Mořská tráva - Seagrass

Mořské trávy jsou (pouze) kvetoucí rostliny které rostou v námořní prostředí. Existuje asi 60 druhů plně mořských mořských řas, které patří ke čtyřem rodiny (Posidoniaceae, Zosteraceae, Hydrocharitaceae a Cymodoceaceae ), vše v pořadí Alismatales (ve třídě jednoděložné rostliny ).[1] Mořské trávy se vyvinuly z suchozemské rostliny která před 70 až 100 miliony let přebarvila oceán.
Název mořská tráva pochází z mnoha druhů s dlouhými a úzkými listy, které rostou oddenkovým rozšířením a často se šíří napříč velkými "louky „připomínající travní porosty; mnoho druhů povrchně připomíná suchozemské trávy z rodiny Poaceae.
Jako všichni autotrofní rostliny, mořské trávy fotosyntetizovat, v ponoření fotonická zóna, a většina se vyskytuje v mělkých a chráněných pobřežních vodách zakotvených v písku nebo bahnitém dně. Většina druhů podstoupí ponorku opylení a dokončit svůj životní cyklus pod vodou.
Mořské trávy tvoří pod vodou husté mořské louky které patří mezi nejproduktivnější ekosystémy na světě. Fungují jako důležité uhlíkové výlevky a poskytovat stanoviště a potravu pro rozmanitost mořský život srovnatelné s korálové útesy.
Vývoj

Suchozemské rostliny se vyvinul snad již před 450 miliony let ze skupiny zelené řasy.[2] Mořské trávy se poté vyvinuly ze suchozemských rostlin, které migrovaly zpět do oceánu.[3][4] Před asi 70 až 100 miliony let se tři nezávislé linie mořských řas (Hydrocharitaceae, Cymodoceaceae komplex a Zosteraceae) vyvinuly z jedné linie jednoděložný kvetoucí rostliny.[5]
Jiné rostliny, které kolonizovaly moře, jako např slaný močál rostliny, mangrovy, a mořské řasy, mají rozmanitější evoluční linie. I přes svou nízkou druhovou rozmanitost se mořským trávám podařilo kolonizovat kontinentální šelfy všech kontinentů kromě Antarktidy.[6]
Taxonomie
| Rodina | obraz | Rody | Popis |
|---|---|---|---|
| Zosteraceae | Rodina Zosteraceae, také známý jako mořská tráva rodina, zahrnuje dva rody obsahující 14 mořských druhů. Nachází se v mírný a subtropický pobřežní vody s nejvyšší rozmanitostí v Koreji a Japonsku. Mezisoučet druhů: | ||
 | Phyllospadix | 6 druhů | |
 | Zostera | 16 druhů | |
| Hydrocharitaceae | Rodina Hydrocharitaceae, také známý jako páskové trávy, zahrnout Kanadská vodní řasa a frogbit. Rodina zahrnuje jak čerstvé, tak mořské vodní sporty, ačkoli ze šestnácti rodů, které jsou v současnosti uznávány, jsou pouze tři mořské. [7] Vyskytují se po celém světě v nejrůznějších stanovištích, jsou však primárně tropická. Mezisoučet druhů: | ||
 | Enhalus | 1 druh | |
 | Halophila | 19 druhů | |
 | Thalassia | 2 druhy | |
| Posidoniaceae | Rodina Posidoniaceae obsahuje jeden rod se dvěma až devíti mořskými druhy nalezenými v mořích Středomoří a kolem jižního pobřeží Austrálie. Mezisoučet druhů: 2 až 9 | ||
 | Posidonia | 2 až 9 druhů | |
| Cymodoceaceae | Rodina Cymodoceaceae, také známý jako kapustňák, zahrnuje pouze mořské druhy.[8] Někteří taxonomové tuto rodinu nepoznali. Mezisoučet druhů: | ||
 | Amphibolis | 2 druhy | |
 | Cymodocea | 4 druhy | |
 | Halodule | 6 druhů | |
 | Stříkačka | 2 druhy | |
 | Thalassodendron | 3 druhy | |
| Celkový počet druhů: | |||
Přílivové a subtidální mořské trávy
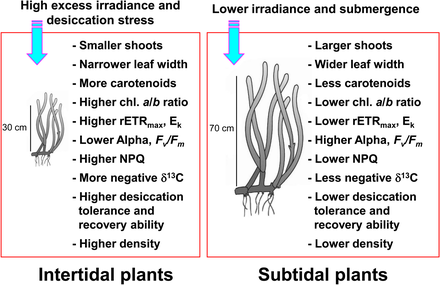
Mořské trávy vyskytující se v přílivových a přílivových pásmech jsou vystaveny velmi proměnlivým podmínkám prostředí v důsledku změn přílivu a odlivu.[10][11] Mořské trávy v přílivová zóna jsou pravidelně vystaveni vzduchu a následně mají extrémně vysoké a nízké teploty, vysokou fotoinhibici ozáření, a vysušení napětí ve vztahu k subtidální mořské trávě.[11][12][13] Takové extrémní teploty mohou vést k výraznému odumírání mořských trav, když jsou mořské trávy vystaveny vzduchu během odlivu.[14][15][16] Vysušovací napětí během odlivu bylo považováno za primární faktor omezující distribuci mořských řas v horní přílivové zóně.[17] Mořské trávy žijící v přílivové zóně jsou obvykle menší než ty v subtidální zóně, aby se minimalizovaly účinky stresového stavu.[18][19] Přílivové mořské trávy také vykazují reakce závislé na světle, jako je snížená fotosyntetická účinnost a zvýšená fotoochrana během období vysokého ozáření a expozice vzduchu.[20][21]

Naproti tomu mořské trávy v subtidální zóna přizpůsobit se sníženým světelným podmínkám způsobeným útlumem světla a rozptylem v důsledku překrývajícího se vodního sloupce a suspendovaných částic.[23][24] Mořské trávy v hluboké subtidální zóně mají obecně delší listy a širší listové čepele než ty v mělké subtidální nebo intertidální zóně, což umožňuje více fotosyntézy, což vede k většímu růstu.[13] Mořské trávy také reagují na snížené světelné podmínky zvýšením chlorofyl obsah a snižování poměr chlorofylu a / b pro zvýšení absorpce světla účinnost pomocí efektivního využití vlnových délek.[25][26][27] Vzhledem k tomu, že mořské trávy v přílivových a přílivových pásmech jsou za velmi odlišných světelných podmínek, vykazují výrazně odlišné fotoaklimatické odezvy, aby maximalizovaly fotosyntetickou aktivitu a ochranu před nadměrným ozářením.
Mořské trávy asimilují velké množství anorganický uhlík k dosažení vysoké úrovně produkce.[28][29] Mořské makrofyty, včetně mořské trávy, použijte oba CO2 a HCO−
3 (hydrogenuhličitan ) pro redukci fotosyntetického uhlíku.[30][31][32] Navzdory expozici vzduchu během odlivu mohou mořské trávy v přílivové zóně pokračovat ve fotosyntéze s využitím CO2 ve vzduchu.[33] Složení anorganických zdrojů uhlíku pro fotosyntézu mořských řas se tedy mezi intertidálními a subtidálními rostlinami pravděpodobně liší. Protože stabilní poměry izotopů uhlíku změn rostlinných tkání na základě anorganických zdrojů uhlíku pro fotosyntézu,[34][35] mořské trávy v přílivových a přílivových pásmech mohou mít různá rozmezí stabilních poměrů izotopů uhlíku.
Mikrobiom z mořské trávy
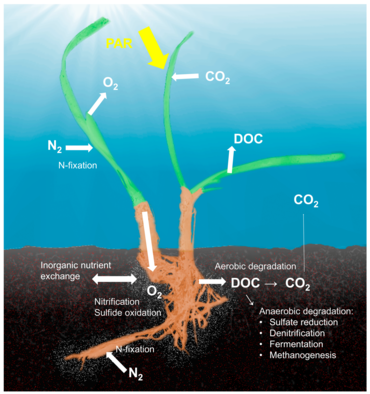
Seagrass holobiont
Koncept holobiont, který zdůrazňuje význam a interakce mikrobiálního hostitele s přidruženými mikroorganismy a viry a popisuje jejich fungování jako jediné biologické jednotky,[38] byla zkoumána a diskutována pro mnoho modelových systémů, i když existuje zásadní kritika konceptu, který definuje různé symbiózy hostitele a mikrobů jako jednu biologickou jednotku.[39] Pojmy holobiont a hologenome se vyvinuly od původní definice,[40] a není pochyb o tom, že symbiotické mikroorganismy jsou klíčové pro biologii a ekologii hostitele tím, že poskytují vitamíny, energii a anorganické nebo organické živiny, podílejí se na obranných mechanismech nebo řídí vývoj hostitele.[41] Ačkoli většina prací o interakcích hostitel-mikrob byla zaměřena na živočišné systémy, jako jsou korály, houby nebo lidé, existuje značná část literatury o rostlinných holobiontech.[42] Mikrobiální společenství spojená s rostlinami ovlivňují obě klíčové složky kondice rostlin, růstu a přežití,[43] a jsou utvářeny dostupností živin a obrannými mechanismy rostlin.[44] Bylo popsáno několik stanovišť, která ukrývají mikroby spojené s rostlinami, včetně rhizoplane (povrch kořenové tkáně), rhizosféra (okraj kořenů), endosféra (uvnitř rostlinné tkáně) a fylosphere (celková nadzemní plocha).[36]
Mořské louky
Postele z mořské trávy / louky mohou být buď monospecifické (tvořené jediným druhem), nebo na smíšených postelích. v mírný oblasti, obvykle dominuje jeden nebo několik druhů (jako úhoř Zostera marina v severním Atlantiku), zatímco tropický postele jsou obvykle rozmanitější, až třináct druh zaznamenané v Filipíny.
Postele z mořské trávy jsou rozmanité a produktivní ekosystémy a může chovat stovky přidružených druhů ze všech phyla, například pro mladistvé a dospělé Ryba, epifytický a volný život makrořasy a mikrořasy, měkkýši, štětinatí červi, a hlístice. Několik druhů bylo původně považováno za krmené přímo mořskou trávou listy (částečně kvůli jejich nízkému obsahu výživy), ale vědecké recenze a vylepšené pracovní metody ukázaly, že mořská tráva bylinožravý je důležitým článkem v potravinovém řetězci, který živí stovky druhů, včetně zelené želvy, dugongové, kapustňáci, Ryba, husy, labutě, mořští ježci a kraby. Některé druhy ryb, které navštěvují / se živí mořskými trávami, vychovávají mláďata v sousedních oblastech mangrovy nebo korálové útesy.
Mořské trávy zachycují sediment a zpomalují pohyb vody, což vede k usazování suspendovaného sedimentu. Výhody zachycování sedimentu korál snížením zatížení sedimentů a zlepšením fotosyntézy pro korály i mořské trávy.[45]

Bílé tečky, často se vyskytující v oblastech mořských řas
Podvodní záběry z mořské louky, býčí hus a mořský úhoř





Ačkoli jsou mořské trávy často přehlíženy, poskytují řadu ekosystémové služby[46][47]. Mořské trávy se berou v úvahu ekosystémoví inženýři.[48][4][3] To znamená, že rostliny mění ekosystém kolem sebe. K této úpravě dochází ve fyzikální i chemické formě. Mnoho druhů mořských řas produkuje rozsáhlou podzemní síť kořenů a oddenek který stabilizuje sediment a snižuje erozi pobřeží.[49] Tento systém také pomáhá při okysličování sedimentu a poskytuje pohostinné prostředí organismy žijící v sedimentech.[48] Mořské trávy také vylepšují Kvalita vody stabilizací těžkých kovů, znečišťujících látek a přebytečných živin.[50][4][3] Dlouhé stébla mořských trav zpomalují pohyb vody, což snižuje energii vln a poskytuje další ochranu proti pobřeží eroze a nárůst bouře. Navíc, protože mořské trávy jsou podvodní rostliny, produkují značné množství kyslíku, který okysličuje vodní sloupec. Tyto louky tvoří více než 10% celkového úložiště uhlíku v oceánu. Na hektar obsahuje dvakrát tolik oxidu uhličitého než deštné pralesy a dokáže zachytit asi 27,4 milionu tun CO2 každoročně.[51] Skladování uhlíku je zásadní ekosystémová služba jak přecházíme do období zvýšené atmosférické hladiny uhlíku. Některé modely změny klimatu však naznačují, že některé mořské trávy vyhynou - Posidonia oceanica Očekává se, že vyhyne, nebo téměř, do roku 2050.
Mořské louky poskytují potravu mnoha mořským býložravcům. Mořské želvy, kapustňáci, papoušci, bodláci, mořští ježci a pinfish se živí mořskými řasami. Mnoho dalších menších zvířat se živí epifity a bezobratlými, kteří žijí na a mezi lopatkami mořských řas.[52] Louky z mořských řas také poskytují fyzické stanoviště v oblastech, které by jinak byly bez jakékoli vegetace. Kvůli této trojrozměrné struktuře ve vodním sloupci mnoho druhů zabírá stanoviště mořských řas jako úkryt a hledání potravy. Odhaduje se, že 17 druhů korálových útesových ryb tráví celou svoji mladistvou životní fázi výhradně na plochách mořských řas.[53] Tato stanoviště také fungují jako školka pro komerčně a rekreačně ceněné druhy rybolovu, včetně roubíka (Mycteroperca microlepis ), červený buben, obyčejný snook, a mnoho dalších.[54][55] Některé druhy ryb využívají mořské louky a různé etapy životního cyklu. V nedávné publikaci Dr. Ross Boucek a jeho kolegové zjistili, že dvě velmi vyhledávané ploché ryby, snook a skvrnitý mořský pstruh poskytují základní potravní stanoviště během reprodukce.[56] Sexuální reprodukce je extrémně energeticky nákladná, aby ji bylo možné dokončit akumulovanou energií; pro úplnou reprodukci proto vyžadují louky z mořských řas v těsné blízkosti.[56] Kromě toho mnoho komerčně důležitých bezobratlých také pobývají na stanovištích mořských řas, včetně lastur (Argopecten irradians ), krabi podkovy, a krevety. Charismatická fauna může být také viděna při návštěvě stanovišť mořských řas. Mezi tyto druhy patří Západoindický kapustňák, zelené mořské želvy a různé druhy žraloků. Vysoká rozmanitost mořských organismů, které lze nalézt na stanovištích mořských řas, je prosazuje jako turistickou atrakci a významný zdroj příjmů pro mnoho pobřežních ekonomik podél Mexického zálivu a v Karibiku.
Vztah k lidem
Historicky byly mořské trávy sbírány jako hnojivo pro písčitou půdu. Toto bylo důležité použití v Laguna Aveiro, Portugalsko, kde byly shromážděné rostliny známé jako Moliço.
Na počátku 20. století ve Francii a v menší míře v Normanské ostrovy, sušené mořské trávy byly použity jako matrace (paillasse) výplň - po těchto matracích byly francouzské síly během roku velmi žádané první světová válka. To bylo také používáno pro obvazy a jiné účely.
V únoru 2017 vědci zjistili, že mořské louky mohou být schopné odstranit různé patogeny z mořské vody. Na malých ostrovech bez zařízení na čištění odpadních vod ve střední Indonésii jsou patogenní mořské bakterie - jako Enterococcus - které ovlivňují lidi, ryby a bezobratlých byly sníženy o 50 procent, když byly přítomny louky z mořských řas, ve srovnání s párovými místy bez mořských řas[57] i když by to mohlo být na újmu jejich přežití.[58]
Poruchy a hrozby
Přírodní poruchy, jako např pastva, bouře, drhnutí ledu a vysušení, jsou nedílnou součástí mořské trávy ekosystém dynamika. Mořské trávy vykazují vysoký stupeň fenotypová plasticita, rychle se přizpůsobující měnícím se podmínkám prostředí.
Mořské trávy jsou na globálním úbytku s přibližně 30 000 km2 (12 000 čtverečních mil) ztraceno během posledních desetiletí. Hlavní příčinou je zejména lidské narušení eutrofizace, mechanické zničení stanoviště a nadměrný rybolov. Nadměrný přísun živin (dusík, fosfor ) je přímo toxický pro mořské trávy, ale co je nejdůležitější, stimuluje růst epifytických a volně plovoucích makro - a mikro -řasy. To oslabuje sluneční světlo, snížení fotosyntéza který vyživuje mořské trávy a prvovýroba Výsledek.
Rostoucí rozpadající se listy mořské trávy a řasy květy řas, což má za následek pozitivní zpětná vazba. To může způsobit dokončení posun režimu od mořské trávy po dominanci řas. Hromadící se důkazy také naznačují, že nadměrný rybolov predátoři (velké dravé ryby) by mohly nepřímo zvýšit růst řas snížením kontroly pastvy prováděné mesograzery, jako korýši a plži, prostřednictvím a trofická kaskáda.
Květy makroskopických řas způsobit úbytek a vymýcení mořských řas. Makrořasy, známé jako obtěžující druhy, rostou ve vláknitých a listových formách a vytvářejí nad mořskou trávou husté nepřipojené rohože, které se na listech mořských řas vyskytují jako epifity. Eutrofizace vede k tvorbě květu, což způsobuje útlum světla ve vodním sloupci, což nakonec vede k anoxickým podmínkám pro mořské trávy a organismy žijící v rostlinách / v jejich okolí. Kromě přímého blokování světla rostlin mají bentické řasy nízký obsah uhlíku / dusíku, což způsobuje, že jejich rozklad stimuluje bakteriální aktivitu, což vede k resuspenzi sedimentů, zvýšení zákalu vody a dalšímu zeslabení světla.[59][60]
Když lidé projíždějí motorovými čluny po mělkých oblastech mořských řas, někdy může vrtulový list mořskou trávu poškodit.
Mezi nejpoužívanější metody ochrany a obnovy louk z mořských řas patří živiny a znečištění snížení, chráněné mořské oblasti a restaurování pomocí mořské trávy přesazování. Mořská tráva není považována za odolnou vůči dopadům budoucích změn prostředí.[61]
Obnovení
Na různých místech se komunity pokoušejí obnovit záhony mořských řas, které byly ztraceny lidskou činností, včetně amerických států Virginie,[62] Florida[63] a Havaj,[64] stejně jako Spojené království.[65] Ukázalo se, že taková znovuzavedení zlepšují ekosystémové služby.[66]
Od roku 2019[Aktualizace] Výzkumné středisko pro pobřežní mořské ekosystémy v Central Queensland University pěstuje mořské trávy již šest let a produkuje semena mořské trávy. Provádějí pokusy o klíčení a setí.[67]
Viz také
- Alismatales
- Modrý uhlík
- Slaniska
- Mangovník
- Prohlížeč dat Ocean: obsahuje globální distribuci datového souboru mořských trav
Reference
- ^ Tomlinson a Vargo (1966). „Na morfologii a anatomii želví trávy, Thalassia testudinum (Hydrocharitaceae). I. Vegetativní morfologie“. Bulletin of Marine Science. 16: 748–761.
- ^ Knauth, L. Paul; Kennedy, Martin J. (2009). „Pozdní prekambrijská ekologizace Země“. Příroda. 460 (7256): 728–732. Bibcode:2009Natur.460..728K. doi:10.1038 / nature08213. PMID 19587681. S2CID 4398942.
- ^ A b C Orth; et al. (2006). „Globální krize pro ekosystémy mořských řas“. BioScience. 56 (12): 987–996. doi:10.1641 / 0006-3568 (2006) 56 [987: AGCFSE] 2.0.CO; 2. hdl:10261/88476.
- ^ A b C Papenbrock, J (2012). "Hlavní body ve fylogenezi, fyziologii a metabolismu mořských řas: co je činí takovými druhy?". International Scholarly Research Network: 1–15.
- ^ Les, D.H., Cleland, M.A. a Waycott, M. (1997) „Fylogenetické studie u Alismatidae, II: vývoj mořských krytosemenných rostlin (mořské trávy) a hydrofilie“. Systematická botanika 22(3): 443–463.
- ^ Orth, Robert J .; Carruthers, TIM J. B .; Dennison, William C .; Duarte, Carlos M .; Fourqurean, James W .; Heck, Kenneth L .; Hughes, A. Randall; Kendrick, Gary A .; Kenworthy, W. Judson; Olyarnik, Suzanne; Krátký, Frederick T .; Waycott, Michelle; Williams, Susan L. (2006). „Globální krize pro mořské trávy ekosystémů“. Bioscience. 56 (12): 987. doi:10.1641 / 0006-3568 (2006) 56 [987: AGCFSE] 2.0.CO; 2. ISSN 0006-3568.
- ^ Christenhusz, Maarten J.M .; Byng, James W. (2016-05-20). „Počet známých druhů rostlin na světě a jeho roční přírůstek“. Phytotaxa. 261 (3): 201. doi:10.11646 / fytotaxa.261.3.1. ISSN 1179-3163.
- ^ Waycott, Michelle; McMahon, Kathryn; Lavery, Paul (2014). Průvodce jižními mírnými mořskými trávami. Publikování CSIRO. ISBN 9781486300150.
- ^ Park, Sang Rul; Kim, Sangil; Kim, Young Kyun; Kang, Chang-Keun; Lee, Kun-Seop (2016). „Fotoklimatické odpovědi Zostera marina v přílivových a subtidálních zónách“. PLOS ONE. 11 (5): e0156214. Bibcode:2016PLoSO..1156214P. doi:10.1371 / journal.pone.0156214. PMC 4881947. PMID 27227327.
 Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0. - ^ Silva, J .; Santos, R. (2003). "Denní variační vzory fotosyntézy mořské trávy podél vertikálního přechodu". Série pokroku v ekologii moří. 257: 37–44. Bibcode:2003MEPS..257 ... 37S. doi:10 3354 / meps257037.
- ^ A b Boese, Bruce L .; Robbins, Bradley D .; Thrby, Glen (2005). „Vysušení je limitujícím faktorem pro distribuci eelgrassu (Zostera marina L.) v přílivové zóně ústí severovýchodního Pacifiku (USA).“ Botanica Marina. 48 (4). doi:10.1515 / BOT.2005.037. S2CID 85105171.
- ^ Durako, M. J .; Kunzelman, J. I .; Kenworthy, W. J .; Hammerstrom, K. K. (2003). „Hloubkově závislá variabilita ve fotobiologii dvou populací Halophila johnsonii a Halophila decipiens“. Mořská biologie. 142 (6): 1219–1228. doi:10.1007 / s00227-003-1038-3. S2CID 85627116.
- ^ A b Olivé, I .; Vergara, J. J .; Pérez-Lloréns, J. L. (2013). "Fotosyntetická a morfologická fotoklimatizace mořské trávy Cymodocea nodosa na sezónu, hloubku a polohu listu". Mořská biologie. 160 (2): 285–297. doi:10.1007 / s00227-012-2087-2. S2CID 86386210.
- ^ Hemminga M. A. a Durate C. M. (2000) Ekologie mořské trávy. Cambridge University Press.
- ^ Seddon, S .; Cheshire, AC (2001). "Fotosyntetická reakce Amphibolis antarctica a Posidonia australis na teplotu a vysychání pomocí chlorofylové fluorescence". Série pokroku v ekologii moří. 220: 119–130. Bibcode:2001MEPS..220..119S. doi:10 3354 / meps220119.
- ^ Hirst A, Ball D, Heislers S, Young P, Blake S, Coots A. Baywide Seagrass Monitoring Program, Milestone Report No. 2 (2008). Fisheries Victoria Technical Report No. 29, January 2009.
- ^ Koch, Evamaria W. (2001). „Beyond Light: fyzikální, geologické a geochemické parametry jako možné požadavky na stanoviště ponořené vodní vegetace“. Ústí řek. 24 (1): 1–17. doi:10.2307/1352808. JSTOR 1352808. S2CID 85287808.
- ^ Tanaka, Y .; Nakaoka, M. (2004). „Nouzový stres a morfologická omezení ovlivňují druhové rozšíření a růst subtropických přílivových mořských řas“. Série pokroku v ekologii moří. 284: 117–131. Bibcode:2004MEPS..284..117T. doi:10 3354 / meps284117.
- ^ Seddon, S .; Cheshire, AC (2001). "Fotosyntetická reakce Amphibolis antarctica a Posidonia australis na teplotu a vysoušení pomocí chlorofylové fluorescence". Série pokroku v ekologii moří. 220: 119–130. Bibcode:2001MEPS..220..119S. doi:10,3 354 / meps220119.
- ^ Björk, M .; Uku, J .; Weil, A .; Beer, S. (1999). "Fotosyntetické tolerance k vysychání tropických přílivových mořských řas". Série pokroku v ekologii moří. 191: 121–126. Bibcode:1999MEPS..191..121B. doi:10,3 354 / meps191121.
- ^ Petrou, K .; Jimenez-Denness, I .; Chartrand, K .; McCormack, C .; Rasheed, M .; Ralph, PJ (2013). „Sezónní heterogenita ve fotofyziologické reakci na expozici vzduchu u dvou tropických přílivových druhů mořských řas“ (PDF). Série pokroku v ekologii moří. 482: 93–106. Bibcode:2013MEPS..482 ... 93P. doi:10 3354 / meps10229.
- ^ Xu, Shaochun; Zhou, Yi; Wang, Pengmei; Wang, Feng; Zhang, Xiaomei; Gu, Ruiting (2016). „Slanost a teplota významně ovlivňují klíčení semen, usazování sazenic a růst sazenice Zostera přístav. PeerJ. 4: e2697. doi:10,7717 / peerj.2697. PMC 5119234. PMID 27896031. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Schwarz, A.-M .; Björk, M .; Buluda, T .; Mtolera, M .; Beer, S. (2000). "Fotosyntetické využití uhlíku a světla dvěma tropickými druhy mořských řas, měřeno in situ". Mořská biologie. 137 (5–6): 755–761. doi:10,1007 / s002270000433. S2CID 86384408.
- ^ Campbell, Stuart J .; McKenzie, Len J .; Kerville, Simon P .; Bité, Juanita S. (2007). "Vzory fotosyntézy tropické mořské trávy ve vztahu ke světlu, hloubce a stanovišti". Estuarine, Coastal and Shelf Science. 73 (3–4): 551–562. Bibcode:2007ECSS ... 73..551C. doi:10.1016 / j.ecss.2007.02.014.
- ^ Lee, Kun-Seop; Dunton, Kenneth H. (1997). „Vliv redukce světla in situ na údržbu, růst a rozdělení zdrojů uhlíku v bankách Thalassia testudinum ex König“. Journal of Experimental Marine Biology and Ecology. 210: 53–73. doi:10.1016 / S0022-0981 (96) 02720-7.
- ^ Longstaff, B.J .; Dennison, W. C. (1999). „Přežití mořské trávy během pulzních zákalů: Účinky deprivace světla na mořské trávy Halodule pinifolia a Halophila ovalis.“ Vodní botanika. 65 (1–4): 105–121. doi:10.1016 / S0304-3770 (99) 00035-2.
- ^ Collier, CJ; Lavery, PS; Ralph, PJ; Masini, RJ (2008). „Fyziologické vlastnosti mořské trávy Posidonia sinuosa podél gradientu dostupnosti světla souvisejícího s hloubkou“. Série pokroku v ekologii moří. 353: 65–79. Bibcode:2008MEPS..353 ... 65C. doi:10 3354 / meps07171.
- ^ Lee, Kun-Seop; Park, Sang Rul; Kim, Young Kyun (2007). "Účinky ozáření, teploty a živin na dynamiku růstu mořských řas: přehled". Journal of Experimental Marine Biology and Ecology. 350 (1–2): 144–175. doi:10.1016 / j.jembe.2007.06.016.
- ^ Nayar, S .; Collings, G.J .; Miller, D.J .; Bryars, S .; Cheshire, A.C. (2009). „Příjem a přidělování zdrojů anorganického uhlíku mírnými mořskými řasami Posidonia a Amphibolis“. Journal of Experimental Marine Biology and Ecology. 373 (2): 87–95. doi:10.1016 / j.jembe.2009.03.010.
- ^ Beer, Sven (1989). "Fotosyntéza a fotorespirace mořských krytosemenných rostlin". Vodní botanika. 34 (1–3): 153–166. doi:10.1016/0304-3770(89)90054-5.
- ^ Larkum AWD, James PL. Směrem k modelu absorpce anorganického uhlíku v mořských řasách zahrnujících karboanhydrázu. V editorech Kuo J, Phillips RC, Walker DI, Kirkman H. Biologie mořské trávy: sborník z mezinárodního semináře. Nedlands: The University of Western Australia; 1996. s. 191–196.
- ^ Pivo, Sven; Rehnberg, Jon (1997). "Získání anorganického uhlíku mořskou trávou Zostera marina". Vodní botanika. 56 (3–4): 277–283. doi:10.1016 / S0304-3770 (96) 01109-6.
- ^ Silva, João; Santos, Rui; Calleja, Maria Ll .; Duarte, Carlos M. (2005). „Ponorená versus vzduchem vystavená intertidální makrofytová produktivita: od fyziologických po hodnocení na úrovni komunity“. Journal of Experimental Marine Biology and Ecology. 317: 87–95. doi:10.1016 / j.jembe.2004.11.010.
- ^ O'Leary, Marion H. (1988). "Uhlíkové izotopy ve fotosyntéze". BioScience. 38 (5): 328–336. doi:10.2307/1310735. JSTOR 1310735.
- ^ Raven, John A .; Johnston, Andrew M .; Kübler, Janet E .; Korb, Rebecca; McInroy, Shona G .; Handley, Linda L .; Scrimgeour, Charlie M .; Walker, Diana I .; Beardall, John; Vanderklift, Mathew; Fredriksen, Stein; Dunton, Kenneth H. (2002). „Mechanická interpretace diskriminace izotopů uhlíku mořskými řasami a mořskými řasami“. Funkční biologie rostlin. 29 (3): 355–378. doi:10.1071 / PP01201. PMID 32689482.
- ^ A b Ugarelli, K., Chakrabarti, S., Laas, P. a Stingl, U. (2017) „Seagrass holobiont and its microbiome“. Mikroorganismy, 5(4): 81. doi:10,3390 / mikroorganismy5040081. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Tarquinio, F., Hyndes, G.A., Laverock, B., Koenders, A. a Säwström, C. (2019) „The seagrass holobiont: pochopení interakcí mořských řas a bakterií a jejich role ve fungování ekosystému mořských řas“. MKS mikrobiologické dopisy, 366(6): fnz057. doi:10.1093 / femsle / fnz057.
- ^ Margulis, Lynn (1991) „Symbiogeneze a symbionticismus“. V: Symbióza jako zdroj evolučních inovací; Margulis, L., Fester, R. (Eds.), Cambridge MIT Press. ISBN 9780262132695.
- ^ Douglas, A.E .; Werren, J.H. (2016) „Holes in the Hologenome: Why Host-Microbe Symbioses are not Holobionts“. mBio, 7: e02099-15. doi:10,1 128 / mBio.02099-15.
- ^ Theis, K.R .; Dheilly, N.M .; Klassen, J.L .; Brucker, R.M .; Baines, J.F .; Bosch, T.C.G .; Cryan, J.F .; Gilbert, S.F .; Dobrou noc, C.J .; Lloyd, E.A.; et al. Správný koncept Hologenome: Ekoevoluční rámec pro hostitele a jejich mikrobiomy. mSystems 2016, 1, e00028-16. doi:10,1 128 / m Systémy 00028-16.
- ^ Rosenberg, E. a Zilber-Rosenberg, I. (2016) „Mikroby řídí vývoj zvířat a rostlin: koncept hologenomu“. MBio, 7(2). doi:10,1 128 / mBio.01395-15.
- ^ Zilber-Rosenberg, I. a Rosenberg, E. (2008) „Role mikroorganismů ve vývoji zvířat a rostlin: teorie evoluce hologenomu“. Recenze mikrobiologie FEMS, 32(5): 723–735. doi:10.1111 / j.1574-6976.2008.00123.x.
- ^ Vandenkoornhuyse, P., Quaiser, A., Duhamel, M., Le Van, A. a Dufresne, A. (2015) „Význam mikrobiomu rostliny holobiont“. Nový fytolog, 206(4): 1196-1206. doi:10,1111 / nph.13312.
- ^ Sánchez-Cañizares, C., Jorrín, B., Poole, P.S. a Tkacz, A. (2017) „Porozumění holobiontu: vzájemná závislost rostlin a jejich mikrobiomu“. Současný názor v mikrobiologii, 38: 188–196. doi:10.1016 / j.mib.2017.07.001.
- ^ Seagrass-Watch: Co je to mořská tráva? Citováno 2012-11-16.
- ^ Nordlund, Lina; Koch, Evamaria W .; Barbier, Edward B .; Creed, Joel C. (2016-10-12). Reinhart, Kurt O. (ed.). „Ekosystémové služby mořských řas a jejich variabilita napříč rody a geografickými oblastmi“. PLOS ONE. 11 (10): e0163091. Bibcode:2016PLoSO..1163091M. doi:10.1371 / journal.pone.0163091. ISSN 1932-6203. PMC 5061329. PMID 27732600.
- ^ Program OSN pro životní prostředí (2020). Z čista jasna: Hodnota mořských trav pro životní prostředí a pro lidi. UNEP, Nairobi. https://www.unenvironment.org/resources/report/out-blue-value-seagrasses-environment-and-people
- ^ A b Jones, Clive G .; Lawton, John H .; Shachak, Moshe (1994). "Organismy jako ekosystémoví inženýři". Oikosi. 69 (3): 373–386. doi:10.2307/3545850. JSTOR 3545850.
- ^ Gray, William; Moffler, Mark (1987). „Kvetení mořské trávy Thalassia testudinum (Hydrocharitacea) v oblasti Tampa Bay na Floridě“. Vodní botanika. 5: 251–259. doi:10.1016/0304-3770(78)90068-2.
- ^ Darnell, Kelly; Dunton, Kenneth (2016). „Reprodukční fenologie subtropických mořských tráv Thalassia testudinum (želví tráva) a Halodule wrightii (hejnová tráva) v severozápadním Mexickém zálivu“. Botanica Marina. 59 (6): 473–483. doi:10.1515 / bot-2016-0080. S2CID 88685282.
- ^ Macreadie, P. I .; Baird, M. E.; Trevathan-Tackett, S. M .; Larkum, A. W. D .; Ralph, P. J. (2013). "Kvantifikace a modelování kapacity sekvestrace uhlíku na loukách z mořských řas". Bulletin o znečištění moří. 83 (2): 430–439. doi:10.1016 / j.marpolbul.2013.07.038. PMID 23948090.
- ^ https://myfwc.com/research/habitat/seagrasses/information/faq/#:~:text=Expand%2FCollapse%20What%20animals%20eat,on%20and%20among%20seagrass%20blades.
- ^ Nagelkerken, I .; Roberts, C. M .; van der Velde, G .; Dorenbosch, M .; van Riel, M. C .; Cocheret de la Morinière, E .; Nienhuis, P. H. (2002). „Jak důležité jsou mangrovy a mořské řasy pro ryby z korálových útesů? Hypotéza školky byla testována na ostrovním měřítku.“ Série pokroku v ekologii moří. 244: 299–305. Bibcode:2002MEPS..244..299N. doi:10 3354 / meps244299.
- ^ Nordlund, L. M .; Unsworth, R. K. F .; Gullstrom, M .; Cullen-Unsworth, L. C. (2018). „Celosvětový význam rybolovné činnosti z mořských řas“. Ryby a rybolov. 19 (3): 399–412. doi:10.1111 / faf.12259.
- ^ Unsworth, R. K. F .; Nordlund, L. M .; Cullen-Unsworth, L. C. (2019). „Mořské louky podporují celosvětovou produkci rybolovu“. Conserv Lett. e12566: e12566. doi:10.1111 / conl.12566.
- ^ A b Boucek, R.E .; Leone, E .; Bickford, J .; Walters-Burnsed, S .; Lowerre-Barbieri, S. (2017). „Více než jen místo tření: Zkoumání jemného měřítka [použití esa dvou druhů ryb v ústí řek v místě tření“. Frontiers in Marine Science (4): 1–9.
- ^ Byington, Cara (2017-02-17). „Nová věda ukazuje, že mořské louky potlačují patogeny“. Nature.org. Členové skupiny NatureNet pro studenou zelenou vědu. Citováno 17. února 2017.
- ^ Jones, BJ; Cullen-Unsworth, L. C .; Unsworth, R. K. F. (2018). „Sledování zdroje dusíku pomocí δ15N odhaluje lidské a zemědělské faktory degradace mořské trávy na Britských ostrovech“. Hranice ve vědě o rostlinách. 9: 133. doi:10.3389 / fpls.2018.00133. PMC 5808166. PMID 29467789.
- ^ McGlathery, KJ (2001). „Květy makroskopických řas přispívají k úbytku mořské trávy v pobřežních vodách obohacených o živiny“ (PDF). Journal of Phycology. 37 (4): 453–456. doi:10.1046 / j.1529-8817.2001.037004453.x. S2CID 38983997.
- ^ Fox SE, YS Olsen a AC Spivak (2010) „Dopady kontrol zdola nahoru a shora dolů a změna klimatu na komunity makrofytů v ústí řek a ekosystémové služby, které poskytují“ In: PF Kemp (Ed) Sborník Eco-DAS Symposium„TAKÉ, kapitola 8: 129–145.
- ^ Unsworth, Richard K.F .; Collier, Catherine J .; Waycott, Michelle; McKenzie, Len J .; Cullen-Unsworth, Leanne C. (2015). „Rámec pro odolnost ekosystémů mořských řas“. Bulletin o znečištění moří. 100 (1): 34–46. doi:10.1016 / j.marpolbul.2015.08.016. PMID 26342389.
- ^ "Obnova eelgrassu | Ochrana přírody ve Virginii". www.nature.org. Citováno 2018-08-06.
- ^ „Obnova mořské trávy“. myfwc.com. Citováno 2018-08-06.
- ^ „Iniciativa pro obnovení mořské trávy - Malama Maunalua“. www.malamamaunalua.org. Citováno 2018-08-06.
- ^ Unsworth, Richard K. F .; McKenzie, Len J .; Collier, Catherine J .; Cullen-Unsworth, Leanne C .; Duarte, Carlos M .; Eklöf, Johan S .; Jarvis, Jessie C .; Jones, Benjamin L .; Nordlund, Lina M. (01.08.2019). „Globální výzvy pro zachování mořských řas“. Ambio. 48 (8): 801–815. doi:10.1007 / s13280-018-1115-r. ISSN 1654-7209. PMC 6541581. PMID 30456457.
- ^ van Katwijk, Marieke M .; Thorhaug, Anitra; Marba, Núria; Orth, Robert J .; Duarte, Carlos M .; Kendrick, Gary A .; Althuizen, Inge H. J .; Balestri, Elena; Bernard, Guillaume (2015-11-25). „Globální analýza obnovy mořských řas: význam rozsáhlé výsadby“. Journal of Applied Ecology. 53 (2): 567–578. doi:10.1111/1365-2664.12562. ISSN 0021-8901.
- ^ Mackay, Jacquie; Stünzner, Inga (24. října 2019). „Mateřská škola pro mořské trávy v centrální Queenslandu by mohla kompenzovat emise uhlíku“. ABC News. Australian Broadcasting Corporation. Citováno 24. října 2019.
Jedna květina může vyprodukovat 15 semen a jedno semeno zasazené ve správných podmínkách může vytvořit hektar mořské trávy.
Další reference
- den Hartog, C. 1970. Mořské trávy světa. Verhandl. der Koninklijke Nederlandse Akademie van Wetenschappen, Afd. Natuurkunde, Č. 59 (1).
- Duarte, Carlos M. a Carina L. Chiscano „Biomasa a produkce mořské trávy: přehodnocení“ Aquatic Botany, svazek 65, čísla 1–4, listopad 1999, strany 159–174.
- Zelená, E.P. & Short, F.T. (eds). 2003. Atlas světa mořských řas. University of California Press, Berkeley, CA. 298 stran
- Hemminga, M.A. a Duarte, C. 2000. Ekologie mořské trávy. Cambridge University Press, Cambridge. 298 stran
- Hogarth, Peter Biologie mangrovů a mořských řas (Oxford University Press, 2007)
- Larkum, Anthony W. D., Robert J. Orth a Carlos M. Duarte (redaktoři) Mořské trávy: biologie, ekologie a ochrana přírody (Springer, 2006)
- Orth, Robert J. a kol. „Globální krize pro ekosystémy mořských řas“ BioScience Prosinec 2006 / sv. 56 č. 12, strany 987–996.
- Krátký, F.T. & Coles, R.G. (eds). 2001. Globální metody výzkumu mořské trávy. Elsevier Science, Amsterdam. 473 stran
- A.W.D. Larkum, R.J. Orth a C.M. Duarte (eds). Biologie mořské trávy: Pojednání. CRC Press, Boca Raton, FL, v tisku.
- A. Schwartz; M. Morrison; I. Hawes; J. Halliday. 2006. Fyzikální a biologické vlastnosti vzácného mořského stanoviště: přílivová mořská mořská dna pobřežních ostrovů. Věda na ochranu přírody 269. 39 stran [1]
- Waycott, M, McMahon, K, & Lavery, P 2014, Průvodce jižními mořskými řasami mírného podnebí, CSIRO Publishing, Melbourne
externí odkazy
- Cullen-Unsworth, Leanne C .; Unsworth, Richard (03.08.2018). „Výzva k ochraně mořských řas“. Věda. 361 (6401): 446–448. Bibcode:2018Sci ... 361..446C. doi:10.1126 / science.aat7318 (neaktivní 2020-11-30). ISSN 0036-8075. PMID 30072524.CS1 maint: DOI neaktivní od listopadu 2020 (odkaz)
- Projekt Seagrass - Charita prosazující ochranu mořských řas prostřednictvím vzdělávání, vlivu, výzkumu a akcí
- SeagrassSpotter - Občanský vědecký projekt zvyšující povědomí o loukách z mořských řas a mapujících jejich umístění
- Seagrass a Seagrass Beds přehled z portálu Smithsonian Ocean
- Článek Nature Geoscience popisující umístění louk z mořských řas po celém světě
- Seagrass-Watch - největší vědecký, nedestruktivní program pro hodnocení a monitorování mořských řas na světě
- Seagrass Ecosystem Research Group na Swansea University - Inter-disciplinární mořský výzkum pro ochranu
- Restore-A-Scar - nezisková kampaň na obnovu louk z mořských řas poškozených rekvizitami lodí
- SeagrassNet - globální monitorovací program pro mořské trávy
- Seagrass Fund v The Ocean Foundation
- Taxonomie mořských řas
- World Seagrass Association
- SeagrassLI
- Věda a management z mořské trávy v Jihočínském moři a Thajském zálivu
- Mořská ekologie (Prosinec 2006) - speciální vydání o mořských řasách
- Kambodžské mořské trávy
- Produktivita na mořské trávě - COST Action ES0906
- Rybolov v západní Austrálii - Informační list o mořské trávě

