisti, které jsou také přitahovány strukturami.
Hydrostatický tlak se zvyšuje o 1 atmosféru na každých 10 metrů do hloubky.[5] Hlubinné organismy mají ve svých tělech stejný tlak, jaký na ně působí zvenčí, takže nejsou extrémním tlakem rozdrceni. Jejich vysoký vnitřní tlak má však za následek sníženou tekutost jejich membrán, protože molekuly jsou stlačovány k sobě. Tekutost v buněčných membránách zvyšuje účinnost biologických funkcí, zejména produkci bílkovin, takže se organismy přizpůsobily této situaci zvýšením podílu nenasycených mastných kyselin v lipidech buněčných membrán.[6] Kromě rozdílů ve vnitřním tlaku si tyto organismy vyvinuly jinou rovnováhu mezi svými metabolickými reakcemi od těch organismů, které žijí v epipelagické zóně. David Wharton, autor Život na hranici možností: Organismy v extrémním prostředíPoznámky: „Biochemické reakce jsou doprovázeny změnami objemu. Pokud má reakce za následek zvětšení objemu, bude potlačena tlakem, zatímco, pokud je spojena se zmenšením objemu, zesílí se“.[7] To znamená, že jejich metabolické procesy musí do určité míry nakonec snížit objem organismu.
Většina ryb, které se vyvinuly v tomto drsném prostředí, nejsou schopny přežít v laboratorních podmínkách a pokusy o jejich udržení v zajetí vedly k jejich smrti. Hlubinné organismy obsahují plyny naplněné prostory (vakuoly).[9] Plyn je stlačován pod vysokým tlakem a expanduje pod nízkým tlakem. Z tohoto důvodu je známo, že tyto organismy vybuchují, pokud se dostanou na povrch.[7]
Vlastnosti
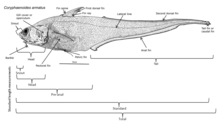



Ryby hlubinných ryb si vyvinuly různé adaptace, aby přežily v této oblasti. Protože mnoho z těchto ryb žije v regionech, kde není přirozené osvětlení, nemohou se spolehnout pouze na svůj zrak při hledání kořisti a kamarádů a vyhýbání se predátorům; hlubinné ryby mají vyvinul přiměřeně extrémní subfotické oblasti, ve které žijí. Mnoho z těchto organismů je slepých a spoléhají na své další smysly, jako je citlivost na změny místního tlaku a vůně, chytání potravy a vyhýbání se tomu, aby byli chyceni. Ti, kteří nejsou slepí, mají velké a citlivé oči, které mohou použít bioluminiscenční světlo. Tyto oči mohou být až 100krát citlivější na světlo než lidské oči. Rhodopsin (Rh1) je protein nacházející se v očních buňkách, který pomáhá zvířatům vidět za tlumeného světla. Zatímco většina obratlovců obvykle má jeden gen opsinu Rh1, některé hlubinné ryby mají několik genů Rh1 a jeden druh, stříbrný spinyfin (Diretmus argenteus), má 38.[10] Toto množení genů Rh1 může pomoci hlubinným rybám vidět v hlubinách oceánu. Aby se zabránilo predaci, je mnoho druhů temných, aby splynuly s prostředím.[11]
Mnoho hlubinných ryb je bioluminiscenční, s extrémně velkýma očima přizpůsobeným tmě. Bioluminiscenční organismy jsou schopné produkovat světlo biologicky mícháním molekul luciferinu, které pak produkují světlo. Tento proces musí být proveden v přítomnosti kyslíku. Tyto organismy jsou běžné v mezopelagické oblasti a níže (200 m a níže). Více než 50% hlubinných ryb, stejně jako některé druhy krevet a chobotnic, je schopno bioluminiscence. Asi 80% těchto organismů má fotofory - světlo produkující žlázové buňky, které obsahují světelné bakterie ohraničené tmavými barvami. Některé z těchto fotoforů obsahují čočky, podobné čočkám v očích lidí, které mohou zesílit nebo snížit vyzařování světla. Schopnost produkovat světlo vyžaduje pouze 1% energie organismu a má mnoho účelů: Používá se k hledání potravy a přitahování kořisti, jako je ďas; nárokovat si území hlídkou; komunikovat a najít partnera a rozptýlit nebo dočasně oslepit predátory, aby unikli. Také v mezopelagice, kde stále proniká nějaké světlo, se některé organismy maskují před predátory pod nimi osvětlením svých břich, aby odpovídaly barvě a intenzitě světla shora, takže nevrhá žádný stín. Tato taktika je známá jako protisvětlo.[12]
Životní cyklus hlubinných ryb může být výhradně hluboká voda, i když některé druhy se rodí v mělčí vodě a po zrání se potápí. Bez ohledu na hloubku, kde se vajíčka a larvy nacházejí, jsou obvykle pelagické. Tento životní styl planktonu - driftování - vyžaduje neutrální vztlak. V zájmu zachování toho vajíčka a larvy často obsahují kapičky oleje ve své plazmě.[13] Když jsou tyto organismy v plně zralém stavu, potřebují další úpravy, aby si udržely své pozice ve vodním sloupci. Obecně platí, že hustota vody způsobuje vztlak - aspekt vztlaku, díky němuž se organismy vznášejí. Aby se tomu zabránilo, musí být hustota organismu větší než hustota okolní vody. Většina živočišných tkání je hustších než voda, takže musí najít rovnováhu, aby se vznášela.[14] Mnoho organismů vyvíjí plavecké měchýře (plynové dutiny), aby zůstaly na hladině, ale kvůli vysokému tlaku svého prostředí hlubinné ryby tento orgán obvykle nemají. Místo toho vykazují struktury podobné křídlovým křídlům, aby poskytly hydrodynamický vztlak. Bylo také zjištěno, že čím hlouběji ryba žije, tím je její maso více želé a tím menší je její kostní struktura. Snižují svou hustotu tkání vysokým obsahem tuku, snížením hmotnosti kostry - docíleno zmenšením velikosti, tloušťky a obsahu minerálů - a akumulací vody [15] díky nim jsou pomalejší a méně hbití než povrchové ryby.
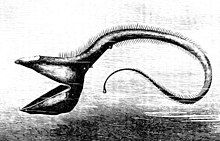
Kvůli špatné úrovni fotosyntetický světlo dosahující hlubinných prostředí, na které se většina ryb musí spolehnout organický hmota klesající z vyšších úrovní, nebo ve vzácných případech hydrotermální průduchy pro živiny. Díky tomu je hlubinný oceán mnohem chudší produktivita než mělčí oblasti. Zvířata v pelagickém prostředí jsou také řídká a jídlo se neobjevuje často. Z tohoto důvodu organismy potřebují úpravy, které jim umožní přežít. Někteří mají dlouhé tykadla, aby jim pomohli najít kořist nebo přilákat kamarády v černohnědé hlubině oceánu. Zejména hlubinná ryba má z obličeje vyčnívající dlouhou rybářskou tyč podobnou adaptaci, na jejímž konci je bioluminiscenční kousek kůže, který se kroutí jako červ, aby nalákal svou kořist. Někteří musí konzumovat jiné ryby stejné velikosti nebo větší než oni a potřebují úpravy, které jim pomohou efektivně trávit. Velké ostré zuby, sklopné čelisti, nepřiměřeně velká tlama a roztažitelná těla jsou některé z charakteristik, které pro tento účel mají hlubinné ryby.[11] The gulper úhoř je jedním příkladem organismu, který vykazuje tyto vlastnosti.
Ryby v různých pelagických a hlubinných bentických zónách jsou fyzicky strukturované a chovají se způsoby, které se od sebe výrazně liší. Zdá se, že skupiny koexistujících druhů v každé zóně fungují podobným způsobem, jako je malá mezopelagika vertikálně migrující krmítka planktonem, bathypelagic ďasovití a hlubinná bentika rattaily. "[16]
Ray ploutev druhy s ostnatými ploutvemi jsou mezi hlubinnými rybami vzácné, což naznačuje, že hlubinné ryby jsou staré a tak dobře přizpůsobené svému prostředí, že invaze modernějších ryb byly neúspěšné.[17] Několik paprskových ploutví, které existují, je hlavně v Beryciformes a Lampriformes, což jsou také starodávné formy. Většina hlubinných pelagických ryb patří do jejich vlastních řádů, což naznačuje dlouhý vývoj v hlubinných prostředích. Naproti tomu hlubinné bentické druhy jsou v řádu, který zahrnuje mnoho souvisejících mělkých vodních ryb.[18]
| Druhy podle pelagické zóny | |
|---|---|
| Mnoho druhů se denně pohybuje mezi zónami při vertikálních migracích. V této tabulce jsou uvedeny ve střední nebo hlubší zóně, kde se pravidelně nacházejí. | |
| Zóna | Mezi druhy a skupiny druhů patří ... |
| Epipelagický[19] | |
| Mezopelagický | Lucerna, opah, lancetfish, Barreleye, Ridgehead, sabretooth, semafor loosejaw, mořské líhně[20] |
| Bathypelagic | Hlavně Bristlemouth a ďas mořský. Taky tesák, zmije, černý vlaštovka, dalekohled, kladivo, daggertooth, barracudina, černá pochva, bobtail snipe úhoř, jednorožec hřeben, pelikán úhoř, ochablé velryby. |
| Benthopelagický[19] | Rattail a nevěstinec jsou obzvláště hojné. |
| Benthic | Platýs velký, hagfish, úhoř, Greeneye úhoř, rejnok, hrudkovitý, a batfish[19] |
| Srovnávací struktura pelagických ryb | ||||
|---|---|---|---|---|
| Epipelagický | Mezopelagický | Bathypelagic | hluboké moře bentický | |
| svaly | svalová těla, zkostnatělé kosti, šupiny, dobře vyvinuté žábry a centrální nervový systém a velká srdce a ledviny. | špatně vyvinutý, ochablý | ||
| kostra | silné, zkostnatělé kosti | slabá, minimální osifikace | ||
| váhy | Ano | žádný | ||
| nervové systémy | dobře vyvinuté | pouze boční linie a čichové | ||
| oči | velké a citlivé | malé a nemusí fungovat | proměnná (dobře vyvinutá až chybí) | |
| fotofóry | chybí | běžný | běžný | obvykle chybí |
| žábry | dobře vyvinuté | |||
| ledviny | velký | malý | ||
| srdce | velký | malý | ||
| plynový měchýř | vertikálně stěhovavé ryby mají plavecké měchy | omezený nebo chybí | proměnná (dobře vyvinutá až chybí) | |
| velikost | obvykle pod 25 cm | variabilní, druhy větší než jeden metr nejsou neobvyklé | ||
Mesopelagické ryby





Pod epipelagickou zónou se podmínky rychle mění. Mezi 200 metry a asi 1000 metry světlo stále mizí, dokud není téměř žádné. Teploty klesají a termoklin na teploty mezi 3,9 ° C (39 ° F) a 7,8 ° C (46 ° F). To je soumrak nebo mezopelagický zóna. Tlak stále roste rychlostí jedné atmosféry každých 10 metrů, zatímco koncentrace živin klesají, spolu s rozpuštěným kyslíkem a rychlostí, jakou voda cirkuluje. “[4]
Provozovatelé sonarů využívající nově vyvinutou technologii sonarů během druhé světové války byli zmateni tím, co se zdálo být falešným mořským dnem 300–500 metrů hluboko ve dne a méně hluboko v noci. Ukázalo se, že to bylo způsobeno miliony mořských organismů, zejména malými mezopelagickými rybami, s plaveckými měchýři odrážejícími sonar. Tyto organismy migrují za soumraku do mělčí vody, aby se živily planktonem. Vrstva je hlubší, když je měsíc venku, a může se stát mělčí, když mraky přecházejí přes měsíc. Tento jev se stal známým jako hluboká rozptylová vrstva.[24]
Většina mesopelagických ryb vyrábí každý den vertikální migrace, pohybující se v noci do epipelagické zóny, často sledující podobné migrace zooplanktonu, a během dne se vracet do hlubin kvůli bezpečnosti.[4][25] Tyto vertikální migrace se často vyskytují na velké vertikální vzdálenosti a jsou prováděny za pomoci a plynový měchýř. Plavecký měchýř je nafouknutý, když se chce ryba posunout nahoru, a vzhledem k vysokému tlaku v mesoplegické zóně to vyžaduje značnou energii. Jak ryby stoupají, musí se tlak v plaveckém měchýři upravit, aby se zabránilo prasknutí. Když se chce ryba vrátit do hlubin, je plavecký měchýř vypuštěn.[26] Některé mesopelagické ryby denně migrují přes termoklin, kde se teplota mění mezi 50 ° F (10 ° C) a 69 ° F (20 ° C), čímž se zobrazují značné tolerance pro změnu teploty.[27]
Tyto ryby mají svalnatá těla, zkostnatělé kosti, šupiny, dobře vyvinuté žábry a centrální nervový systém a velké srdce a ledviny. Mezopelagický podavače planktonu mít malá ústa s jemnými žábry, zatímco piscivores mají větší ústa a hrubší žábry.[4] Vertikálně stěhovavé ryby mají plavecké měchy.[17]
Mesopelagické ryby jsou přizpůsobeny pro aktivní život za špatných světelných podmínek. Většina z nich jsou vizuální predátoři s velkýma očima. Některé ryby s hlubší vodou mají tubulární oči s velkými čočkami a pouze tyčové buňky které vypadají vzhůru. Ty poskytují binokulární vidění a velkou citlivost na malé světelné signály.[4] Tato adaptace poskytuje vylepšené terminální vidění na úkor laterálního vidění a umožňuje dravci vyzvednout se oliheň, sépie obecná a menší ryby, které jsou siluety proti temnotě nad nimi.
Mesopelagické ryby obvykle postrádají obranné trny a používají barvu maskovat sami od ostatních ryb. Ambush dravci jsou tmavé, černé nebo červené. Jelikož delší, červené vlnové délky světla nedosahují hlubokého moře, funguje červená efektivně stejně jako černá. Použití migračních formulářů zahloubený stříbřité barvy. Na břiše se často zobrazují fotofóry produkující slabé světlo. Pro dravce zdola, dívajícího se nahoru, tohle bioluminiscence maskuje siluetu ryb. Někteří z těchto predátorů však mají žluté čočky, které filtrují (červené) nedostatek okolního světla, takže bioluminiscence je viditelná.[28]
The brownsnout spookfish, druh Barreleye, je jediným obratlovcem, o kterém je známo, že používá zrcadlo, na rozdíl od čočky, aby zaostřil obraz v očích.[29][30]
Odběr vzorků pomocí hloubky vlečné sítě naznačuje to lucerna tvoří až 65% všech hlubinných ryb biomasa.[31] Ve skutečnosti patří lucerna mezi nejrozšířenější, nejlidnatější a nejrozmanitější ze všech obratlovců hraje důležitou roli ekologický role kořisti pro větší organismy. Odhadovaná globální biomasa lucerny je 550 - 660 milionů metrických tun, několikrát chytil celý svět rybolovu. Lucerna také představuje velkou část biomasy odpovědné za hluboká rozptylová vrstva světových oceánů. Sonar odráží se od milionů luceren plavat měchýře, což vypadá jako falešné dno.[32]
Tuňák velkooký jsou epipelagické / mezopelagické druhy, které jí jiné ryby. Satelitní značení ukázalo, že tuňák velkooký často tráví delší dobu plavbou hluboko pod hladinou během dne, někdy se potápí až 500 metrů. Předpokládá se, že tyto pohyby reagují na vertikální migraci kořistních organismů v hluboká rozptylová vrstva.

The semafor loosejaw má spodní čelist o čtvrtinu delší než jeho tělo. Čelist nemá podlahu a je připevněna pouze závěsem a upravenou kostí jazyka. Velké zuby podobné zubům vpředu jsou následovány mnoha malými ostnatými zuby.[33][34]

The semafor loosejaw je také jednou z mála ryb, které produkují červenou barvu bioluminiscence. Jelikož většina jejich kořisti nedokáže vnímat červené světlo, umožňuje to lovit v podstatě neviditelným paprskem světla.[33]

Lancetfish s dlouhým nosem. Lancetfish jsou přepadení predátoři, kteří tráví veškerý čas v mezopelagické zóně. Patří mezi největší mesopelagické ryby (až 2 metry).[35]

The daggertooth paralyzuje jiné mezopelagické ryby, když je kousne zuby podobnými dýce.[36]



Bathypelagické ryby
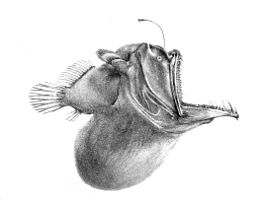



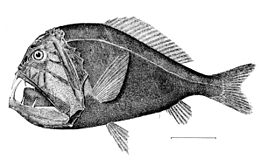
Pod mezopelagickou zónou je temná tma. Toto je půlnoc (nebo bathypelagická zóna ), sahající od 1 000 metrů do spodní hluboké vody bentická zóna. Pokud je voda výjimečně hluboká, pelagické pásmo pod 4000 metrů se někdy nazývá nižší půlnoc (nebo abyssopelagic zóna ). Teploty v této zóně se pohybují od 1 do 4 stupňů Celsia a jsou zcela aphotické.
Podmínky jsou v těchto zónách poněkud jednotné; tma je úplná, tlak je drtivý a teploty, živiny a hladiny rozpuštěného kyslíku jsou nízké.[4]
Bathypelagické ryby mají speciální úpravy vyrovnat se s těmito podmínkami - mají pomalé metabolismus a nespecializovaná strava, ochota jíst všechno, co přijde. Raději sedí a čekají na jídlo, než aby ztráceli energii hledáním. Chování bathypelagických ryb lze srovnávat s chováním mezopelagických ryb. Mesopelagické ryby jsou často vysoce mobilní, zatímco batypelagické ryby jsou téměř všichni draví lži-na-počkání, obvykle vynakládají na pohyb málo energie.[44]
Dominantní bathypelagické ryby jsou malé Bristlemouth a ďas mořský; tesák, zmije, daggertooth a barracudina jsou také běžné. Tyto ryby jsou malé, mnoho asi 10 centimetrů dlouhé a ne mnoho delší než 25 cm. Většinu času tráví trpělivým čekáním ve vodním sloupci, aby se objevila kořist nebo aby ji lákali jejich luminofory. Jak málo energie je k dispozici v bathypelagických zónách filtruje shora v podobě detritu, fekálního materiálu a příležitostných bezobratlých nebo mezopelagických ryb.[44] Asi 20 procent potravin, které mají původ v epipelagické zóně, spadá do mezopelagické zóny,[24] ale jen asi 5 procent filtruje dolů do bathypelagické zóny.[37]
Bathypelagické ryby jsou sedavé, přizpůsobené k produkci minimální energie v prostředí s velmi malým množstvím potravy nebo dostupnou energií, dokonce ani sluneční světlo, pouze bioluminiscence. Jejich těla jsou protáhlý se slabými, vodnatými svaly a kosterní struktur. Jelikož tolik ryb je voda, nejsou v těchto hloubkách stlačovány velkými tlaky. Často jsou výsuvné, otočné čelisti se zakřivenými zuby. Jsou slizké, bez váhy. Centrální nervový systém je omezen na laterální linii a čichové systémy, oči jsou malé a nemusí fungovat, a žábry, ledviny a srdce a plavecké měchy jsou malé nebo chybí.[37][45]
Jedná se o stejné rysy, jaké se vyskytují u ryb larvy, což naznačuje, že bathypelagické ryby během svého vývoje tyto vlastnosti získaly neoteny. Stejně jako u larev tyto vlastnosti umožňují rybám zůstat zavěšeny ve vodě s malým výdajem energie.[46]
Navzdory svému divokému vzhledu jsou tato zvířata hlubin většinou miniaturní ryby se slabým svalstvem a jsou příliš malá na to, aby představovala jakoukoli hrozbu pro člověka.
Plavecké bazény hlubinných ryb buď chybí, nebo jsou sotva funkční a batypelagické ryby obvykle neprovádějí vertikální migrace. Plnění měchýřů při tak velkých tlacích přináší obrovské náklady na energii. Některé hlubinné ryby mají plavecké měchy, které fungují, zatímco jsou mladé, a obývají horní epipelagickou zónu, ale když se ryby přesunou dolů do svého dospělého prostředí, uschnou nebo se naplní tukem.[47]
Nejdůležitějšími smyslovými systémy jsou obvykle vnitřní ucho, který reaguje na zvuk, a boční čára, který reaguje na změny tlaku vody. The čichový Systém může být také důležitý pro muže, kteří najdou ženy podle čichu.[48]Bathypelagické ryby jsou černé nebo někdy červené a málo fotofóry. Když se používají fotofóry, obvykle je to nalákat kořist nebo přilákat partnera. Jelikož je jídlo tak vzácné, batypelagičtí predátoři nejsou ve svých stravovacích návycích selektivní, ale chytají vše, co se k nim dost blíží. Dosahují toho tím, že mají velkou tlamu s ostrými zuby, aby popadli velkou kořist a překrývali se žábry které zabraňují úniku malé spolknuté kořisti.[45]
V této zóně není snadné najít partnera. Některé druhy závisí na bioluminiscence, kde jsou bioluminiscenční vzory jedinečné pro konkrétní druhy. Ostatní jsou hermafroditi, což zdvojnásobuje jejich šance na produkci vajíček i spermií, když dojde k setkání.[37] Samice ďasů se uvolní feromony přilákat drobné muže. Když ji muž najde, kouše se na ni a nikdy ji nepustí. Když muž z druhu ďasovitých Haplophryne mollis kousne do kůže ženy, uvolní enzym který tráví pokožku jeho úst a jejího těla a spojuje pár do bodu, kdy se oba oběhové systémy spojí. Samec pak atrofuje na nic víc než pár pohlavní žlázy. Tento extrém sexuální dimorfismus zajišťuje, že když je žena připravena k rozmnožování, má okamžitě k dispozici partnera.[49]
V batypelagické zóně žije mnoho jiných forem než ryby, například chobotnice, velké velryby, chobotnice, houby, brachiopody, mořské hvězdy a echinoidy, ale v této zóně je pro ryby obtížné žít.

The pelikán úhoř používá ústa jako síť, otevírá velká ústa a plave na svou kořist. Má luminiscenční orgán na špičce ocasu, aby přilákal kořist.

The černý vlaštovka, se svým roztažitelným žaludkem, je pozoruhodný svou schopností polykat, celý, kostnaté ryby desetinásobek jeho hmotnosti.[50][51]

ženský Haplophryne mollis ďasovitý vlečený připojenými muži, kteří atrofovali do páru pohlavní žlázy, pro použití, když je žena připravena se rozmnožit.


Přizpůsobení vysokému tlaku
Jak se ryba pohybuje hlouběji do moře, váha vody nad hlavou se zvyšuje hydrostatický tlak na ryby. Tento zvýšený tlak je asi jeden standardní atmosféra na každých 10 metrů do hloubky (jedna atmosféra je tlak vyvíjený na hladinu moře atmosférou nahoře). Pro rybu na dně bathypelagická zóna, tento tlak činí asi 400 atmosfér (téměř 6000 liber na čtvereční palec).[52]
Hlubinné organismy mají adaptace na buněčné a fyziologické úrovni, které jim umožňují přežít v prostředích s velkým tlakem. Nemít tyto úpravy omezuje hloubky, ve kterých mohou druhy mělkých vod fungovat. Vysoká úroveň vnějšího tlaku ovlivňuje, jak probíhají metabolické procesy a biochemické reakce. Rovnováha mnoha chemických reakcí je narušena tlakem a tlak může inhibovat procesy, které vedou ke zvýšení objemu. Voda, klíčový zastánce mnoha biologických procesů, je velmi náchylná k objemovým změnám, hlavně proto, že složky buněčné tekutiny mají vliv na vodní strukturu. Enzymatické reakce, které vyvolávají změny v organizaci vody, tedy účinně mění objem systému.[53] Proteiny zodpovědné za katalytické reakce jsou obvykle drženy pohromadě slabými vazby a reakce obvykle zahrnují zvýšení objemu.[54] Aby se přizpůsobila této změně, byla upravena proteinová struktura a reakční kritéria hlubinných ryb tak, aby vydržely tlak, aby bylo možné za těchto podmínek provádět reakce. Ve vysokotlakém prostředí dvouvrstvá buněčné membrány ztráta plynulosti. Hlubinné buněčné membrány upřednostňují fosfolipidové dvojvrstvy s vyšším podílem nenasycené mastné kyseliny, které vyvolávají vyšší tekutost než jejich protějšky na hladině moře.

Hlubinné druhy vykazují nižší změny entropie a entalpie ve srovnání s organismy na povrchové úrovni, protože prostředí s vysokým tlakem a nízkou teplotou upřednostňuje negativní změny entalpie a snižuje závislost na reakcích řízených entropií. Ze strukturálního hlediska globulární proteiny hlubinných ryb terciární struktura G-aktin je relativně tuhý ve srovnání s hladinou ryb.[55] Že bílkoviny v hlubinných rybách se strukturálně liší od povrchových ryb, je patrné z pozorování, které aktin z svalová vlákna hlubinných ryb jsou extrémně odolné vůči teplu; podobný tomu, co se nachází v ještěrky. Tyto proteiny jsou strukturálně posíleny modifikací vazeb v terciární struktuře proteinu, což také vyvolává vysokou úroveň tepelné stability.[56] Proteiny jsou strukturně zesíleny, aby odolávaly tlaku modifikací vazeb v terciární struktuře.[57] Proto vysoká hladina hydrostatického tlaku, podobně jako vysoké tělesné teploty termofilních pouštních plazů, upřednostňuje rigidní proteinové struktury.
Na + / K + -ATPáza je lipoprotein enzym, který hraje významnou roli v osmoregulace a je silně ovlivněn hydrostatickým tlakem. Inhibice Na + / K + -ATPázy je způsobena zvýšenou kompresí v důsledku tlaku. Krok omezující rychlost reakce Na + / K + -ATPázy indukuje expanzi ve dvojvrstvě obklopující protein, a proto zvětšení objemu. Zvýšení objemu činí reaktivitu Na + / K + -ATPázy citlivou na vyšší tlaky. Přestože aktivita Na + / K + -ATPázy na gram žaberní tkáň je nižší pro hlubinné ryby, Na + / K + -ATPázy hlubinných ryb vykazují mnohem vyšší toleranci hydrostatického tlaku ve srovnání s jejich protějšky v mělké vodě. Toto je ilustrováno mezi druhy C. acrolepis (asi 2000 metrů hluboko) a jeho hadalpelagický protějšek C. armatus (asi 4000 metrů hluboko), kde jsou Na + / K + -ATPázy z C. armatus jsou mnohem méně citlivé na tlak. Tuto odolnost vůči tlaku lze vysvětlit adaptacemi v proteinu a lipidu skupiny Na + / K + -ATPázy.[58]
Lucerna

Odběr vzorků pomocí hloubky vlečné sítě naznačuje to lucerna tvoří až 65% všech hlubinných ryb biomasa.[31] Ve skutečnosti patří lucerna mezi nejrozšířenější, nejlidnatější a nejrozmanitější ze všech obratlovců hraje důležitou roli ekologický role kořisti pro větší organismy. S odhadovanou globální biomasou 550 - 660 milionů metrické tuny, několikrát za celým světovým rybolovem, tvoří lucerny také velkou část biomasy odpovědné za hluboká rozptylová vrstva světových oceánů. V Jižní oceán, Myctophids poskytují alternativní zdroj potravy krill pro predátory, jako je oliheň a královský tučňák. I když jsou tyto ryby hojné a plodné, v současné době jen několik komerčních luceren rybolov existují: Patří sem omezené vypnuté operace Jižní Afrika, v sub-Antarktida a v Ománský záliv.
Ohrožené druhy
2006 studie podle kanadský vědci našel pět druhů hlubinných ryb - modrý štikozubec, ostnatý úhoř - být na pokraji vyhynutí v důsledku posunu o komerční rybolov z kontinentální police na svahy řeky kontinentální police, až do hloubky 1600 metrů. Pomalý reprodukce z nich Ryba - dosáhnou sexuální dospělost zhruba ve stejném věku jako lidské bytosti - je jedním z hlavních důvodů, proč se nemohou zotavit z nadměrný rybolov.[59]
Viz také
- Sčítání mořského života
- Hluboká oceánská voda
- Hluboké moře
- Hlubinná společenství
- Hlubinné ryby
- Ryby žijící při dně
- Pelagické ryby
Citace
- ^ Trujillo, Alan P .; Harold V. Thurman (2011). Essentials of Oceanography 10. vydání. Boston: Prentice Hall. p. 354. ISBN 978-0321668127.
- ^ Trujillo, Alan P .; Harold V. Thurman (2011). Essentials of Oceanography 10. vydání. Boston: Prentice Hall. p. 365. ISBN 978-0321668127.
- ^ Trujillo, Alan P .; Harold V. Thurman (2011). Essentials of Oceanography 10. vydání. Boston: Prentice Hall. 457, 460. ISBN 978-0321668127.
- ^ A b C d E F Moyle a Cech, 2004, strana 585
- ^ Whartone, Davide. (2002). Život na hranici možností: Organismy v extrémním prostředí. Cambridge, Velká Británie: Cambridge UP. str.198. ISBN 978-0521782128.
- ^ Wharton, David (2002). Život na hranici možností: Organismy v extrémním prostředí. Cambridge, Velká Británie: Cambridge UP. 199, 201–202. ISBN 978-0521782128.
- ^ A b Whartone, Davide. (2002). Život na hranici možností: Organismy v extrémním prostředí. Cambridge, Velká Británie: Cambridge UP. str.199. ISBN 978-0521782128.
- ^ Compagno, L.J.V. (1984). Sharks of the World: Anotovaný a ilustrovaný katalog druhů žraloků známých dodnes. Organizace pro výživu a zemědělství OSN. str. 14–15. ISBN 92-5-101384-5.
- ^ Wharton, David A. (2007-07-23). Život na hranici možností: Organismy v extrémním prostředí. Cambridge University Press. ISBN 9781139431941.
- ^ Musilova, Zuzana; Cortesi, Fabio; Matschiner, Michael; Davies, Wayne; Patel, Jagdish; Stieb, Sara; de Busserolles, Fanny; Malmstrøm, Martin; Tørresen, Ole; Brown, Celeste; Mountford, Jessica; Hanel, Reinhold; Stenkamp, Deborah; Jakobsen, Kjetill; Carleton, Karen; Jentoft, Sissel; Marshall, Justin; Salzburger, Walter. „Vize využívající několik odlišných opsinů prutů v hlubinných rybách“. Sciencemag. Americká asociace pro rozvoj vědy. Citováno 10. května 2019.
- ^ A b Trujillo, Alan P .; Harold V. Thurman (2011). Essentials of Oceanography 10. vydání. Boston: Prentice Hall. p. 415. ISBN 978-0321668127.
- ^ Trujillo, Alan P .; Harold V. Thurman (2011). Essentials of Oceanography 10. vydání. Boston: Prentice Hall. 414–415. ISBN 978-0321668127.
- ^ Randall, David J .; Anthony Peter Farrell (1997). Hlubinné ryby. San Diego: Academic. p. 217. ISBN 978-0123504401.
- ^ Randall, David J .; Anthony Peter Farrell (1997). Hlubinné ryby. San Diego: Academic. p. 195. ISBN 978-0123504401.
- ^ Randall, David J .; Anthony Peter Farrell (1997). Hlubinné ryby. San Diego: Academic. 196, 225. ISBN 978-0123504401.
- ^ Moyle a Čech, 2004, s. 591
- ^ A b Haedrich RL (1996) „Hlubinné ryby: vývoj a adaptace v největších životních prostorech Země“ Journal of Fish Biology 49(sA): 40-53.
- ^ Moyle a Cech, 2004, strana 586
- ^ A b C Moyle a Cech, 2004, strana 571
- ^ Froese, Rainer a Pauly, Daniel, eds. (2009). "Argyropelecus aculeatus" v FishBase. Verze ze srpna 2009.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2009). "Dissostichus mawsoni" v FishBase. Verze ze srpna 2009.
- ^ Tajemství hlubinných ryb s tubulárníma očima a průhlednou hlavou vyřešeno ScienceDaily, 24. února 2009.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2010). "Gigantura chuni" v FishBase. Verze z října 2010.
- ^ A b Ryan P „Hlubinná stvoření: mezopelagická zóna“ Te Ara - encyklopedie Nového Zélandu. Aktualizováno 21. září 2007.
- ^ Bone & Moore 2008, s. 38.
- ^ Douglas EL, Friedl WA, Pickwell GV (1976). „Ryby v minimálních zónách kyslíku: charakteristiky okysličení krve“. Věda. 191 (4230): 957–959. doi:10.1126 / science.1251208. PMID 1251208.
- ^ Moyle a Čech, 2004, s. 590
- ^ Munz WRA (1976). "Na žlutých čočkách u mesopelagických zvířat". Marine Biological Association of the UK. 56 (4): 963–976. doi:10.1017 / S0025315400021019.
- ^ Wagner, H.J .; Douglas, R.H .; Frank, T. M.; Roberts, N.W. & Partridge, J.C. (27. ledna 2009). „Nové oko obratlovců využívající refrakční i reflexní optiku“. Aktuální biologie. 19 (2): 108–114. doi:10.1016 / j.cub.2008.11.061. PMID 19110427. S2CID 18680315.
- ^ Smith, L. (8. ledna 2009). „Ryby se čtyřmi očima vidí skrz hlubinný šero“. Times Online. Times Newspapers Ltd. Citováno dne 14. března 2009.
- ^ A b Hulley, P. Alexander (1998). Paxton, J.R .; Eschmeyer, W.N. (eds.). Encyklopedie ryb. San Diego: Academic Press. str. 127–128. ISBN 0-12-547665-5.
- ^ R. Cornejo; R. Koppelmann; T. Sutton. „Diverzita hlubinných ryb a ekologie v bentické mezní vrstvě“.
- ^ A b Kenaley, CP (2007). "Revize semaforu rodu Loosejaw." Malacosteus (Teleostei: Stomiidae: Malacosteinae), s popisem nového druhu z mírné jižní polokoule a Indického oceánu “. Copeia. 2007 (4): 886–900. doi:10.1643 / 0045-8511 (2007) 7 [886: ROTSLG] 2.0.CO; 2.
- ^ Sutton, T. T. (listopad 2005). „Trofická ekologie hlubinných ryb Malacosteus niger (Pisces: Stomiidae): Tajemná ekologie krmení pro usnadnění jedinečného vizuálního systému? “. Deep-Sea Research Part I: Oceanographic Research Papers. 52 (11): 2065–2076. doi:10.1016 / j.dsr.2005.06.011.
- ^ Moyle a Čech, 2004, s. 336
- ^ Froese, Rainer a Pauly, Daniel, eds. (2010). "Anotopterus pharao" v FishBase. Verze z dubna 2010.
- ^ A b C d Ryan P „Hlubinná stvoření: batypelagická zóna“ Te Ara - encyklopedie Nového Zélandu. Aktualizováno 21. září 2007.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2006). "Gonostoma bathyphilum" v FishBase. Verze z ledna 2006.
- ^ Froese, Rainer a Daniel Pauly, eds. (2009). "Gonostoma" v FishBase. Verze ze srpna 2009.
- ^ „Spojování znalostí a lidí více než 10 let“. Archivovány od originál dne 09.07.2012.
- ^ „Vědci řeší záhadu: 3 ryby jsou stejné“. 22. ledna 2009. Citováno 2009-01-22.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2010). "Chauliodus sloani" v FishBase. Verze z dubna 2010.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2009). "Anoplogaster cornuta" v FishBase. Verze ze srpna 2009.
- ^ A b Moyle a Čech, 2004, s. 594
- ^ A b Moyle a Čech, 2004, s. 587
- ^ Marshall (1984) „Progenetic tendences in deep-sea fishes“, str. 91-101, Potts GW a Wootton RJ (eds.) (1984) Reprodukce ryb: strategie a taktika Rybářská společnost Britských ostrovů.
- ^ Horn MH (1970). „Plavecký měchýř jako mladistvý orgán u stromateoidních ryb“. Breviora. 359: 1–9.
- ^ Jumper GY, Bair RC (1991). „Lokalizace podle čichu: model a aplikace na problém páření v hlubinných Hatchetfish Argyropelecus hemigymnus". Americký přírodovědec. 138 (6): 1431–1458. doi:10.1086/285295. JSTOR 2462555. S2CID 84386858.
- ^ Theodore W. Pietsch (1975). „Precocious sexual parasitism in the deep sea ceratioid dialerfish, Cryptopsaras couesi Gill“. Příroda. 256 (5512): 38–40. doi:10.1038 / 256038a0. S2CID 4226567. Citováno 31. července 2008.
- ^ Jordan, D.S. (1905). Průvodce studiem ryb. H. Holt and Company.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2009). "Chiasmodon niger" v FishBase. Verze ze srpna 2009.
- ^ Scott, Thomas R .; Powell, James (2018-09-24). Vesmír takový, jaký ve skutečnosti je: Země, vesmír, hmota a čas. Columbia University Press. doi:10,7312 / skot18494. ISBN 978-0-231-54576-1.
- ^ HOCHACHKA, PETER W .; SOMERO, GEORGE N. (1984). Biochemická adaptace. Princeton University Press. JSTOR j.ctt7zv9d4.
- ^ Priede, Imants G., ed. (2017), „Přizpůsobení hlubinnému moři“, Hlubinné ryby: biologie, rozmanitost, ekologie a rybolov, Cambridge: Cambridge University Press, s. 87–138, ISBN 978-1-316-01833-0, vyvoláno 2020-10-31
- ^ HOCHACHKA, PETER W .; SOMERO, GEORGE N. (1984). Biochemická adaptace. Princeton University Press. JSTOR j.ctt7zv9d4.
- ^ Priede, Imants G., ed. (2017), „Přizpůsobení hlubinnému moři“, Hlubinné ryby: biologie, rozmanitost, ekologie a rybolov, Cambridge: Cambridge University Press, str. 87–138, ISBN 978-1-316-01833-0, vyvoláno 2020-10-31
- ^ Priede, Imants G., ed. (2017), „Přizpůsobení hlubinnému moři“, Hlubinné ryby: biologie, rozmanitost, ekologie a rybolov, Cambridge: Cambridge University Press, s. 87–138, ISBN 978-1-316-01833-0, vyvoláno 2020-10-31
- ^ Somero, G N (říjen 1992). „Přizpůsobení vysokému hydrostatickému tlaku“. Roční přehled fyziologie. 54 (1): 557–577. doi:10.1146 / annurev.ph.54.030192.003013. ISSN 0066-4278. PMID 1314046.
- ^ Devine Jennifer A., Baker Krista D., Haedrich Richard L. (2006). „Rybolov: Hlubinné ryby se kvalifikují jako ohrožené“. Příroda. 439 (7072): 29. doi:10.1038 / 439029a. PMID 16397489. S2CID 4428618.CS1 maint: více jmen: seznam autorů (odkaz)
Reference
- Moyle, PB a Cech, JJ (2004) Ryby, Úvod do ichtyologie. 5. vydání, Benjamin Cummings. ISBN 978-0-13-100847-2
Další čtení
- Gordon J. D. M. (2001) „Hlubinné ryby“ In: John H. Steele, Steve A. Thorpe, Karl K. Turekian (Eds) Prvky fyzické oceánografie, strany 227–233, Academic Press. ISBN 9780123757241.
- Hoar WS, Randall DJ a Farrell AP (Eds) (1997) Hlubinné ryby, Academic Press. ISBN 9780080585406.
- Shotton, Ross (1995) „Hlubinný rybolov“ V: Přezkum stavu světových mořských rybolovných zdrojů, Technický papír FAO pro rybolov 457, FAO, Řím. ISBN 92-5-105267-0.
- Tandstad M, Shotton R, Sanders J a Carocci F (2011) „Hlubinný rybolov“ V: Přezkum stavu světových mořských rybolovných zdrojů, strany 265–278, technický papír FAO Fisheries 569, FAO, Řím. ISBN 978-92-5-107023-9.
externí odkazy
| Externí video | |
|---|---|
- https://www.pbs.org/wgbh/nova/abyss/life/bestiary.html
- http://ocean.nationalgeographic.com/ocean/photos/deep-sea-creatures/
- Hlubinná stvoření - Články, fakta a obrázky hlubinných zvířat


| O Ryba |  | ||||
|---|---|---|---|---|---|
| Anatomie a fyziologie | |||||
| Smyslové systémy | |||||
| Reprodukce | |||||
| Pohyb | |||||
| jiný chování | |||||
| Podle místo výskytu | |||||
| Jiné typy | |||||
| Komerční |
| ||||
| Hlavní skupiny | |||||
| Seznamy |
| ||||
| |||||
| Kontrolní úřad |
|---|