Choanoflagellate - Choanoflagellate
| Choanoflageláty Časová řada: 900–0 Ma Nejsou známy žádné fosílie, důkazy molekulárních hodin pro původ 1050-800 Ma[1] | |
|---|---|
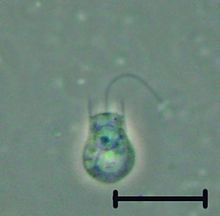 | |
| Codosiga sp. | |
| Vědecká klasifikace | |
| Doména: | Eukaryota |
| (bez hodnocení): | Unikonta |
| (bez hodnocení): | Obazoa |
| (bez hodnocení): | Opisthokonta |
| (bez hodnocení): | Holozoa |
| (bez hodnocení): | Filozoa |
| (bez hodnocení): | Choanozoa |
| Třída: | Choanoflagellatea Cavalier-Smith, 1998[2][3] |
| Zadejte druh | |
| Monosiga brevicollis[4] | |
| Objednávky a rodiny | |
sestra: Metazoa | |
| Synonyma | |
| |
The choanoflagellates jsou skupina volně žijících jednobuněčných a koloniálních bičíkatý eukaryoty považována za nejbližší žijící příbuzné zvířata. Choanoflagellates jsou límečky s límečkem, které mají vzájemně propojený nálevkovitý límec mikrovilli na základně bičíku. Choanoflageláty jsou schopné jak nepohlavní, tak sexuální reprodukce.[7] Mají výraznou buňku morfologie charakterizované vejcovitým nebo sférickým buněčným tělem 3–10µm v průměru s jedním apikálním bičíkem obklopeným límcem 30-40 mikroklků (viz obrázek). Pohyb bičíku vytváří vodní proudy, které mohou pohánět volné plavání choanoflagelátů přes vodní sloupec a pasti bakterie a detritus proti límci mikroklků, kde jsou tyto potraviny pohlceny. Toto krmení poskytuje kritické spojení v globálním měřítku uhlíkový cyklus, propojení trofické úrovně. Kromě svých kritických ekologických rolí jsou choanoflageláty obzvláště zajímavé pro evoluční biologové, kteří studují původ mnohobuněčnosti u zvířat. Jako nejbližší žijící příbuzní zvířat slouží choanoflageláty jako užitečný model pro rekonstrukce posledního jednobuněčného předka zvířat.
Etymologie
Choanoflagellate je hybridní slovo z řečtiny χοάνη khoánē význam "trychtýř"(kvůli tvaru límce) a latinského slova bičík.
Vzhled

Každý choanoflagellate má jeden bičík, obklopen prstenem z aktin - vyplněné výčnělky zvané mikroklky, tvořící válcovitý nebo kuželovitý límec (choanos v řečtině). Pohyb bičíku čerpá vodu přes límec a bakterie a detritus jsou zachyceny mikroklky a přijímány.[8] Vodní proudy generované bičík také tlačte volně plavecké buňky, jako v zvíře spermie. Naproti tomu většina ostatních bičíkovců je vytáhl jejich bičíky.
Kromě jediného apikálního bičíku obklopeného aktin -plněné mikrovilli který charakterizuje choanoflagellates, vnitřní organizaci organely v cytoplazma je konstantní.[9] Bičíkovec bazální tělo sedí u základny apikální bičík a druhý, ne bičíkový bazální tělo spočívá v pravém úhlu k bičíkové základně. The jádro zaujímá v buňce polohu od apikálu do středu a potravinové vakuoly jsou umístěny v bazální oblast cytoplazma.[9][10] Kromě toho je buněčné tělo mnoha choanoflagelátů obklopeno rozlišovacím znakem extracelulární matrix nebo periplast. Tyto buněčné kryty se velmi liší ve struktuře a složení a jsou používány taxonomy pro účely klasifikace. Mnoho choanoflagellates staví složité „domy“ ve tvaru koše, tzv lorica, z několika silikátových pásů slepených dohromady.[9] Funkční význam periplastu není znám, ale u přisedlých organismů se předpokládá, že napomáhá připojení k substrátu. U planktonických organismů se spekuluje, že periplast zvyšuje odpor, čímž působí proti síle generované bičíkem a zvyšuje účinnost krmení.[11]
Choanoflagellates jsou buď volně plavat ve vodním sloupci, nebo přisedlý, ulpívající na substrátu přímo nebo prostřednictvím periplastu nebo tenkého stopky.[12] Ačkoli se choanoflageláty považují za přísně volně žijící a heterotrofní, řada příbuzných choanoflagellate, jako jsou členové Ichthyosporea nebo Mesomycetozoa, následujte a parazitický nebo patogenní životní styl.[13] Životní historie choanoflagelátů jsou špatně pochopeny. Mnoho druhů je považováno za osamělé; zdá se však, že kolonialita ve skupině několikrát nezávisle vznikala a koloniální druhy si udržují osamělé stádium.[12]
Ekologie

Více než 125 existujících druhů choanoflagelátů[8] jsou známé, celosvětově distribuovány v námořní, brakický a sladkovodní prostředí od Arktidy po tropy, zabírající obě pelagický a bentický zóny. Ačkoli většina odběrů choanoflagelátů se objevila mezi 0 ma 25 m, byly získány z hloubky až 300 m ve volné vodě[14] a 100 m pod antarktickými ledovými příkrovy.[15] Předpokládá se, že existuje mnoho druhů kosmopolitní v globálním měřítku [např. Diaphanoeca grandis bylo nahlášeno z Severní Amerika, Evropa a Austrálie (OBIS)], zatímco u jiných druhů se uvádí, že mají omezené regionální distribuce.[16] Společně distribuované druhy choanoflagelátů mohou zaujímat zcela odlišná mikroprostředí, ale obecně je třeba ještě objasnit faktory, které ovlivňují distribuci a disperzi choanoflagelátů.
Počet druh, jako jsou ty v rod Proterospongie, forma jednoduchá kolonie,[8] planktonika shluky, které se podobají miniaturnímu shluku hroznů, ve kterém je každá buňka v kolonii bičována, nebo shluky buněk na jedné stopce.[9][17] V říjnu 2019 našli vědci nové chování choanoflagelátů v pásmu: zřejmě se mohou koordinovat a reagovat na světlo.[18]
Choanoflageláty se živí bakterie a propojit jinak nepřístupné formy uhlík na organismy vyšší v trofickém řetězci.[19] I dnes jsou důležité v uhlíkový cyklus a mikrobiální web s potravinami.[8] Existují určité důkazy o tom, že choanoflageláty také hodují na viry.[20]
Životní cyklus

Choanoflageláty rostou vegetativně, přičemž více druhů prochází podélným štěpením;[10] je však třeba ještě objasnit reprodukční životní cyklus choanoflagelátů. Dokument zveřejněný v srpnu 2017 ukázal, že změny prostředí, včetně přítomnosti určitých bakterií, vyvolávají rojení a následnou sexuální reprodukci choanoflagelátů.[7] The ploidie úroveň není známa;[22] nicméně objev retrotranspozonů a klíčových genů zapojených do meiózy[23] dříve navrhli, aby použili sexuální reprodukce jako součást jejich životního cyklu. Některé choanoflagellates mohou podstoupit encystment, což zahrnuje zatažení bičíku a límce a zapouzdření v elektronově husté fibrilární stěně. Při přenosu na čerstvé médium dochází k vylučování; ačkoli to zbývá přímo pozorovat.[24] Další vyšetření choanoflagelátu životní cyklus bude informativní o mechanismech tvorby kolonií a atributech přítomných před vývoj živočišné mnohobuněčnosti.
Důkazy o sexuální reprodukci byly hlášeny u druhů choanoflagellate Salpingoeca rosetta.[25][26] Byly hlášeny také důkazy o přítomnosti konzervovaných meiotické geny v choanoflagellates Monosiga brevicollis a Monosiga ovata.[27]
Biomineralizace křemíku
Acanthoecid choanoflagellates produkují extracelulární strukturu koše známou jako lorica. Lorica se skládá z jednotlivých pobřežních proužků vyrobených z biokompozitu oxidu křemičitého. Každý pobřežní proužek je vytvořen v buňce choanoflagelátu a poté je vylučován na povrch buňky. U nudiform choanoflagellates probíhá lorica montáž pomocí řady chapadel, jakmile bylo vyrobeno dostatečné množství pobřežních proužků, které by zahrnovaly celou lorica. U tektiformních choanoflagelátů se pobřežní proužky shromažďují v nastaveném uspořádání pod límcem. Během buněčného dělení nová buňka bere tyto pobřežní proužky jako součást cytokineze a sestavuje vlastní loricu pouze pomocí těchto dříve vyrobených pásů.[28]
Choanoflagellátová biosilicifikace vyžaduje koncentraci kyselina křemičitá uvnitř buňky. To se provádí pomocí proteinů Silicon Transporter (SIT). Analýza choanoflagellátových SIT ukazuje, že jsou podobné křemíkovým transportérům typu SIT rozsivky a další tváření křemene stramenopiles. Rodina genů SIT vykazuje malou nebo žádnou homologii s jinými geny, dokonce i s geny v nesilikátových choanoflagelátech nebo stramenopilech. To naznačuje, že rodina genů SIT se vyvinula prostřednictvím události postranního přenosu genů mezi Acanthoecids a Stramenopiles. Toto je pozoruhodný případ horizontální přenos genů mezi dvěma vzdáleně příbuznými eukaryotickými skupinami a poskytl vodítka pro biochemii a interakce křemíku a proteinu jedinečné rodiny genů SIT.[29]
Klasifikace
Vztah k metazoanům
Dujardin, francouzský biolog zajímající se o vývoj prvoků, zaznamenal morfologické podobnosti choanoflagelátů a houbových choanocytů a navrhl možnost blízkého vztahu již v roce 1841.[11] V posledním desetiletí byl tento předpokládaný vztah mezi choanoflageláty a zvířaty potvrzen nezávislými analýzami více nespojených sekvencí: 18S rDNA, geny kódující nukleární proteiny a mitochondriální genomy (Steenkamp a kol., 2006; Burger a kol., 2003;[13] Wainright a kol., 1993). Důležité je, že srovnání sekvencí mitochondriálního genomu z choanoflagelátu a tří hub potvrzuje umístění choanoflagelátů jako outgroup Metazoa a popírají možnost, že se choanoflageláty vyvinuly z metazoanů (Lavrov, et al., 2005). Konečně nedávné studie genů exprimovaných v choanoflagellatech odhalily, že choanoflageláty syntetizují homology metazoanových buněčných signálních a adhezivních genů.[30] (King, 2003) Sekvenování genomu ukazuje, že mezi živými organismy jsou choanoflageláty nejtěsněji příbuzné zvířatům.[8]Vzhledem k tomu, že choanoflageláty a metazoany spolu úzce souvisejí, slibují srovnání mezi oběma skupinami, že poskytnou vhled do biologie jejich posledního společného předka a prvních událostí v metazoan vývoj. The choanocyty (také známé jako "buňky s límečkem") houby (považovány za nejzákladnější metazoa) mají stejnou základní strukturu jako choanoflageláty. Límečkové buňky se nacházejí v jiných zvířecích skupinách, jako je např stužkové,[31] což naznačuje, že to byl morfologie jejich posledního společného předka. Poslední společný předek zvířat a choanoflagelátů byl jednobuněčný, snad tvořil jednoduché kolonie; na rozdíl od toho poslední společný předek všech eumetazoan zvířata byl mnohobuněčný organismus s diferencovanými tkáněmi, jednoznačným „tělesným plánem“ a embryonálním vývojem (včetně gastrulace).[8] Načasování rozdělení těchto linií je obtížné omezit, ale bylo to pravděpodobně v pozdním prekambrii,>600 před miliony let.[8]
Některé nedávné práce nepodporují sesterský vztah Choanoflagellates se zvířaty. Choanoflageláty mohou souviset s Ichthyosporea.[32]
Vnější vztahy Choanoflagellatea.[33]
| Opisthokonta |
| ||||||||||||||||||||||||||||||||||||
Fylogenetické vztahy
Choanoflageláty byly zahrnuty do Chrysophyceae až do Hibberda, 1975.[34] Nedávné molekulární fylogenetické rekonstrukce vnitřních vztahů choanoflagelátů umožňuje polarizaci vývoje postavy v kladu. Velké fragmenty atomu SSU a LSU ribozomální RNA, alfa tubulin, a protein tepelného šoku K vyřešení vnitřních vztahů a polarity znaků v choanoflagellate bylo použito 90 kódujících genů.[17] Každý ze čtyř genů vykazoval nezávisle podobné výsledky a analýzu kombinovaného souboru dat (zřetězené ) spolu se sekvencemi z jiných blízce příbuzných druhů (zvířata a houby ) ukazují, že choanoflageláty jsou silně podporovány jako monofyletický a potvrdit jejich pozici nejbližšího známého jednobuněčného žijícího příbuzného zvířat.
Dříve byla Choanoflagellida rozdělena do těchto tří rodin na základě složení a struktury jejich periplastů: Codonosigidae, Salpingoecidae a Acanthoecidae. Členům čeledi Codonosigidae se zdá, že postrádají periplast při zkoumání světelným mikroskopem, ale mohou mít jemnou vnější vrstvu viditelnou pouze elektronová mikroskopie. Rodina Salpingoecidae se skládá z druhů, jejichž buňky jsou uzavřeny v pevné tece, která je viditelná jak světelným, tak elektronovým mikroskopem. The theca je vylučovaná krytina převážně složená z celulóza nebo jiný polysacharidy.[35] O těchto divizích je nyní známo, že jsou paraphyletic, s konvergentním vývojem těchto forem rozšířeným. Třetí rodina choanoflagelátů, Acanthoecidae, byla podporována jako monofyletická skupina. Tento clade má a synapomorfie buněk nalezených v košovitém lorica, poskytujícím alternativní název „Loricate Choanoflagellates“. Acanthoecid lorica se skládá z řady křemičitý pobřežní pruhy uspořádané do druhově specifického lorica vzoru. “[9][11]
Choanoflagelátový strom založený na molekulární fylogenetice se dělí na tři dobře podporované klady.[17] Clade 1 a Clade 2 se skládají z kombinace druhů tradičně připisovaných Codonosigidae a Salpingoecidae, zatímco Clade 3 zahrnuje druhy ze skupiny taxonomicky klasifikované jako Acanthoecidae.[17] Mapování charakterových vlastností této fylogeneze naznačuje, že poslední společný předek choanoflagellates byl mořský organismus s diferencovaným životní cyklus s sedavý a pohyblivý etapy.[17]
Taxonomie
Choanoflageláty[3]
- Objednat Craspedida Cavalier-Smith 1997 em. Nitsche a kol. 2011
- Rodina Salpingoecidae Kent 1880-1882
- ?Dicraspedella Ellis 1930
- ?Diploeca Ellis 1930
- ?Diplosigopsis Francé 1897
- ?Pachysoeca Ellis 1930
- ?Piropsis Meunier 1910
- ?Salpingorhiza Klug 1936
- ?Sphaerodendron Zhukov, Mylnikov & Moiseev 1976 non Seemann 1865
- ?Stelexomonas Lackey 1942
- Astrosiga Kent 1880-1882
- Aulomonas Lackey 1942
- Choanoeca Ellis 1930
- Kladospongie Iyengar a Ramathan 1940
- Codonosigopsis Senn 1900
- Codosiga James-Clark 1866
- Diplosiga Frenzel 1891
- Desmarella Kent 1880-1882
- Hartaetosiga Carr, Richter & Nitsche 2017
- Kentrosiga Schiller 1953
- Monosiga Kent 1880-1882
- Mylnosiga Carr, Richter & Nitsche 2017
- Lagenoeca Kent 1881
- Microstomoeca Carr, Richter & Nitsche 2017
- Paramonosiga Jeuck, Arndt & Nitsche 2014
- Proterospongie Kent 1882
- Salpingoeca James-Clark 1868 non Ellis 1933
- Sphaeroeca Lauterborn 1894 non Meyrick 1895
- Stagondoeca Carr, Richter & Nitsche 2017
- Stylochromonas Lackey 1940
- Rodina Salpingoecidae Kent 1880-1882
- Objednat Acanthoecida Norris 1965 em. Nitsche a kol. 2011 (Laikát choanoflagellates)
- Conioeca Thomsen & Ostergaard 2019
- Rodina Acanthoecidae Norris 1965 em. Nitsche a kol. 2011 (Nudiform choanoflagellates)
- Acanthoeca Ellis 1930
- Enibas Schiwitza, Arndt & Nitsche 2019
- Helgoeca Leadbeater 2008
- Polyoeca Kent 1880
- Savillea Loeblich III 1967
- Rodina Stephanoecidae Leadbeater 2011 (Tectiform choanoflagellates)
- ?Conion Thomsen 1982
- ?Spiraloecion Marchant & Perrin 1986
- Acanthocorbis Hara & Takahashi 1984
- Amoenoscopa Hara & Takahashi 1987
- Apheloecion Thomsen 1983
- Bicosta Leadbeater 1978
- Calliacantha Leadbeater 1978
- Calotheca Thomsen & Moestrup 1983 non Desv. 1810 bez Spreng. 1817 non Heyden 1887
- Cosmoeca Thomsen 1984
- Crinolina Thomsen 1976 non Smetana 1982
- Crucispina Espeland & Throndsen 1986
- Diaphanoeca Ellis 1930
- Didymoeca Doweld 2003
- Kakoeca Buck & Marchant 1991
- Monocosta Thomsen 1979 non Monocostus Schumann 1904
- Nannoeca Thomsen 1988
- Parvicorbicula Deflandre 1960
- Pleurasiga Schiller 1925
- Polyfibula Manton 1981
- Saepicula Leadbeater 1980
- Saroca Thomsen 1979
- Spinoeca Thomsen, Ostergaard & Hansen 1995 non Poulsen 1973
- Stephanacantha Thomsen 1983
- Stephanoeca Ellis 1930
- Syndetophyllum Thomsen & Moestrup 1983
- Thomsenella Özdikmen 2009
Genomy a transkriptomy
Dva druhy choanoflagellate mají své genomy plně sekvenovány, další dva druhy měly přepis data zveřejněna.
Monosiga brevicollis genom
Genom Monosiga brevicollis, se 41,6 miliony párů bází,[8] má podobnou velikost jako vláknité houby a jiné volně žijící jednobuněčné eukaryoty, ale je mnohem menší než u typických zvířat.[8] V roce 2010 fylogenomická studie odhalila přítomnost několika genů řas v genomu Monosiga brevicollis. To by mohlo být způsobeno skutečností, že v rané evoluční historii choanoflageláty konzumovaly řasy jako potravu prostřednictvím fagocytóza.[36] Carr a kol. (2010)[27] promítl M. brevicollis genom pro známé eukaryotický geny meiózy. Z 19 známých testovaných eukaryotických meiotických genů (včetně 8, které fungují v žádném jiném procesu kromě meiózy), bylo 18 identifikováno M. brevicollis. Přítomnost meiotických genů, včetně genů specifických pro meiózu, naznačuje, že meióza, a implicitně, sex je přítomen v choanoflagellates.
Salpingoeca rosetta genom
Genom Salpingoeca rosetta má velikost 55 megabází.[37] V genomu jsou přítomny homology buněčné adheze, geny metabolismu neuropeptidů a glykosfingolipidů.S. rosetta má sexuální životní cyklus a přechody mezi ním haploidní a diploidní fáze.[26] V reakci na omezení živin haploidní kultury S. rosetta stát se diploidním. Tento posun ploidie se shoduje s pářením, během kterého se malé, bičíkaté buňky spojují s většími bičíkatými buňkami. Existují také důkazy o historickém páření a rekombinace v S. rosetta.
S. rosetta je indukován k sexuální reprodukci mořskou bakterií Vibrio fischeri.[25] Jediný V. fischeri protein, EroS plně rekapituluje živou aktivitu podobnou afrodiziaku V. fisheri.
Monosiga ovata přepis
Datová sada EST z Monosiga ovata byla zveřejněna v roce 2006.[38] Hlavním nálezem tohoto transkriptomu byla choanoflagellátová Hogletova doména a osvětlila roli přeskupování domén ve vývoji Ježková signální dráha. M. ovata má alespoň čtyři eukaryotické meiotické geny.[27]
Stephanoeca diplocostata přepis
První transkriptom loricate choanoflagellate[29] vedlo k objevu choanoflagelátových křemíkových transportérů. Následně byly podobné geny identifikovány u druhého druhu loricate, Diaphanoeca grandis. Analýza těchto genů zjistila, že choanoflagellátové SIT vykazují homologii s křemíkovými transportéry typu SIT rozsivky a vyvinuli se skrz horizontální přenos genů.
Galerie

Salpingoeca pod PCM

Salpingoeca sp. sekce pod TEM

Desmarella moniliformis kolonie pod PCM

Codosiga kolonie pod světelná mikroskopie

Sphaeroeca kolonie (přibližně 230 jedinců) pod světelnou mikroskopií.






Reference
- ^ Parfrey LW, Lahr DJ, Knoll AH, Katz LA (srpen 2011). „Odhad načasování časné eukaryotické diverzifikace pomocí multigenových molekulárních hodin“. Sborník Národní akademie věd Spojených států amerických. 108 (33): 13624–9. Bibcode:2011PNAS..10813624P. doi:10.1073 / pnas.1110633108. PMC 3158185. PMID 21810989.
- ^ Cavalier-Smith T (1998). „Neomonada a původ zvířat a hub.“. In Coombs GH, Vickerman K, Sleigh MA, Warren A (eds.). Evoluční vztahy mezi prvoky. Londýn: Kluwer. 375–407.
- ^ A b Leadbeater BS (2015). Choanoflageláty: evoluce, biologie a ekologie. University of Birmingham. ISBN 978-0-521-88444-0.
- ^ King N, Westbrook MJ, Young SL, Kuo A, Abedin M, Chapman J a kol. (Únor 2008). „Genom choanoflagellate Monosiga brevicollis a původ metazoanů“. Příroda. 451 (7180): 783–8. Bibcode:2008Natur.451..783K. doi:10.1038 / nature06617. PMC 2562698. PMID 18273011.
- ^ Saville-Kent, W. (1880). Manuál Infusoria. London, sv. 1, s. 324, [1].
- ^ Nitsche F, Carr M, Arndt H, Leadbeater BS (2011). "Vyšší úroveň taxonomie a molekulární fylogenetiky Choanoflagellatea". The Journal of Eukaryotic Microbiology. 58 (5): 452–62. doi:10.1111 / j.1550-7408.2011.00572.x. PMID 21895836.
- ^ A b „Bakteriální protein působí jako afrodiziakum pro choanoflageláty“.
- ^ A b C d E F G h i Král N „Westbrook MJ, Young SL, Kuo A, Abedin M, Chapman J a kol. (Únor 2008). „Genom choanoflagellate Monosiga brevicollis a původ metazoanů“. Příroda. 451 (7180): 783–8. Bibcode:2008Natur.451..783K. doi:10.1038 / nature06617. PMC 2562698. PMID 18273011.
- ^ A b C d E Leadbeater BS, Thomsen H (2000). „Objednat Choanoflagellida“. Ilustrovaný průvodce protozoa, druhé vydání. Lawrence: Společnost protozoologů. 451: 14–38.
- ^ A b Karpov S, Leadbeater BS (květen 1998). "Struktura a složení cytoskeletu v choanoflagelátech". Journal of Eukaryotic Microbiology. 45 (3): 361–367. doi:10.1111 / j.1550-7408.1998.tb04550.x.
- ^ A b C Leadbeater BS, Kelly M (2001). "Vývoj zvířat choanoflagellates a houby". Voda a atmosféra online. 9 (2): 9–11.
- ^ A b Leadbeater BS (únor 1983). „Životní historie a ultrastruktura nových mořských druhů Proterospongie (Choanoflagellida) ". J. Mar. Biol. Doc. SPOJENÉ KRÁLOVSTVÍ. 63 (1): 135–160. doi:10.1017 / S0025315400049857.
- ^ A b Mendoza L, Taylor JW, Ajello L (2002). „Třída mezomycetozoea: heterogenní skupina mikroorganismů na hranici mezi zvířaty a houbami“. Výroční přehled mikrobiologie. 56: 315–44. doi:10.1146 / annurev.micro.56.012302.160950. PMID 12142489. S2CID 14764188.
- ^ Thomsen H (1982). Planktonické choanoflageláty z Disko Bugtu v západním Grónsku s průzkumem mořského nanoplanktonu v této oblasti. Meddelelser om Gronland, Bioscience. 8. s. 3–63. ISBN 978-87-635-1149-0.
- ^ Buck KR, Garrison DL (červen 1988). „Distribuce a hojnost choanoflagelátů (Acanthoecidae) v zóně ledové hrany ve Weddellově moři v Antarktidě.“ Mar. Biol. 98 (2): 263–269. doi:10.1007 / BF00391204.
- ^ Thomsen H, Buck K, Chavez F (1991). „Choanoflagellates ve vodách střední Kalifornie: taxonomie, morfologie a seskupení druhů“. Ofélie. 33 (2): 131–164. doi:10.1080/00785326.1991.10429736.
- ^ A b C d E Carr M, Leadbeater BS, Hassan R, Nelson M, Baldauf SL (říjen 2008). „Molekulární fylogeneze choanoflagelátů, sesterská skupina Metazoa“. Sborník Národní akademie věd Spojených států amerických. 105 (43): 16641–6. Bibcode:2008PNAS..10516641C. doi:10.1073 / pnas.0801667105. PMC 2575473. PMID 18922774.
- ^ „Nově objevené mikroorganismy se spojují, vyklopte'". HHMI.org. Citováno 2019-10-29.
- ^ Butterfield NJ (1. dubna 1997). „Ekologie planktonu a přechod proterozoik-fanerozoik“. Paleobiologie. 23 (2): 247–262. doi:10.1017 / S009483730001681X.
- ^ Wu, Katherine J. (24. září 2020). „Nic nejí viry, je to tak? Seznamte se s některými hladovými protisty: Nové genetické důkazy staví případ, že jednobuněčné mořské mikroby mohou virům sekat“. The New York Times. str. D3 (29. září 2020, tisk). Citováno 1. prosince 2020.
- ^ De Loof, A. a Schoofs, L. (2019) „Způsob působení farnesolu,„ ušlechtilého “neznámého zejména v homeostáze Ca (2+) a jeho juvenilních esterů hormonů v evoluční retrospektivě“. Frontiers in Neuroscience, 13: 141. doi:10,3389 / fnins.2019.00141.
- ^ Claus Nielsen. Evoluce zvířat: Vzájemné vztahy živé fyly. 3. vyd. Claus Nielsen. Oxford, Velká Británie: Oxford University Press, 2012, s. 14.
- ^ Carr M, Leadbeater BS, Baldauf SL (2002). "Zachované meiotické geny poukazují na sex v choanoflagellates". The Journal of Eukaryotic Microbiology. 57 (1): 56–62. doi:10.1111 / j.1550-7408.2009.00450.x. PMID 20015185.
- ^ Leadbeater BS, Karpov SA (září – říjen 2000). „Tvorba cyst ve sladkovodním kmeni choanoflagellate Desmarella moniliformis Kent“. The Journal of Eukaryotic Microbiology. 47 (5): 433–9. doi:10.1111 / j.1550-7408.2000.tb00071.x. PMID 11001139.
- ^ A b Woznica A, Gerdt JP, Hulett RE, Clardy J, King N (září 2017). „Páření v nejbližším příbuzném zvířat je vyvoláno bakteriální chondroitinázou“. Buňka. 170 (6): 1175–1183.e11. doi:10.1016 / j.cell.2017.08.005. PMC 5599222. PMID 28867285.
- ^ A b Levin TC, král N (listopad 2013). „Důkazy o sexu a rekombinaci v choanoflagellate Salpingoeca rosetta“. Aktuální biologie. 23 (21): 2176–80. doi:10.1016 / j.cub.2013.08.061. PMC 3909816. PMID 24139741.
- ^ A b C Carr M, Leadbeater BS, Baldauf SL (2010). "Zachované meiotické geny poukazují na sex v choanoflagellates". The Journal of Eukaryotic Microbiology. 57 (1): 56–62. doi:10.1111 / j.1550-7408.2009.00450.x. PMID 20015185.
- ^ Leadbeater BS, Yu Q, Kent J, Stekel DJ (leden 2009). „Trojrozměrné obrazy choanoflagellate loricae“. Řízení. Biologické vědy. 276 (1654): 3–11. doi:10.1098 / rspb.2008.0844. PMC 2581655. PMID 18755674.
- ^ A b Marron AO, Alston MJ, Heavens D, Akam M, Caccamo M, Holland PW, Walker G (duben 2013). „Rodina křemíkových transportérů křemeliny v křemičitých loriátových choanoflagelátech“. Řízení. Biologické vědy. 280 (1756): 20122543. doi:10.1098 / rspb.2012.2543. PMC 3574361. PMID 23407828.
- ^ Král N, Carroll SB (Prosinec 2001). „Receptorová tyrosinkináza z choanoflagelátů: molekulární pohledy na časný vývoj zvířat“. Sborník Národní akademie věd Spojených států amerických. 98 (26): 15032–7. Bibcode:2001PNAS ... 9815032K. doi:10.1073 / pnas.261477698. PMC 64978. PMID 11752452.
- ^ Cantell C, Franzén Å, Sensenbaugh T (říjen 1982). "Ultrastruktura multicilárních límcových buněk v pilidiové larvě Lineus bilineatus (Nemertini) ". Zoomorfologie. 101 (1): 1–15. doi:10.1007 / BF00312027.
- ^ Corsaro D, Walochnik J, Venditti D, Müller KD, Hauröder B, Michel R (prosinec 2014). „Znovuobjevení Nucleophaga amoebae, nového člena Rozellomycota“. Parazitologický výzkum. 113 (12): 4491–8. doi:10.1007 / s00436-014-4138-8. PMID 25258042.
- ^ Torruella, Guifré; Mendoza, Alex de; Grau-Bové, Xavier; Antó, Meritxell; Chaplin, Mark A .; Campo, Javier del; Eme, Laura; Pérez-Cordón, Gregorio; Whipps, Christopher M. (2015). „Fylogenomika odhaluje konvergentní vývoj životního stylu u blízkých příbuzných zvířat a hub“. Aktuální biologie. 25 (18): 2404–2410. doi:10.1016 / j.cub.2015.07.053. PMID 26365255.
- ^ Reviers, B. de. (2006). Biologia e Filogenia das Algas. Editora Artmed, Porto Alegre, str. 156.
- ^ (Adl a kol., 2005)
- ^ Sun G, Yang Z, Ishwar A, Huang J (prosinec 2010). "Řasy geny v nejbližších příbuzných zvířat". Molekulární biologie a evoluce. 27 (12): 2879–89. doi:10.1093 / molbev / msq175. PMID 20627874.
- ^ Fairclough SR, Chen Z, Kramer E, Zeng Q, Young S, Robertson HM, Begovic E, Richter DJ, Russ C, Westbrook MJ, Manning G, Lang BF, Haas B, Nusbaum C, King N (únor 2013). „Vývoj premetazoanového genomu a regulace buněčné diferenciace v choanoflagellate Salpingoeca rosetta“. Genome Biology. 14 (2): R15. doi:10.1186 / gb-2013-14-2-r15. PMC 4054682. PMID 23419129.
- ^ Snell EA, Brooke NM, Taylor WR, Casane D, Philippe H, Holland PW (Únor 2006). „Neobvyklý choanoflagelátový protein uvolněný autokatalytickým zpracováním Hedgehog“. Řízení. Biologické vědy. 273 (1585): 401–7. doi:10.1098 / rspb.2005.3263. PMC 1560198. PMID 16615205.
externí odkazy
- ChoanoWiki zdroj pro spolupráci udržovaný výzkumnou komunitou Choanoflagellate
- Webová stránka Tree of Life pro choanoflagellates
- Monosiga brevicollis prohlížeč genomu
- Vaše mozková chemie existovala dříve, než zvířata
- Choanobase, genetická knihovna Choanoflagellate, vyvinutá a udržovaná Nicole King laboratoř v University of California, Berkeley
