Mnohobuněčný organismus - Multicellular organism

Mnohobuněčné organismy jsou organismy které se skládají z více než jednoho buňka, na rozdíl od jednobuněčné organismy.[1]
Všechny druhy zvířata, suchozemské rostliny a většina houby jsou mnohobuněčné, stejně jako mnoho jiných řasy, zatímco několik organismů je částečně jednobuněčných a částečně mnohobuněčných slizové formy a sociální améby, jako je rod Dictyostelium.[2][3]
Mnohobuněčné organismy vznikají různými způsoby, například buněčné dělení nebo agregací mnoha jednotlivých buněk.[4][3] Koloniální organismy jsou výsledkem mnoha identických jedinců, kteří se spojili a vytvořili a kolonie. Často však může být obtížné oddělit koloniální protisty od pravých mnohobuněčných organismů, protože tyto dva pojmy nejsou odlišné; koloniální protisté dostali přezdívku „vícebuněčný“ spíše než „mnohobuněčný“.[5][6]
Evoluční historie
Výskyt
Mnohobuněčnost se u eukaryot vyvinula nezávisle nejméně 25krát,[7][8] a také v některých prokaryoty, jako sinice, myxobakterie, aktinomycety, Magnetoglobus multicellularis nebo Methanosarcina.[3] Složité mnohobuněčné organismy se však vyvinuly pouze v šesti eukaryotický skupiny: zvířata, houby, hnědé řasy, červené řasy, zelené řasy, a suchozemské rostliny.[9] Vyvíjelo se to opakovaně Chloroplastida (zelené řasy a suchozemské rostliny), jednou nebo dvakrát pro zvířata, jednou pro hnědé řasy, třikrát v houby (chytrids, ascomycetes a bazidiomycety )[10] a možná několikrát za slizové formy a červené řasy.[11] První důkaz mnohobuněčnosti je z sinice - jako organismy, které žily před 3–3,5 miliardami let.[7] Pro reprodukci musí opravdové mnohobuněčné organismy vyřešit problém regenerace celého organismu zárodečné buňky (tj., spermie a vejce buněk), problém, který je studován v evoluční vývojová biologie. U zvířat se vyvinula značná rozmanitost typy buněk v mnohobuněčném těle (100–150 různých typů buněk), ve srovnání s 10–20 v rostlinách a houbách.[12]
Ztráta mnohobuněčnosti
U některých skupin došlo ke ztrátě mnohobuněčnosti.[13] Houby jsou převážně mnohobuněčné, ačkoli časně odlišné linie jsou převážně jednobuněčné (např. Microsporidia ) a došlo k četným obrácením jednobuněčnosti mezi houbami (např. Saccharomycotina, Cryptococcus, a další droždí ).[14][15] Může se také vyskytnout u některých červené řasy (např., Porfyridium ), ale je možné, že jsou primitivně jednobuněčné.[16] Ztráta mnohobuněčnosti je také u některých považována za pravděpodobnou zelené řasy (např., Chlorella vulgaris a nějaký Ulvophyceae ).[17][18] U jiných skupin, obecně parazitů, došlo ke snížení mnohobuněčnosti v počtu nebo typech buněk (např myxozoans mnohobuněčné organismy, o nichž se dříve myslelo, že jsou jednobuněčné, jsou pravděpodobně extrémně omezené cnidarians ).[19]
Rakovina
Mnohobuněčné organismy, zejména dlouhověká zvířata, čelí výzvě rakovina, ke kterému dochází, když buňky nedokážou regulovat svůj růst v rámci normálního vývojového programu. Během tohoto procesu lze pozorovat změny morfologie tkáně. Rakovina u zvířat (metazoans ) byl často popisován jako ztráta mnohobuněčnosti.[20] Diskutuje se o možnosti existence rakoviny u jiných mnohobuněčných organismů[21][22] nebo dokonce u prvoků.[23] Například, rostlinné hálky byly charakterizovány jako nádory,[24] ale někteří autoři tvrdí, že u rostlin se nevyvíjí rakovina.[25]
Separace somatických a zárodečných buněk
V některých mnohobuněčných skupinách, které se nazývají Weismannists, oddělení mezi sterilním somatická buňka linka a zárodečná buňka linka se vyvinula. Weismannistický vývoj je však relativně vzácný (např. Obratlovci, členovci, Volvox ), protože velká část druhů má kapacitu pro somatická embryogeneze (např. suchozemské rostliny, většina řas, mnoho bezobratlých).[26][27]
Hypotézy o původu

Jedna hypotéza o původu mnohobuněčnosti spočívá v tom, že skupina funkčně specifických buněk agregovaná do hlemýžďové hmoty zvané a grex, která se pohybovala jako mnohobunková jednotka. To je v podstatě co slizové formy dělat. Další hypotézou je, že primitivní buňka podstoupila dělení jádra, čímž se stala a koenocyt. Poté by se kolem každého jádra (a buněčného prostoru a organel zabraných v prostoru) vytvořila membrána, což by vedlo ke skupině spojených buněk v jednom organismu (tento mechanismus je pozorovatelný v Drosophila ). Třetí hypotéza spočívá v tom, že když se jednobuněčný organismus rozdělil, dceřiné buňky se neoddělily, což mělo za následek konglomeraci identických buněk v jednom organismu, který by později mohl vyvinout specializované tkáně. To je to, co rostlina a zvíře embrya stejně dobře jako koloniální choanoflagellates.[28][29]
Protože první mnohobuněčné organismy byly jednoduché, měkké organismy postrádající kost, skořápku nebo jiné tvrdé části těla, nejsou ve fosilním záznamu dobře zachovány.[30] Jedinou výjimkou může být demosponge, které ve starých horninách mohly zanechat chemický podpis. K nejranějším fosiliím mnohobuněčných organismů patří napadené Grypania spiralis a fosilie černých břidlic v Paleoproterozoikum Fosilní skupina Francevillian Formace B v Gabon (Gabonionta ).[31] The Formace Doushantuo přineslo 600 milionů let staré mikrofosílie s důkazem mnohobuněčných vlastností.[32]
Až do nedávné doby, fylogenetické rekonstrukce prošla anatomický (zejména embryologické ) podobnosti. To je nepřesné, protože živé mnohobuněčné organismy, jako je zvířata a rostliny jsou více než 500 milionů let odstraněny z jejich jednobuněčných předků. Takový čas umožňuje obojí odlišný a konvergentní evoluční čas napodobovat podobnosti a hromadit rozdíly mezi skupinami moderních a vyhynulých rodových druhů. Moderní fylogenetika využívá sofistikované techniky, jako je aloenzymy, satelitní DNA a další molekulární markery popisující vlastnosti, které jsou sdíleny mezi vzdáleně příbuznými liniemi.[Citace je zapotřebí ]
K vývoji mnohobuněčnosti mohlo dojít mnoha různými způsoby, z nichž některé jsou popsány níže:
Symbiotická teorie
Tato teorie naznačuje, že první mnohobuněčné organismy se vyskytly od symbióza (spolupráce) různých druhů jednobuněčných organismů, z nichž každý má různé role. Postupem času se tyto organismy staly natolik závislými na sobě, že by nebyly schopny samostatně přežít, což by nakonec vedlo k začlenění jejich genomů do jednoho mnohobuněčného organismu.[33] Každý příslušný organismus by se stal samostatnou linií diferencovaných buněk v nově vytvořeném druhu.
Tento druh silně závislé symbiózy lze často vidět, například ve vztahu mezi klaun ryby a Riterri sasanky. V těchto případech je extrémně pochybné, zda by jeden z druhů přežil velmi dlouho, kdyby druhý vyhynul. Problém této teorie však spočívá v tom, že stále není známo, jak by mohla být DNA každého organismu začleněna do jednoho genom představovat je jako jediný druh. Ačkoli se předpokládá, že k takové symbióze došlo (např. mitochondrie a chloroplasty v živočišných a rostlinných buňkách—endosymbióza ), stalo se to jen extrémně zřídka ai tehdy si genomy endosymbiontů zachovaly rozlišovací prvek, který samostatně replikoval svoji DNA během mitóza hostitelského druhu. Například dva nebo tři symbiotické organismy tvořící kompozit lišejník, i když jsou na sobě navzájem závislí na přežití, musí se samostatně rozmnožovat a poté znovu formovat, aby znovu vytvořili jeden individuální organismus.
Cellularizační (syncyciální) teorie
Tato teorie uvádí, že jediný jednobuněčný organismus s více jádra, se mohly vyvinout vnitřní membrána oddíly kolem každého z jeho jader.[34] Mnoho protistů, jako je náčelníci nebo slizové formy může mít několik jader, což mu poskytuje podporu hypotéza. Jednoduchá přítomnost více jader však na podporu teorie nestačí. Několik jader nálevníků je odlišných a má jasné diferencované funkce. The makronukleus slouží potřebám organismu, zatímco mikrojader se používá k sexuální reprodukci s výměnou genetického materiálu. Slizové formy syncitia se tvoří z jednotlivých améboidních buněk, jako jsou syncitiální tkáně některých mnohobuněčných organismů, nikoli naopak. Aby byla tato teorie považována za platnou, potřebuje prokazatelný příklad a mechanismus generování mnohobuněčného organismu z již existujícího syncytia.
Koloniální teorie
Koloniální teorie Haeckel, 1874 navrhuje symbiózu mnoha organismů stejného druhu (na rozdíl od symbiotická teorie, což naznačuje symbiózu různých druhů) vedlo k mnohobuněčnému organismu. Přinejmenším u některých se předpokládá, že se vyvinuly na pevnině, vícebuněčnost nastává oddělením a následným připojením buněk (např. buněčné slizové formy ) vzhledem k tomu, že u většiny mnohobuněčných typů (těch, které se vyvinuly ve vodním prostředí) dochází k mnohobuněčnosti jako důsledek toho, že se buňky po dělení nerozdělí.[35] Mechanismus této druhé tvorby kolonií může být stejně jednoduchý jako neúplný cytokineze, ačkoli mnohobuněčnost se také obvykle považuje za zahrnutou buněčná diferenciace.[36]
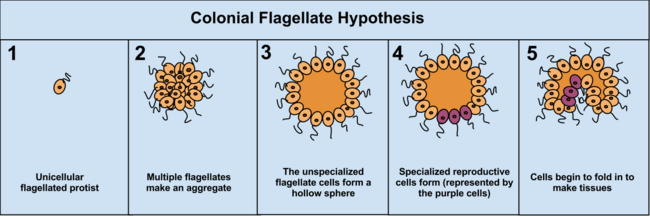
Výhodou hypotézy Colonial Theory hypotéza je, že bylo pozorováno, že se vyskytuje nezávisle u 16 různých phyla protoctistan. Například během nedostatku potravy améba Dictyostelium seskupuje se v kolonii, která se jako jedna přesune na nové místo. Některé z těchto améb se poté navzájem mírně odlišují. Další příklady koloniální organizace v protista jsou Volvocaceae, jako Eudorina a Volvox, z nichž druhá se skládá až z 500–50 000 buněk (v závislosti na druhu), z nichž se rozmnožuje pouze zlomek.[37] Například u jednoho druhu se množí 25–35 buněk, 8 nepohlavně a přibližně 15–25 sexuálně. Často však může být těžké oddělit koloniální protistů od skutečných mnohobuněčných organismů, protože tyto dva pojmy se neliší; koloniální protisté dostali přezdívku „vícebuněčný“ spíše než „mnohobuněčný“.[5]
Teorie synzoospor
Někteří autoři naznačují, že původ mnohobuněčnosti, přinejmenším v Metazoa, nastal v důsledku přechodu od časové k prostorové diferenciace buněk, spíše než postupným vývojem buněčné diferenciace, jak je potvrzeno v Haeckel Je Teorie gastraea.[38]
GK-PID
Asi před 800 miliony let,[39] malá genetická změna v jedné molekule zvané guanylát kináza doména interakce proteinů (GK-PID) mohla umožnit organismům přejít z jednoho buněčného organismu do jedné z mnoha buněk.[40]
Role virů
Geny vypůjčené od viry a mobilní genetické prvky (MGE) byly nedávno identifikovány jako látky hrající klíčovou roli v diferenciaci mnohobuněčných tkání a orgánů a dokonce i v sexuální reprodukci, ve fúzi vaječných buněk a spermií.[41][42]Takto fúzované buňky se také podílejí na metazoanových membránách, jako jsou ty, které zabraňují chemickým látkám pronikat přes placenta a oddělení mozkových těl.[41] Byly identifikovány dvě virové složky. První je syncytin, který pochází z viru.[43]Druhý identifikovaný v roce 2007 se nazývá EFF1, který pomáhá formovat pokožku Caenorhabditis elegans, součást celé rodiny FF proteinů. Felix Rey z Pasteurova institutu v Paříži zkonstruoval 3D strukturu proteinu EFF1[44] a ukázal, že funguje při spojování jedné buňky s druhou při virových infekcích. Skutečnost, že všechny známé buněčné fúzní molekuly mají virový původ, naznačuje, že byly životně důležité pro mezibuněčné komunikační systémy, které umožňovaly mnohobuněčnost. Bez schopnosti buněčné fúze by se mohly tvořit kolonie, ale nebylo by možné nic tak složitého jako houba.[45]
Hypotéza dostupnosti kyslíku
Tato teorie naznačuje, že kyslík dostupný v atmosféře rané Země mohl být limitujícím faktorem pro vznik mnohobuněčného života.[46] Tato hypotéza je založena na korelaci mezi vznikem mnohobuněčného života a zvýšením hladin kyslíku během této doby. K tomu by došlo po Skvělá oxidační událost (GOE), ale před posledním zvýšením kyslíku. Mlýny[47] dochází k závěru, že množství kyslíku přítomného během Ediacaran není nezbytný pro složitý život, a proto je nepravděpodobné, že by byl hnacím faktorem pro vznik mnohobuněčnosti.
Hypotéza Země sněhové koule
A sněhová koule Země je geologická událost, kdy je celý povrch Země pokryt sněhem a ledem. Poslední sněhová koule Země se odehrála během Kryogenní období a sestával ze dvou globálních událostí zalednění známých jako Sturtian a Marinoan zalednění. Xiao[48] naznačuje, že mezi obdobím známým jako „Nudná miliarda „a Země sněhové koule, jednoduchý život mohl mít čas na inovace a vývoj, který by později mohl vést k vývoji mnohobuněčnosti. Hypotéza Země sněhové koule v souvislosti s mnohobuněčností naznačuje, že cyrogénské období v historii Země mohlo být katalyzátorem evoluce komplexního mnohobuněčného života. Brocks[49] naznačuje, že doba mezi sturtianským glaciánem a novějším marinoanským glaciánem umožnila, aby v mořích dominovaly planktonické řasy, což umožnilo rychlou rozmanitost života rostlinných i živočišných linií. Krátce po marinoanu se rychle objevil složitý život, který se diverzifikoval v tzv Kambrijská exploze.
Hypotéza predace
Hypotéza predace naznačuje, že aby se jedlíci vyhnuli konzumaci, vyvinuli jednoduché jednobuněčné organismy mnohobuněčnost, aby bylo obtížnější být konzumovány jako kořist. Herron a kol [50] provedl laboratorní evoluční experimenty na jednobuněčné zelené řase, C. reinhardtii, používající paramecium jako predátora. Zjistili, že v přítomnosti tohoto predátora C. reinhardtii skutečně vyvíjí jednoduché mnohobunkové funkce.
Výhody
Mnohobuněčnost umožňuje organismu překročit limity velikosti, které normálně stanoví difúze: jednotlivé buňky se zvětšenou velikostí mají snížený poměr povrchu k objemu a mají potíže s absorpcí dostatečného množství živin a jejich transportem po celé buňce. Mnohobuněčné organismy tedy mají konkurenční výhody zvětšení velikosti bez jejích omezení. Mohou mít delší životnost, protože mohou pokračovat v životě, když jednotlivé buňky zemřou. Mnohobuněčnost také umožňuje zvýšení složitosti povolením diferenciace typů buněk v jednom organismu.
Zda je to však možné považovat za výhody, je diskutabilní. Drtivá většina živých organismů je jednobuněčná ai z hlediska biomasy jsou jednobuněčné organismy mnohem úspěšnější než zvířata, i když ne rostliny.[51] Spíše než vidět vlastnosti jako delší životnost a větší velikost jako výhodu, mnoho biologů je vidí jen jako příklady rozmanitosti s přidruženými kompromisy.
Viz také
Reference
- ^ Becker, Wayne M .; et al. (2008). Svět buňky. Pearson Benjamin Cummings. str. 480. ISBN 978-0-321-55418-5.
- ^ Chimileski, Scott; Kolter, Roberto (2017). Život na hranici zraku: Fotografický průzkum mikrobiálního světa. Harvard University Press. ISBN 9780674975910.
- ^ A b C Lyons, Nicholas A .; Kolter, Roberto (duben 2015). „O vývoji bakteriální mnohobuněčnosti“. Současný názor v mikrobiologii. 24: 21–28. doi:10.1016 / j.mib.2014.12.007. ISSN 1879-0364. PMC 4380822. PMID 25597443.
- ^ S. M. Miller (2010). „Volvox, Chlamydomonas a vývoj mnohobuněčnosti“. Přírodní výchova. 3 (9): 65.
- ^ A b Brian Keith Hall; Benedikt Hallgrímsson; Monroe W. Strickberger (2008). Strickbergerova evoluce: integrace genů, organismů a populací (4. vydání). Hall / Hallgrímsson. str.149. ISBN 978-0-7637-0066-9.
- ^ Adl, Sina; et al. (Říjen 2005). „Nová vyšší klasifikace eukaryot s důrazem na taxonomii protistů“. J. Eukaryot. Microbiol. 52 (5): 399–451. doi:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- ^ A b Grosberg, RK; Strathmann, RR (2007). „Vývoj mnohobuněčnosti: Menší zásadní přechod?“ (PDF). Annu Rev Ecol Evol Syst. 38: 621–654. doi:10.1146 / annurev.ecolsys.36.102403.114735.
- ^ Parfrey, L.W .; Lahr, D.J.G. (2013). „Mnohobuněčnost vznikala několikrát ve vývoji eukaryot“ (PDF). BioEssays. 35 (4): 339–347. doi:10.1002 / bies.201200143. PMID 23315654. S2CID 13872783.
- ^ http://public.wsu.edu/~lange-m/Documnets/Teaching2011/Popper2011.pdf
- ^ Niklas, KJ (2014). „Evoluční a vývojové počátky mnohobuněčnosti“. Dopoledne. J. Bot. 101 (1): 6–25. doi:10,3732 / ajb.1300314. PMID 24363320.
- ^ Bonner, John Tyler (1998). „Počátky mnohobuněčnosti“ (PDF). Integrativní biologie. 1 (1): 27–36. doi:10.1002 / (SICI) 1520-6602 (1998) 1: 1 <27 :: AID-INBI4> 3.0.CO; 2-6. ISSN 1093-4391. Archivovány od originálu 8. března 2012.CS1 maint: unfit url (odkaz)
- ^ Margulis, L. & Chapman, M.J. (2009). Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth ([4th ed.]. Ed.). Amsterdam: Academic Press / Elsevier. str. 116.
- ^ Seravin L. N. (2001) Princip protisměrné morfologické revoluce a jeho význam pro konstrukci megasystému protistů a dalších eukaryot. Protistologie 2: 6–14, [1].
- ^ Parfrey, L.W. & Lahr, D.J.G. (2013), s. 344.
- ^ Medina, M .; Collins, A. G .; Taylor, J. W .; Valentine, J. W .; Lipps, J. H .; Zettler, L. A. Amaral; Sogin, M. L. (2003). „Fylogeneze opistokononty a vývoj mnohobuněčnosti a složitosti hub a metazoí“. International Journal of Astrobiology. 2 (3): 203–211. Bibcode:2003IJAsB ... 2..203M. doi:10.1017 / s1473550403001551.
- ^ Seckbach, Joseph, Chapman, David J. [eds.]. (2010). Červené řasy v genomickém věku. New York, NY, USA: Springer, s. 252, [2].
- ^ Cocquyt, E .; Verbruggen, H .; Leliaert, F .; De Clerck, O. (2010). „Evoluce a cytologická diverzifikace zelených mořských řas (Ulvophyceae)“. Mol. Biol. Evol. 27 (9): 2052–2061. doi:10,1093 / molbev / msq091. ISSN 0737-4038. PMID 20368268.
- ^ Richter, Daniel Joseph: Obsah genů různých choanoflagelátů osvětluje zvířecí původ, 2013.
- ^ "Myxozoa". tolweb.org. Citováno 14. dubna 2018.
- ^ Davies, P. C. W .; Lineweaver, C. H. (2011). „Rakovinové nádory jako Metazoa 1.0: klepání na geny dávných předků“. Fyzikální biologie. 8 (1): 015001. Bibcode:2011PhBio ... 8a5001D. doi:10.1088/1478-3975/8/1/015001. PMC 3148211. PMID 21301065.
- ^ Richter, D. J. (2013), str. 11.
- ^ Gaspar, T .; Hagege, D .; Kevers, C .; Penel, C .; Crèvecoeur, M .; Engelmann, I .; Greppin, H .; Foidart, J. M. (1991). „Když se rostlinné teratomy za nepřítomnosti patogenů změní na rakovinu“. Physiologia Plantarum. 83 (4): 696–701. doi:10.1111 / j.1399-3054.1991.tb02489.x.
- ^ Lauckner, G. (1980). Nemoci prvoků. V: Nemoci mořských živočichů. Kinne, O. (ed.). Sv. 1, s. 84, John Wiley & Sons, Chichester, Velká Británie.
- ^ Riker, A. J. (1958). "Rostlinné nádory: Úvod". Sborník Národní akademie věd Spojených států amerických. 44 (4): 338–9. Bibcode:1958PNAS ... 44..338R. doi:10.1073 / pnas.44.4.338. PMC 335422. PMID 16590201.
- ^ Doonan, J .; Hunt, T. (1996). „Buněčný cyklus. Proč rostliny nedostanou rakovinu?“. Příroda. 380 (6574): 481–2. doi:10.1038 / 380481a0. PMID 8606760. S2CID 4318184.
- ^ Ridley M (2004) Evolution, 3. vydání. Blackwell Publishing, str. 295-297.
- ^ Niklas, K. J. (2014) Evoluční a vývojové počátky mnohobuněčnosti.
- ^ Fairclough, Stephen R .; Dayel, Mark J .; King, Nicole (26. října 2010). „Mnohobuněčný vývoj v choanoflagelátu“. Aktuální biologie. 20 (20): R875 – R876. doi:10.1016 / j.cub.2010.09.014. PMC 2978077. PMID 20971426. Citováno 14. dubna 2018.
- ^ V jednobuněčném dravci se dozvíte o narození zvířecího království
- ^ A Knoll, 2003. Život na mladé planetě. Princeton University Press. ISBN 0-691-00978-3 (tvrdý obal), ISBN 0-691-12029-3 (brožura). Vynikající kniha o rané historii života, velmi přístupná pro neprofesionála; zahrnuje rozsáhlé diskuse o časných podpisech, fosilizaci a organizaci života.
- ^ El Albani, Abderrazak; et al. (1. července 2010). „Velké koloniální organismy s koordinovaným růstem v okysličeném prostředí před 2,1 rokem“. Příroda. 466 (7302): 100–104. Bibcode:2010Natur.466..100A. doi:10.1038 / nature09166. ISSN 0028-0836. PMID 20596019. S2CID 4331375.
- ^ Chen, L .; Xiao, S .; Pang, K .; Zhou, C .; Yuan, X. (2014). „Diferenciace buněk a separace zárodků-soma u fosilií podobných zvířatům Ediacaran“. Příroda. 516 (7530): 238–241. Bibcode:2014 Natur.516..238C. doi:10.1038 / příroda13666. PMID 25252979. S2CID 4448316.
- ^ Margulis, Lynn (1998). Symbiotická planeta: Nový pohled na evoluci. New York: Základní knihy. str. 160. ISBN 978-0-465-07272-9.
- ^ Hickman CP, Hickman FM (8. července 1974). Integrované principy zoologie (5. vydání). Mosby. str. 112. ISBN 978-0-8016-2184-0.
- ^ Wolpert, L .; Szathmáry, E. (2002). "Mnohobuněčnost: Evoluce a vejce". Příroda. 420 (6917): 745. Bibcode:2002 Natur.420..745W. doi:10.1038 / 420745a. PMID 12490925. S2CID 4385008.
- ^ Kirk, D. L. (2005). „Dvanáctikrokový program pro rozvoj mnohobuněčnosti a dělby práce“. BioEssays. 27 (3): 299–310. doi:10.1002 / bies.20197. PMID 15714559.
- ^ AlgaeBase. Volvox Linnaeus, 1758: 820.
- ^ Mikhailov K. V., Konstantinova A. V., Nikitin M. A., Troshin P. V., Rusin L., Lyubetsky V., Panchin Y., Mylnikov A. P., Moroz L. L., Kumar S. & Aleoshin V. V. (2009). Původ Metazoa: přechod od časové diferenciace k diferenciaci prostorových buněk. Biologické testy, 31(7), 758–768, [3] Archivováno 2016-03-05 na Wayback Machine.
- ^ Erwin, Douglas H. (9. listopadu 2015). „Časný život metazoanů: divergence, životní prostředí a ekologie“. Phil. Trans. R. Soc. B. 370 (20150036): 20150036. doi:10.1098 / rstb.2015.0036. PMC 4650120. PMID 26554036.
- ^ Zimmer, Carle (7. ledna 2016). „Genetické převrácení pomohlo organizmům přejít z jedné buňky do mnoha“. New York Times. Citováno 7. ledna 2016.
- ^ A b Eugene V. Koonin: Viry a mobilní prvky jako hnací síly evolučních přechodů. In: Philos Trans R Soc Lond B Biol Sci., 2016 19. srpna, doi: 10.1098 / rstb.2015.0442
- ^ Rafi Letzter: Starověký virus může být odpovědný za lidské vědomí, in: Live Science, 2. února 2018
- ^ Mi S1, Lee X, Li X, Veldman GM, Finnerty H, Racie L, LaVallie E, Tang XY, Edouard P, Howes S, Keith JC Jr, McCoy JM .: Syncytin je retrovirový obalový protein v zajetí podílející se na morfogenezi lidské placenty. V přírodě. 2000 17. února; 403 (6771): 785-789. doi: 10.1038 / 35001608, PMID 10693809
- ^ Jamin, M, H Raveh-Barak, B Podbilewicz, FA Rey et al. (2014) „Structural base of eukaryotic cell-cell fusion“ (Cell, svazek 157, číslo 2, 10. dubna 2014), strany 407–419, doi: 10.1016 / j.cell.2014.02.020
- ^ Slezak, Michael (2016), „Žádné viry? Ani kůže, ani kosti“ (New Scientist, No. 2958, 1. března 2014), s. 16
- ^ Nursall, J. R. (duben 1959). „Kyslík jako předpoklad původu metazoí“. Příroda. 183 (4669): 1170–1172. Bibcode:1959 Natur.183.1170N. doi:10.1038 / 1831170b0. ISSN 1476-4687. S2CID 4200584.
- ^ Mills, D. B .; Ward, L. M .; Jones, C .; Sweeten, B .; Forth, M .; Treusch, A. H .; Canfield, D. E. (2014-02-18). „Požadavky na kyslík u prvních zvířat“. Sborník Národní akademie věd. 111 (11): 4168–4172. Bibcode:2014PNAS..111.4168M. doi:10.1073 / pnas.1400547111. ISSN 0027-8424. PMC 3964089. PMID 24550467.
- ^ Lyons, Timothy W .; Droser, Mary L .; Lau, Kimberly V .; Porter, Susannah M .; Xiao, Shuhai; Tang, Qing (2018-09-28). „Po nudné miliardě a před mrazivými miliony: evoluční vzorce a inovace v toniánském období“. Nová témata v biologických vědách. 2 (2): 161–171. doi:10.1042 / ETLS20170165. hdl:10919/86820. ISSN 2397-8554. PMID 32412616.
- ^ Brocks, Jochen J .; Jarrett, Amber J. M .; Sirantoine, Eva; Hallmann, Christian; Hoshino, Yosuke; Liyanage, Tharika (srpen 2017). „Vzestup řas v kryogenních oceánech a výskyt zvířat“. Příroda. 548 (7669): 578–581. Bibcode:2017Natur.548..578B. doi:10.1038 / příroda23457. ISSN 1476-4687. PMID 28813409. S2CID 205258987.
- ^ Herron, Matthew D .; Borin, Joshua M .; Boswell, Jacob C .; Walker, Jillian; Chen, I.-Chen Kimberly; Knox, Charles A .; Boyd, Margrethe; Rosenzweig, Frank; Ratcliff, William C. (2019-02-20). „De novo počátky mnohobuněčnosti v reakci na predaci“. Vědecké zprávy. 9 (1): 2328. Bibcode:2019NatSR ... 9.2328H. doi:10.1038 / s41598-019-39558-8. ISSN 2045-2322. PMC 6382799. PMID 30787483.
- ^ Bar-On, Yinon M .; Phillips, Rob; Milo, Ron (19. 6. 2018). „Distribuce biomasy na Zemi“. PNAS. 115 (25): 6506–6511. doi:10.1073 / pnas.1711842115. PMC 6016768. PMID 29784790.
externí odkazy
| Vývoj |  | |
|---|---|---|
| Populace genetika | ||
| Rozvoj | ||
| Z taxony | ||
| Z orgány | ||
| Z procesy | ||
| Tempo a režimy | ||
| Speciace | ||
| Dějiny | ||
| Filozofie | ||
| Příbuzný | ||
| ||