Mořské mikroorganismy - Marine microorganisms
Mořské mikroorganismy jsou definovány svým stanovištěm jako mikroorganismy žijící v mořské prostředí, tj. v slaná voda moře nebo oceánu nebo brakický voda pobřežní ústí. Mikroorganismus (nebo mikrob) je libovolný mikroskopický živobytí organismus, tj. jakákoli forma života příliš malá na to, aby ji pouhé lidské oko skutečně vidělo, potřebující mikroskop. Mikroorganismy jsou velmi rozmanité. Oni mohou být jednobuněčný[1] nebo mnohobuňečný a zahrnout všechny bakterie a archaea a většina prvoky, stejně jako některé druhy houby, řasy a některá mikroskopická zvířata, jako např vířníky a copepods. Mnoho makroskopické zvířata a rostliny mít mikroskopické juvenilní fáze. Někteří mikrobiologové také klasifikují biologicky aktivní entity, jako jsou viry a viroidy jako mikroorganismy, ale ostatní je považují za neživé.[2][3]
Odhaduje se, že mořské mikroorganismy tvoří asi 70%,[4] nebo asi 90%,[5][6] biomasy v oceánu. Společně tvoří mořské mikrobiom. V průběhu miliard let se z tohoto mikrobiomu vyvinulo mnoho životních stylů a adaptací a účastní se ho globální cyklistika téměř všech chemických prvků.[7] Mikroorganismy mají zásadní význam pro recyklaci živin ekosystémy jak se chovají rozkladače. Jsou také zodpovědní za téměř všechny fotosyntéza která se vyskytuje v oceánu, stejně jako jízda na kole uhlík, dusík, fosfor a další živiny a stopové prvky.[8] Mořské mikroorganismy oddělují velké množství uhlíku a produkují většinu světového kyslíku.
Malý podíl mořských mikroorganismů je patogenní, způsobující nemoci a dokonce smrt mořských rostlin a živočichů.[9] Mořské mikroorganismy recyklovat hlavní chemické prvky, a to jak produkcí, tak spotřebováním přibližně poloviny veškeré organické hmoty generované na planetě každý rok. Jako obyvatelé největšího prostředí na Zemi řídí mikrobiální námořní systémy změny v každém globálním systému.
V červenci 2016 vědci uvedli, že identifikovali sadu 355 geny z poslední univerzální společný předek (LUCA) ze všech život na planetě, včetně mořských mikroorganismů.[10] Navzdory své rozmanitosti je mikroskopický život v oceánech stále špatně pochopen. Například role viry v mořských ekosystémech nebyl sotva prozkoumán ani na počátku 21. století.[11]
| Část série přehledů o |
| mořský život |
|---|
| Část série na |
| Plankton |
|---|
 |
Přehled


1. Co je tam venku? Které mikroorganismy jsou přítomny v našich mořích a oceánech a v jakém počtu se vyskytují?
2. Co dělají? Jaké funkce plní každý z těchto mikroorganismů v mořském prostředí a jak přispívají ke globálním cyklům energie a hmoty?
– Evropská vědecká nadace, 2012[12]:14
Mikroorganismy tvoří asi 70% mořská biomasa.[4] A mikroorganismus, nebo mikrob, je mikroskopický organismus příliš malý na to, aby byl dostatečně rozeznatelný pouhým okem. V praxi to zahrnuje organismy menší než asi 0,1 mm.[12]:13
Takové organismy mohou být jednobuněčný[1] nebo mnohobuňečný. Mikroorganismy jsou rozmanité a zahrnují všechny bakterie a archaea, většina protistů počítaje v to řasy, prvoky a plísňové protists, stejně jako určitá mikroskopická zvířata, jako jsou vířníky. Mnoho makroskopické zvířata a rostliny mít mikroskopické juvenilní fáze. Někteří mikrobiologové také klasifikují viry (a viroidy ) jako mikroorganismy, ale ostatní je považují za neživé.[2][3]
Mikroorganismy mají zásadní význam pro recyklaci živin ekosystémy jak se chovají rozkladače. Některé mikroorganismy jsou patogenní, způsobující nemoci a dokonce smrt u rostlin a zvířat.[9] Jako obyvatelé největšího prostředí na Zemi řídí mikrobiální námořní systémy změny v každém globálním systému. Mikroby jsou zodpovědné za prakticky všechno fotosyntéza která se vyskytuje v oceánu, stejně jako jízda na kole uhlík, dusík, fosfor a další živiny a stopové prvky.[13]
| Mořské mikroorganismy | |||||||||||||||||||||||||




Mikroskopický podmořský život je různorodý a stále špatně pochopený, například pokud jde o roli viry v mořských ekosystémech.[14] Většina mořských virů je bakteriofágy, které jsou neškodné pro rostliny a živočichy, ale jsou nezbytné pro regulaci slaných a sladkovodních ekosystémů.[15] Infikují a ničí bakterie ve vodních mikrobiálních komunitách a jsou nejdůležitějším mechanismem recyklace uhlíku v mořském prostředí. Organické molekuly uvolněné z mrtvých bakteriálních buněk stimulují růst čerstvých bakterií a řas.[16] Virová aktivita může také přispět k biologické čerpadlo proces, kterým uhlík je oddělený v hlubokém oceánu.[17]
Proud vzdušných mikroorganismů obíhá planetu nad meteorologickými systémy, ale pod komerčními vzdušnými pruhy.[18] Některé peripatetické mikroorganismy jsou smeteny ze suchozemských prachových bouří, ale většina pochází z mořských mikroorganismů v mořský sprej. V roce 2018 vědci uvedli, že na každý metr čtvereční kolem planety se denně ukládají stovky milionů virů a desítky milionů bakterií.[19][20]
Mikroskopické organismy žijí po celém světě biosféra. Hmotnost prokaryot mikroorganismy - což zahrnuje bakterie a archea, ale nikoliv nukleované eukaryotické mikroorganismy - může to být až 0,8 bilionu tun uhlíku (z celkové biosféry Hmotnost, odhaduje se na 1 až 4 biliony tun).[21] Jednobuněčný barofilní mořské mikroby byly nalezeny v hloubce 10 900 m (35 800 ft) v Mariana příkop, nejhlubší místo v oceánech Země.[22][23] Mikroorganismy žijí uvnitř hornin 580 m (1900 ft) pod mořským dnem pod 2590 m (8,500 ft) oceánu u pobřeží severozápadní Spojené státy,[22][24] stejně jako 2400 m (7 900 ft; 1,5 mi) pod mořským dnem mimo Japonsko.[25] Největší známá teplota, při které může existovat mikrobiální život, je 122 ° C (252 ° F) (Methanopyrus kandleri ).[26] V roce 2014 vědci potvrdili existenci mikroorganismů žijících 800 m (2 600 ft) pod ledem Antarktida.[27][28] Podle jednoho vědce „Mikroby najdete všude - jsou extrémně přizpůsobivé podmínkám a přežijí, ať jsou kdekoli.“[22] Mořské mikroorganismy slouží jako „základ všech mořských potravinářských sítí, recyklace hlavních prvků a produkce a spotřeba asi poloviny organické hmoty generované na Zemi každý rok“.[29][30]
Mořské viry
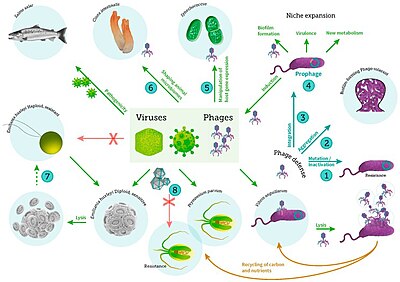
včetně virové infekce bakterií, fytoplanktonu a ryb[31]
A virus je malý infekční agens že replikuje jen uvnitř živých buňky dalších organismy. Viry mohou infikovat všechny typy formy života, z zvířata a rostliny na mikroorganismy, počítaje v to bakterie a archaea.[32]
Pokud nejsou uvnitř infikované buňky nebo v procesu infikování buňky, existují viry ve formě nezávislých částic. Tyto virové částice, také známé jako viriony, se skládá ze dvou nebo tří částí: (i) genetický materiál (genom) vyrobený z obou DNA nebo RNA, dlouho molekuly které nesou genetickou informaci; ii) a protein kabát zvaný kapsid, který obklopuje a chrání genetický materiál; a v některých případech (iii) an obálka z lipidy který obklopuje proteinový obal, když jsou mimo buňku. Tvary těchto virových částic se pohybují od jednoduchých spirálovitý a icosahedral formy pro některé druhy virů na složitější struktury pro jiné. Většina druhů virů má viriony, které jsou příliš malé na to, aby je bylo možné vidět pomocí optický mikroskop. Průměrný virion je asi jedna setina velikosti průměru bakterie.
Počátky virů v EU evoluční historie života jsou nejasné: některé mohou mít vyvinul z plazmidy - kousky DNA, které se mohou pohybovat mezi buňkami - zatímco jiné se mohly vyvinout z bakterií. V evoluci jsou viry důležitým prostředkem horizontální přenos genů, což se zvyšuje genetická rozmanitost.[33] Viry jsou některými považovány za životní formu, protože přenášejí genetický materiál, množí se a vyvíjejí se přírodní výběr. Chybí jim však klíčové vlastnosti (jako je buněčná struktura), které jsou obecně považovány za nezbytné k tomu, aby se považovaly za život. Protože mají některé, ale ne všechny takové vlastnosti, byly viry popsány jako „organismy na pokraji života“[34] a jako replikátoři.[35]

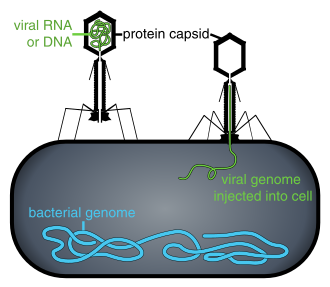

Viry se vyskytují všude tam, kde je život, a pravděpodobně existovaly od doby, kdy se nejprve vyvinuly živé buňky.[36] Původ virů je nejasný, protože netvoří fosilie, takže molekulární techniky byly použity k porovnání DNA nebo RNA virů a jsou užitečným prostředkem ke zkoumání toho, jak vznikly.[37]
Viry jsou nyní považovány za starodávné a mají původ, který předcházel divergenci života v tři domény.[38]
Názory na to, zda jsou viry formou, se liší život nebo organické struktury, které interagují s živými organismy.[39] Někteří je považují za formu života, protože nesou genetický materiál, množí se vytvářením více kopií sebe sama prostřednictvím sebezhromáždění a vyvíjejí se skrz přírodní výběr. Chybí jim však klíčové vlastnosti, jako je buněčná struktura, která se obecně považuje za nezbytnou k tomu, aby se mohla považovat za život. Protože mají některé, ale ne všechny takové vlastnosti, byly viry popsány jako replikátory[40] a jako „organismy na pokraji života“.[34]
Fágy


Bakteriofágy, často právě volal fágy, jsou viry, které parazit bakterie a archaea. Mořské fágy parazity mořských bakterií a archea, jako např sinice.[41] Jsou běžnou a různorodou skupinou virů a jsou nejhojnější biologickou entitou v mořském prostředí, protože jejich hostitelé, bakterie, jsou obvykle numericky dominantním buněčným životem v moři. Obecně je v každém ml mořské vody asi 1 až 10 milionů virů, nebo asi desetkrát více dvouvláknových DNA virů, než jsou buněčné organismy,[42][43] ačkoli odhady množství virů v mořské vodě se mohou v širokém rozmezí lišit.[44][45] Na dlouhou dobu, sledoval fágy objednávky Caudovirales Zdálo se, že dominuje mořským ekosystémům v počtu a rozmanitosti organismů.[41]V důsledku více nespokojeného výzkumu se však viry bez ocasu zdají být dominantní ve více hloubkách a oceánských oblastech, následované Caudovirales rodiny myovirů, podovirů a siphovirů.[46] Fágy patřící do rodin Corticoviridae,[47] Inoviridae,[48] Microviridae,[49] a Autolykiviridae[50][51][52][53]je také známo, že infikují různé mořské bakterie.
Existují také archaeanské viry, které se replikují uvnitř archaea: jedná se o dvouvláknové DNA viry neobvyklých a někdy jedinečných tvarů.[54][55] Tyto viry byly podrobně studovány v dokumentu termofilní archaea, zejména objednávky Sulfolobales a Termoprotealy.[56]
Úloha virů
Mikroorganismy tvoří přibližně 70% mořské biomasy.[4] Odhaduje se, že viry každý den zabijí 20% této biomasy a že v oceánech je 15krát více virů než bakterií a archea. Viry jsou hlavními činiteli odpovědnými za rychlé ničení škodlivých látek květy řas,[57] které často zabíjejí další mořský život.[58]Počet virů v oceánech klesá dále na moři a hlouběji do vody, kde je méně hostitelských organismů.[17]
Viry jsou důležitým přírodním prostředkem přenos genů mezi různými druhy, což se zvyšuje genetická rozmanitost a řídí evoluci.[33] Předpokládá se, že viry hrály ústřední roli v časném vývoji, před diverzifikací bakterií, archea a eukaryot, v době poslední univerzální společný předek života na Zemi.[59] Viry jsou stále jedním z největších rezervoárů neprozkoumané genetické rozmanitosti na Zemi.[17]
Obří viry


Viry mají obvykle délku od 20 do 300 nanometrů. To může být v kontrastu s délkou bakterií, která začíná na přibližně 400 nanometrech. Jsou tu také obří viry, často volané girusy, obvykle asi 1000 nanometrů (jeden mikron) na délku. Všechny obří viry patří kmen Nucleocytoviricota (NCLDV), společně s neštovice Největší známý z nich je Tupanvirus. Tento rod obřího viru byl objeven v roce 2018 v hlubokém oceánu i v sodném jezeře a může dosáhnout celkové délky až 2,3 mikronu.[60]
Objev a následná charakterizace obřích virů vyvolala určitou debatu o jejich evolučním původu. Dvě hlavní hypotézy o jejich původu spočívají v tom, že se buď vyvinuly z malých virů, sbíraly DNA z hostitelských organismů, nebo že se vyvinuly z velmi komplikovaných organismů do současné podoby, která není pro reprodukci soběstačná.[61] Od jakého druhu komplikovaných organismů se mohou obrovské viry lišit, je také předmětem debaty. Jeden návrh je, že počáteční bod ve skutečnosti představuje čtvrtý doména života,[62][63] ale to bylo do značné míry zlevněno.[64][65]
Prokaryotes
Mořské bakterie



Bakterie tvoří velkou doména z prokaryotický mikroorganismy. Typicky několik mikrometry na délku mají bakterie řadu tvarů, od koulí po tyče a spirály. Bakterie byly mezi prvními formami života, které se objevily Země, a jsou přítomny ve většině z nich stanoviště. Bakterie obývají půdu, vodu, kyselé horké prameny, radioaktivní odpad,[66] a hluboké části zemská kůra. Bakterie také žijí symbiotický a parazitický vztahy s rostlinami a zvířaty.
Kdysi považováno za rostliny tvořící třídu Schizomycetes, bakterie jsou nyní klasifikovány jako prokaryoty. Na rozdíl od buněk zvířat a dalších eukaryoty, bakteriální buňky neobsahují a jádro a zřídka přístav vázané na membránu organely. Ačkoli termín bakterie tradičně zahrnoval všechny prokaryoty, vědecká klasifikace se změnilo po objevu v 90. letech, že prokaryoty sestávají ze dvou velmi odlišných skupin organismů vyvinul od starodávného společného předka. Tyto evoluční domény se nazývají Bakterie a Archaea.[67]
Předchůdci moderních bakterií byli jednobuněčné mikroorganismy, které byly první formy života se objeví na Zemi asi před 4 miliardami let. Po dobu asi 3 miliard let byla většina organismů mikroskopická a dominantní formou života byly bakterie a archea.[68][69] Ačkoli bakteriální fosilie existují, jako např stromatolity, jejich nedostatek rozlišovací způsobilosti morfologie brání tomu, aby byly použity k prozkoumání historie bakteriálního vývoje nebo k datu času vzniku konkrétního bakteriálního druhu. Genové sekvence však mohou být použity k rekonstrukci bakterie fylogeneze a tyto studie naznačují, že bakterie se nejprve lišily od linie archaeal / eukaryotic.[70] Bakterie se také účastnily druhé velké evoluční divergence, archaea a eukaryot. Zde eukaryota vyústila do vstupu starodávných bakterií do endosymbiotikum sdružení s předky eukaryotických buněk, které samy o sobě pravděpodobně souvisely s Archaea.[71][72] To zahrnovalo pohlcení proto-eukaryotickými buňkami alfaproteobakteriální symbionty mitochondrie nebo hydrogenosomy, které se stále nacházejí ve všech známých Eukarya. Později některé eukaryoty, které již obsahovaly mitochondrie, také pohltily organismy podobné sinicím. To vedlo k vytvoření chloroplasty v řasách a rostlinách. Existují také některé řasy, které pocházejí z ještě pozdějších endosymbiotických událostí. Zde eukaryota pohltila eukaryotické řasy, které se vyvinuly v plastid „druhé generace“.[73][74] Toto je známé jako sekundární endosymbióza.

Mariňák Thiomargarita namibiensis, největší známá bakterie

Sinice květy může obsahovat smrtelné cyanotoxiny

The chloroplasty z glaukofyty mít peptidoglykan vrstva, důkazy naznačující jejich endosymbiotikum původ z sinice.[75]

Bakterie Marinomonas arctica roste uvnitř arktického mořského ledu při teplotách pod bodem mrazu




Pelagibacter všudypřítomný a jeho příbuzní mohou být nejhojnějšími organismy v oceánu a tvrdí se, že jsou pravděpodobně nejhojnějšími bakteriemi na světě. Tvoří asi 25% všech mikrobiálních látek plankton buňky a v létě mohou představovat přibližně polovinu buněk přítomných v povrchové vodě mírného oceánu. Celková hojnost P. ubique a příbuzní se odhadují na přibližně 2 × 1028 mikroby.[76] Bylo však hlášeno v Příroda v únoru 2013, že bakteriofág HTVC010P, který útočí P. ubique, byl objeven a „je to pravděpodobně opravdu nejběžnější organismus na planetě“.[77][78]
Největší známá bakterie, mořská Thiomargarita namibiensis, mohou být viditelné pouhým okem a někdy dosahují 0,75 mm (750 μm).[79][80]
Mořské archaea

The archaea (Řek pro starověký[82]) tvoří a doména a království z jednobuněčný mikroorganismy. Tyto mikroby jsou prokaryoty, což znamená, že nemají žádné buněčné jádro nebo jakýkoli jiný vázaný na membránu organely ve svých celách.
Archaea byly původně klasifikovány jako bakterie, ale tato klasifikace je zastaralá.[83] Buňky archaealu mají jedinečné vlastnosti oddělující je od ostatních dvou domén života, Bakterie a Eukaryota. Archaea se dále dělí na několik uznaných phyla. Klasifikace je obtížná, protože většina nebyla izolována v laboratoři a byla detekována pouze jejich analýzou nukleové kyseliny ve vzorcích z jejich prostředí.
Archea a bakterie mají obecně podobnou velikost a tvar, i když několik archaeí má velmi podivné tvary, například ploché a čtvercové buňky Haloquadratum walsbyi.[84] Přes tuto morfologickou podobnost s bakteriemi mají archaea geny a několik metabolické cesty které jsou více příbuzné těm z eukaryot, zejména enzymy zahrnutý do něčeho, zůčastnit se čeho transkripce a překlad. Jiné aspekty archaeální biochemie jsou jedinečné, například jejich spoléhání se na etherové lipidy v jejich buněčné membrány, jako archaeols. Archaea využívá více zdrojů energie než eukaryota: ty se pohybují od organické sloučeniny, jako jsou cukry, do amoniak, ionty kovů nebo dokonce vodíkový plyn. Sůl-tolerantní archaea (dále jen Haloarchaea ) používají sluneční světlo jako zdroj energie a další druhy archea opravit uhlík; na rozdíl od rostlin a sinice, žádný známý druh archaea nedělá obojí. Archaea rozmnožovat nepohlavně podle binární dělení, fragmentace nebo pučící; na rozdíl od bakterií a eukaryot se netvoří žádné známé druhy výtrusy.
Archaea jsou zvláště četné v oceánech a archaea v plankton může být jednou z nejhojnějších skupin organismů na planetě. Archaea jsou hlavní součástí života Země a mohou hrát roli v obou uhlíkový cyklus a dusíkový cyklus. Crenarchaeota (eocyty) jsou kmenem archea, o kterém se předpokládá, že je v mořském prostředí velmi hojný a je jedním z hlavních přispěvatelů k fixaci uhlíku.[85]

Eocyty může být nejhojnější z mořských archaeí

Halobakterie, které se nacházejí ve vodě téměř nasycené solí, jsou nyní uznávány jako archaea.

Ploché buňky archaea ve tvaru čtverce Haloquadratum walsbyi

Methanosarcina barkeri, mořská archaea, která produkuje metan

Termofily, jako Pyrolobus fumarii přežít dobře přes 100 ° C




Eukaryoty

Všechny živé organismy lze rozdělit do obou skupin prokaryoty nebo eukaryoty. Život vznikl jako jednobuněčný prokaryoty a později se vyvinuly ve složitější eukaryoty. Na rozdíl od prokaryotických buněk jsou eukaryotické buňky vysoce organizované. Prokaryoty jsou bakterie a archaea, zatímco eukaryoty jsou další formy života - protistů, rostliny, houby a zvířata. Protisti jsou obvykle jednobuněční, zatímco rostliny, houby a zvířata jsou obvykle mnohobuněčný.
Zdá se velmi pravděpodobné, že kořen eukaryot leží v archaea; nejbližšími příbuznými dnes známými mohou být Heimdallarchaeota kmen navrhovaného Asgard nadkmen. Tato teorie je moderní verzí scénáře původně navrženého v roce 1984 jako Eocytová hypotéza, když Crenarchaeota (alias Eocyty) byli tehdy nejbližšími známými archaickými příbuznými eukaryot. Možné přechodná forma mikroorganismu mezi prokaryotem a eukaryotem objevili v roce 2012 japonští vědci. Parakaryon myojinensis je jedinečný mikroorganismus větší než typický prokaryot, ale s jaderným materiálem uzavřeným v membráně jako v eukaryotu a přítomností endosymbionty. Toto je považováno za první věrohodnou evoluční formu mikroorganismu, která ukazuje fázi vývoje od prokaryotu po eukaryot.[86][87]
Námořní protisti
Protisti jsou eukaryoty, které nelze klasifikovat jako rostliny, houby nebo zvířata. Obvykle jsou jednobuněčné a mikroskopické. Život vznikl jako jednobuněčné prokaryoty (bakterie a archea) a později se z nich vyvinula složitější eukaryoty. Eukaryoty jsou rozvinutější formy života známé jako rostliny, zvířata, houby a prvoci. Termín protist se historicky začal používat jako výhodný termín pro eukaryoty, které nelze striktně klasifikovat jako rostliny, zvířata nebo houby. Nejsou součástí moderní kladistiky, protože jsou paraphyletic (chybí společný předek).
Trofickým režimem
Protisty lze rozdělit do čtyř skupin podle toho, zda je jejich výživa rostlinná, zvířecí, plísňová,[88] nebo jejich směs.[89]
Protisti podle toho, jak dostávají jídlo | |||||||
|---|---|---|---|---|---|---|---|
| Typ protisty | Popis | Příklad | Některé další příklady | ||||
| Rostlinný | Autotrofní protisti, kteří si vyrábějí vlastní jídlo, aniž by museli konzumovat jiné organismy, obvykle pomocí fotosyntézy |  | Zelené řasy, Pyramimonas | Červené a hnědé řasy, rozsivky a nějaký dinoflageláty. Rostlinní protisté jsou důležitými složkami fytoplanktonu diskutováno níže. | |||
| Jako zvíře | Heterotrofní protisti, kteří dostávají potravu konzumující jiné organismy (bakterie, archea a malé řasy) |  | Radiolarian protist, jak je tažen Haeckel | Foraminiferans a některé námořní améby, náčelníci a bičíky. | |||
| Plísňové | Saprotrofní protisti, kteří získávají potravu ze zbytků organismů, které se rozpadly a rozpadly |  | Marine slizové sítě tvoří labyrintové sítě trubic, ve kterých mohou cestovat améby bez pseudopodů | Mořské lišejníky | |||
| Mixotrops | Rozličný | Mixotrofní a osmotrofický protisti, kteří získávají jídlo kombinací výše uvedeného |  | Euglena mutabilis, fotosyntetický bičíkatý | Mnoho mořských mixotropů se vyskytuje mezi protisty, zejména mezi nálevníky a dinoflageláty[90] | ||


Protisti jsou velmi různorodé organismy, které jsou v současné době organizovány do 18 kmenů, ale není snadné je zařadit.[92][93] Studie prokázaly, že v oceánech, hlubinných průduchech a říčních sedimentech existuje vysoká protistátní rozmanitost, což naznačuje, že ještě není nutné objevit velké množství eukaryotických mikrobiálních komunit.[94][95] Došlo k malému výzkumu mixotrofní protistům, ale nedávné studie v mořském prostředí zjistily, že mixotrofní protesty přispívají významnou částí protistů biomasa.[90] Protože protisté jsou eukaryoti, mají ve své buňce alespoň jeden jádro, stejně jako organely jako mitochondrie a Golgiho těla. Protisti jsou nepohlavní, ale dokážou se rychle reprodukovat mitóza nebo fragmentace.
- Jednobuněčné a mikroskopické protisty

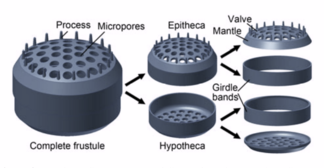
Diatomy mají sklo podobné buněčným stěnám oxid křemičitý a zavolal frustule.[97]

Fosilní roztoč frustule od 32-40 mya

Jednobuněčná řasa, Gephyrocapsa oceanica

Dva dinoflageláty

Zooxanthellae je fotosyntetická řasa, která žije uvnitř hostitelů korál

Jednobuněčný ciliate se zelenou zoochlorellae žijící uvnitř endosymbioticky










| Externí video | |
|---|---|
Ciliate požití rozsivky
Améba pohlcující rozsivku


Na rozdíl od buněk prokaryot jsou buňky eukaryot vysoce organizované. Rostliny, zvířata a houby jsou obvykle mnohobuněčný a jsou typicky makroskopické. Většina protistů je jednobuněčná a mikroskopická. Existují ale výjimky. Někteří jednobuněční mořští protisti jsou makroskopičtí. Některé mořské slizové formy mají jedinečné životní cykly, které zahrnují přepínání mezi jednobuněčnými, koloniální a mnohobuněčné formy.[98] Jiní mořští protisti nejsou ani jednobuněční, ani mikroskopičtí, jako např Mořská řasa.
- Makroskopické protisty (viz také jednobuněčné makrořasy → )

Jednobuněčný obří améba má až 1000 jádra a dosahuje délky 5 mm

Gromia sphaerica je velká sférická teste améba což vede k blátě. Jeho průměr je až 3,8 cm.[99]

Spiculosiphon oceana jednobuněčný foraminiferan se vzhledem a životním stylem, který napodobuje a mycí houba, dorůstá do délky 5 cm.

The xenofyor, žije další jednobuněčný foraminiferan hlubinné zóny. Má obrovskou skořápku o průměru až 20 cm.[100]

Obří řasa, a hnědé řasy, není skutečná rostlina, přesto je mnohobuněčná a může dorůst až 50 metrů





Protisti byli popsáni jako taxonomické pytle ztracených existencí, kde vše, co se nevejde do jednoho z hlavních biologická království Lze umístit.[101] Někteří moderní autoři upřednostňují vyloučit mnohobuněčné organismy z tradiční definice protisty, čímž omezují protisty na jednobuněčné organismy.[102][103] Tato omezenější definice vylučuje mnoho hnědý, mnohobuňečný Červené a zelené řasy, a slizové formy.[104]
Lokomoce
Další způsob kategorizace protistů je podle jejich způsobu lokomoce. Mnoho jednobuněčných protistů, zejména prvoků, je pohyblivý a může generovat pohyb použitím bičíky, řasy nebo pseudopody. Buňky, které používají k pohybu bičíky, se obvykle označují jako bičíky, buňky, které používají řasinky, se obvykle označují jako náčelníci a buňky, které používají pseudopody, se obvykle označují jako améba nebo améboidy. Ostatní protisti jsou ne pohyblivý, a proto nemají žádný pohybový mechanismus.
Protisti podle toho, jak se pohybují | ||||||||
|---|---|---|---|---|---|---|---|---|
| Typ protisty | Pohybový mechanismus | Popis | Příklad | Další příklady | ||||
| Pohyblivý | Bičíky |  | A bičík (Latinsky pro bič) je řasovitý přívěsek, který vyčnívá z buněčného těla některých protistů (stejně jako některých bakterií). Bičíky používají od jednoho do několika bičíků k pohybu a někdy jako krmení a smyslové vnímání organela. |  | Kryptofyty | Všechno dinoflageláty a nanoflageláty (choanoflagellates, silicoflagellates, většina zelené řasy )[105][106] (Ostatní protisté procházejí fází jako gamety když mají dočasné bičík - některé radiolariáni, foraminiferans a Apicomplexa ) | ||
| Ciliates |  | A cilium (Latinsky pro řasa) je malé bičík. Ciliates používají několik řasinek, které mohou mít stovky, aby se napájely vodou. |  | Paramecium bursaria kliknutím zobrazíte řasinky | Foraminiferans a některé námořní améby, náčelníci a bičíky. | |||
| Améby (améboidy) |  | Améba mají schopnost měnit tvar vysunutím a zasunutím pseudopody (Řek pro falešné nohy).[107] |  | Améba | Nalezeno v každém významném protistovi linie. Améboidní buňky se vyskytují mezi prvoky, ale také v řasy a houby.[108][109] | |||
| Ne pohyblivý | žádný |  | Diatom | Diatomy, coccolithophores a nepohyblivé druhy Phaeocystis[106] Mezi prvoky parazitující Apicomplexa jsou nemotorní. | ||||
Bičíky zahrnují bakterie i protisty. Model rotačního motoru používaný bakteriemi používá protony z elektrochemický gradient aby mohli přesunout své bičíky. Točivý moment v bičíku bakterií je tvořen částicemi, které vedou protony kolem základny bičíku. Směr otáčení bičíku v bakteriích pochází z obsazení protonových kanálů podél obvodu bičíkového motoru.[110]
Ciliates obecně mají stovky až tisíce řasinek, které jsou hustě zabalené v polích. Během pohybu se jednotlivý cilium deformuje pomocí výkonového zdvihu s vysokým třením, po kterém následuje obnovovací zdvih s nízkým třením. Vzhledem k tomu, že existuje více řasinek zabalených dohromady na jednotlivém organismu, zobrazují kolektivní chování v a metachronální rytmus. To znamená, že deformace jednoho cilium je ve fázi s deformací jeho souseda, což způsobuje deformační vlny, které se šíří po povrchu organismu. Tyto množící se řasinky umožňují organismu používat řasinky koordinovaně k pohybu. Typickým příkladem řasinkového mikroorganismu je Paramecium, jednobuněčný, řasinkatý prvok, pokrytý tisíci řasinek. Řasinky společně mlátí Paramecium pohánět vodou rychlostí 500 mikrometrů za sekundu.[111]
| Externí video | |
|---|---|
- Bičíkatý, nálevník a améba

Bakteriální bičík otočený o molekulární motor na jeho základně

Losos spermie

Paramecium krmení bakteriemi

Náměstek Oxytricha trifallax s řasinkami jasně viditelnými

Améba s požitými rozsivkami





Mořské houby


Více než 1 500 druhů houby jsou známy z mořského prostředí.[112] Jsou parazitující na mořské řasy nebo zvířata, nebo jsou saproby krmení mrtvými organickými látkami z řas, korálů, cyst prvoků, mořských trav a dalších substrátů.[113] Spóry mnoha druhů mají speciální přídavky, které usnadňují připevnění k substrátu.[114] Mořské houby lze nalézt také v mořská pěna a kolem hydrotermální oblasti oceánu.[115] Různorodá škála neobvyklých sekundárních metabolity je produkován mořskými houbami.[116]
Mykoplankton jsou saprotropní členové plankton společenství námořní a sladkovodní ekosystémy.[117][118] Skládají se z vláknitý volně žijící houby a kvasinky spojené s planktonovými částicemi nebo fytoplankton.[119] Podobný bakterioplankton, tyto vodní houby hrají významnou roli v heterotrofní mineralizace a koloběh živin.[120] I když je většinou mikroskopický, některý mykoplankton může mít průměr až 20 mm a délku přes 50 mm.[121]
Typický mililitr mořské vody obsahuje asi 103 do 104 buňky hub.[122] Toto číslo je větší v pobřežních ekosystémech a ústí řek kvůli nutričnímu odtoku ze suchozemských komunit. Vyšší rozmanitost mykoplanktonu se vyskytuje kolem pobřeží a v povrchových vodách až do 1000 metrů, s a svislý profil to záleží na tom, jak hojné fytoplankton je.[123][124] Tento profil se mezi sezónami mění kvůli změnám v dostupnosti živin.[125] Mořské houby přežívají v prostředí s konstantním nedostatkem kyslíku, a proto závisí na difúzi kyslíku turbulence a kyslík generovaný fotosyntetické organismy.[126]
Mořské houby lze klasifikovat jako:[126]
- Nižší houby - přizpůsobené mořským stanovištím (zoosporický houby, včetně mastigomycetů: oomycety a chytridiomycetes )
- Vyšší houby - vláknité, upravené na planktonický životní styl (hyphomycetes, ascomycetes, bazidiomycety ). Většina druhů mykoplanktonu jsou vyšší houby.[123]
Lišejníky jsou mutualistic asociace mezi houbou, obvykle an ascomycete a řasa nebo a sinice. Několik lišejníků se vyskytuje v mořském prostředí.[127] Mnoho dalších se vyskytuje v stříkající zóna, kde zaujímají různé vertikální zóny podle toho, jak tolerantní jsou vůči ponoření.[128] Některé lišejníky žijí dlouho; jeden druh byl datován na 8 600 let.[129] Jejich životnost je však obtížně měřitelná, protože to, co definuje stejný lišejník, není přesné.[130] Lišejníky rostou vegetativním odlomením kousku, který může nebo nemusí být definován jako stejný lišejník, a mohou se sloučit dva lišejníky různého věku, což vyvolává otázku, zda se jedná o stejný lišejník.[130]
The mořský šnek Littoraria irrorata poškozuje rostliny Spartina v mořských bažinách, kde žije, což umožňuje kolonizaci rostliny spory přílivových ascomycetous hub. Hlemýžď pak sžírá růst hub přednostně před samotnou trávou.[131]
Podle fosilních záznamů se houby datují pozdě Proterozoikum éra před 900-570 miliony let. V Číně byly objeveny fosilní mořské lišejníky staré 600 milionů let.[132] Předpokládá se, že mykoplankton se vyvinul z suchozemských hub, pravděpodobně v Paleozoikum éry (před 390 miliony let).[133]
Mořští živočichové
| Externí video | |
|---|---|
Jako mladiství se zvířata vyvíjejí z mikroskopických stádií, která mohou zahrnovat výtrusy, vejce a larvy. Alespoň jedna skupina mikroskopických zvířat, parazitický cnidarian Myxozoa, je jednobuněčný ve své dospělé formě a zahrnuje mořské druhy. Ostatní dospělí mořští mikroorganismy jsou mnohobuněčné. Mikroskopický dospělý členovci se běžněji vyskytují ve vnitrozemí ve sladké vodě, ale existují i mořské druhy. Mikroskopický dospělý námořní korýši zahrnout některé copepods, perloočka a tardigrades (vodní medvědi). Nějaké námořní hlístice a vířníky jsou také příliš malé na to, aby je bylo možné rozpoznat pouhým okem, stejně jako mnoho jiných loricifera, včetně nedávno objevených anaerobní druhy, které tráví život v anoxický životní prostředí.[134][135] Copepods více přispívají k sekundární produktivita a uhlíkový dřez světových oceánů než jakákoli jiná skupina organismů.
- Mořští živočichové

Darkfield fotka a gastrotrich, 0,06–3,0 mm dlouhý, červovitý živočich žijící mezi částicemi sedimentu

Obrněný Pliciloricus enigmaticus, dlouhý asi 0,2 mm, žije v prostorech mezi mořským štěrkem

Vířníky, obvykle 0,1–0,5 mm dlouhé, mohou vypadat jako protisti, ale jsou mnohobuněčné a patří do skupiny Animalia

Tardigrades (vodní medvědi), asi 0,5 mm dlouhý, patří k nejodolnějším známým zvířatům




Prvovýrobci

Prvovýrobci jsou autotrofní organismy, které vytvářejí své vlastní jídlo, místo aby jedly jiné organismy. To znamená, že primární producenti se stávají výchozím bodem v EU potravní řetězec pro heterotrofní organismy, které jedí jiné organismy. Někteří mořští primární producenti jsou specializované bakterie a archea, které jsou chemotrofy, připravují si vlastní jídlo shromažďováním hydrotermální průduchy a studené prosakuje a pomocí chemosyntéza. Nicméně většina námořní prvovýroba pochází z organismů, které používají fotosyntéza na oxidu uhličitém rozpuštěném ve vodě. Tento proces využívá energii ze slunečního záření k přeměně vody a oxid uhličitý[136]:186–187 na cukry, které lze použít jak jako zdroj chemické energie, tak i organických molekul, které se používají ve strukturních složkách buněk.[136]:1242 Prvotní mořští producenti jsou důležití, protože podporují téměř celý život mořských živočichů generováním většiny kyslík a jídlo, které poskytuje jiným organismům chemickou energii, kterou potřebují k existenci.
Hlavními mořskými primárními producenty jsou sinice, řasy a mořské rostliny. The kyslík uvolněný jako vedlejší produkt fotosyntézy je zapotřebí téměř všechno živé buněčné dýchání. Primární producenti mají navíc celosvětový vliv uhlík a voda cykly. Stabilizují pobřežní oblasti a mohou poskytovat stanoviště mořským živočichům. Termín divize byl tradičně používán místo kmen když diskutujeme o prvovýrobcích, ale Mezinárodní kód nomenklatury pro řasy, houby a rostliny nyní přijímá oba výrazy jako ekvivalenty.[137]
Sinice



| Externí video | |
|---|---|
Sinice byly prvními organismy, které vyvinuly schopnost přeměňovat sluneční světlo na chemickou energii. Tvoří kmen (rozdělení) bakterií, které se pohybují od jednobuněčných po vláknitý a zahrnout koloniální druhy. Vyskytují se téměř všude na Zemi: ve vlhké půdě, ve sladkovodním i mořském prostředí a dokonce i na antarktických skalách.[139] Zejména některé druhy se vyskytují jako unášené buňky plovoucí v oceánu a jako takové byly mezi prvními z fytoplankton.
První primární producenti, kteří používali fotosyntézu, byli oceánští sinice asi před 2,3 miliardami let.[140][141] Uvolňování molekulárních kyslík podle sinice jako vedlejší produkt fotosyntézy vyvolaných globálních změn v prostředí Země. Protože kyslík byl v té době toxický pro většinu života na Zemi, vedlo to k téměř vyhynutí organismy netolerující kyslík, a dramatická změna který přesměroval vývoj hlavních živočišných a rostlinných druhů.[142]
Malý (0,6 µm ) mořské sinice Prochlorococcus, objevený v roce 1986, dnes tvoří důležitou součást základny oceánu potravní řetězec a představuje velkou část fotosyntézy otevřeného oceánu[143] a odhadem 20% kyslíku v zemské atmosféře.[144] Je to pravděpodobně nejhojnější rod na Zemi: jeden mililiter povrchové mořské vody může obsahovat 100 000 buněk nebo více.[145]
Biologové si původně mysleli sinice was algae, and referred to it as "blue-green algae". The more recent view is that cyanobacteria is a bacteria, and hence is not even in the same Království as algae. Most authorities exclude all prokaryoty, and hence cyanobacteria from the definition of algae.[146][147]
Řasy
Řasy is an informal term for a widespread and diverse group of photosynthetic protistů which are not necessarily closely related and are thus polyfyletický. Marine algae can be divided into six groups: zelený, Červené a hnědé řasy, euglenophytes, dinoflageláty a rozsivky.
Dinoflagellates and diatoms are important components of marine algae and have their own sections below. Euglenophytes are a phylum of unicellular flagellates with only a few marine members.
Not all algae are microscopic. Green, red and brown algae all have multicellular macroscopic forms that make up the familiar mořské řasy. Zelené řasy, an informal group, contains about 8,000 recognised species.[148] Many species live most of their lives as single cells or are filamentous, while others form kolonie made up from long chains of cells, or are highly differentiated macroscopic seaweeds. Červené řasy, a (disputed) phylum contains about 7,000 recognised species,[149] většinou mnohobuňečný and including many notable seaweeds.[149][150] Hnědé řasy formulář a třída containing about 2,000 recognised species,[151] většinou mnohobuňečný and including many seaweeds such as kelp.Unlike higher plants, algae lack roots, stems, or leaves. They can be classified by size as mikrořasy nebo makrořasy.
Mikrořasy are the microscopic types of algae, not visible to the naked eye. Většinou jsou jednobuněčný species which exist as individuals or in chains or groups, though some are mnohobuňečný. Microalgae are important components of the marine protists discussed above, as well as the phytoplankton diskutováno níže. Oni jsou velmi různorodý. It has been estimated there are 200,000-800,000 species of which about 50,000 species have been described.[152] Depending on the species, their sizes range from a few micrometers (µm) to a few hundred micrometers. They are specially adapted to an environment dominated by viscous forces.

Chlamydomonas globosa, a unicellular green alga with two bičíky just visible at bottom left

Chlorella vulgaris, a common green mikrořasy, v endosymbióza s ciliate[153]

Centric diatom

Dinoflageláty





Makrořasy are the larger, mnohobuňečný and more visible types of algae, commonly called mořské řasy. Seaweeds usually grow in shallow coastal waters where they are anchored to the seafloor by a sevření. Like microalgae, macroalgae (seaweeds) can be regarded as marine protists since they are not true plants. But they are not microorganisms, so they are not within the scope of this article.
Unicellular organisms are usually microscopic, less than one tenth of a millimeter long. Existují výjimky. Mermaid's wineglass, a genus of subtropical zelené řasy, is single-celled but remarkably large and complex in form with a single large nucleus, making it a model organism for studying buněčná biologie.[154] Another single-celled algae, Caulerpa taxifolia, has the appearance of a vascular plant including "leaves" arranged neatly up stalks like a fern. Selective breeding in aquariums to produce hardier strains resulted in an accidental release into the Mediterranean where it has become an invazivní druhy known colloquially as killer algae.[155]
Marine microplankton


Plankton (from Greek for wanderers) are a diverse group of organisms that live in the vodní sloup of large bodies of water but cannot swim against a current. As a result, they wander or drift with the currents.[156] Plankton are defined by their ekologický výklenek, not by any fylogenetické nebo taxonomické klasifikace. They are a crucial source of food for many marine animals, from píce ryby na velryby. Plankton can be divided into a plant-like component and an animal component.
Fytoplankton

Fytoplankton are the plant-like components of the plankton community ("phyto" comes from the Greek for rostlina). Oni jsou autotrofní (self-feeding), meaning they generate their own food and do not need to consume other organisms.
Phytoplankton perform three crucial functions: they generate nearly half of the world atmospheric oxygen, they regulate ocean and atmospheric carbon dioxide levels, and they form the base of the marine web s potravinami. When conditions are right, květy of phytoplankton algae can occur in surface waters. Fytoplankton jsou r-stratégové which grow rapidly and can double their population every day. The blooms can become toxic and deplete the water of oxygen. However, phytoplankton numbers are usually kept in check by the phytoplankton exhausting available nutrients and by grazing zooplankton.[159]
Phytoplankton consist mainly of microscopic photosynthetic eukaryoty which inhabit the upper sunlit layer in all oceans. They need sunlight so they can photosynthesize. Most phytoplankton are single-celled algae, but other phytoplankton are bacteria and some are protistů.[160] Phytoplankton include cyanobacteria (above), rozsivky, various other types of řasy (red, green, brown, and yellow-green), dinoflageláty, euglenoids, coccolithophorids, kryptomonády, chlorofyty, prasinophytes, a silicoflagellates. They form the base of the prvovýroba that drives the ocean web s potravinami, and account for half of the current global primary production, more than the terrestrial forests.[161]
- Fytoplankton

Phytoplankton are the foundation of the ocean food chain

They come in many shapes and sizes.

Koloniální fytoplankton

The cyanobacterium Prochlorococcus accounts for much of the ocean's primary production

Zelený sinice scum washed up on a rock in California




Diatomy


Diatomy form a (disputed) phylum containing about 100,000 recognised species of mainly unicellular algae. Diatoms generate about 20 percent of the oxygen produced on the planet each year,[96] take in over 6.7 billion metric tons of křemík each year from the waters in which they live,[162] and contribute nearly half of the organic material found in the oceans.

Diatomy are one of the most common types of phytoplankton

They come in many shapes

Their protective shells (frustles) are made of silicon







Diatoms are enclosed in protective silica (glass) shells called frustule. Each frustule is made from two interlocking parts covered with tiny holes through which the diatom exchanges nutrients and wastes.[159] The frustules of dead diatoms drift to the ocean floor where, over millions of years, they can build up as much as half a mile deep.[163]
Silicified frustule of a pennate diatom with two overlapping halves

Fossil diatom

There are over 100,000 species of rozsivky which account for 50% of the ocean's primary production



| Externí video | |
|---|---|
Coccolithophores
Coccolithophores are minute unicellular photosynthetic protists with two flagella for locomotion. Most of them are protected by a shell covered with ornate circular plates or scales called kokcithové. The coccoliths are made from calcium carbonate. The term coccolithophore derives from the Greek for a seed carrying stone, referring to their small size and the coccolith stones they carry. Under the right conditions they bloom, like other phytoplankton, and can turn the ocean milky white.[166]





Kvetou řasy z Emiliania huxleyi off the southern coast of England




Mikrobiální rhodopsin

(2) it changes its configuration so a proton is expelled from the cell
(3) the chemical potential causes the proton to flow back to the cell
(4) thus generating energy
(5) in the form of adenosintrifosfát.[168]
Phototrophic metabolism relies on one of three energy-converting pigments: chlorofyl, bacteriochlorophyll, a sítnice. Retinal is the chromofor nalezen v rhodopsiny. The significance of chlorophyll in converting light energy has been written about for decades, but phototrophy based on retinal pigments is just beginning to be studied.[169]

In 2000 a team of microbiologists led by Edward DeLong made a crucial discovery in the understanding of the marine carbon and energy cycles. They discovered a gene in several species of bacteria[171][172] responsible for production of the protein rhodopsin, previously unheard of in bacteria. These proteins found in the cell membranes are capable of converting light energy to biochemical energy due to a change in configuration of the rhodopsin molecule as sunlight strikes it, causing the pumping of a proton from inside out and a subsequent inflow that generates the energy.[173] The archaeal-like rhodopsins have subsequently been found among different taxa, protists as well as in bacteria and archaea, though they are rare in complex mnohobuněčné organismy.[174][175][176]
Research in 2019 shows these "sun-snatching bacteria" are more widespread than previously thought and could change how oceans are affected by global warming. "The findings break from the traditional interpretation of marine ecology found in textbooks, which states that nearly all sunlight in the ocean is captured by chlorophyll in algae. Instead, rhodopsin-equipped bacteria function like hybrid cars, powered by organic matter when available — as most bacteria are — and by sunlight when nutrients are scarce."[177][169]
Tady je astrobiological conjecture called the Hypotéza Fialové Země which surmises that original life forms on Earth were retinal-based rather than chlorophyll-based, which would have made the Earth appear purple instead of green.[178][179]
Redfield and f- ratios
Během třicátých let Alfred C. Redfield found similarities between the composition of elements in phytoplankton and the major dissolved nutrients in the deep ocean.[180] Redfield proposed that the ratio of carbon to nitrogen to phosphorus (106:16:1) in the ocean was controlled by the phytoplankton’s requirements, as phytoplankton subsequently release nitrogen and phosphorus as they remineralize. This ratio has become known as the Redfieldův poměr, and is used as a fundamental principle in describing the stechiometrie of seawater and phytoplankton evolution.[181]
However, the Redfield ratio is not a universal value and can change with things like geographical latitude.[182] Based on allocation of resources, phytoplankton can be classified into three different growth strategies: survivalist, bloomer and generalist. Survivalist phytoplankton has a high N:P ratio (>30) and contains an abundance of resource-acquisition machinery to sustain growth under scarce resources. Bloomer phytoplankton has a low N:P ratio (<10), contains a high proportion of growth machinery and is adapted to exponential growth. Obecný fytoplankton má podobný poměr N: P jako Redfieldův poměr a obsahuje relativně stejný mechanismus získávání a růstu zdrojů.[181]
The f-ratio is the fraction of total prvovýroba fuelled by dusičnan (as opposed to that fuelled by other dusík sloučeniny jako amonný ). The ratio was originally defined by Richard Eppley and Bruce Peterson in one of the first doklady estimating global oceanic production.[183]
Zooplankton
Zooplankton are the animal component of the planktonic community ("zoo" comes from the Greek for zvíře). Oni jsou heterotrofní (jiné krmení), což znamená, že nemohou vyrábět své vlastní jídlo a musí místo toho konzumovat jako jídlo jiné rostliny nebo zvířata. To zejména znamená, že jedí fytoplankton.
Zooplankton je obecně větší než fytoplankton, většinou stále mikroskopický, ale některé lze pozorovat pouhým okem. Mnoho prvoky (jednobuněčné protistů že kořist na jiný mikroskopický život) jsou zooplankton, včetně zooflageláty, foraminiferans, radiolariáni, někteří dinoflageláty a mořská mikroorganismy. Macroscopic zooplankton (not generally covered in this article) include pelagic cnidarians, ctenofory, měkkýši, členovci a pláštěnci, stejně jako plankton šípové červy a štětinatí červi.

Microzooplankton: hlavní pastevci planktonu ...


Mnoho druhů prvoky (eukaryoty ) a bakterie (prokaryoty ) prey on other microorganisms; the feeding mode is evidently ancient, and evolved many times in both groups.[184][185][186] Among freshwater and marine zooplankton, whether single-celled or multi-cellular, predatory grazing on fytoplankton and smaller zooplankton is common, and found in many species of nanoflagellates, dinoflageláty, ciliates, vířníky, a diverse range of meroplankton animal larvae, and two groups of crustaceans, namely copepods a perloočky.[187]
Radiolariáni


Radiolariáni jsou jednobuněční predátoři protistů obalené propracovanými kulovitými skořápkami obvykle vyrobenými z oxidu křemičitého a propíchnuté otvory. Jejich název pochází z latiny pro „radius“. Chytí kořist prodloužením částí těla skrz otvory. Stejně jako křemíkové frustule rozsivek, mohou radiolarianské skořápky klesnout na oceánské dno, když radiolarianové zemřou a stanou se zachovány jako součást oceánského sedimentu. Tyto pozůstatky, jako mikrofosílie, poskytnout cenné informace o minulých oceánských podmínkách.[188]

Stejně jako rozsivky mají radiolariáni mnoho tvarů

Stejně jako rozsivky jsou radiolarianské skořápky obvykle vyrobeny ze silikátu

nicméně acantharian radiolariáni mají granáty vyrobené z síran strontnatý krystaly

Výřez schematického sférického radiolarianského pláště





closely replicate some radiolarian shell patterns[189]
| Externí video | |
|---|---|

Cladococcus abietinus
Foraminiferans


Jako radiolariáni foraminiferans (forams zkráceně) jsou jednobuněční draví protisti, také chráněni mušlemi, které mají otvory. Jejich název pochází z latiny pro „nositele děr“. Jejich granáty, často volané testy, jsou komorové (forams přidávají další komory, jak rostou). Mušle jsou obvykle vyrobeny z kalcitu, ale někdy jsou vyrobeny z aglutinovaný částice sedimentu nebo chiton a (zřídka) oxid křemičitý. Většina foramů je bentická, ale asi 40 druhů je planktických.[190] Jsou široce prozkoumány s dobře zavedenými fosilními záznamy, které vědcům umožňují odvodit hodně o minulých prostředích a podnebí.[188]


| Externí video | |
|---|---|

část zobrazující komory spirálového foramu

Žít Amoniak tepida streamovaná granulovaná ektoplazma pro chytání potravy

Skupina planktonických foramů

Fosilní nummulitid forams of various sizes from the Eocen

The Egyptské pyramidy byly vyrobeny z vápence, který obsahoval nummulity.[191]




A number of forams are mixotrofní (viz. níže ). Ty mají jednobuněčné řasy tak jako endosymbionty z různých linií, jako je zelené řasy, červené řasy, zlaté řasy, rozsivky, a dinoflageláty.[190] Mixotrofní foraminifery jsou zvláště běžné v oceánských vodách chudých na živiny.[192] Některé forams jsou kleptoplast, zachování chloroplasty od požití řas po chování fotosyntéza.[193]
Améba



Naked amoeba showing food vacuoles and ingested diatom

Shell nebo test a teste améba, Arcella sp.

Xenogenní testate amoeba covered in diatoms (from Penard's Amoeba Collection )




Ciliates

Tintinnopsis campanula

Holophyra vajíčko





Mesodinium rubrum produce deep red blooms using enslaved chloroplasts from their algal prey [194]
Several taxa of ciliates interacting
Blepharisma americanum swimming in a drop of pond water with other microorganisms



Mixotrofy


A mixotrof je organismus, který může používat různé kombinace zdroje energie a uhlíku, místo toho, aby měl jeden trofický režim na kontinuu od úplného autotrofie na jednom konci do heterotrofie na druhé straně. Odhaduje se, že mixotrofy tvoří více než polovinu veškerého mikroskopického planktonu.[195] Existují dva typy eukaryotických mixotrofů: ty s vlastními chloroplasty a ti s endosymbionty —A další, kteří je získávají prostřednictvím kleptoplastika nebo zotročením celé fototrofní buňky.[196]
Rozdíl mezi rostlinami a zvířaty se často rozpadá u velmi malých organismů. Možné kombinace jsou fotografie- a chemotrofie, litho- a organotrofie, auto- a heterotrofie nebo jiné jejich kombinace. Mixotrofy mohou být buď eukaryotický nebo prokaryotický.[197] Mohou využívat různé podmínky prostředí.[198]
Nedávné studie mořského mikrozooplanktonu zjistily, že 30–45% hojnosti nálevníků bylo mixotrofních a až 65% améboidů, foramů a radiolarianů biomasa byl mixotrofní.[90]
Phaeocystis je důležitý rod řas, který se vyskytuje jako součást moří fytoplankton okolo světa. Má to polymorfní životní cyklus, od volně žijících buněk až po velké kolonie.[199] Má schopnost vytvářet plovoucí kolonie, kde jsou stovky buněk vloženy do gelové matrice, která se může během květy.[200] Jako výsledek, Phaeocystis je důležitým přispěvatelem do moří uhlík[201] a cykly síry.[202] Phaeocystis druhy jsou endosymbionty acantharian radiolariáni.[203][204]
Mixotrophic plankton that combine phototrophy and heterotrophy – table based on Stoecker et. al., 2017 [205] | |||||||
|---|---|---|---|---|---|---|---|
| General types | Popis | Příklad | Další příklady | ||||
| Bacterioplankton | Photoheterotrophic bakterioplankton |  | Vibrio cholerae | Roseobacter spp. Erythrobacter spp. Gammaproteobacterial clade OM60 Widespread among bacteria and archaea | |||
| Fytoplankton | Volala constitutive mixotrophs Mitra et. al., 2016.[206] Phytoplankton that eat: photosynthetic protists with inherited plastidy and the capacity to ingest prey. |  | Ochromonas druh | Ochromonas spp. Prymnesium parvum Dinoflagellate examples: Fragilidium subglobosum,Heterocapsa triquetra,Karlodinium veneficum,Neoceratium furca,Prorocentrum minimální | |||
| Zooplankton | Volala nekonstitutivní mixotrofy Mitra et. al., 2016.[206] Zooplankton, který je fotosyntetický: mikrozooplankton nebo metazoan zooplankton, který získává fototrofii retencí chloroplastůA nebo údržba endosymbiontů řas. | ||||||
| Generalisté | Protisti, kteří zadržují chloroplasty a zřídka jiné organely z mnoha taxonů řas |  | Většina oligotrich nálevníci, kteří zadržují plastidyA | ||||
| Specialisté | 1. Protisti, kteří zadržují chloroplasty a někdy i jiné organely z jednoho druhu řas nebo velmi blízkých druhů řas |  | Dinophysis acuminata | Dinofýza spp. Myrionecta rubra | |||
| 2. Protisti nebo zooplankton s řasovými endosymbionty pouze jednoho druhu řas nebo velmi blízce příbuzných druhů řas |  | Noctiluca scintillans | Metazooplankton s řasami endosymbionty Většina mixotrofních Rhizaria (Acantharea, Polycystinea, a Foraminifera ) Zelený Noctiluca scintillans | ||||
- Mixoplankton

Tintinnid ciliate Favella

Euglena mutabilis, fotosyntetický bičíkatý

Zoochlorellae (zelená) žijící uvnitř ciliate Stichotricha secunda


Dinoflageláty


Dinoflageláty jsou součástí skupina řas, a tvoří kmen jednobuněčných bičíkovců s asi 2 000 mořskými druhy.[207] Název pochází z řeckého slova „dinos“ víření a latina "flagellum" znamená a bič nebo šlehat. To se týká dvou bičovitých nástavců (bičíků) používaných pro pohyb vpřed. Většina dinoflagelátů je chráněna červenohnědým celulózovým pancířem. Stejně jako ostatní fytoplankton jsou i dinoflageláty r-stratégové které za správných podmínek mohou květ a tvořit červené přílivy. Vykopává může být nejvíce bazální bičíkovitá linie.[105]
By trophic orientation dinoflagellates cannot be uniformly categorized. Je známo, že některé dinoflageláty jsou fotosyntetický, ale velká část z nich je ve skutečnosti mixotrofní kombinující fotosyntézu s požitím kořisti (fagotrofie ).[208] Některé druhy jsou endosymbionty mořských živočichů a dalších protistů a hrají důležitou roli v biologii korálové útesy. Jiné předcházejí jiným prvokům a několik forem je parazitických. Mnoho dinoflagelátů je mixotrofní a lze je také klasifikovat jako fytoplankton.
Toxický dinoflagelát Dinophysis acuta získat chloroplasty ze své kořisti. "It cannot catch the cryptophytes byitself, and instead relies on ingesting ciliates such as the red Myrionecta rubra, which sequester their chloroplasts from aspecific cryptophyte clade (Geminigera/Plagioselmis/Teleaulax)".[205]

Gyrodinium, jeden z mála nahých dinoflagelátů, kterým chybí brnění

Dinoflagelát Protoperidinium vytlačuje velký krmný závoj, aby zachytil kořist

Nassellarian radiolariáni mohou být v symbióze s dinoflageláty

Dinoflagelát Dinophysis acuta








Dinoflageláty často žijí symbióza s jinými organismy. Mnoho nassellarian radiolarianský dům dinoflagellate symbionty v rámci jejich testů.[210] Nassellarian poskytuje amonný a oxid uhličitý pro dinoflagelát, zatímco dinoflagelát poskytuje nassellarianovi sliznici užitečnou pro lov a ochranu před škodlivými útočníky.[211] Existují důkazy z DNA analýza, že symbióza dinoflagelátů s radiolariány se vyvinula nezávisle na jiných dinoflagelátových symbiózách, jako například foraminifera.[212]
Some dinoflagellates are bioluminiscenční. At night, ocean water can light up internally and sparkle with blue light because of these dinoflagellates.[213][214] Bioluminescent dinoflagellates possess scintillons, individual cytoplazmatický bodies which contain dinoflagellate luciferase, the main enzyme involved in the luminescence. The luminescence, sometimes called the phosphorescence of the sea, occurs as brief (0.1 sec) blue flashes or sparks when individual scintillons are stimulated, usually by mechanical disturbances from, for example, a boat or a swimmer or surf.[215]

Tripos muelleri je rozeznatelný podle rohů ve tvaru písmene U.

Oodinium, rod parazitický dinoflageláty, příčiny sametová nemoc v rybách[216]

Karenia brevis produkuje červené přílivy vysoce toxické pro člověka[217]

Noctiluca scintillans, a bioluminescent dinoflagellate[218]





Marine microfossils
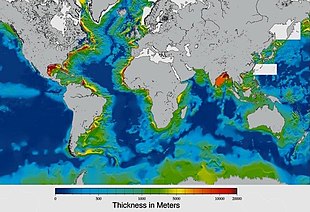
Sediments at the bottom of the ocean have two main origins, terrigenous and biogenous.
Terrigenous sediments account for about 45% of the total marine sediment, and originate in the erosion of skály on land, transported by rivers and land runoff, windborne dust, volcanoes, or grinding by glaciers.
Biogenous sediments account for the other 55% of the total sediment, and originate in the skeletal remains of marine protists (single-celled plankton and benthos microorganisms). Much smaller amounts of precipitated minerals and meteoric dust can also be present. Sliz, in the context of a marine sediment, does not refer to the consistency of the sediment but to its biological origin. The term ooze was originally used by John Murray, the "father of modern oceanography", who proposed the term radiolarian ooze for the silica deposits of radiolarian shells brought to the surface during the Expedice Challenger.[219] A biogenic ooze je pelagický sediment containing at least 30 percent from the skeletal remains of marine organisms.
Main types of biogenic ooze | ||||||||
|---|---|---|---|---|---|---|---|---|
| typ | minerální formuláře | protist zapojen | name of skeleton | typical size (mm) | ||||
| Křemičitý bláto | SiO2 oxid křemičitý křemen sklenka opál chert | rozsivka |  | frustrace | 0.002 to 0.2[220] |  | rozsivka mikrofosilní from 40 million years ago | |
| radiolarian |  | test or shell | 0.1 to 0.2 |  | elaborate silica shell of a radiolarian | |||
| Calcareous ooze | CaCO3 kalcit aragonit vápenec mramor křída | foraminiferan |  | test or shell | under 1 |  | Vápenatý test of a planktic foraminiferan. There are about 10,000 living species of foraminiferans[221] | |
| coccolithophore |  | kokcithové | under 0.1[222] |  | Coccolithophores are the largest global source of biogenic calcium carbonate, and significantly contribute to the global carbon cycle.[223] They are the main constituent of chalk deposits such as the bílé útesy Doveru. | |||

Křemelina is a soft, křemičitý, sedimentární hornina made up of microfossils in the form of the frustule (shells) of single cell rozsivky (click to magnify)

Ilustrace a Globigerina sliz

Mušle (testy ), usually made of calcium carbonate, from a foraminiferal ooze on the deep ocean floor





Opal can contain protist microfossils of diatoms, radiolarians, silicoflagellates and ebridians [225]

Marble can contain protist microfossils of foraminiferans, coccolithophores, calcareous nannoplankton and algae, ostracodes, pteropody, calpionellids and bryozoa [225]





Within each colored area, the type of material shown is what dominates, although other materials are also likely to be present.
For further information, viz zde
Marine microbenthos

Marine microbenthos are microorganisms that live in the bentická zóna of the ocean – that live near or on the seafloor, or within or on surface seafloor sediments. Slovo bentos comes from Greek, meaning "depth of the sea". Mikrobentos se vyskytuje všude na mořském dně kontinentálních šelfů nebo kolem nich, stejně jako v hlubších vodách s větší rozmanitostí v sedimentech na mořském dně nebo na nich. V mělkých vodách poskytují mořské louky, korálové útesy a řasy obzvláště bohaté stanoviště. v fotonické zóny bentické rozsivky dominují jako fotosyntetické organismy. v přílivové zóny měnící se přílivy a odlivy silně kontrolovat příležitosti pro mikrobentos.

Elphidium rozšířený hojný rod bentických foramů

Heterohelix, vyhynulý rod bentických foramů


Foraminifera i rozsivky mají planktonika a bentický formy, to znamená, že se mohou pohybovat v vodní sloup nebo žít na sedimentu na dně oceánu. Ať tak či onak, jejich skořápky skončí na mořském dně poté, co zemřou. Tyto skořápky jsou široce používány jako klimatické proxy. Chemické složení skořápek je důsledkem chemického složení oceánu v době, kdy byly skořápky vytvořeny. Minulé teploty vody lze odvodit také z poměrů stabilní izotopy kyslíku ve skořápkách, protože lehčí izotopy se snadněji odpařují v teplejší vodě a zanechávají těžší izotopy ve skořápkách. Informace o minulém podnebí lze odvodit dále z množství foramů a rozsivek, protože v teplé vodě bývají hojnější.[226]
Náhlé událost zániku který před 66 miliony let zabil dinosaury, také vyhynuli tři čtvrtiny všech ostatních živočišných a rostlinných druhů. Následně však vzkvétaly hlubinné bentické foramy. V roce 2020 bylo oznámeno, že vědci zkoumali chemické složení tisíců vzorků těchto bentických foramů a pomocí svých poznatků vytvořili nejpodrobnější klimatický záznam Země vůbec.[227][228]
Nějaký endolity mají extrémně dlouhý život. V roce 2013 uvedli vědci důkazy o endolitech v oceánském dně, starých možná miliony let, s generační dobou 10 000 let.[229] Ty se pomalu metabolizují a nejsou v klidovém stavu. Nějaký Aktinobakterie nalezen v Sibiř se odhadují na půl milionu let.[230][231][232]
Mořské mikrobiomy
Symbióza a holobionti
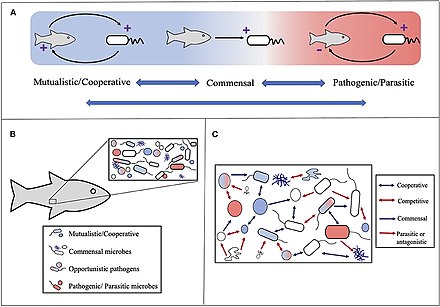
Pojem holobiont původně definoval Dr. Lynn Margulis ve své knize z roku 1991 Symbióza jako zdroj evolučních inovací jako shromáždění a hostitel a mnoho dalších druhů žijících v něm nebo v jeho okolí, které společně tvoří diskrétní ekologická jednotka.[234] Složky holobiontu jsou jednotlivé druhy nebo bionty, zatímco kombinované genom ze všech biontů je hologenome.[235]
Koncept se od této původní definice následně vyvinul,[236] se zaměřením pohybujícím se na mikrobiální druhy spojené s hostitelem. Holobiont tedy zahrnuje hostitele, virom, mikrobiom, a další členové, kteří všichni nějakým způsobem přispívají k fungování celku.[237][238] Holobiont obvykle zahrnuje a eukaryot hostitel a všechny symbiotický viry, bakterie, houby atd., které žijí uvnitř nebo uvnitř.[239]
Existuje však kontroverze o tom, zda lze holobionty považovat za jednotlivé evoluční jednotky.[240]


Korály vytvářející útes jsou dobře studované holobionty, které zahrnují samotný korál (eukaryotický bezobratlý ve třídě Anthozoa ), fotosyntetický dinoflageláty volala zooxanthellae (Symbiodinium ) a související bakterie a viry.[245] Koevoluční vzorce existují pro korálová mikrobiální společenství a korálovou fylogenezi.[246]
Mořské jídlo web
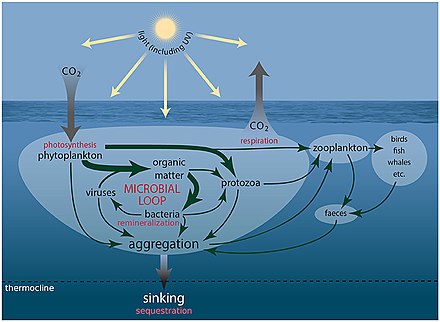
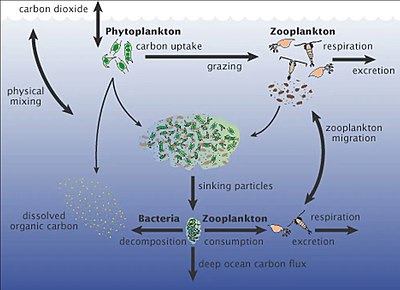
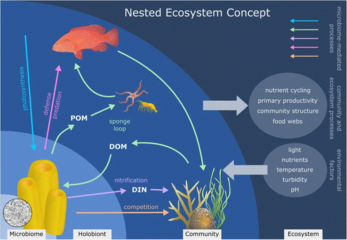
Mořské mikroorganismy hrají ústřední roli v web mořských potravin.
The virový zkrat Pathway je mechanismus, který zabraňuje mořským mikrobiálním látkám částice organické hmoty (POM) z migrace nahoru trofické úrovně jejich recyklací do rozpuštěná organická hmota (DOM), které mohou mikroorganismy snadno absorbovat.[247] Virový posun pomáhá udržovat rozmanitost v mikrobiálním ekosystému tím, že brání jedinému druhu mořských mikrobů v ovládnutí mikroprostředí.[248] DOM recyklovaný cestou virového zkratu je srovnatelný s množstvím generovaným jinými hlavními zdroji mořského DOM.[249]


| Externí video | |
|---|---|

Pelagibacter všudypřítomný, nejhojnější bakterie v oceánu, hraje v celosvětovém měřítku významnou roli uhlíkový cyklus.
 Mořský sníh je sprcha organických částic, která padá z horních vod do hlubokého oceánu [251] Je významným vývozcem uhlíku.
Mořský sníh je sprcha organických částic, která padá z horních vod do hlubokého oceánu [251] Je významným vývozcem uhlíku.



Mezikontinentální komunity



Mikrobiální společenství mořského ledu (SIMCO) odkazují na skupiny mikroorganismy žijící uvnitř a na rozhraní mořský led u pólů. Ledová matice, kterou obývají, má silné vertikální přechody slanosti, světla, teploty a živin. Chemie mořského ledu je nejvíce ovlivněna slaností solanky, která ovlivňuje pH a koncentrace rozpuštěných živin a plynů. The solanka vytvořený během tání mořského ledu vytváří póry a kanály v mořském ledu, ve kterých mohou tyto mikroby žít. V důsledku těchto gradientů a dynamických podmínek se ve spodní vrstvě ledu nachází vyšší množství mikrobů, i když některé se nacházejí ve střední a horní vrstvě.[254]
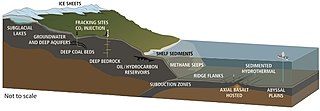
Hydrotermální průduchy jsou umístěny tam, kde tektonické desky se oddělují a šíří. To umožňuje vodě z oceánu vstoupit do zemské kůry, kde je zahříváno magmatem. Zvyšující se tlak a teplota tlačí vodu zpět z těchto otvorů, na cestě voda hromadí rozpuštěné minerály a chemikálie z hornin, se kterými se setkává. Průduchy lze charakterizovat teplotou a chemickým složením jako difuzní průduchy které uvolňují čistou relativně chladnou vodu obvykle pod 30 ° C, jako bílí kuřáci které emitují mléčně zbarvenou vodu při teplejších teplotách, asi 200 - 330 ° C a podobně černí kuřáci které emitují vodu ztemnělou nahromaděnými sraženinami sirníku při vysokých teplotách, asi 300-400 ° C.[255]
Hydrotermální ventilační mikrobiální společenství jsou mikroskopické jednobuněčné organismy, které žijí a množí se v chemicky odlišné oblasti kolem hydrotermálních průduchů. Patří mezi ně organismy v mikrobiální rohože, volné plovoucí buňky a bakterie v endosymbiotikum vztahy se zvířaty. Protože v těchto hloubkách není sluneční světlo, dodává energii chemosyntéza kde symbiotické bakterie a archea tvoří dno potravinového řetězce a jsou schopny podporovat různé organismy, jako je obří trubkoví červi a Pompejské červy. Tyto organismy využívají tento symbiotický vztah k využívání a získávání chemické energie, která se uvolňuje v těchto hydrotermálních ventilačních oblastech.[256] Chemolithoautotrofní bakterie získávají živiny a energii z geologické aktivity v hydrotermálním otvoru, aby fixovaly uhlík do organických forem.
Viry jsou také součástí mikrobiální komunity hydrotermálních ventilací a jejich vliv na mikrobiální ekologii v těchto ekosystémech je rostoucí oblastí výzkumu.[257] Viry jsou nejhojnějším životem v oceánu a skrývají největší rezervoár genetické rozmanitosti.[258] Protože jejich infekce jsou často smrtelné, představují významný zdroj úmrtnosti, a mají tak rozsáhlý vliv na biologické oceánografické procesy, vývoj a biogeochemické cyklování v oceánu.[259] Byly však nalezeny důkazy o tom, že viry nalezené ve větracích stanovištích přijaly více mutualistic než parazitický evoluční strategie za účelem přežití extrémního a nestálého prostředí, ve kterém existují.[260] Bylo zjištěno, že hlubinné hydrotermální průduchy mají vysoký počet virů, což naznačuje vysokou produkci virů.[261] Stejně jako v jiných mořských prostředích hlubinné hydrotermální viry ovlivnit hojnost a rozmanitost prokaryoty a proto ovlivňují mikrobiální biogeochemické cyklování o lyžování jejich hostitelé k replikaci.[262] Na rozdíl od jejich role jako zdroje úmrtnosti a kontroly populace se však předpokládá, že viry zlepšují přežití prokaryot v extrémních prostředích a fungují jako rezervoáry genetické informace. Interakce virosféry s mikroorganismy pod tlakem prostředí se proto považují za pomoc při přežití mikroorganismů prostřednictvím rozptýlení hostitelských genů prostřednictvím horizontální přenos genů.[263]
Hluboká biosféra a temná hmota

The hluboká biosféra je ta část biosféra který se nachází pod několika prvními metry povrchu. Rozkládá se nejméně 5 kilometrů pod kontinentálním povrchem a 10,5 kilometru pod mořským povrchem s teplotami, které mohou překročit 100 ° C.
Živé organismy nad povrchem konzumují organickou hmotu a kyslík. Níže nejsou k dispozici, takže využívají „poživatiny“ (dárci elektronů ), jako je vodík uvolňovaný z hornin různými chemickými procesy, metan, sloučeniny se sníženým obsahem síry a amonium. „Dýchají“ akceptory elektronů jako jsou dusičnany a dusitany, oxidy manganu a železa, oxidované sloučeniny síry a oxid uhličitý.
Ve větších hloubkách je velmi málo energie a metabolismus může být až milionkrát pomalejší než na povrchu. Buňky mohou žít tisíce let před rozdělením a není znám žádný limit jejich věku. Podpovrch tvoří asi 90% biomasa v bakteriích a archaeách a 15% z celkové biomasy pro biosféru. Rovněž se nalézají eukaryoty, většinou mikroskopické, ale zahrnující vícebuněčný život. Viry jsou také přítomny a infikují mikroby.
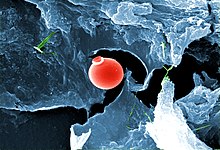
V roce 2018 vědci z Deep Carbon Observatory oznámil to formy života, včetně 70% bakterií a archea na Zemi, v celkovém objemu biomasy 23 miliard tun uhlík, dostát 4,8 km (3,0 mil) hluboko v podzemí, včetně 2,5 km (1,6 mil) pod mořským dnem.[264][265][266] V roce 2019 byly objeveny mikrobiální organismy žijící 7 900 stop (2400 m) pod povrchem, dýchající síru a jíst kameny jako např pyrit jako jejich pravidelný zdroj potravy.[267][268][269] K tomuto objevu došlo v nejstarší známé vodě na Zemi.[270]
V roce 2020 vědci uvedli, že našli nejdelší formy života, jaké kdy existovaly: aerobní mikroorganismy, které byly v kvazi pozastavená animace až 101,5 milionu let. Mikroorganismy byly nalezeny v organicky chudé sedimenty 68,9 metrů (226 stop) pod mořské dno v Jižní Pacifik Gyre (SPG), „nejsmrtelnější místo v oceánu“.[271][272]
Biologové to dosud nedokázali kultura v laboratoři drtivá většina mikroorganismů. To platí zejména pro bakterie a archea a je to způsobeno nedostatkem znalostí nebo schopností zajistit požadované růstové podmínky.[273][274] Termín mikrobiální temná hmota se začalo používat k popisu mikroorganismů, o kterých vědci vědí, že tam jsou, ale nebyli schopni kultivovat, a jejichž vlastnosti proto zůstávají nepolapitelné.[273] Mikrobiální temná hmota nesouvisí s temná hmota fyziky a kosmologie, ale je to tzv. obtížnost efektivního studia. Je těžké odhadnout jeho relativní velikost, ale akceptovaný hrubý odhad je, že méně než jedno procento mikrobiálních druhů v daném ekologický výklenek je kultivovatelný. V posledních letech se vyvíjí snaha dešifrovat více mikrobiální temné hmoty pomocí učení jejich genom DNA sekvence ze vzorků prostředí[275] a poté získáním poznatků o jejich metabolismu z jejich sekvenovaného genomu a podporou znalostí potřebných pro jejich kultivaci.
Mikrobiální rozmanitost


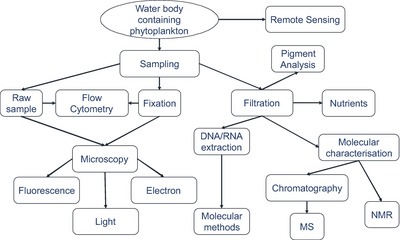
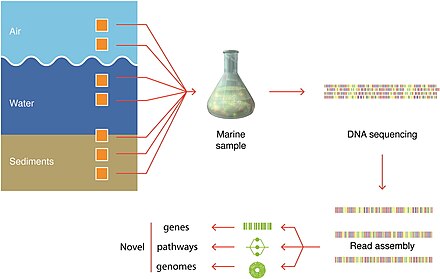
Techniky odběru vzorků

Modré pozadí označuje filtrovaný objem potřebný k získání dostatečného počtu organismů pro analýzu.
Skutečné objemy, ze kterých se odebírají vzorky organismů, se vždy zaznamenávají.[279]
Identifikace mikroorganismů

pomocí moderních zobrazovacích technik.[279]

Tradičně fylogeneze mikroorganismů bylo odvozeno a jejich taxonomie byla založena na základě studií o morfologie. Vývoj v roce 2006 molekulární fylogenetika umožnily stanovit evoluční vztah druhů analýzou hlubších charakteristik, jako jsou jejich DNA a protein sekvence, například ribozomální DNA.[280] Nedostatek snadno dostupných morfologických znaků, jako jsou ty, které jsou přítomny v zvířata a rostliny, zvláště brání časnému úsilí při klasifikaci bakterií a archea. To vedlo k chybné, zkreslené a zmatené klasifikaci, jejíž příklad je uveden Carl Woese, je Pseudomonas jehož etymologie ironicky odpovídala jeho taxonomii, konkrétně „falešné jednotce“.[281] Mnoho bakteriálních taxonů bylo překlasifikováno nebo předefinováno pomocí molekulární fylogenetiky.
Poslední vývoj v roce 2006 molekulární sekvenování umožnily navrácení genomy in situ, přímo ze vzorků životního prostředí a vyhýbání se nutnosti kultivace. To vedlo například k rychlému rozšíření znalostí o rozmanitosti bakteriální kmen. Tyto techniky jsou vyřešeny genomem metagenomika a jednobuněčná genomika.



| Externí video | |
|---|---|
Metody identifikace mikroorganismů [284] | |||
|---|---|---|---|
| Chromogenní média | Mikroskopické techniky | Biochemické techniky | Molekulární techniky |
Tradiční média | Světlé pole Tmavé pole SEM TEM CLSM bankomat Invertovaná mikroskopie | Spektrometrie – FTIR – Ramanova spektrometrie Hmotnostní spektrometrie – GC – LC – MALDI-TOF – ESI Elektrokinetická separace Mikrofluidní čip Metody slušnosti - Wickerhamova karta – API - Křišťál BBL - Vitek - Biolog | PCR QPCR v reálném čase Rychlá PCR Sekvenování PCR RFLP PFGE Ribotyping WGS MALDI-TOF MS  |
Nové technologie sekvenování a hromadění sekvenčních dat vedly k posunu paradigmatu, který zdůraznil jak všudypřítomnost mikrobiálních společenství ve spojení s vyššími organismy, tak kritickou roli mikrobů ve zdraví ekosystému.[285] Tyto nové možnosti přinesly revoluci mikrobiální ekologie, protože analýza genomů a metagenomů vysoce výkonným způsobem poskytuje účinné metody řešení funkčního potenciálu jednotlivých mikroorganismů i celých komunit v jejich přirozeném prostředí.[286][287][288]
Používání omických dat
Omics je termín používaný neformálně k označení poboček biologie jejichž jména končí příponou -omika, jako genomika, proteomika, metabolomika, a glykomika. Cílem Omics je kolektivně charakterizovat a kvantifikovat zásoby biologických molekul, které se promítají do struktury, funkce a dynamiky organismu nebo organismů. Například, funkční genomika si klade za cíl identifikovat funkce co největšího počtu genů daného organismu. Kombinuje různé-omické techniky jako jsou transkriptomika a proteomika s nasycenými mutant sbírky.[290][291]
Mnoho věcí nad rámec originálu genom se staly užitečnými a v posledních letech byly široce přijaty výzkumnými vědci. Přípona -omika může poskytnout snadnou zkratku pro zapouzdření pole; například an interaktomika studie je rozumně rozeznatelná v souvislosti s rozsáhlými analýzami interakcí gen-gen, protein-protein nebo protein-ligand, zatímco proteomika se stal pojmem pro studium bílkoviny ve velkém měřítku.
Jakákoli daná technika omiky, použitá sama o sobě, nemůže dostatečně rozmotat složitost hostitelského mikrobiomu. K uspokojivému rozluštění složitosti interakcí hostitel-mikrobiom jsou zapotřebí přístupy multi-„komiksové“.[292] Například, metagenomika, metatranscriptomics, metaproteomika a metabolomika všechny metody se používají k poskytování informací o metagenom.[293]

Vidět...
- Brüwer, J.D. a Buck-Wiese, H. (2018) „Čtení knihy života - Omics jako univerzální nástroj napříč obory“. V: YOUMARES 8 - Oceans Across Boundaries: Learning from each other, strany 73–82. Springer. ISBN 9783319932842.
Antropogenní dopady

V mořském prostředí, mikrobiální prvovýroba podstatně přispívá k CO2 sekvestrace. Mořské mikroorganismy také recyklují živiny pro použití v web mořských potravin a v procesu uvolnění CO2 do atmosféry. Mikrobiální biomasa a další organické látky (zbytky rostlin a živočichů) se v průběhu milionů let přeměňují na fosilní paliva. Naproti tomu spalování fosilní paliva osvobozuje skleníkové plyny za malý zlomek času. V důsledku toho uhlíkový cyklus je nevyvážený a atmosférický CO2 úrovně budou nadále stoupat, dokud budou fosilní paliva nadále spalována.[6]
Viz také
- mořský život
- Hluboké maximum chlorofylu
- Havajské oceánské časové řady
- Mezinárodní sčítání mořských mikrobů
- Mořská mikrobiální symbióza
- Mikrobiální biogeografie
- Mikrobiální společenství
- Mikrobiální ekologie
- Mikrobiální potravinový web
- Mikrobiální smyčka
- Mikrobiální oxidace síry
- Mikrobiálně indukovaná sedimentární struktura
- Mikrobiologie minimálních zón kyslíku
- Oceánský uhlíkový cyklus
- Ooid
- Picoeukaryote
- Roseobacter
Reference
- ^ A b Madigan M; Martinko J, vyd. (2006). Brockova biologie mikroorganismů (13. vydání). Pearson Education. p. 1096. ISBN 978-0-321-73551-5.
- ^ A b Rybicki EP (1990). "Klasifikace organismů na pokraji života nebo problémy se systematikou virů". Jihoafrický žurnál vědy. 86: 182–6. ISSN 0038-2353.
- ^ A b Lwoff A (1956). „Koncept viru“. Journal of General Microbiology. 17 (2): 239–53. doi:10.1099/00221287-17-2-239. PMID 13481308.
- ^ A b C Bar-On, YM; Phillips, R; Milo, R (2018). „Distribuce biomasy na Zemi“. PNAS. 115 (25): 6506–6511. doi:10.1073 / pnas.1711842115. PMC 6016768. PMID 29784790.
- ^ Sčítání mořského života Zpřístupněno 29. října 2020.
- ^ A b C Cavicchioli, Ricardo; Ripple, William J .; Timmis, Kenneth N .; Azam, Farooq; Bakken, Lars R .; Baylis, Matthew; Behrenfeld, Michael J .; Boetius, Antje; Boyd, Philip W .; Classen, Aimée T .; Crowther, Thomas W .; Danovaro, Roberto; Foreman, Christine M .; Huisman, Jef; Hutchins, David A .; Jansson, Janet K .; Karl, David M .; Koskella, Britt; Mark Welch, David B .; Martiny, Jennifer B. H .; Moran, Mary Ann; Orphan, Victoria J .; Reay, David S .; Remais, Justin V .; Rich, Virginie I .; Singh, Brajesh K .; Stein, Lisa Y .; Stewart, Frank J .; Sullivan, Matthew B .; et al. (2019). „Varování vědců pro lidstvo: mikroorganismy a změna klimatu“. Příroda Recenze Mikrobiologie. 17 (9): 569–586. doi:10.1038 / s41579-019-0222-5. PMC 7136171. PMID 31213707.
 Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0. - ^ Bolhuis, H. a Cretoiu, M.S. (2016) „Co je na mořských mikroorganismech tak zvláštního?“. In: L. J. Stal a M. S. Cretoiu (Eds.) Mořský mikrobiom, strany 3–20, Springer. ISBN 9783319330006
- ^ „Funkce globálního oceánského mikrobiomu klíč k pochopení změn prostředí“. www.sciencedaily.com. University of Georgia. 10. prosince 2015. Citováno 11. prosince 2015.
- ^ A b Údaje o úmrtnosti WHO z roku 2002 Přístupné 20. ledna 2007
- ^ Wade, Nicholas (25. července 2016). „Seznamte se s Lucem, předkem všeho živého“. New York Times. Citováno 25. července 2016.
- ^ Suttle, C.A. (2005). "Viry v moři". Příroda. 437 (9): 356–361. Bibcode:2005 Natur.437..356S. doi:10.1038 / nature04160. PMID 16163346. S2CID 4370363.
- ^ A b Glöckner, F.O., Gasol, J.M., McDonough, N., Calewaert, J.B. a kol. (2012) Mořská mikrobiální rozmanitost a její role ve fungování ekosystému a při změně prostředí. Evropská vědecká nadace, Poziční papír 17. ISBN 978-2-918428-71-8
- ^ „Funkce globálního oceánského mikrobiomu klíč k pochopení změn prostředí“. www.sciencedaily.com. University of Georgia. 10. prosince 2015. Citováno 11. prosince 2015.
- ^ Suttle, C.A. (2005). "Viry v moři". Příroda. 437 (9): 356–361. Bibcode:2005 Natur.437..356S. doi:10.1038 / nature04160. PMID 16163346. S2CID 4370363.
- ^ Shors p. 5
- ^ Shors p. 593
- ^ A b C Suttle CA. Mořské viry - hlavní hráči v globálním ekosystému. Příroda Recenze Mikrobiologie. 2007;5(10):801–12. doi:10.1038 / nrmicro1750. PMID 17853907.
- ^ Živé bakterie jezdí na vzdušných proudech Země Smithsonian Magazine, 11. ledna 2016.
- ^ Robbins, Jim (13. dubna 2018). „Biliony za biliony virů každý den padají z nebe“. The New York Times. Citováno 14. dubna 2018.
- ^ Reche, Isabel; D’Orta, Gaetano; Mladenov, Natalie; Winget, Danielle M; Suttle, Curtis A (29. ledna 2018). „Míra ukládání virů a bakterií nad mezní vrstvou atmosféry“. ISME Journal. 12 (4): 1154–1162. doi:10.1038 / s41396-017-0042-4. PMC 5864199. PMID 29379178.
- ^ Zaměstnanci (2014). „Biosféra“. Aspen Global Change Institute. Citováno 10. listopadu 2014.
- ^ A b C Choi, Charles Q. (17. března 2013). „Mikrobům se daří v nejhlubším místě na Zemi“. LiveScience. Citováno 17. března 2013.
- ^ Glud, Ronnie; Wenzhöfer, Frank; Middelboe, Mathias; Oguri, Kazumasa; Turnewitsch, Robert; Canfield, Donald E .; Kitazato, Hiroshi (17. března 2013). „Vysoká rychlost obratu mikrobiálního uhlíku v sedimentech v nejhlubším oceánském příkopu na Zemi“. Nature Geoscience. 6 (4): 284–288. Bibcode:2013NatGe ... 6..284G. doi:10.1038 / ngeo1773.
- ^ Oskin, Becky (14. března 2013). „Intraterrestrials: Life Thrives in Ocean Floor“. LiveScience. Citováno 17. března 2013.
- ^ Morelle, Rebecca (15. prosince 2014). "Analyzovány mikroby objevené nejhlubším námořním cvičením". BBC novinky. Citováno 15. prosince 2014.
- ^ Takai K; Nakamura K; Toki T; Tsunogai U; et al. (2008). „Proliferace buněk při 122 ° C a izotopově těžká produkce CH4 hypertermofilním methanogenem při vysokotlaké kultivaci“. Sborník Národní akademie věd Spojených států amerických. 105 (31): 10949–54. Bibcode:2008PNAS..10510949T. doi:10.1073 / pnas.0712334105. PMC 2490668. PMID 18664583.
- ^ Fox, Douglas (20. srpna 2014). „Jezera pod ledem: tajná zahrada Antarktidy“. Příroda. 512 (7514): 244–246. Bibcode:2014 Natur.512..244F. doi:10.1038 / 512244a. PMID 25143097.
- ^ Mack, Eric (20. srpna 2014). „Život potvrzen pod antarktickým ledem; je vesmír další?“. Forbes. Citováno 21. srpna 2014.
- ^ Armbrust, E.V .; Palumbi, S.R. (2015). „Odhalení skrytých světů biologické rozmanitosti oceánů“. Věda. 348 (6237): 865–867. doi:10.1126 / science.aaa7378. PMID 25999494. S2CID 36480105.
- ^ Azam, F .; Malfatti, F. (2007). „Mikrobiální struktura mořských ekosystémů“. Příroda Recenze Mikrobiologie. 5 (10): 782–791. doi:10.1038 / nrmicro1747. PMID 17853906. S2CID 10055219.
- ^ Middelboe, M .; Brussaard, C. (2017). „Mořské viry: klíčoví hráči v mořských ekosystémech“. Viry. 9 (10): 302. doi:10,3390 / v9100302. PMC 5691653. PMID 29057790.
- ^ Koonin EV, Senkevich TG, Dolja VV. Starověký virový svět a vývoj buněk. Biology Direct. 2006;1:29. doi:10.1186/1745-6150-1-29. PMID 16984643.
- ^ A b Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H. Fág jako agenti laterálního přenosu genů. Současný názor v mikrobiologii. 2003;6(4):417–24. doi:10.1016 / S1369-5274 (03) 00086-9. PMID 12941415.
- ^ A b Rybicki, EP. Klasifikace organismů na pokraji života nebo problémy se systematikou virů. Jihoafrický žurnál vědy. 1990;86:182–186.
- ^ Koonin, E. V .; Starokadomskyy, P. (říjen 2016). „Jsou viry naživu? Paradigma replikátoru vrhá rozhodující světlo na starou, ale zavádějící otázku“. Studie historie a filozofie vědy Část C: Studie historie a filozofie biologických a biomedicínských věd. 59: 125–134. doi:10.1016 / j.shpsc.2016.02.016. PMC 5406846. PMID 26965225.
- ^ Iyer LM, Balaji S, Koonin EV, Aravind L. Evoluční genomika nukleo-cytoplazmatických velkých DNA virů. Virový výzkum. 2006;117(1):156–84. doi:10.1016 / j.virusres.2006.01.009. PMID 16494962.
- ^ Sanjuán R, Nebot MR, Chirico N, Mansky LM, Belshaw R. Míry virových mutací. Journal of Virology. 2010;84(19):9733–48. doi:10.1128 / JVI.00694-10. PMID 20660197.
- ^ Mahy WJ & Van Regenmortel MHV (eds). Desk Encyclopedia of General Virology. Oxford: Academic Press; 2009. ISBN 0-12-375146-2. p. 28.
- ^ Jsou viry naživu? Paradigma replikátoru vrhá rozhodující světlo na starou, ale zavádějící otázku. Studie z historie a filozofie biologických a biomedicínských věd. 2016. doi:10.1016 / j.shpsc.2016.02.016. PMID 26965225.
- ^ Koonin, E. V .; Starokadomskyy, P. (7. března 2016). „Jsou viry naživu? Paradigma replikátoru vrhá rozhodující světlo na starou, ale zavádějící otázku“. Studie z historie a filozofie biologických a biomedicínských věd. 59: 125–34. doi:10.1016 / j.shpsc.2016.02.016. PMC 5406846. PMID 26965225.
- ^ A b Mann, NH (17. května 2005). „Třetí věk fága“. PLOS Biology. 3 (5): 753–755. doi:10.1371 / journal.pbio.0030182. PMC 1110918. PMID 15884981.
- ^ Wommack KE, Colwell RR. Virioplankton: viry ve vodních ekosystémech. Recenze mikrobiologie a molekulární biologie. 2000;64(1):69–114. doi:10.1128 / MMBR.64.1.69-114.2000. PMID 10704475.
- ^ Suttle CA. Viry v moři. Příroda. 2005;437:356–361. doi:10.1038 / nature04160. PMID 16163346. Bibcode:2005 Natur.437..356S.
- ^ Bergh O, Børsheim KY, Bratbak G, Heldal M. Vysoký výskyt virů ve vodním prostředí. Příroda. 1989;340(6233):467–8. doi:10.1038 / 340467a0. PMID 2755508. Bibcode:1989 Natur.340..467B.
- ^ Wigington CH, Sonderegger D, Brussaard CPD, Buchan A, Finke JF, Fuhrman JA, Lennon JT, Middelboe M, Suttle CA, Stock C, Wilson WH, Wommack KE, Wilhelm SW, Weitz JS. Znovu přezkoumání vztahu mezi mořským virem a množstvím mikrobiálních buněk. Přírodní mikrobiologie. 2016;1:15024. doi:10.1038 / nmicrobiol.2015.24. PMID 27572161.
- ^ Brum JR, Schenck RO, Sullivan MB (září 2013). „Globální morfologická analýza mořských virů ukazuje minimální regionální variace a dominanci bezocasých virů“. Časopis ISME. 7 (9): 1738–51. doi:10.1038 / ismej.2013.67. PMC 3749506. PMID 23635867.
- ^ Krupovic M, Bamford DH (2007). „Předpokládané profágy související s lytickým bezocasým mořským dsDNA fágem PM2 jsou rozšířené v genomech vodních bakterií“. BMC Genomics. 8: 236. doi:10.1186/1471-2164-8-236. PMC 1950889. PMID 17634101.
- ^ Xue H, Xu Y, Boucher Y, Polz MF (2012). „Vysoká frekvence nového filamentózního fága, VCY φ, v populaci prostředí Vibrio cholerae“. Aplikovaná a environmentální mikrobiologie. 78 (1): 28–33. doi:10.1128 / AEM.06297-11. PMC 3255608. PMID 22020507.
- ^ Roux S, Krupovic M, Poulet A, Debroas D, Enault F (2012). „Evoluce a rozmanitost virové rodiny Microviridae prostřednictvím sbírky 81 nových kompletních genomů shromážděných z čtení viromu“. PLOS ONE. 7 (7): e40418. Bibcode:2012PLoSO ... 740418R. doi:10.1371 / journal.pone.0040418. PMC 3394797. PMID 22808158.
- ^ Kauffman, Kathryn M .; Hussain, Fatima A .; Yang, Joy; Arevalo, Philip; Brown, Julia M .; Chang, William K .; VanInsberghe, David; Elsherbini, Joseph; Sharma, Radhey S .; Cutler, Michael B .; Kelly, Libusha; Polz, Martin F. (2018). „Hlavní linie neosoustředěných dsDNA virů jako nerozpoznaných zabijáků mořských bakterií“. Příroda. 554 (7690): 118–122. Bibcode:2018Natur.554..118K. doi:10.1038 / příroda25474. PMID 29364876. S2CID 4462007.
- ^ Vědci našli nový typ viru ve světových oceánech: Autolykiviridae, dne: sci-news, 25. ledna 2018
- ^ V oceánu byly právě objeveny nikdy předtím neviděné viry s podivnou DNA, na: vědavýstraha, 25. ledna 2018
- ^ NCBI: Autolykiviridae (rodina) - nezařazené dsDNA viry
- ^ Lawrence CM, Menon S, Eilers BJ, et al.. Strukturální a funkční studie archaálních virů. Journal of Biological Chemistry. 2009;284(19):12599–603. doi:10,1074 / jbc.R800078200. PMID 19158076.
- ^ Prangishvili D, Forterre P, Garrett RA. Viry Archea: sjednocující pohled. Příroda Recenze Mikrobiologie. 2006;4(11):837–48. doi:10.1038 / nrmicro1527. PMID 17041631.
- ^ Prangishvili D, Garrett RA. Výjimečně rozmanité morfotypy a genomy crenarcheálních hypertermofilních virů. Transakce s biochemickou společností. 2004; 32 (Pt 2): 204–8. doi:10.1042 / BST0320204. PMID 15046572.
- ^ Suttle CA. Viry v moři. Příroda. 2005;437(7057):356–61. doi:10.1038 / nature04160. PMID 16163346. Bibcode:2005 Natur.437..356S.
- ^ www.cdc.gov. Škodlivé květy řas: Červený příliv: Domů [Citováno 2014-12-19].
- ^ Forterre P, Philippe H. Poslední univerzální společný předek (LUCA), jednoduchý nebo komplexní ?. Biologický bulletin. 1999; 196 (3): 373–5; diskuse 375–7. doi:10.2307/1542973. PMID 11536914.
- ^ Abrahão, Jônatas; Silva, Lorena; Silva, Ludmila Santos; Khalil, Jacques Yaacoub Bou; Rodrigues, Rodrigo; Arantes, Thalita; Assis, Felipe; Boratto, Paulo; Andrade, Miguel; Kroon, Erna Geessien; Ribeiro, Bergmann; Bergier, Ivan; Seligmann, Herve; Ghigo, Eric; Colson, Philippe; Levasseur, Anthony; Kroemer, Guido; Raoult, Didier; Scola, Bernard La (27. února 2018). „Tailanvirus obrovský má nejkompletnější translační aparát známé virosféry“. Příroda komunikace. 9 (1): 749. Bibcode:2018NatCo ... 9..749A. doi:10.1038 / s41467-018-03168-1. PMC 5829246. PMID 29487281.
- ^ Bichell RE. „V genech obrovských virů naráží na jejich tajemný původ“. Konec konců.
- ^ Van Etten JL (červenec – srpen 2011). „Obří viry“. Americký vědec. 99 (4): 304–311. doi:10.1511/2011.91.304.
- ^ Legendre M, Arslan D, Abergel C, Claverie JM (leden 2012). „Genomika megaviru a nepolapitelná čtvrtá doména života“. Komunikativní a integrativní biologie. 5 (1): 102–6. doi:10,4161 / cib.18624. PMC 3291303. PMID 22482024.
- ^ Schulz F, Yutin N, Ivanova NN, Ortega DR, Lee TK, Vierheilig J, Daims H, Horn M, Wagner M, Jensen GJ, Kyrpides NC, Koonin EV, Woyke T (duben 2017). „Obří viry s rozšířeným doplňkem komponent překladového systému“ (PDF). Věda. 356 (6333): 82–85. Bibcode:2017Sci ... 356 ... 82S. doi:10.1126 / science.aal4657. PMID 28386012. S2CID 206655792.
- ^ Bäckström D, Yutin N, Jørgensen SL, Dharamshi J, Homa F, Zaremba-Niedwiedzka K, Spang A, Wolf YI, Koonin EV, Ettema TJ (březen 2019). „Virové geny z hlubinných sedimentů rozšiřují oceánský megavrom a podporují nezávislé počátky virového gigantismu“. mBio. 10 (2): e02497-02418. doi:10,1 128 / mBio.02497-18. PMC 6401483. PMID 30837339.
- ^ Fredrickson JK, Zachara JM, Balkwill DL, Kennedy D, Li SM, Kostandarithes HM, Daly MJ, Romine MF, Brockman FJ (2004). „Geomikrobiologie vysoce vadózních sedimentů kontaminovaných jaderným odpadem v lokalitě Hanford ve státě Washington“. Aplikovaná a environmentální mikrobiologie. 70 (7): 4230–41. doi:10.1128 / AEM.70.7.4230-4241.2004. PMC 444790. PMID 15240306.
- ^ Woese CR, Kandler O, Wheelis ML (1990). „Směrem k přirozenému systému organismů: návrh domén Archaea, Bacteria a Eucarya“. Sborník Národní akademie věd Spojených států amerických. 87 (12): 4576–9. Bibcode:1990PNAS ... 87,4576 W.. doi:10.1073 / pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ Schopf JW (1994). „Nesourodé sazby, různé osudy: tempo a způsob vývoje se změnily z prekambria na phanerozoik“. Sborník Národní akademie věd Spojených států amerických. 91 (15): 6735–42. Bibcode:1994PNAS ... 91.6735S. doi:10.1073 / pnas.91.15.6735. PMC 44277. PMID 8041691.
- ^ DeLong EF, Pace NR (2001). "Environmentální rozmanitost bakterií a archea". Systematická biologie. 50 (4): 470–8. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Brown JR, Doolittle WF (1997). „Archaea a přechod mezi prokaryotem a eukaryotem“. Recenze mikrobiologie a molekulární biologie. 61 (4): 456–502. doi:10.1128/.61.4.456-502.1997. PMC 232621. PMID 9409149.
- ^ Dyall, Sabrina D .; Brown, Mark T .; Johnson, Patricia J. (9. dubna 2004). „Starověké invaze: Od endosymbiontů po organely“. Věda. 304 (5668): 253–257. Bibcode:2004Sci ... 304..253D. doi:10.1126 / science.1094884. ISSN 0036-8075. PMID 15073369. S2CID 19424594.
- ^ Poole AM, Penny D (2007). „Hodnocení hypotéz o původu eukaryot“. BioEssays. 29 (1): 74–84. doi:10.1002 / bies.20516. PMID 17187354. S2CID 36026766.
- ^ Lang BF, Gray MW, Burger G (1999). "Vývoj mitochondriálního genomu a původ eukaryot". Výroční přehled genetiky. 33: 351–97. doi:10.1146 / annurev.genet.33.1.351. PMID 10690412.
- ^ McFadden GI (1999). "Endosymbióza a vývoj rostlinné buňky". Aktuální názor na biologii rostlin. 2 (6): 513–9. doi:10.1016 / S1369-5266 (99) 00025-4. PMID 10607659.
- ^ Patrick J. Keeling (2004). „Rozmanitost a evoluční historie plastidů a jejich hostitelů“. American Journal of Botany. 91 (10): 1481–1493. doi:10.3732 / ajb.91.10.1481. PMID 21652304.
- ^ „Candidatus Pelagibacter Ubique.“ Evropský bioinformatický institut. Evropský bioinformatický institut, 2011. Web. 8. ledna 2012. http://www.ebi.ac.uk/2can/genomes/bacteria/Candidatus_Pelagibacter_ubique.html Archivováno 1. prosince 2008, na Wayback Machine
- ^ „Bleší trh: Nově objevený virus může být nejhojnějším organismem na planetě“. Ekonom. 16. února 2013. Citováno 16. února 2013.
- ^ Zhao, Y .; Temperton, B .; Thrash, J. C .; Schwalbach, M. S .; Vergin, K. L .; Landry, Z. C .; Ellisman, M .; Deerinck, T .; Sullivan, M. B .; Giovannoni, S. J. (2013). "Hojné viry SAR11 v oceánu". Příroda. 494 (7437): 357–360. Bibcode:2013Natur.494..357Z. doi:10.1038 / příroda1121. PMID 23407494. S2CID 4348619.
- ^ „Největší bakterie: Vědec objevuje novou formu bakteriálního života u afrického pobřeží“, Max Planck Institute for Marine Microbiology, 8. dubna 1999, archivovány od originál dne 20. ledna 2010
- ^ Seznam prokaryotických jmen se Stálým v nomenklatuře - rod Thiomargarita
- ^ Bang C, Schmitz RA (2015). „Archaea spojená s lidským povrchem: nelze podceňovat“. Recenze mikrobiologie FEMS. 39 (5): 631–48. doi:10.1093 / femsre / fuv010. PMID 25907112.
- ^ Archaea Online slovník etymologie. Vyvolány 17 August je 2016.
- ^ Tempo NR (květen 2006). "Čas na změnu". Příroda. 441 (7091): 289. Bibcode:2006 Natur.441..289P. doi:10.1038 / 441289a. PMID 16710401. S2CID 4431143.
- ^ Stoeckenius W (1. října 1981). „Walsbyho čtvercová bakterie: jemná struktura ortogonálního prokaryota“. Journal of Bacteriology. 148 (1): 352–60. doi:10.1128 / JB.148.1.352-360.1981. PMC 216199. PMID 7287626.
- ^ Madigan M. a Martinko J. (ed.) (2005). Brockova biologie mikroorganismů (11. vydání). Prentice Hall. ISBN 978-0-13-144329-7.CS1 maint: další text: seznam autorů (odkaz)
- ^ "Hlubinné mikroorganismy a původ eukaryotické buňky" (PDF). Citováno 24. října 2017.
- ^ Yamaguchi, Masashi; et al. (1. prosince 2012). „Prokaryot nebo eukaryot? Unikátní mikroorganismus z hlubin moře“. Journal of Electron Microscopy. 61 (6): 423–431. doi:10.1093 / jmicro / dfs062. PMID 23024290.
- ^ Whittaker, R.H .; Margulis, L. (1978). "Protist klasifikace a království organismů". Biosystémy. 10 (1–2): 3–18. doi:10.1016/0303-2647(78)90023-0. PMID 418827.
- ^ Faure, E; Ne, F; Benoiston, AS; Labadie, K; Bittner, L; Ayata, SD (2019). „Mixotrofní protisté zobrazují kontrastní biogeografie v globálním oceánu“. ISME Journal. 13 (4): 1072–1083. doi:10.1038 / s41396-018-0340-5. PMC 6461780. PMID 30643201.
- ^ A b C Leles, S.G .; Mitra, A .; Flynn, K.J .; Stoecker, D.K .; Hansen, P.J .; Calbet, A .; McManus, G.B .; Sanders, R.W .; Caron, D. A.; Ne, F .; Hallegraeff, G.M. (2017). „Oceánští protisté s různými formami získané fototrofie vykazují kontrastní biogeografie a hojnost“. Sborník Královské společnosti B: Biologické vědy. 284 (1860): 20170664. doi:10.1098 / rspb.2017.0664. PMC 5563798. PMID 28768886.
- ^ Budd, Graham E; Jensen, Sören (2017). „Původ zvířat a hypotéza„ Savannah “pro časnou bilaterální evoluci“. Biologické recenze. 92 (1): 446–473. doi:10.1111 / brv.12239. PMID 26588818.
- ^ Cavalier-Smith T (prosinec 1993). „Prvoky království a jejich 18 kmenů“. Mikrobiologické recenze. 57 (4): 953–94. doi:10.1128 / mmbr. 57.4.953-994.1993. PMC 372943. PMID 8302218.
- ^ Corliss JO (1992). „Měl by pro protisty existovat samostatný kód nomenklatury?“. BioSystems. 28 (1–3): 1–14. doi:10.1016 / 0303-2647 (92) 90003-H. PMID 1292654.
- ^ Slapeta J, Moreira D, López-García P (2005). „Rozsah protistické rozmanitosti: poznatky z molekulární ekologie sladkovodních eukaryot“. Sborník Královské společnosti B: Biologické vědy. 272 (1576): 2073–81. doi:10.1098 / rspb.2005.3195. PMC 1559898. PMID 16191619.
- ^ Moreira D, López-García P (2002). „Molekulární ekologie mikrobiálních eukaryot odhaluje skrytý svět“ (PDF). Trendy v mikrobiologii. 10 (1): 31–8. doi:10.1016 / S0966-842X (01) 02257-0. PMID 11755083.
- ^ A b Vzduch, který dýcháte? Diatom to udělal
- ^ „Více o Diatoms“. Muzeum paleontologie University of California. Archivovány od originál dne 4. října 2012. Citováno 11. října 2019.
- ^ Devreotes P (1989). „Dictyostelium discoideum: modelový systém pro vývoj buněčných interakcí“. Věda. 245 (4922): 1054–8. Bibcode:1989Sci ... 245.1054D. doi:10.1126 / science.2672337. PMID 2672337.
- ^ Matz, Michail V .; Tamara M. Frank; Justin Marshall; Edith A. Widder; Sonke Johnsen (9. prosince 2008). „Giant Deep-Sea Protist Produces Bilaterian-like Traces“ (PDF). Aktuální biologie. Elsevier Ltd. 18 (23): 1849–1854. doi:10.1016 / j.cub.2008.10.028. PMID 19026540. S2CID 8819675.
- ^ Gooday, A. J .; Aranda da Silva, A .; Pawlowski, J. (1. prosince 2011). „Xenophyophores (Rhizaria, Foraminifera) z Nazaré Canyonu (okraj Portugalska, SV Atlantik)“. Hlubokomořský výzkum, část II: Aktuální studie v oceánografii. Geologie, geochemie a biologie podmořských kaňonů západně od Portugalska. 58 (23–24): 2401–2419. Bibcode:2011DSRII..58.2401G. doi:10.1016 / j.dsr2.2011.04.005.
- ^ Neil A C, Reece J B, Simon E J (2004) Základní biologie s fyziologií Pearson / Benjamin Cummings, strana 291. ISBN 9780805375039
- ^ O'Malley MA, Simpson AG, Roger AJ (2012). „Ostatní eukaryoty ve světle evoluční protistologie“. Biologie a filozofie. 28 (2): 299–330. doi:10.1007 / s10539-012-9354-r. S2CID 85406712.
- ^ Adl SM, Simpson AG, Farmer MA, Andersen RA, Anderson OR, Barta JR, Bowser SS, Brugerolle G, Fensome RA, Fredericq S, James TY, Karpov S, Kugrens P, Krug J, Lane CE, Lewis LA, Lodge J Lynn DH, Mann DG, McCourt RM, Mendoza L, Moestrup O, Mozley-Standridge SE, Nerad TA, Shearer CA, Smirnov AV, Spiegel FW, Taylor MF (2005). „Nová vyšší klasifikace eukaryot s důrazem na taxonomii protistů“. The Journal of Eukaryotic Microbiology. 52 (5): 399–451. doi:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- ^ Margulis L, Chapman MJ (19. března 2009). Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth. Akademický tisk. ISBN 9780080920146.
- ^ A b Dawson, Scott C; Paredez, Alexander R (2013). „Alternativní cytoskeletální krajiny: cytoskeletální novinka a vývoj v bazálních vykopávkách protistů“. Aktuální názor na buněčnou biologii. 25 (1): 134–141. doi:10.1016 / j.ceb.2012.11.005. PMC 4927265. PMID 23312067.
- ^ A b Atkinson, A .; Polimene, L .; Fileman, E.S .; Widdicombe, C.E.; McEvoy, A.J.; Smyth, T.J.; Djeghri, N.; Sailley, S.F.; Cornwell, L.E. (2018). ""Komentář. What drives plankton seasonality in a stratifying shelf sea? Some competing and complementary theories"]" (PDF). Limnologie a oceánografie. 63 (6): 2877–2884. Bibcode:2018LimOc..63.2877A. doi:10.1002/lno.11036.
- ^ Singleton, Paul (2006). Slovník mikrobiologie a molekulární biologie, 3. vydání, revidováno. Chichester, Velká Británie: John Wiley & Sons. p. 32. ISBN 978-0-470-03545-0.
- ^ David J. Patterson. „Améby: Protisté, kteří se pohybují a krmí pomocí Pseudopodia“. Webový projekt Tree of Life.
- ^ „Améby“. University of Edinburgh. Archivovány od originál dne 10. června 2009.
- ^ Brady, Richard M. (1993). "Torque and switching in the bacterial flagellar motor. An electrostatic model". Biofyzikální deník. 64 (4): 961–973. Bibcode:1993BpJ....64..961B. doi:10.1016/S0006-3495(93)81462-0. PMC 1262414. PMID 7684268.
- ^ Lauga, Eric; Thomas R Powers (25 August 2009). "The hydrodynamics of swimming microorganisms". Zprávy o pokroku ve fyzice. 72 (9): 096601. arXiv:0812.2887. Bibcode:2009RPPh...72i6601L. doi:10.1088/0034-4885/72/9/096601. S2CID 3932471.
- ^ Hyde, K.D.; E.B.J. Jones; E. Leaño; S.B. Pointing; A.D. Poonyth; L.L.P. Vrijmoed (1998). "Role of fungi in marine ecosystems". Biodiverzita a ochrana. 7 (9): 1147–1161. doi:10.1023/A:1008823515157. S2CID 22264931.
- ^ Kirk, P.M., Cannon, P.F., Minter, D.W. and Stalpers, J. "Dictionary of the Fungi". Edn 10. CABI, 2008
- ^ Hyde, K.D.; E.B.J. Jones (1989). "Spore attachment in marine fungi". Botanica Marina. 32 (3): 205–218. doi:10.1515/botm.1989.32.3.205. S2CID 84879817.
- ^ Le Calvez T, Burgaud G, Mahé S, Barbier G, Vandenkoornhuyse P (October 2009). "Fungal diversity in deep-sea hydrothermal ecosystems". Aplikovaná a environmentální mikrobiologie. 75 (20): 6415–21. doi:10.1128/AEM.00653-09. PMC 2765129. PMID 19633124.
- ^ San-Martín, A.; S. Orejanera; C. Gallardo; M. Silva; J. Becerra; R. Reinoso; M.C. Chamy; K. Vergara; J. Rovirosa (2008). "Steroids from the marine fungus Geotrichum sp". Journal of the Chilean Chemical Society. 53 (1): 1377–1378. doi:10.4067/S0717-97072008000100011.
- ^ Jones, E.B.G., Hyde, K.D., & Pang, K.-L., eds. (2014). Freshwater fungi: and fungal-like organisms. Berlín / Boston: De Gruyter.
- ^ Jones, E.B.G.; Pang, K.-L., eds. (2012). Marine Fungi, and Fungal-like Organisms. Marine and Freshwater Botany. Berlin, Boston: De Gruyter (published August 2012). doi:10.1515/9783110264067. ISBN 978-3-11-026406-7. Citováno 3. září 2015.
- ^ Wang, Xin; Singh, Purnima; Gao, Zheng; Zhang, Xiaobo; Johnson, Zackary I.; Wang, Guangyi (2014). "Distribution and Diversity of Planktonic Fungi in the West Pacific Warm Pool". PLOS ONE. 9 (7): e101523. Bibcode:2014PLoSO...9j1523W. doi:10.1371/journal.pone.0101523.s001. PMC 4081592. PMID 24992154.
- ^ Wang, G .; Wang, X .; Liu, X .; Li, Q. (2012). "Diversity and biogeochemical function of planktonic fungi in the ocean". In Raghukumar, C. (ed.). Biology of marine fungi. Berlin, Heidelberg: Springer-Verlag. pp. 71–88. doi:10.1007/978-3-642-23342-5. ISBN 978-3-642-23341-8. S2CID 39378040. Citováno 3. září 2015.
- ^ Damare, Samir; Raghukumar, Chandralata (11 November 2007). "Fungi and Macroaggregation in Deep-Sea Sediments". Mikrobiální ekologie. 56 (1): 168–177. doi:10.1007/s00248-007-9334-y. ISSN 0095-3628. PMID 17994287. S2CID 21288251.
- ^ Kubanek, Julia; Jensen, Paul R.; Keifer, Paul A.; Sullards, M. Cameron; Collins, Dwight O.; Fenical, William (10 June 2003). "Seaweed resistance to microbial attack: A targeted chemical defense against marine fungi". Sborník Národní akademie věd. 100 (12): 6916–6921. Bibcode:2003PNAS..100.6916K. doi:10.1073/pnas.1131855100. ISSN 0027-8424. PMC 165804. PMID 12756301.
- ^ A b Gao, Zheng; Johnson, Zackary I.; Wang, Guangyi (30 July 2009). "Molecular characterization of the spatial diversity and novel lineages of mycoplankton in Hawaiian coastal waters". Časopis ISME. 4 (1): 111–120. doi:10.1038/ismej.2009.87. ISSN 1751-7362. PMID 19641535.
- ^ Panzer, Katrin; Yilmaz, Pelin; Weiß, Michael; Reich, Lothar; Richter, Michael; Wiese, Jutta; Schmaljohann, Rolf; Labes, Antje; Imhoff, Johannes F. (30 July 2015). "Identification of Habitat-Specific Biomes of Aquatic Fungal Communities Using a Comprehensive Nearly Full-Length 18S rRNA Dataset Enriched with Contextual Data". PLOS ONE. 10 (7): e0134377. Bibcode:2015PLoSO..1034377P. doi:10.1371/journal.pone.0134377. PMC 4520555. PMID 26226014.
- ^ Gutierrez, Marcelo H; Pantoja, Silvio; Quinones, Renato a and Gonzalez, Rodrigo R. First record of flamentous fungi in the coastal upwelling ecosystem off central Chile. Gayana (Concepc.) [online]. 2010, vol.74, n.1, pp. 66-73. ISSN 0717-6538.
- ^ A b Sridhar, K.R. (2009). "10. Aquatic fungi – Are they planktonic?". Plankton Dynamics of Indian Waters. Jaipur, India: Pratiksha Publications. 133–148.
- ^ Species of Higher Marine Fungi Archivováno 22 April 2013 at the Wayback Machine University of Mississippi. Retrieved 2012-02-05.
- ^ Freshwater and marine lichen-forming fungi Retrieved 2012-02-06.
- ^ "Lichens". National Park Service, US Department of the Interior, Government of the United States. 22. května 2016. Citováno 4. dubna 2018.
- ^ A b "The Earth Life Web, Growth and Development in Lichens". earthlife.net. 14. února 2020.
- ^ Silliman B. R. & S. Y. Newell (2003). "Fungal farming in a snail". PNAS. 100 (26): 15643–15648. Bibcode:2003PNAS..10015643S. doi:10.1073/pnas.2535227100. PMC 307621. PMID 14657360.
- ^ Yuan X, Xiao S (2005). "Lichen-Like Symbiosis 600 Million Years Ago". Věda. 308 (5724): 1017–1020. Bibcode:2005Sci...308.1017Y. doi:10.1126/science.1111347. PMID 15890881. S2CID 27083645.
- ^ Jones, E. B. Gareth; Pang, Ka-Lai (31 August 2012). Marine Fungi: and Fungal-like Organisms. Walter de Gruyter. ISBN 9783110264067.
- ^ Janet Fang (6 April 2010). "Animals thrive without oxygen at sea bottom". Příroda. 464 (7290): 825. doi:10.1038/464825b. PMID 20376121.
- ^ "Briny deep basin may be home to animals thriving without oxygen". Vědecké zprávy. 23. září 2013.
- ^ A b Campbell, Neil A .; Reece, Jane B .; Urry, Lisa Andrea; Cain, Michael L .; Wasserman, Steven Alexander; Minorsky, Peter V .; Jackson, Robert Bradley (2008). Biologie (8 ed.). San Francisco: Pearson – Benjamin Cummings. ISBN 978-0-321-54325-7.
- ^ McNeill, J .; et al., eds. (2012). Mezinárodní kód nomenklatury pro řasy, houby a rostliny (Melbourne Code), přijatý osmnáctým mezinárodním botanickým kongresem Melbourne, Austrálie, červenec 2011 (elektronické vydání). Mezinárodní asociace pro taxonomii rostlin. Citováno 14. května 2017.
- ^ Changes in oxygen concentrations in our ocean can disrupt fundamental biological cycles Phys.org, 25 November 2019.
- ^ Walsh PJ, Smith S, Fleming L, Solo-Gabriele H, Gerwick WH, eds. (2. září 2011). "Cyanobacteria and cyanobacterial toxins". Oceans and Human Health: Risks and Remedies from the Seas. Akademický tisk. str. 271–296. ISBN 978-0-08-087782-2.
- ^ "The Rise of Oxygen - Astrobiology Magazine". Astrobiologický časopis. 30. července 2003. Citováno 6. dubna 2016.
- ^ Flannery, D. T.; R.M. Walter (2012). "Archean tufted microbial mats and the Great Oxidation Event: new insights into an ancient problem". Australian Journal of Earth Sciences. 59 (1): 1–11. Bibcode:2012AuJES..59....1F. doi:10.1080/08120099.2011.607849. S2CID 53618061.
- ^ Rothschild, Lynn (September 2003). "Understand the evolutionary mechanisms and environmental limits of life". NASA. Archivovány od originál dne 11. března 2012. Citováno 13. července 2009.
- ^ Nadis S (December 2003). "The cells that rule the seas" (PDF). Scientific American. 289 (6): 52–3. Bibcode:2003SciAm.289f..52N. doi:10.1038/scientificamerican1203-52. PMID 14631732. Archivovány od originál (PDF) dne 19. dubna 2014. Citováno 2. června 2019.
- ^ "The Most Important Microbe You've Never Heard Of". npr.org.
- ^ Flombaum, P.; Gallegos, J. L.; Gordillo, R. A.; Rincon, J.; Zabala, L. L.; Jiao, N.; Karl, D. M.; Li, W. K. W.; Lomas, M. W.; Veneziano, D.; Vera, C. S.; Vrugt, J. A.; Martiny, A. C. (2013). "Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus". Sborník Národní akademie věd. 110 (24): 9824–9829. Bibcode:2013PNAS..110.9824F. doi:10.1073/pnas.1307701110. PMC 3683724. PMID 23703908.
- ^ Nabors, Murray W. (2004). Introduction to Botany. San Francisco, CA: Pearson Education, Inc. ISBN 978-0-8053-4416-5.
- ^ Allaby, M., ed. (1992). "Algae". The Concise Dictionary of Botany. Oxford: Oxford University Press.
- ^ Guiry MD (October 2012). "How many species of algae are there?". Journal of Phycology. 48 (5): 1057–63. doi:10.1111/j.1529-8817.2012.01222.x. PMID 27011267. S2CID 30911529.
- ^ A b Guiry, M.D .; Guiry, G.M. (2016). "Algaebase". www.algaebase.org. Citováno 20. listopadu 2016.
- ^ D. Thomas (2002). Mořské řasy. Life Series. Muzeum přírodní historie, Londýn. ISBN 978-0-565-09175-0.
- ^ Hoek, Christiaan; den Hoeck, Hoeck Van; Mann, David; Jahns, H.M. (1995). Algae : an introduction to phycology. Cambridge University Press. p. 166. ISBN 9780521316873. OCLC 443576944.
- ^ Starckx, Senne (31 October 2012) A place in the sun - Algae is the crop of the future, according to researchers in Geel Flanders Today, Retrieved 8 December 2012
- ^ Duval, B.; Margulis, L. (1995). "The microbial community of Ophrydium versatile colonies: endosymbionts, residents, and tenants". Symbióza. 18: 181–210. PMID 11539474.
- ^ Mandoli, DF (1998). "Elaboration of Body Plan and Phase Change during Development of Acetabularia: How Is the Complex Architecture of a Giant Unicell Built?". Roční přehled fyziologie rostlin a molekulární biologie rostlin. 49: 173–198. doi:10.1146/annurev.arplant.49.1.173. PMID 15012232. S2CID 6241264.
- ^ Pierre Madl; Maricela Yip (2004). "Literature Review of Caulerpa taxifolia". BUFUS-Info. 19 (31).
- ^ Lalli, C.; Parsons, T. (1993). Biological Oceanography: An Introduction. Butterworth-Heinemann. ISBN 0-7506-3384-0.
- ^ Eerie red glow traces ocean plant health Věda NASA, 28. května 2009.
- ^ Behrenfeld, M.J.; Westberry, T.K.; Boss, E.S.; O'Malley, R.T.; Siegel, D.A.; Wiggert, J.D.; Franz, B.A .; McLain, C.R.; Feldman, G.C.; Doney, S.C.; Moore, J.K. (2009). "Satellite-detected fluorescence reveals global physiology of ocean phytoplankton". Biogeovědy. 6 (5): 779–794. Bibcode:2009BGeo....6..779B. doi:10.5194/bg-6-779-2009.
- ^ A b Wassilieff, Maggy (2006) "Plankton - Plant plankton", Te Ara - encyklopedie Nového Zélandu. Přístup: 2. listopadu 2019.
- ^ Lindsey, R., Scott, M. a Simmon, R. (2010) „Co jsou to fytoplankton“. NASA Earth Observatory.
- ^ Field, C. B.; Behrenfeld, M. J.; Randerson, J. T .; Falwoski, P. G. (1998). "Primary production of the biosphere: Integrating terrestrial and oceanic components". Věda. 281 (5374): 237–240. Bibcode:1998Sci...281..237F. doi:10.1126/science.281.5374.237. PMID 9657713.
- ^ Treguer, P.; Nelson, D. M.; Van Bennekom, A. J.; Demaster, D. J.; Leynaert, A.; Queguiner, B. (1995). "The Silica Balance in the World Ocean: A Reestimate". Věda. 268 (5209): 375–9. Bibcode:1995Sci...268..375T. doi:10.1126/science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ "King's College London - Lake Megachad". www.kcl.ac.uk. Citováno 5. května 2018.
- ^ Arsenieff, L.; Simon, N.; Rigaut-Jalabert, F.; Le Gall, F.; Chaffron, S.; Corre, E.; Com, E.; Bigeard, E.; Baudoux, A.C. (2018). "First Viruses Infecting the Marine Diatom Guinardia delicatula". Hranice v mikrobiologii. 9: 3235. doi:10.3389/fmicb.2018.03235. PMC 6334475. PMID 30687251.
- ^ Zhang, D .; Wang, Y .; Cai, J .; Pan, J.; Jiang, X .; Jiang, Y. (2012). "Bio-manufacturing technology based on diatom micro- and nanostructure". Bulletin čínské vědy. 57 (30): 3836–3849. Bibcode:2012ChSBu..57.3836Z. doi:10.1007/s11434-012-5410-x.
- ^ Wassilieff, Maggy (2006) "A coccolithophore", Te Ara - encyklopedie Nového Zélandu. Přístup: 2. listopadu 2019.
- ^ Rost, B. and Riebesell, U. (2004) "Coccolithophores and the biological pump: responses to environmental changes". V: Coccolithophores: From Molecular Processes to Global Impact, pages 99–125, Springer. ISBN 9783662062784.
- ^ DeLong, E.F.; Beja, O. (2010). "The light-driven proton pump proteorhodopsin enhances bacterial survival during tough times". PLOS Biology. 8 (4): e1000359. doi:10.1371/journal.pbio.1000359. PMC 2860490. PMID 20436957. e1000359.
- ^ A b Gómez-Consarnau, L.; Raven, J.A.; Levine, N.M.; Cutter, L.S.; Wang, D .; Seegers, B.; Arístegui, J.; Fuhrman, J.A.; Gasol, J.M.; Sañudo-Wilhelmy, S.A. (2019). "Microbial rhodopsins are major contributors to the solar energy captured in the sea". Vědecké zálohy. 5 (8): eaaw8855. Bibcode:2019SciA....5.8855G. doi:10.1126/sciadv.aaw8855. PMC 6685716. PMID 31457093.
- ^ Oren, Aharon (2002). "Molecular ecology of extremely halophilic Archaea and Bacteria". Ekologie mikrobiologie FEMS. 39 (1): 1–7. doi:10.1111/j.1574-6941.2002.tb00900.x. ISSN 0168-6496. PMID 19709178.
- ^ Béja, O.; Aravind, L .; Koonin, E.V.; Suzuki, M.T.; Hadd, A.; Nguyen, L.P.; Jovanovich, S.B.; Gates, C.M.; Feldman, R.A.; Spudich, J.L.; Spudich, E.N. (2000). "Bacterial rhodopsin: evidence for a new type of phototrophy in the sea". Věda. 289 (5486): 1902–1906. Bibcode:2000Sci...289.1902B. doi:10.1126/science.289.5486.1902. PMID 10988064. S2CID 1461255.
- ^ "Interviews with Fellows: Ed Delong". Americká akademie mikrobiologie. Archivovány od originál dne 7. srpna 2016. Citováno 2. července 2016.
- ^ Bacteria with Batteries, Popular Science, January 2001, Page 55.
- ^ Béja, O.; Aravind, L .; Koonin, E.V.; Suzuki, M.T.; Hadd, A.; Nguyen, L.P.; Jovanovich, S.B.; Gates, C.M.; Feldman, R.A.; Spudich, J.L.; Spudich, E.N. (2000). "Bacterial rhodopsin: evidence for a new type of phototrophy in the sea". Věda. 289 (5486): 1902–1906. Bibcode:2000Sci...289.1902B. doi:10.1126/science.289.5486.1902. PMID 10988064.
- ^ Boeuf, Dominique; Audic, Stéphane; Brillet-Guéguen, Loraine; Caron, Christophe; Jeanthon, Christian (2015). "MicRhoDE: a curated database for the analysis of microbial rhodopsin diversity and evolution". Databáze. 2015: bav080. doi:10.1093/database/bav080. ISSN 1758-0463. PMC 4539915. PMID 26286928.
- ^ Yawo, Hiromu; Kandori, Hideki; Koizumi, Amane (5 June 2015). Optogenetics: Light-Sensing Proteins and Their Applications. Springer. s. 3–4. ISBN 978-4-431-55516-2. Citováno 30. září 2015.
- ^ A tiny marine microbe could play a big role in climate change University of Southern California, Press Room, 8 August 2019.
- ^ DasSarma, Shiladitya; Schwieterman, Edward W. (11 October 2018). "Early evolution of purple retinal pigments on Earth and implications for exoplanet biosignatures". International Journal of Astrobiology: 1–10. arXiv:1810.05150. Bibcode:2018arXiv181005150D. doi:10.1017/S1473550418000423. ISSN 1473-5504. S2CID 119341330.
- ^ Sparks, William B.; DasSarma, S.; Reid, I. N. (December 2006). "Evolutionary Competition Between Primitive Photosynthetic Systems: Existence of an early purple Earth?". American Astronomical Society Meeting Abstracts. 38: 901. Bibcode:2006AAS...209.0605S.
- ^ Redfield, Alfred C. (1934). „O podílech organických derivátů v mořské vodě a jejich vztahu ke složení planktonu“. V Johnstone, James; Daniel, Richard Jellicoe (eds.). James Johnstone Memorial Volume. Liverpool: University Press of Liverpool. str. 176–92. OCLC 13993674.
- ^ A b Arrigo, Kevin R. (2005). "Mořské mikroorganismy a globální výživové cykly". Příroda. 437 (7057): 349–55. Bibcode:2005 Natur.437..349A. doi:10.1038 / nature04159. PMID 16163345. S2CID 62781480.
- ^ Martiny, A.C.; Pham, C.T.; Primeau, F.W.; Vrugt, J.A.; Moore, J.K.; Levin, S.A.; Lomas, M.W. (2013). "Strong latitudinal patterns in the elemental ratios of marine plankton and organic matter". Nature Geoscience. 6 (4): 279–283. Bibcode:2013NatGe...6..279M. doi:10.1038/ngeo1757.
- ^ Eppley, R.W.; Peterson, B.J. (1979). "Particulate organic matter flux and planktonic new production in the deep ocean". Příroda. 282 (5740): 677–680. Bibcode:1979Natur.282..677E. doi:10.1038/282677a0. S2CID 42385900.
- ^ Velicer, Gregory J.; Mendes-Soares, Helena (2007). "Bacterial predators" (PDF). Buňka. 19 (2): R55–R56. doi:10.1016 / j.cub.2008.10.043. PMID 19174136. S2CID 5432036.
- ^ Stevens, Alison N. P. (2010). "Predation, Herbivory, and Parasitism". Znalosti o výchově k přírodě. 3 (10): 36.
- ^ Jurkevitch, Edouard; Davidov, Yaacov (2006). "Phylogenetic Diversity and Evolution of Predatory Prokaryotes". Predatory Prokaryotes. Springer. str.11 –56. doi:10.1007/7171_052. ISBN 978-3-540-38577-6.
- ^ Hansen, Per Juel; Bjørnsen, Peter Koefoed; Hansen, Benni Winding (1997). "Zooplankton grazing and growth: Scaling within the 2-2,-μm body size range". Limnologie a oceánografie. 42 (4): 687–704. doi:10.4319/lo.1997.42.4.0687. summarizes findings from many authors.
- ^ A b Wassilieff, Maggy (2006) "Plankton - zvířecí plankton", Te Ara - encyklopedie Nového Zélandu. Přístup: 2. listopadu 2019.
- ^ Varea, C.; Aragon, J.L.; Barrio, R.A. (1999). "Turing patterns on a sphere". Fyzický přehled E. 60 (4): 4588–92. Bibcode:1999PhRvE..60.4588V. doi:10.1103/PhysRevE.60.4588. PMID 11970318.
- ^ A b Hemleben, C .; Anderson, O.R .; Spindler, M. (1989). Moderní planktonická foraminifera. Springer-Verlag. ISBN 978-3-540-96815-3.
- ^ Foraminifera: Historie studia, University College v Londýně. Citováno: 18. listopadu 2019.
- ^ Advances in Microbial Ecology, Volum 11
- ^ Bernhard, J. M .; Bowser, S.M. (1999). „Benthic Foraminifera of dysoxic sediment: chloroplast sekvestrace a funkční morfologie“. Recenze vědy o Zemi. 46 (1): 149–165. Bibcode:1999ESRv ... 46..149B. doi:10.1016 / S0012-8252 (99) 00017-3.
- ^ Dierssen, Heidi; McManus, George B.; Chlus, Adam; Qiu, Dajun; Gao, Bo-Cai; Lin, Senjie (2015). "Space station image captures a red tide ciliate bloom at high spectral and spatial resolution". Sborník Národní akademie věd. 112 (48): 14783–14787. Bibcode:2015PNAS..11214783D. doi:10.1073/pnas.1512538112. PMC 4672822. PMID 26627232.
- ^ Dejte si pozor na mixotrofy - mohou zničit celé ekosystémy „během několika hodin“
- ^ Mikroskopické únosce těla zamořují naše oceány - Phys.org
- ^ Eiler A (prosinec 2006). „Důkazy o všudypřítomnosti mixotrofních bakterií v horním oceánu: důsledky a důsledky“. Appl Environ Microbiol. 72 (12): 7431–7. doi:10.1128 / AEM.01559-06. PMC 1694265. PMID 17028233.
- ^ Katechakis A, Stibor H (červenec 2006). „Mixotrof Ochromonas tuberculata může napadnout a potlačit specializovaná společenství fago- a fototrofních planktonů v závislosti na podmínkách živin “. Ekologie. 148 (4): 692–701. Bibcode:2006Oecol.148..692K. doi:10.1007 / s00442-006-0413-4. PMID 16568278. S2CID 22837754.
- ^ Schoemann, Véronique; Becquevort, Sylvie; Stefels, Jacqueline; Rousseau, Véronique; Lancelot, Christiane (1 January 2005). „Květy Phaeocystis v globálním oceánu a jejich kontrolní mechanismy: recenze“. Journal of Sea Research. Železné zdroje a oceánské živiny - pokrok v globálních environmentálních simulacích. 53 (1–2): 43–66. Bibcode:2005JSR .... 53 ... 43S. CiteSeerX 10.1.1.319.9563. doi:10.1016 / j.seares.2004.01.008.
- ^ „Vítejte na domovské stránce projektu sekvenování genomu Phaeocystis antarctica.“.
- ^ DiTullio, G. R .; Grebmeier, J. M .; Arrigo, K. R .; Lizotte, M. P .; Robinson, D. H .; Leventer, A .; Barry, J. P .; VanWoert, M. L .; Dunbar, R. B. (2000). „Rychlý a včasný vývoz květů Phaeocystis antarctica kvete v Rossově moři v Antarktidě“. Příroda. 404 (6778): 595–598. doi:10.1038/35007061. PMID 10766240. S2CID 4409009.
- ^ J, Stefels; L, Dijkhuizen; WWC, Gieskes (20 July 1995). „Aktivita DMSP-lyázy v jarním květu fytoplanktonu u holandského pobřeží, související s hojností Phaeocystis sp.“ (PDF). Série pokroku v ekologii moří. 123: 235–243. Bibcode:1995MEPS..123..235S. doi:10,3 354 / meps123235.
- ^ Decelle, Johan; Simó, Rafel; Galí, Martí; Vargas, Colomban de; Colin, Sébastien; Desdevises, Yves; Bittner, Lucie; Probert, Ian; Not, Fabrice (30 October 2012). „Originální režim symbiózy v otevřeném oceánu planktonu“. Sborník Národní akademie věd. 109 (44): 18000–18005. Bibcode:2012PNAS..10918000D. doi:10.1073 / pnas.1212303109. ISSN 0027-8424. PMC 3497740. PMID 23071304.
- ^ Mars Brisbin, Margaret; Grossmann, Mary M .; Mesrop, Lisa Y .; Mitarai, Satoshi (2018). „In-host Symbiont Diversity and Extended Symbiont rozmanitost ve Photosymbiotické Acantharea (Clade F)“. Hranice v mikrobiologii. 9: 1998. doi:10.3389 / fmicb.2018.01998. ISSN 1664-302X. PMC 6120437. PMID 30210473.
- ^ A b Stoecker, D.K .; Hansen, P.J .; Caron, D. A.; Mitra, A. (2017). "Mixotrophy in the marine plankton". Výroční zpráva o námořní vědě. 9: 311–335. Bibcode:2017 ARMS .... 9..311S. doi:10.1146 / annurev-marine-010816-060617. PMID 27483121.
- ^ A b Mitra, A; Flynn, KJ; Tillmann, U; Raven, J; Caron, D; et al. (2016). „Definování funkčních skupin protonů planktonu na mechanismech pro získávání energie a živin; začlenění různých mixotrofních strategií“. Protist. 167 (2): 106–20. doi:10.1016 / j.protis.2016.01.003. PMID 26927496.
- ^ Gómez F (2012). „Kontrolní seznam a klasifikace živých dinoflagelátů (Dinoflagellata, Alveolata)“ (PDF). CICIMAR Océanides. 27 (1): 65–140. Archivovány od originál (PDF) dne 27. listopadu 2013.
- ^ Stoecker DK (1999). "Mixotrofie mezi dinoflageláty". The Journal of Eukaryotic Microbiology. 46 (4): 397–401. doi:10.1111 / j.1550-7408.1999.tb04619.x. S2CID 83885629.
- ^ Suggested Explanation for Glowing Seas--Including Currently Glowing California Seas Národní vědecká nadace, 18. října 2011.
- ^ Boltovskoy, Demetrio; Anderson, O. Roger; Correa, Nancy M. (2017). Příručka protistů. Springer, Cham. str. 731–763. doi:10.1007/978-3-319-28149-0_19. ISBN 9783319281476.
- ^ Anderson, O. R. (1983). Radiolaria. Springer Science & Business Media.
- ^ Gast, R. J .; Caron, D. A. (1 November 1996). "Molekulární fylogeneze symbiotických dinoflagelátů z planktonické foraminifera a radiolaria". Molekulární biologie a evoluce. 13 (9): 1192–1197. doi:10.1093 / oxfordjournals.molbev.a025684. ISSN 0737-4038. PMID 8896371.
- ^ Castro P, Huber ME (2010). Mořská biologie (8. vydání). McGraw Hill. p. 95. ISBN 978-0071113021.
- ^ Hastings JW (1996). "Chemistries and colors of bioluminescent reactions: a review". Gen. 173 (1 Spec No): 5–11. doi:10.1016/0378-1119(95)00676-1. PMID 8707056.
- ^ Haddock SH, Moline MA, Case JF (2009). "Bioluminescence in the sea". Výroční zpráva o námořní vědě. 2: 443–93. Bibcode:2010 ARMS .... 2..443H. doi:10.1146 / annurev-marine-120308-081028. PMID 21141672. S2CID 3872860.
- ^ "Protozoa infikující žáby a kůži". Veterinární příručka společnosti Merck. Archivovány od originál dne 3. března 2016. Citováno 4. listopadu 2019.
- ^ Brand, Larry E .; Campbell, Lisa; Bresnan, Eileen (2012). "Karenia: Biologie a ekologie toxického rodu ". Škodlivé řasy. 14: 156–178. doi:10.1016 / j.hal.2011.10.020.
- ^ Buskey, E.J. (1995). "Growth and bioluminescence of Noctiluca scintillans on varying algal diets". Journal of Plankton Research. 17 (1): 29–40. doi:10.1093/plankt/17.1.29.
- ^ Thomson, Charles Wyville (2014) Voyage of the Challenger: The Atlantic Cambridge University Press, page235. ISBN 9781108074759.
- ^ Grethe R. Hasle; Erik E. Syvertsen; Karen A. Steidinger; Karl Tangen (25 January 1996). "Marine Diatoms". In Carmelo R. Tomas (ed.). Identifying Marine Diatoms and Dinoflagellates. Akademický tisk. pp. 5–385. ISBN 978-0-08-053441-1. Citováno 13. listopadu 2013.
- ^ Ald, S.M.; et al. (2007). "Diversity, Nomenclature, and Taxonomy of Protists" (PDF). Syst. Biol. 56 (4): 684–689. doi:10.1080/10635150701494127. PMID 17661235. Archivovány od originál (PDF) dne 31. března 2011. Citováno 11. října 2019.
- ^ Moheimani, N.R .; Webb, J.P .; Borowitzka, M.A. (2012), „Bioremediace a další potenciální aplikace řas kokokolithophoridů: přehled. Bioremediace a další potenciální aplikace řas kokokolithophoridů: přehled“, Výzkum řas, 1 (2): 120–133, doi:10.1016 / j.algal.2012.06.002
- ^ Taylor, A.R .; Chrachri, A .; Wheeler, G .; Goddard, H .; Brownlee, C. (2011). „Napěťově řízený kanál H +, který je základem homeostázy pH v kalcifikujících kokokolithophorech“. PLOS Biology. 9 (6): e1001085. doi:10.1371 / journal.pbio.1001085. PMC 3119654. PMID 21713028.
- ^ Wierer, U .; Arrighi, S .; Bertola, S .; Kaufmann, G .; Baumgarten, B .; Pedrotti, A .; Pernter, P .; Pelegrin, J. (2018). „Iceman's lithic toolkit: Suroviny, technologie, typologie a použití“. PLOS ONE. 13 (6): e0198292. Bibcode:2018PLoSO..1398292W. doi:10.1371 / journal.pone.0198292. PMC 6010222. PMID 29924811.
- ^ A b Haq B.U. and Boersma A. (Eds.) (1998) Úvod do mořské mikropaleontologie Elsevier. ISBN 9780080534961
- ^ Bruckner, Monica (2020) „Paleoklimatologie: Jak můžeme odvodit minulé podnebí?“ SERC, Carleton College. Upraveno 23. července 2020. Citováno 10. září 2020.
- ^ Země se blíží ke stavu „skleníku“, který nebyl zaznamenán za 50 milionů let, ukazují epické nové klimatické záznamy LiveScience, 10. září 2020.
- ^ Westerhold, T., Marwan, N., Drury, AJ, Liebrand, D., Agnini, C., Anagnostou, E., Barnet, JS, Bohaty, SM, Vleeschouwer, D., Florindo, F. a Frederichs, T . (2020) „Astronomicky datovaný záznam klimatu Země a jeho předvídatelnosti za posledních 66 milionů let“. Věda, 369(6509): 1383–1387. doi:10.1126 / science.aba6853.
- ^ Bob Yirka 29. srpna 2013
- ^ Sussman: Nejstarší rostliny, Opatrovník, 2. května 2010
- ^ „Archivovaná kopie“. Archivovány od originál dne 13. července 2018. Citováno 13. července 2018.CS1 maint: archivovaná kopie jako titul (odkaz)
- ^ Willerslev, Eske; Froese, Duane; Gilichinsky, David; Ronn, Regin; Bunce, Michael; Zuber, Maria T .; Gilbert, M. Thomas P .; Značka, Tina; Munch, Kasper; Nielsen, Rasmus; Mastepanov, Michail; Christensen, Torben R .; Hebsgaard, Martin B .; Johnson, Sarah Stewart (4. září 2007). „Starověké bakterie ukazují důkazy o opravě DNA. Sborník Národní akademie věd. 104 (36): 14401–14405. Bibcode:2007PNAS..10414401J. doi:10.1073 / pnas.0706787104. PMC 1958816. PMID 17728401.
- ^ Egan, S., Fukatsu, T. a Francino, M.P., 2020. Příležitosti a výzvy pro výzkum mikrobiální symbiózy v éře mikrobiomu. Frontiers in Microbiology, 11. doi:10.3389 / fmicb.2020.01150. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Margulis, Lynn; Fester, René (1991). Symbióza jako zdroj evolučních inovací. MIT Stiskněte. ISBN 9780262132695.
- ^ Bordenstein, Seth R .; Theis, Kevin R. (2015). „Hostitelská biologie ve světle mikrobiomu: deset principů holobiontů a hologenomů“. PLOS Biol. 13 (8): e1002226. doi:10.1371 / journal.pbio.1002226. ISSN 1545-7885. PMC 4540581. PMID 26284777.
- ^ Theis, K.R., Dheilly, N.M., Klassen, J.L., Brucker, R.M., Baines, J.F., Bosch, T.C., Cryan, J.F., Gilbert, S.F., Goodnight, C.J., Lloyd, E.A. a Sapp, J. (2016) „Správný koncept hologenomu: ekologicko-evoluční rámec pro hostitele a jejich mikrobiomy“. Msystems, 1(2). doi:10,1 128 / m Systémy 00028-16. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Bulgarelli, D., Schlaeppi, K., Spaepen, S., Van Themaat, E.V.L. a Schulze-Lefert, P. (2013) „Struktura a funkce bakteriální mikrobioty rostlin“. Roční přehled biologie rostlin, 64: 807–838. doi:10.1146 / annurev-arplant-050312-120106.
- ^ Vandenkoornhuyse, Philippe; Quaiser, Achim; Duhamel, Marie; Le Van, Amandine; Dufresne, Alexis (1. června 2015). „Význam mikrobiomu rostliny holobiont“. Nový fytolog. 206 (4): 1196–1206. doi:10,1111 / nph.13312. ISSN 1469-8137. PMID 25655016.
- ^ Sánchez-Cañizares, C., Jorrín, B., Poole, P.S. a Tkacz, A. (2017) „Porozumění holobiontu: vzájemná závislost rostlin a jejich mikrobiomu“. Současný názor v mikrobiologii, 38: 188–196. doi:10.1016 / j.mib.2017.07.001.
- ^ Douglas, A.E. a Werren, J.H. (2016) „Díry v hologenomu: proč symbiózy hostitel-mikrob nejsou holobionty“. MBio, 7(2). doi:10,1 128 / mBio.02099-15.
- ^ Thompson, J.R., Rivera, H.E., Closek, C.J. a Medina, M. (2015) „Mikroby v korálovém holobiontu: partneři prostřednictvím evoluce, vývoje a ekologických interakcí“. Hranice v buněčné a infekční mikrobiologii, 4: 176. doi:10.3389 / fcimb.2014.00176.
- ^ Pita, L., Rix, L., Slaby, B.M., Franke, A. a Hentschel, U. (2018) „Houba holobiont v měnícím se oceánu: od mikrobů po ekosystémy“. Mikrobiom, 6(1): 46. doi:10.1186 / s40168-018-0428-1. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Ugarelli, K., Chakrabarti, S., Laas, P. a Stingl, U. (2017) „The seagrass holobiont and its microbiome“. Mikroorganismy, 5(4): 81. doi:10,3390 / mikroorganismy5040081. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Cavalcanti, G.S., Shukla, P., Morris, M., Ribeiro, B., Foley, M., Doane, M.P., Thompson, C.C., Edwards, M.S., Dinsdale, E.A. a Thompson, F.L. (2018) „Rhodoliths holobionts in the changing ocean: host-microbes interactions mediace coralline algae resilience under ocean acid“. BMC Genomics, 19(1): 1–13. doi:10.1186 / s12864-018-5064-4. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Knowlton, N. a Rohwer, F. (2003) „Vícedruhový mikrobiální vzájemný vztah na korálových útesech: hostitel jako stanoviště“. Americký přírodovědec, 162(S4): S51-S62. doi:10.1086/378684.
- ^ Pollock, F. Joseph; McMinds, Ryan; Smith, Styles; Bourne, David G .; Willis, Bette L .; Medina, Mónica; Thurber, Rebecca Vega; Zaneveld, Jesse R. (22. listopadu 2018). „Bakterie spojené s korály vykazují fylosymbiózu a kopylogenezi“. Příroda komunikace. 9 (1): 1–13. doi:10.1038 / s41467-018-07275-x. ISSN 2041-1723. PMID 30467310.
- ^ Wilhelm, Steven W .; Suttle, Curtis A. (1999). „Viry a cykly živin v moři“. BioScience. 49 (10): 781–788. doi:10.2307/1313569. ISSN 1525-3244. JSTOR 1313569.
- ^ Weinbauer, Markus G .; et al. (2007). „Synergické a antagonistické účinky virové lýzy a pastvy protistanu na bakteriální biomasu, produkci a rozmanitost“. Mikrobiologie prostředí. 9 (3): 777–788. doi:10.1111 / j.1462-2920.2006.01200.x. PMID 17298376.
- ^ Robinson, Carol a Nagappa Ramaiah. „Mikrobiální heterotrofní metabolické rychlosti omezují mikrobiální uhlíkovou pumpu.“ Americká asociace pro pokrok ve vědě, 2011.
- ^ Gutierrez, MH; Jara, AM; Pantoja, S (2016). „Plísňové parazity infikují mořské rozsivky v upwellingovém ekosystému Humboldtova současného systému mimo centrální Chile“. Environ Microbiol. 18 (5): 1646–1653. doi:10.1111/1462-2920.13257. hdl:10533/148260. PMID 26914416.
- ^ Co je mořský sníh? NOAA National Ocean Service. Aktualizováno: 25.06.18.
- ^ Monroy, P., Hernández-García, E., Rossi, V. a López, C. (2017) „Modelování dynamického potopení biogenních částic v oceánském toku“. Nelineární procesy v geofyzice, 24(2): 293–305. doi:10.5194 / npg-24-293-2017. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Creative Commons Attribution 3.0 International License.
- ^ Simon, M., Grossart, H., Schweitzer, B. a Ploug, H. (2002) „Mikrobiální ekologie organických agregátů ve vodních ekosystémech“. Vodní mikrobiální ekologie, 28: 175–211. doi:10 3354 / ame028175.
- ^ Boetius, Antje; Anesio, Alexandre M .; Deming, Jody W .; Mikucki, Jill A .; Rapp, Josephine Z. (1. listopadu 2015). „Mikrobiální ekologie kryosféry: mořský led a ledovcová stanoviště“. Příroda Recenze Mikrobiologie. 13 (11): 677–690. doi:10.1038 / nrmicro3522. PMID 26344407. S2CID 20798336.
- ^ Lutz, Richard A .; Kennish, Michael J. (1993). „Ekologie hlubinných hydrotermálních větracích komunit: přehled“. Recenze geofyziky. 31 (3): 211. Bibcode:1993RvGeo..31..211L. doi:10.1029 / 93rg01280. ISSN 8755-1209.
- ^ Kádár, Enikõ; Costa, Valentina; Santos, Ricardo S .; Powell, Jonathan J. (červenec 2006). „Tkáňové dělení mikroesenciálních kovů ve větrném mlži Bathymodiolus azoricus a přidružených organizmech (bakterie endosymbiont a parazit polychaete) z geochemicky odlišných průduchů Středoatlantického hřbetu“. Journal of Sea Research. 56 (1): 45–52. Bibcode:2006JSR .... 56 ... 45 tis. doi:10.1016 / j.seares.2006.01.002. ISSN 1385-1101.
- ^ Anderson, Rika E .; Brazelton, William J .; Baross, John A. (2011). „Je genetická krajina hluboké podpovrchové biosféry ovlivněna viry?“. Hranice v mikrobiologii. 2: 219. doi:10.3389 / fmicb.2011.00219. ISSN 1664-302X. PMC 3211056. PMID 22084639.
- ^ Suttle, Curtis A. (září 2005). "Viry v moři". Příroda. 437 (7057): 356–361. Bibcode:2005 Natur.437..356S. doi:10.1038 / nature04160. ISSN 0028-0836. PMID 16163346. S2CID 4370363.
- ^ Suttle, Curtis A. (říjen 2007). „Mořské viry - hlavní hráči v globálním ekosystému“. Příroda Recenze Mikrobiologie. 5 (10): 801–812. doi:10.1038 / nrmicro1750. ISSN 1740-1526. PMID 17853907. S2CID 4658457.
- ^ Anderson, Rika E .; Sogin, Mitchell L .; Baross, John A. (3. října 2014). „Evoluční strategie virů, bakterií a archea v hydrotermálních ventilačních ekosystémech odhalená prostřednictvím metagenomiky“. PLOS ONE. 9 (10): e109696. Bibcode:2014PLoSO ... 9j9696A. doi:10.1371 / journal.pone.0109696. ISSN 1932-6203. PMC 4184897. PMID 25279954.
- ^ Ortmann, Alice C .; Suttle, Curtis A. (srpen 2005). „Vysoký výskyt virů v hlubinném hydrotermálním ventilačním systému naznačuje mikrobiální úmrtnost zprostředkovanou viry“. Deep Sea Research Part I: Oceanographic Research Papers. 52 (8): 1515–1527. Bibcode:2005DSRI ... 52.1515O. doi:10.1016 / j.dsr.2005.04.002. ISSN 0967-0637.
- ^ Breitbart, Mya (15. ledna 2012). „Marine Viruses: Truth or Dare“. Výroční zpráva o námořní vědě. 4 (1): 425–448. Bibcode:2012 ARMS .... 4..425B. doi:10.1146 / annurev-marine-120709-142805. ISSN 1941-1405. PMID 22457982.
- ^ Goldenfeld, Nigel; Woese, Carl (leden 2007). „Další revoluce biologie“. Příroda. 445 (7126): 369. arXiv:q-bio / 0702015. Bibcode:2007 Natur.445..369G. doi:10.1038 / 445369a. ISSN 0028-0836. PMID 17251963. S2CID 10737747.
- ^ Deep Carbon Observatory (10. prosince 2018). „Život na hluboké Zemi činí celkem 15 až 23 miliard tun uhlíku - stokrát více než u lidí“. EurekAlert!. Citováno 11. prosince 2018.
- ^ Dockrill, Peter (11. prosince 2018). „Vědci odhalují masivní biosféru života ukrytou pod povrchem Země“. Vědecké varování. Citováno 11. prosince 2018.
- ^ Gabbatiss, Josh (11. prosince 2018). „Masivní studie„ hlubokého života “odhalila miliardy tun mikrobů žijících hluboko pod povrchem Země“. Nezávislý. Citováno 11. prosince 2018.
- ^ Lollar, Garnet S .; Warr, Oliver; Vyprávění, Jon; Osburn, Magdalena R .; Sherwood Lollar, Barbara (2019). "'Sledujte vodu ': Hydrogeochemická omezení při mikrobiálním výzkumu 2,4 km pod povrchem na observatoři Deep Fluid and Deep Life Observatory v Kidd Creek “. Geomikrobiologický deník. 36: 859–872. doi:10.1080/01490451.2019.1641770. S2CID 199636268.
- ^ Nejstarší podzemní voda na světě podporuje život chemií Water-Rock, 29. července 2019, deepcarbon.net.
- ^ Podivné formy života nalezené hluboko v důlním bodě na obrovských „podzemních Galapágách“, Autor: Corey S. Powell, 7. září 2019, nbcnews.com.
- ^ Nejstarší voda na Zemi nalezená hluboko v kanadském štítu, 14. prosince 2016, Maggie Romuld
- ^ Wu, Katherine J. (28. července 2020). „Tyto mikroby mohly přežít 100 milionů let pod mořským dnem - bakterie se zachránily ze svých chladných, stísněných domovů a míst chudých na živiny, probudily se v laboratoři a rostly“. Citováno 31. července 2020.
- ^ Morono, Yuki; et al. (28. července 2020). „Aerobní mikrobiální život přetrvává v oxickém mořském sedimentu starém až 101,5 milionu let“. Příroda komunikace. 11 (3626): 3626. doi:10.1038 / s41467-020-17330-1. PMC 7387439. PMID 32724059.
- ^ A b Filee, J .; Tetart, F .; Suttle, C. A .; Krisch, H. M. (2005). „Mořské bakteriofágy typu T4, všudypřítomná součást temné hmoty biosféry“. Sborník Národní akademie věd. 102 (35): 12471–12476. Bibcode:2005PNAS..10212471F. doi:10.1073 / pnas.0503404102. ISSN 0027-8424. PMC 1194919. PMID 16116082.
- ^ University of Tennessee v Knoxville (25. září 2018). „Studie: Mikrobiální temná hmota dominuje prostředím Země“. Eurekalert!. Citováno 26. září 2018.
- ^ Hedlund, Brian P .; Dodsworth, Jeremy A .; Murugapiran, Senthil K .; Rinke, Christian; Woyke, Tanja (2014). „Dopad jednobuněčné genomiky a metagenomiky na vznikající pohled na extremofilní“ mikrobiální temnou hmotu"". Extremophiles. 18 (5): 865–875. doi:10.1007 / s00792-014-0664-7. ISSN 1431-0651. PMID 25113821. S2CID 16888890.
- ^ Květen, R.M. (1988). „Kolik druhů je na zemi?“. Věda. 241 (4872): 1441–1449. Bibcode:1988Sci ... 241.1441M. doi:10.1126 / science.241.4872.1441. PMID 17790039. S2CID 34992724.
- ^ Locey, K.J .; Lennon, J.T. (2016). „Zákony o měřítku předpovídají globální mikrobiální rozmanitost“. Sborník Národní akademie věd. 113 (21): 5970–5975. Bibcode:2016PNAS..113,5970L. doi:10.1073 / pnas.1521291113. PMC 4889364. PMID 27140646.
- ^ A b Vitorino, L.C .; Bessa, L.A. (2018). „Mikrobiální rozmanitost: rozdíl mezi odhadovanou a známou“. Rozmanitost. 10 (2): 46. doi:10,3390 / d10020046. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ A b C Karsenti, E .; Acinas, S.G .; Bork, P .; Bowler, C .; De Vargas, C .; Raes, J .; Sullivan, M .; Arendt, D .; Benzoni, F .; Claverie, J.M .; Následuje, M. (2011). „Holistický přístup k biologii mořských ekosystémů“. PLOS Biology. 9 (10): e1001177. doi:10.1371 / journal.pbio.1001177. PMC 3196472. PMID 22028628.
- ^ Olsen GJ, Woese CR, Overbeek R (1994). „Větry (evolučních) změn: vdechnutí nového života mikrobiologii“. Journal of Bacteriology. 176 (1): 1–6. doi:10.2172/205047. PMC 205007. PMID 8282683.
- ^ Woese, ČR (1987). "Bakteriální evoluce". Mikrobiologické recenze. 51 (2): 221–71. doi:10.1128 / MMBR.51.2.221-271.1987. PMC 373105. PMID 2439888.
- ^ A b Collins, R. Eric (2019) Malý a mocný - mikrobiální život NOAA: Průzkum oceánu a výzkumExpedice Gulf of Alaska Seamounts 2019.
 Tento článek včlení text z tohoto zdroje, který je v veřejná doména.
Tento článek včlení text z tohoto zdroje, který je v veřejná doména. - ^ Käse L, Geuer JK. (2018) „Reakce fytoplanktonu na změnu mořského podnebí - úvod“. In Jungblut S., Liebich V., Bode M. (Eds) YOUMARES 8 – Oceans Across Boundaries: Learning from each other, strany 55–72, Springer. doi:10.1007/978-3-319-93284-2_5. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Franco-Duarte, R .; Černáková, L .; Kadam, S .; Kaushik, K .; Salehi, B .; Bevilacqua, A .; Corbo, M.R .; Antolak, H .; Dybka-Stępień, K .; Leszczewicz, M .; Relison Tintino, S. (2019). „Pokroky v chemických a biologických metodách k identifikaci mikroorganismů - od minulosti po současnost“. Mikroorganismy. 7 (5): 130. doi:10,3390 / mikroorganismy7050130. PMC 6560418. PMID 31086084. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Lozupone, C.A., Stombaugh, J.I., Gordon, J.I., Jansson, J.K. a Knight, R. (2012) „Diverzita, stabilita a odolnost lidské střevní mikrobioty“. Příroda, 489(7415): 220–230. doi:10.1038 / příroda 11550.
- ^ Venter, J. C., Remington, K., Heidelberg, J. F., Halpern, A. L., Rusch, D., Eisen, J. A., Wu, D., Paulsen, I., Nelson, K. E., Nelson, W. a Fouts, D. E. (2004) „Sekvenování brokovnice environmentálního genomu Sargasového moře“. Věda, 304(5667): 66–74. doi:10.1126 / science.1093857.
- ^ Liu, L., Li, Y., Li, S., Hu, N., He, Y., Pong, R., Lin, D., Lu, L. a Law, M. (2012) „Srovnání sekvenční systémy nové generace “. BioMed Research International, 2012: 251364. doi:10.1155/2012/251364.
- ^ Berg, Gabriele; Daria Rybakova, Doreen Fischer, Tomislav Cernava, Marie-Christine Champomier Vergès, Trevor Charles, Xiaoyulong Chen, Luca Cocolin, Kellye Eversole, Gema Herrero Corral, Maria Kazou, Linda Kinkel, Lene Lange, Nelson Lima, Alexander Loy, James A. Macklin Emmanuelle Maguin, Tim Mauchline, Ryan McClure, Birgit Mitter, Matthew Ryan, Inga Sarand, Hauke Smidt, Bettina Schelkle, Hugo Roume, G. Seghal Kiran, Joseph Selvin, Rafael Soares Correa de Souza, Leo van Overbeek, Brajesh K. Singh , Michael Wagner, Aaron Walsh, Angela Sessitsch a Michael Schloter (2020) „Znovu navštívená definice mikrobiomu: staré koncepty a nové výzvy“. Mikrobiom, 8(103): 1–22. doi:10.1186 / s40168-020-00875-0. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Kodzius, R .; Gojobori, T. (2015). „Marine metagenomics as a source for biospecting“. Marine Genomics. 24: 21–30. doi:10.1016 / j.margen.2015.07.001. PMID 26204808. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Holtorf, Hauke; Guitton, Marie-Christine; Reski, Ralfe (2002). "Rostlinná funkční genomika". Naturwissenschaften. 89 (6): 235–249. doi:10.1007 / s00114-002-0321-3. PMID 12146788. S2CID 7768096.
- ^ Cain, A.K., Barquist, L., Goodman, A.L., Paulsen, I.T., Parkhill, J. a van Opijnen, T. (2020) „Desetiletí pokroku v sekvenování transpozon-inzerce“. Genetika hodnocení přírody, 21: 526–540. doi:10.1038 / s41576-020-0244-x.
- ^ Zhang, W., Li, F. a Nie, L. (2010) „Integrating multiple 'omics' analysis for microbial biologics: application and metodologies“. Mikrobiologie, 156(2): 287–301. doi:10.1099 / mic.0.034793-0.
- ^ Singh, R.P. a Kothari, R. (2017) „Éra omiků a hostitelské mikrobiomy“. In: Singh RP, Kothari R, Koringa PG a Singh SP (Vyd.) Porozumění interakcím hostitel-mikrobiom - přístup Omics, strany 3–12, Springer. ISBN 9789811050503. doi:10.1007/978-981-10-5050-3_1
- ^ Capotondi, A., Jacox, M., Bowler, C., Kavanaugh, M., Lehodey, P., Barrie, D., Brodie, S., Chaffron, S., Cheng, W., Dias, DF, Eveillard , D. a 20 dalších (2019) „Pozorovací potřeby podporující modelování a předpovídání mořských ekosystémů: od globálního oceánu po regionální a pobřežní systémy“. Frontiers in Marine Science, 6: 623. doi:10.3389 / fmars.2019.00623. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Sunagawa, S., Coelho, L. P., Chaffron, S., Kultima, J. R., Labadie, K., Salazar, G. a kol. (2015). „Oceánský plankton. Struktura a funkce mikrobiomu globálního oceánu“. Věda, 348: 1261359. doi:10.1126 / science.1261359.
- ^ Guidi, L., Chaffron, S., Bittner, L., Eveillard, D., Larhlimi, A., Roux, S. a kol. (2016). „Sítě planktonu podporující export uhlíku v oligotrofním oceánu“. Příroda, 532: 465–470. doi:10.1038 / příroda16942.
- ^ Guyennon, A., Baklouti, M., Diaz, F., Palmieri, J., Beuvier, J., Lebaupin-Brossier, C., et al. (2015). „Nové poznatky o vývozu organického uhlíku ve Středozemním moři z trojrozměrného modelování“. Biogeovědy, 11: 425–442. doi:10.5194 / bg-12-7025-2015.
- ^ Louca, S., Parfrey, L. W. a Doebeli, M. (2016). "Oddělení funkce a taxonomie v mikrobiomu globálního oceánu". Věda, 353: 1272–1277. doi:10.1126 / science.aaf4507.
- ^ Coles, V. J., Stukel, M. R., Brooks, M. T., Burd, A., Crump, B. C., Moran, M. A. a kol. (2017). „Oceánská biogeochemie modelována s objevující se genomikou založenou na zvláštnostech“. Věda, 358: 1149–1154. doi:10.1126 / science.aan5712.
- ^ Mock, T., Daines, S.J., Geider, R., Collins, S., Metodiev, M., Millar, A.J., Moulton, V. a Lenton, T.M. (2016) „Překlenutí propasti mezi omikou a vědou o zemském systému, abychom lépe pochopili, jak změna životního prostředí ovlivňuje mořské mikroby“. Globální biologie změn, 22(1): 61–75. doi:10.1111 / gcb.12983. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ A b Toseland A, Daines SJ a Clark JR a kol. (2013) „Vliv teploty na alokaci a metabolismus mořských fytoplanktonů“. Přírodní změna podnebí, 3: 979– 984. doi:10.1038 / nclimate1989.
- ^ Martiny AC, Pham CTA, Primeau FW, Vrugt JA, Moore JK, Levin SA a Lomas MW (2013) „Silné zeměpisné šířky v poměru elementů mořského planktonu a organické hmoty“. Nature Geoscience, 6: 279–283. doi:10.1038 / ngeo1757.
Obecné odkazy
- Haq B.U. a Boersma A. (Eds.) (1998) Úvod do mořské mikropaleontologie Elsevier. ISBN 9780080534961
- Kirchman, David L. (2018) Procesy v mikrobiální ekologii Oxford University Press. ISBN 9780192506474
- Helmreich, Stefan (2009) Alien Ocean: Anthropological Voyages in Microbial Seas University of California Press. ISBN 9780520942608
- Munn, Colin B. (2019) Mořská mikrobiologie: ekologie a aplikace CRC Press. ISBN 9780429592362
- Stal LJ a Cretoiu MS (2016) Mořský mikrobiom: nevyužitý zdroj biologické rozmanitosti a biotechnologického potenciálu Springer. ISBN 9783319330006
- Middelboe M a Corina Brussaard CPD (vydání) Mořské viry 2016, MDPI. ISBN 978-3-03842-621-9 Stáhnout PDF
- Ohtsuka S, Suzaki T, Horiguchi T, Suzuki N and Not F (Eds.) (2015) Marine Protists: Diversity and Dynamics Springer. ISBN 9784431551300
- Johnson MD a V. Moeller HV (2019) Mixotrophy v Protists: Od modelových systémů k matematickým modelům, 2. vydání, Frontiers Media. ISBN 9782889631483
- Radiolarian umění
