Mořské jídlo web - Marine food web - Wikipedia

| Část série přehledů o |
| mořský život |
|---|
Ve srovnání s pozemským prostředím má mořské prostředí pyramidy z biomasy které jsou obrácené na základně. Zejména biomasa spotřebitelů (kozonožci, krill, krevety, píce) je větší než biomasa prvovýrobci. Stává se to proto, že primární producenti oceánu jsou malí fytoplankton které rychle rostou a množí se, takže malá hmota může mít vysokou rychlost prvovýroba. Naproti tomu mnoho významných suchozemských primárních producentů, jako např zralé lesy, rostou a množí se pomalu, takže k dosažení stejné rychlosti primární produkce je zapotřebí mnohem většího množství.
Kvůli této inverzi je to zooplankton které tvoří většinu mořského živočicha biomasa. Tak jako primární spotřebitelé, zooplankton je zásadní vazbou mezi primárními producenty (zejména fytoplanktonem) a zbytkem web mořských potravin (sekundární spotřebitelé ).[1]
Pokud fytoplankton zemře dříve, než se sní, sestupuje skrz eufotická zóna jako součást mořský sníh a usazuje se do hlubin moře. Tímto způsobem fytoplankton každoročně odděluje do oceánu asi 2 miliardy tun oxidu uhličitého, což způsobí, že oceán se stane záchytem oxidu uhličitého, který drží asi 90% veškerého zachyceného uhlíku.[2] Oceán produkuje asi polovinu světového kyslíku a ukládá 50krát více oxidu uhličitého než atmosféra.[3]
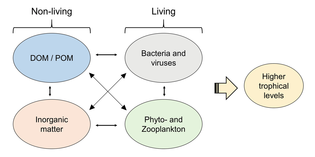
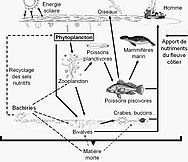
Ekosystém nelze pochopit bez znalosti toho, jak jeho potravní síť určuje tok materiálů a energie. Fytoplankton autotrofně produkuje biomasu přeměnou anorganické sloučeniny do organický ty. Tímto způsobem funguje fytoplankton jako základ mořské potravní sítě tím, že podporuje veškerý další život v oceánu. Druhým ústředním procesem v síti mořských potravin je mikrobiální smyčka. Tato smyčka se zhoršuje mořské bakterie a archaea, remineralizuje organické a anorganické látky a poté recykluje produkty buď v pelagické potravinové síti, nebo je ukládá jako sediment na mořském dně.[4]
Potravinové řetězce a trofické úrovně
| mořský potravinový řetězec (typický) |
|---|
| ↓ |
| fytoplankton |
| ↓ |
| býložravý zooplankton |
| ↓ |
| masožravý zooplankton |
| ↓ |
| ↓ |
Potravinové weby jsou postaveny z potravinové řetězce. Všechny formy života v moři mají potenciál stát se potravou pro další formu života. V oceánu začíná potravní řetězec obvykle energií ze slunce fytoplankton, a navazuje na kurz jako:
fytoplankton → býložravý zooplankton → masožravý zooplankton → podavač filtrů → dravé obratlovce
Fytoplankton nepotřebují k jídlu jiné organismy, protože mají schopnost vyrábět si vlastní jídlo přímo z anorganického uhlíku za použití slunečního záření jako zdroje energie. Tento proces se nazývá fotosyntéza a vede k tomu, že fytoplankton přeměňuje přirozeně se vyskytující uhlík na protoplazma. Z tohoto důvodu se o fytoplanktonu říká prvovýrobci na dně nebo na první úrovni mořského potravinového řetězce. Protože jsou na první úrovni, říká se, že mají tropická úroveň z 1 (z řecký trophē což znamená jídlo). Fytoplankton jsou pak konzumovány na další trofické úrovni v potravinovém řetězci mikroskopickými zvířaty zvanými zooplankton.
Zooplankton tvoří druhou trofickou hladinu v potravinovém řetězci a zahrnují mikroskopické jednobuněčné organismy zvané prvoky i malé korýši, jako copepods a krill a larva ryb, chobotnic, humrů a krabů. Organismy na této úrovni lze považovat za primární spotřebitelé.
Menší býložravý zooplankton zase konzumují větší masožravé zooplantery, jako jsou větší dravé prvoky a krill, a tím píce ryby, které jsou malé, vyučování, krmení filtrem Ryba. Toto tvoří třetí trofickou úroveň v potravinovém řetězci.
Čtvrtá trofická úroveň se skládá z dravé ryby, mořští savci a mořští ptáci které konzumují krmné ryby. Příklady jsou mečoun, těsnění a terejové.
Apexové predátoři, jako např kosatky, které mohou konzumovat těsnění, a žraloci mako, který může konzumovat mečouny, tvoří pátou trofickou úroveň. Baleen velryby může konzumovat zooplankton a krill přímo, což vede k potravnímu řetězci pouze se třemi nebo čtyřmi trofickými úrovněmi.
V praxi jsou trofické úrovně ne obvykle jednoduchá celá čísla protože stejný konzumní druh se často živí na více než jedné trofické úrovni.[8][9] Například velký mořští obratlovci může jíst menší dravé ryby, ale může také jíst krmítka pro filtry; the rejnok jí korýši, ale kladivoun jí korýše i rejnoky. Zvířata se mohou také navzájem jíst; the treska jí menší tresku a také rak a raky jedí larvy tresky. Stravovací návyky mladistvého zvířete a v důsledku toho jeho trofická úroveň se mohou s růstem měnit.
Vědec o rybolovu Daniel Pauly nastavuje hodnoty trofických úrovní na jednu u prvovýrobců a detritus, dva u býložravců a detritivores (primární spotřebitelé), tři u sekundárních spotřebitelů atd. Definice trofické úrovně TL pro jakýkoli konzumní druh je:[10]

kde je zlomková trofická úroveň kořisti j, a představuje zlomek j ve stravě i. V případě mořských ekosystémů má trofická úroveň většiny ryb a jiných mořských spotřebitelů hodnotu mezi 2,0 a 5,0. Horní hodnota, 5,0, je neobvyklá, dokonce i pro velké ryby,[11] ačkoli se vyskytuje u vrcholných predátorů mořských savců, jako jsou lední medvědi a kosatky.[12] Kontrastem je, že lidé mají střední trofickou hladinu přibližně 2,21, přibližně stejnou jako prase nebo sardela.[13][14]


Podle taxonu
Prvovýrobci

Na základně oceánské potravní sítě jsou jednobuněčné řasy a další rostlinné organismy známé jako fytoplankton. Stejně jako rostliny na souši se používá fytoplankton chlorofyl a další světelné pigmenty provést fotosyntéza, absorbující atmosférický oxid uhličitý k výrobě cukrů pro palivo. Chlorofyl ve vodě mění způsob, jakým voda odráží a absorbuje sluneční světlo, což vědcům umožňuje mapovat množství a umístění fytoplanktonu. Tato měření poskytují vědcům cenné poznatky o zdraví oceánského prostředí a pomáhají vědcům studovat oceánský uhlíkový cyklus.[15]
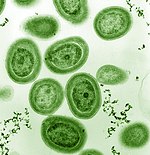
Mezi fytoplankton jsou členové z kmene bakterií zvaných sinice. Mořské sinice zahrnují nejmenší známé fotosyntetické organismy. Nejmenší ze všech, Prochlorococcus, je napříč jen 0,5 až 0,8 mikrometrů.[16] Pokud jde o jednotlivá čísla, Prochlorococcus je pravděpodobně nejhojnějším druhem na Zemi: jeden mililiter povrchové mořské vody může obsahovat 100 000 buněk nebo více. Celosvětově se odhaduje na několik oktilion (1027) Jednotlivci.[17] Prochlorococcus je všudypřítomný mezi 40 ° severní šířky a 40 ° jižní šířky a dominuje v oligotrofní (na živiny chudé) oblasti oceánů.[18] Bakterie tvoří asi 20% kyslíku v zemské atmosféře.[19]
- Fytoplankton tvoří základ oceánského potravinového řetězce

Fytoplankton

Dinoflagellate

Diatomy



V oceánech nejvíce prvovýroba provádí řasy. To je kontrast k pozemkům, kde většinu primární produkce provádí cévnatých rostlin. Řasy se pohybují od jednotlivých plovoucích buněk po připojené mořské řasy, zatímco cévnaté rostliny jsou v oceánu zastoupeny skupinami jako mořské trávy a mangrovy. Větší producenti, jako jsou mořské trávy a mořské řasy, jsou většinou omezeny na pobřežní zóna a mělké vody, kde jsou připojit na podkladový podklad a jsou stále uvnitř fotonická zóna. Většinu primární produkce řas však provádí fytoplankton.
V oceánském prostředí je tedy první spodní trofická úroveň obsazena hlavně fytoplankton mikroskopické driftující organismy jednobuněčný řasy, které se vznášejí v moři. Většina fytoplanktonu je příliš malá na to, aby ji bylo možné jednotlivě vidět pouhým okem. Mohou se jevit jako (často zelené) zabarvení vody, pokud jsou přítomny v dostatečně vysokém počtu. Protože svoji biomasu zvětšují hlavně fotosyntézou, žijí ve sluncem osvětlené povrchové vrstvě (eufotická zóna ) moře.
Mezi nejdůležitější skupiny fytoplanktonu patří rozsivky a dinoflageláty. Diatomy jsou zvláště důležité v oceánech, kde podle některých odhadů tvoří až 45% celkové primární produkce oceánu.[20] Diatomy jsou obvykle mikroskopický, i když některé druhy mohou dosahovat délky až 2 milimetrů.
Primární spotřebitelé
Druhá trofická úroveň (primární spotřebitelé ) je obsazen zooplankton které se živí fytoplankton. Spolu s fytoplanktonem tvoří základ potravní pyramidy, která podporuje většinu velkých světových rybářských revírů. Zooplankton jsou drobná zvířata nalezená s fytoplanktonem oceánský povrchové vody a zahrnují drobné korýši a larvy ryb a potěr (nedávno vylíhnutá ryba). Většina zooplanktonu je podavače filtrů a pomocí přídavků napínají fytoplankton ve vodě. Některé větší zooplanktony se také živí menším zooplanktonem. Některý zooplankton může trochu skákat, aby se vyhnul predátorům, ale neumí plavat. Stejně jako fytoplankton se vznášejí místo proudů, přílivu a odlivu. Zooplanktony se mohou rychle množit, jejich populace se může za příznivých podmínek zvýšit až o třicet procent denně. Mnoho lidí žije krátkým a produktivním životem a rychle dosáhne dospělosti.
- Zooplankton tvoří druhou úroveň v oceánském potravinovém řetězci

Segmentovaný červ

Drobní korýši podobní krevetám

Juvenilní planktonová chobotnice

Obzvláště důležité skupiny zooplanktonu jsou copepods a krill. Copepods jsou skupina malých korýši nalezený v oceánu a sladkovodní stanoviště. Jsou největším zdrojem bílkovin v moři,[21] a jsou důležitou kořistí pícnin. Krill představují další největší zdroj bílkovin. Krill jsou obzvláště velcí dravci zooplanktonu, kteří se živí menším zooplanktonem. To znamená, že skutečně patří ke třetí trofické úrovni, sekundárním spotřebitelům, spolu s pícninami.

Spolu, fytoplankton a zooplankton tvoří většinu z plankton v moři. Plankton je termín používaný pro jakýkoli malý drift organismy které se vznášejí v moři (řecký planktos = tulák nebo tulák). Podle definice organismy klasifikované jako plankton nejsou schopny plavat proti oceánským proudům; nemohou odolat okolnímu proudu a ovládat svou polohu. V oceánském prostředí jsou první dvě trofické úrovně obsazeny hlavně plankton. Plankton se dělí na výrobce a spotřebitele. Producenty jsou fytoplankton (řecky phyton = rostlina) a spotřebiteli, kteří konzumují fytoplankton, je zooplankton (řecky zoon = zvíře).
Medúza jsou pomalí plavci a většina druhů tvoří součást planktonu. Medúzy byly tradičně považovány za trofické slepé uličky, drobné hráče v síti mořských potravin, želatinové organismy s tělesný plán z velké části založen na vodě, která nabízí malou nutriční hodnotu nebo zájem o jiné organismy, kromě několika specializovaných predátorů, jako je mořské mušle a kožená mořská želva.[23][22] Tento názor byl nedávno zpochybněn. Medúzy a obecněji želatinový zooplankton který zahrnuje salpy a ctenofory, jsou velmi různorodé, křehké bez tvrdých částí, obtížně viditelné a monitorovatelné, podléhají rychlým výkyvům populace a často žijí nepohodlně daleko od pobřeží nebo hluboko v oceánu. Pro vědce je obtížné detekovat a analyzovat medúzy ve vnitřnostech predátorů, protože se při konzumaci změní na kaši a jsou rychle strávitelné.[23] Ale medúzy kvetou v obrovském množství a bylo prokázáno, že tvoří hlavní složky ve stravě tuňák, oštěp a mečoun stejně jako různí ptáci a bezobratlí, jako např chobotnice, mořské okurky, kraby a obojživelníci.[24][22] „I přes nízkou energetickou hustotu může být podíl medúz na energetickém rozpočtu predátorů mnohem větší, než se předpokládá, a to kvůli rychlému trávení, nízkým nákladům na zachycení, dostupnosti a selektivnímu krmení energeticky bohatších složek. mořští predátoři náchylní k požití plastů. “[22]
Spotřebitelé vyšších objednávek

- Mořští bezobratlí
- Ryba
- Krmivo pro ryby: Krmné ryby zaujímají centrální polohy v oceánu potravinářské weby. Organismy, které jí, jsou nižší tropická úroveň a organismy, které ji jedí, jsou na vyšší trofické úrovni. Krmné ryby zaujímají střední úroveň v potravním pásu a slouží jako dominantní kořist pro ryby, mořské ptáky a savce vyšší úrovně.[25]
- Predátor ryby
- Mleté ryby
- Ostatní mořští obratlovci
V roce 2010 vědci zjistili, že velryby přenášejí živiny z hlubin oceánu zpět na povrch pomocí procesu, kterému říkali velrybí čerpadlo.[26] Velryby se krmí na hlubších úrovních v oceánu, kde krill je nalezen, ale pravidelně se vracejte na povrch, abyste dýchali. Tam velryby vyprázdněte tekutinu bohaté na dusík a železo. Místo ponoření zůstává kapalina na povrchu, kde fytoplankton konzumovat to. V Mainském zálivu poskytuje velrybí čerpadlo více dusíku než řeky.[27]

Keporkaky výpad zdola krmit se pícninami

Terejové ponorný ponor shora chytat pícniny
 Velrybí čerpadlo cyklus živin
Velrybí čerpadlo cyklus živin

Mikroorganismy

V posledních letech dochází k rostoucímu uznání mořské mikroorganismy hrají v mořských ekosystémech mnohem větší roli, než se dříve myslelo. Vývoj v roce 2006 metagenomika dává vědcům schopnost odhalit dříve skryté rozmanitosti mikroskopického života a nabízí výkonnou čočku pro sledování mikrobiálního světa a potenciál revoluce v chápání živého světa.[29] Metabarcodingová dietní analýza techniky se používají k rekonstrukci potravinářských webů na vyšších úrovních taxonomického rozlišení a odhalují hlubší složitost v síti interakcí.[30]
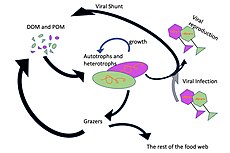
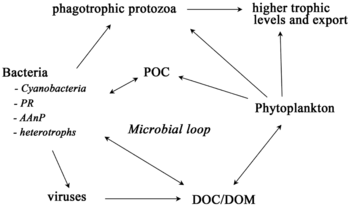
Mikroorganismy hrají klíčové role v sítích mořských potravin. The virový zkrat Pathway je mechanismus, který zabraňuje mořským mikrobiálním látkám částice organické hmoty (POM) z migrace nahoru trofické úrovně jejich recyklací do rozpuštěná organická hmota (DOM), které mohou mikroorganismy snadno absorbovat.[31] Virový posun pomáhá udržovat rozmanitost v mikrobiálním ekosystému tím, že brání jedinému druhu mořských mikrobů v ovládnutí mikroprostředí.[32] DOM recyklovaný cestou virového zkratu je srovnatelný s množstvím generovaným jinými hlavními zdroji mořského DOM.[33]
Obecně, rozpuštěný organický uhlík (DOC) se zavádí do oceánského prostředí bakteriální lýzou, únikem nebo exsudací fixovaného uhlíku z fytoplanktonu (např. Slizovitý exopolymer z rozsivky ), náhlá stárnutí buněk, nedbalé krmení zooplanktonem, vylučování odpadních produktů vodními živočichy nebo rozklad nebo rozpouštění organických částic ze suchozemských rostlin a půd.[34] Bakterie v mikrobiální smyčka rozložit tento částicový detritus a využít tuto energeticky bohatou hmotu pro růst. Vzhledem k tomu, že více než 95% organické hmoty v mořských ekosystémech sestává z polymerní, vysoké molekulární váha (HMW) sloučeniny (např. Protein, polysacharidy, lipidy), pouze malá část z celkového počtu rozpuštěná organická hmota (DOM) je snadno použitelný pro většinu mořských organismů na vyšších trofických úrovních. To znamená, že rozpuštěný organický uhlík není k dispozici přímo většině mořských organismů; mořské bakterie zavádějí tento organický uhlík do potravinářské sítě, což má za následek dostupnost další energie pro vyšší trofické úrovně.
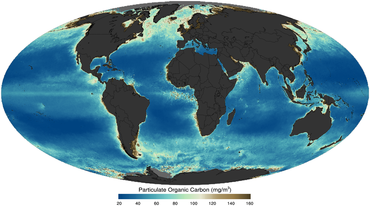
tak jako zobrazeno satelitem v roce 2011




Obří mořský virus CroV infikuje a způsobuje smrt lýza mořského zooflagelátu Cafeteria roenbergensis.[38] To má vliv na pobřežní ekologii, protože Cafeteria roenbergensis se živí bakteriemi nacházejícími se ve vodě. Pokud je nízký počet Cafeteria roenbergensis v důsledku rozsáhlých infekcí CroV bakteriální populace exponenciálně rostou.[39] Dopad CroV o přirozené populaci C. roenbergensis zůstává neznámý; Bylo však zjištěno, že virus je velmi specifický pro hostitele a neinfikuje jiné blízce příbuzné organismy.[40] Cafeteria roenbergensis je také infikována druhým virem, Virový virofág, což je satelitní virus, což znamená, že je schopen replikovat se pouze v přítomnosti jiného specifického viru, v tomto případě v přítomnosti CroV.[41] Tento virus interferuje s replikací CroV, což vede k přežití C. roenbergensis buňky. Mavirus je schopen integrovat se do genomu buněk C. roenbergensis, a tím uděluje obyvatelstvu imunitu.[42]
Houby


Podle stanoviště
Pelagické weby
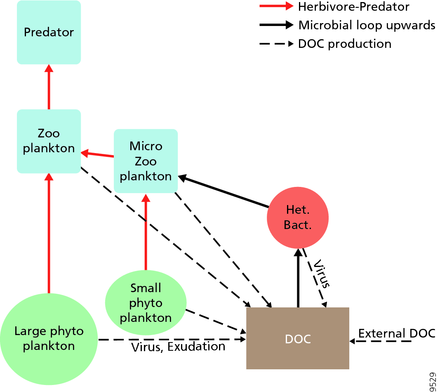
Pro pelagické ekosystémy navrhli Legendre a Rassoulzadagan v roce 1995 kontinuum trofických cest s býložravým potravinovým řetězcem a mikrobiální smyčka jako koncoví členové potravinářského webu.[47] Klasický lineární koncový člen potravinového řetězce zahrnuje pastvu zooplanktonu na větším fytoplanktonu a následnou predaci zooplanktonu buď větším zooplanktonem nebo jiným predátorem. V takovém lineárním potravinovém řetězci může predátor vést buď k vysokému fytoplanktonu biomasa (v systému s fytoplanktonem, býložravcem a predátorem) nebo se sníženou biomasou fytoplanktonu (v systému se čtyřmi úrovněmi). Mohou tedy vést ke změnám v množství dravců trofické kaskády.[48] Koncový člen mikrobiální smyčky zahrnuje nejen fytoplankton jako základní zdroj, ale také rozpuštěný organický uhlík.[49] Rozpuštěný organický uhlík je používán heterotrofními bakteriemi k růstu, předcházeného větším zooplanktonem. V důsledku toho se rozpuštěný organický uhlík transformuje pomocí smyčky bakteriální-mikrozooplankton na zooplankton. Tyto dvě dráhy zpracování uhlíku koncových členů jsou spojeny na více úrovních. Malý fytoplankton lze konzumovat přímo mikrozooplanktonem.[46]
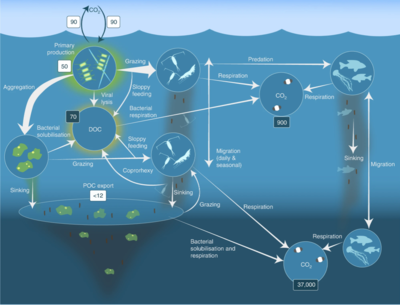
Jak je znázorněno na obrázku vpravo, rozpuštěný organický uhlík je produkován různými způsoby a různými organismy, a to jak primárními producenty, tak spotřebiteli organického uhlíku. K uvolňování DOC u primárních producentů dochází pasivně únikem a aktivně během nevyváženého růstu během omezení živin.[50][51] Další přímá cesta od fytoplanktonu k rozpuštěnému organickému poolu zahrnuje virová lýza.[52] Mořské viry jsou hlavní příčinou úmrtnosti fytoplanktonu v oceánu, zejména v teplejších vodách s nízkou šířkou. Nedbalé krmení býložravci a neúplné trávení kořisti spotřebiteli jsou dalšími zdroji rozpuštěného organického uhlíku. Heterotrofní mikroby používají k solubilizaci extracelulární enzymy částicový organický uhlík a použít tento a další zdroje rozpuštěného organického uhlíku pro růst a údržbu. Část mikrobiální heterotrofní produkce je využívána mikrozooplanktonem; jiná část heterotrofní komunity podléhá intenzivní virové lýze a to způsobuje opětovné uvolňování rozpuštěného organického uhlíku. Účinnost mikrobiální smyčky závisí na mnoha faktorech, ale zejména na relativní důležitosti predace a virové lýzy pro úmrtnost heterotrofních mikrobů.[46]
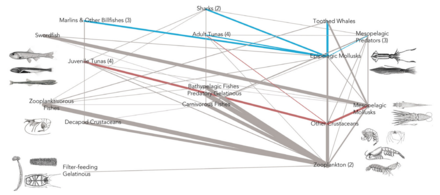
- Pelagický web s potravinami

Pelagická potravní síť a biologická pumpa. Spojení mezi oceánskou biologickou pumpou a pelagickou potravinářskou sítí a schopnost vzdáleně ochutnat tyto komponenty z lodí, satelitů a autonomních vozidel. Světle modré vody jsou eufotická zóna, zatímco tmavší modré vody představují zóna soumraku.[53]

- Mesopelagic food web

Dopad mezopelagických druhů na globální rozpočet na uhlík[54]
DVM = díl vertikální migrace NM = nemigrace


Mezopelagický bristlemouths mohou být nejhojnějšími obratlovci na planetě, i když je o nich známo jen málo.[55]

Želatinové dravce takhle narcomedusan konzumují největší rozmanitost mezopelagické kořisti


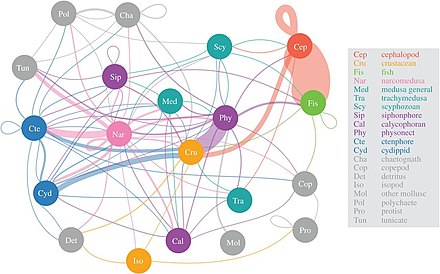
Vědci začínají podrobněji zkoumat z velké části neznámou zónu soumraku mezopelagický, Hluboký 200 až 1 000 metrů. Tato vrstva je odpovědná za každoroční odstraňování asi 4 miliard tun oxidu uhličitého z atmosféry. Mezopelagická vrstva je obývána většinou mořských ryb biomasa.[55]
Podle studie z roku 2017 narcomedusae konzumovat největší rozmanitost mezopelagické kořisti, následovaný physonect sifonofory, ctenofory a hlavonožci. Význam takzvaného „rosolovitého rouna“ teprve začíná být pochopen, ale zdá se, že medúzy, ctenofory a sifonofory mohou být klíčovými predátory v hlubokých pelagických potravních sítích s ekologickými dopady podobnými rybám dravců a chobotnic. Tradičně želatinové dravce byly považovány za neúčinné poskytovatele mořských trofických cest, ale zdá se, že mají zásadní a nedílnou roli v hlubokých pelagických potravních sítích.[56] Diel vertikální migrace, důležitý aktivní transportní mechanismus, umožňuje mesozooplankton izolovat oxid uhličitý z atmosféry a dodávat uhlík pro další mesopelagické organismy.[57]
Studie z roku 2020 uvádí, že do roku 2050 by se globální oteplování mohlo šířit v hlubokém oceánu sedmkrát rychleji než nyní, i když se sníží emise skleníkových plynů. Zahřívá se mezopelagický a hlubší vrstvy mohou mít zásadní důsledky pro potravní síť hlubokých oceánů, protože druhy oceánů se budou muset pohybovat, aby zůstaly na teplotách přežití.[58][59]
- Ryby za soumraku vrhají nové světlo na oceánský ekosystém Konverzace, 10. února 2014.
- Ocean Mystery in the Trillions The New York Times, 29. června 2015.
- Mesopelagické ryby - Malaspina objížďka expedice roku 2010.[60][61]
Na povrchu oceánu

Oceánská povrchová stanoviště sedí na rozhraní mezi oceánem a atmosférou. The jako biofilm stanoviště na povrchu oceánu ukrývá mikroorganismy žijící na povrchu, běžně označované jako neuston. Toto rozsáhlé rozhraní vzduch-voda leží na křižovatce hlavních procesů výměny vzduch-voda, které pokrývají více než 70% globální povrchové plochy. Bakterie v povrchové mikrovrstvě oceánu, tzv bakterioneuston, jsou zajímavé díky praktickým aplikacím, jako je výměna skleníkových plynů mezi vzduchem a mořem, výroba klimaticky aktivních mořských aerosolů a dálkový průzkum oceánu.[63] Zvláštní pozornost je věnována produkci a degradaci povrchově aktivní látky (povrchově aktivní materiály) mikrobiálními biochemickými procesy. Mezi hlavní zdroje povrchově aktivních látek v otevřeném oceánu patří fytoplankton,[64] pozemský odtok a depozice z atmosféry.[63]
Na rozdíl od barevných květů řas nemusí být bakterie spojené s povrchově aktivní látkou viditelné na barevných obrázcích oceánů. Mít schopnost detekovat tyto „neviditelné“ bakterie spojené s povrchově aktivní látkou pomocí radar se syntetickou clonou má obrovské výhody za každého počasí, bez ohledu na mrak, mlhu nebo denní světlo.[63] To je zvláště důležité ve velmi silném větru, protože to jsou podmínky, kdy dochází k nejintenzivnějším výměnám vzduch-moře a produkci mořského aerosolu. Proto kromě barevných satelitních snímků mohou satelitní snímky SAR poskytnout další pohled na globální obraz biofyzikálních procesů na hranici mezi oceánem a atmosférou, výměny skleníkových plynů vzduch-moře a produkce mořských aerosolů aktivních z hlediska klimatu.[63]
Na dně oceánu
Dno oceánu (bentický ) stanoviště sedí na rozhraní mezi oceánem a vnitřkem Země.
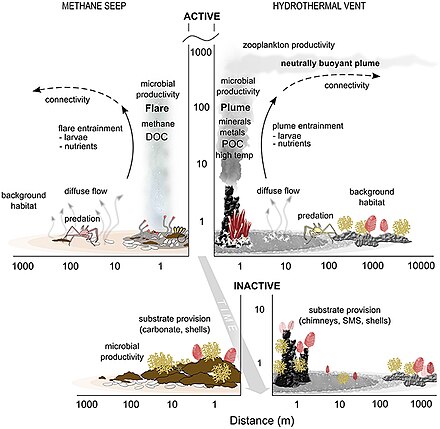
- Průsaky a průduchy
Pobřežní sítě

Mezi pobřežní vody patří vody v ústí řek a znovu kontinentální police. Zabírají asi 8 procent z celkové oceánské oblasti[70] a tvoří asi polovinu veškeré produktivity oceánu.
Klíčové živiny určující eutrofizace jsou dusík v pobřežních vodách a fosfor v jezerech. Oba se nacházejí ve vysokých koncentracích v guano (výkaly mořských ptáků), které působí jako hnojivo pro okolní oceán nebo přilehlé jezero. Kyselina močová je dominantní sloučeninou dusíku a během její mineralizace vznikají různé formy dusíku. V diagramu vpravo: (1) amonifikace produkuje NH3 a NH4 + a (2) nitrifikace produkuje NO3− oxidací NH4 +. Za alkalických podmínek, typických pro výkaly mořských ptáků, je NH3 rychle těkavý (3) a transformován na NH4 +, který je transportován z kolonie, a mokrou depozicí exportován do vzdálených ekosystémů, které jsou eutrofizovány (4). Cyklus fosforu je jednodušší a má sníženou pohyblivost. Tento prvek se nachází v řadě chemických forem ve fekálním materiálu mořského ptáka, ale nejpohyblivější a biologicky dostupný je ortofosfát, který může být vyluhuje podzemními nebo povrchovými vodami (5).[69]

na kontinentálním šelfu
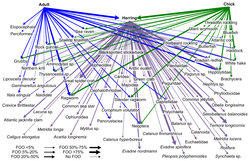
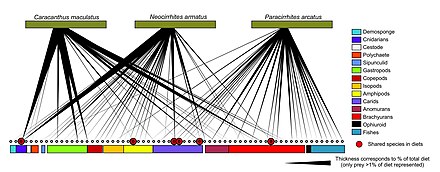
Čárový kód DNA lze použít ke konstrukci struktur potravinářského webu s lepším taxonomickým rozlišením v uzlech webu. To poskytuje konkrétnější identifikaci druhů a větší jasnost o tom, kdo koho jí. „Čárové kódy DNA a informace o DNA mohou umožnit nové přístupy ke konstrukci větších interakčních sítí a překonat některé překážky při dosahování přiměřené velikosti vzorku.“[30]
Nově používanou metodou pro identifikaci druhů je metabarcoding DNA. Identifikace druhů prostřednictvím morfologie je poměrně obtížné a vyžaduje spoustu času a odborných znalostí.[79][80] Vysoce výkonné sekvenování Metabarcoding DNA umožňuje taxonomické přiřazení a tedy identifikaci celého vzorku, pokud jde o specifické pro skupinu primery vybráno pro předchozí DNA zesílení.

- Čárový kód mikrobiální DNA
- Čárové kódy DNA řas
- Čárový kód rybí DNA
- Čárový kód DNA při hodnocení stravy
- Řasy lesy
- Byrnes, J.E., Reynolds, P.L. a Stachowicz, J.J. (2007) „Invaze a vymírání přetvářejí pobřežní mořské potravní sítě“. PLOS ONE, 2(3): e295. doi:10.1371 / journal.pone.0000295
Polární weby
obklopen oceány
obklopen pevninami
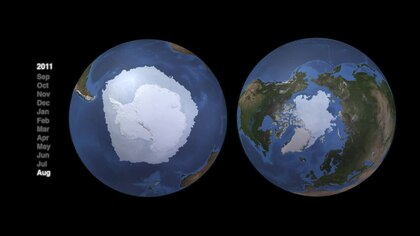
Arktické a antarktické mořské systémy se velmi liší topografické struktury a v důsledku toho mají velmi odlišné struktury potravinového webu.[82] Arktické i antarktické pelagické potravní sítě mají charakteristické energetické toky řízené převážně několika klíčovými druhy. Ani pro jeden však neexistuje žádný obecný web. Pro odolnost a udržení energetických toků jsou důležité alternativní cesty. Tyto komplikovanější alternativy však poskytují menší tok energie druhům vyšších trofických úrovní. „Struktura potravní sítě může být v různých oblastech podobná, ale jednotlivé druhy, které dominují středním trofickým úrovním, se v polárních oblastech liší.“[83]



- Arktický
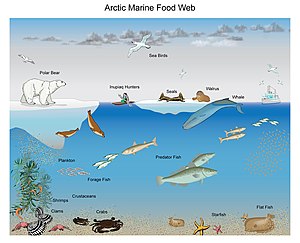
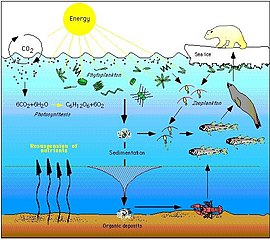
Arktický web s potravinami je složitý. Ztráta mořského ledu může v konečném důsledku ovlivnit celou potravní síť, od řas a planktonu přes ryby až po savce. Dopad změny klimatu na konkrétní druh se může vlnit potravní sítí a ovlivnit celou řadu dalších organismů ... Nejenže pokles mořského ledu poškozuje populace ledních medvědů snížením rozsahu jejich primárního prostředí, ale také negativně je ovlivňovat prostřednictvím efektů potravinového webu. Poklesy trvání a rozsahu mořského ledu v Arktidě vedou k poklesu množství ledových řas, kterým se v ledech daří v kapsách bohatých na živiny. Tyto řasy konzumuje zooplankton, který je zase konzumován arktickou treskou, důležitým zdrojem potravy pro mnoho mořských savců, včetně tuleňů. Těsnění požírají lední medvědi. Pokles ledových řas proto může přispět k poklesu populací ledních medvědů.[84]
V roce 2020 vědci uvedli, že měření za poslední dvě desetiletí pokračovala prvovýroba v Severní ledový oceán vykazují nárůst o téměř 60% v důsledku vyšších koncentrací fytoplankton. Předpokládají, že nové živiny proudí z jiných oceánů, a naznačují, že to znamená, že Severní ledový oceán může být schopen podporovat vyšší trofická úroveň produkce a další uhlíková fixace v budoucnu.[85][86]


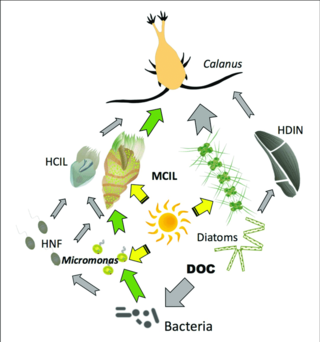
Šedé šipky: tok uhlíku k heterotrofům
Zelené šipky: hlavní cesty toku uhlíku do nebo z mixotrofů
HCIL: heterotrofní nájemníci; MCIL: mixotrofní nálevníci; HNF: heterotrofní nanoflageláty; DOC: rozpuštěný organický uhlík; HDIN: heterotrofní dinoflageláty[87]

- antarktický

Antarktické medúzy Diplulmaris antarctica pod ledem

Kolonie řas Phaeocystis antarctica, důležitý fytoplanktér v Rossově moři, který dominuje květům rané sezóny poté, co mořský led ustoupí a vyváží významný uhlík.[89]

The pennate rozsivka Fragilariopsis kerguelensis, který se nachází v celém antarktickém cirkumpolárním proudu, je klíčovou hnací silou globálního světa silikátové čerpadlo.[90]

Skupina kosatky pokus o uvolnění a crabeater těsnění na ledová kra




a uhlíkové hmoty (Gt C) v tmavých polích


Foundation and keystone species



Druhy základů are species that have a dominant role structuring an ekologická komunita, shaping its environment and defining its ecosystem. Such ecosystems are often named after the foundation species, such as seagrass meadows, oyster beds, coral reefs, kelp forests and mangrove forests.[97] Například červená mangrovová is a common foundation species in mangrove forests. The mangrove’s root provides nursery grounds for young fish, such as snapper.[98] A foundation species can occupy any trophic level in a food web but tend to be a producer.[99]
The term was coined in 1972 by Paul K. Dayton,[100] who applied it to certain members of marine bezobratlý a řasy komunity. It was clear from studies in several locations that there were a small handful of species whose activities had a disproportionate effect on the rest of the marine community and they were therefore key to the resilience of the community. Dayton’s view was that focusing on foundation species would allow for a simplified approach to more rapidly understand how a community as a whole would react to disturbances, such as pollution, instead of attempting the extremely difficult task of tracking the responses of all community members simultaneously.


Keystone druhy are species that have large effects, disproportionate to their numbers, within ecosystem food webs.[101] An ecosystem may experience a dramatic shift if a keystone species is removed, even though that species was a small part of the ecosystem by measures of biomasa nebo produktivita.[102]
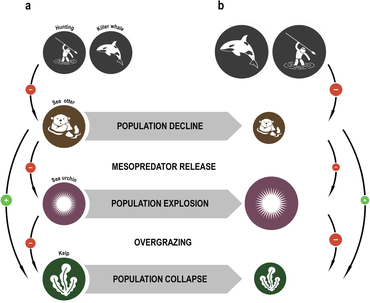
The concept of the keystone species was introduced in 1969 by the zoologist Robert T. Paine.[103][104] Paine developed the concept to explain his observations and experiments on the relationships between mořští bezobratlí z přílivová zóna (between the high and low tide lines), including mořská hvězdice a mušle. Nějaký mořské hvězdy kořist na mořští ježci, mušle, a další měkkýši that have no other natural predators. If the sea star is removed from the ecosystem, the mussel population explodes uncontrollably, driving out most other species.[105]
Mořské vydry limit the damage sea urchins inflict on řasy. When the sea otters of the North American west coast were hunted commercially for their fur, their numbers fell to such low levels that they were unable to control the sea urchin population. The urchins in turn grazed the zadržení z kelp so heavily that the kelp forests largely disappeared, along with all the species that depended on them. Reintroducing the sea otters has enabled the kelp ecosystem to be restored.[106][107]
Cryptic interactions

Cryptic interactions, interactions which are "hidden in plain sight", occur throughout the marine planktonic foodweb but are currently largely overlooked by established methods, which mean large‐scale data collection for these interactions is limited. Despite this, current evidence suggests some of these interactions may have perceptible impacts on foodweb dynamics and model results. Incorporation of cryptic interactions into models is especially important for those interactions involving the transport of nutrients or energy.[108]
Simplifications such as “zooplankton consume phytoplankton,” “phytoplankton take up inorganic nutrients,” “gross primary production determines the amount of carbon available to the foodweb,” etc. have helped scientists explain and model general interactions in the aquatic environment. Traditional methods have focused on quantifying and qualifying these generalizations, but rapid advancements in genomics, sensor detection limits, experimental methods, and other technologies in recent years have shown that generalization of interactions within the plankton community may be too simple. These enhancements in technology have exposed a number of interactions which appear as cryptic because bulk sampling efforts and experimental methods are biased against them.[108]
Complexity and stability
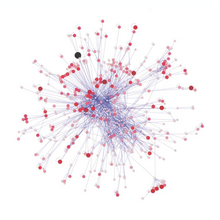
Potravinové weby provide a framework within which a complex network of predator–prey interactions can be organised. A food web model is a network of potravinové řetězce. Each food chain starts with a primární producent nebo autotrofní, an organism, such as an alga or a plant, which is able to manufacture its own food. Next in the chain is an organism that feeds on the primary producer, and the chain continues in this way as a string of successive predators. The organisms in each chain are grouped into trofické úrovně, based on how many links they are removed from the primary producers. The length of the chain, or trophic level, is a measure of the number of species encountered as energy or nutrients move from plants to top predators.[111] Energie jídla flows from one organism to the next and to the next and so on, with some energy being lost at each level. At a given trophic level there may be one species or a group of species with the same predators and prey.[112]
V roce 1927 Charles Elton published an influential synthesis on the use of food webs, which resulted in them becoming a central concept in ecology.[113] In 1966, interest in food webs increased after Robert Paine's experimental and descriptive study of intertidal shores, suggesting that food web complexity was key to maintaining species diversity and ecological stability.[114] Many theoretical ecologists, including Robert May a Stuart Pimm, were prompted by this discovery and others to examine the mathematical properties of food webs. According to their analyses, complex food webs should be less stable than simple food webs.[115]:75–77[116]:64 The apparent paradox between the complexity of food webs observed in nature and the mathematical fragility of food web models is currently an area of intensive study and debate. The paradox may be due partially to conceptual differences between persistence of a food web and equilibrial stabilita of a food web.[115][116]
A trofická kaskáda can occur in a food web if a trophic level in the web is suppressed.
For example, a top-down cascade can occur if predators are effective enough in predation to reduce the abundance, or alter the behavior, of their kořist, thereby releasing the next lower trophic level from predation. A top-down cascade is a trophic cascade where the top consumer/predator controls the primární spotřebitel populace. In turn, the primary producer population thrives. The removal of the top predator can alter the food web dynamics. In this case, the primary consumers would overpopulate and exploit the primary producers. Eventually there would not be enough primary producers to sustain the consumer population. Top-down food web stability depends on competition and predation in the higher trophic levels. Invasive species can also alter this cascade by removing or becoming a top predator. This interaction may not always be negative. Studies have shown that certain invasive species have begun to shift cascades; and as a consequence, ecosystem degradation has been repaired.[117][118] An example of a cascade in a complex, open-ocean ecosystem occurred in the northwest Atlantik během 80. a 90. let. Odstranění Treska obecná (Gadus morhua) and other ground fishes by sustained nadměrný rybolov resulted in increases in the abundance of the prey species for these ground fishes, particularly smaller krmit ryby and invertebrates such as the northern krab sněhu (Chionoecetes opilio) a severní krevety (Pandalus borealis). The increased abundance of these prey species altered the community of zooplankton that serve as food for smaller fishes and invertebrates as an indirect effect.[119] Top-down cascades can be important for understanding the knock-on effects of removing nejlepší dravci from food webs, as humans have done in many places through lov a rybolov.
In a bottom-up cascade, the population of primary producers will always control the increase/decrease of the energy in the higher trophic levels. Primary producers are plants, phytoplankton and zooplankton that require photosynthesis. Although light is important, primary producer populations are altered by the amount of nutrients in the system. This food web relies on the availability and limitation of resources. All populations will experience growth if there is initially a large amount of nutrients.[120][121]
Terrestrial comparisons

Compared to terrestrial biomass pyramids, aquatic pyramids are generally inverted at the base


Marine environments can have inversions in their biomass pyramids. In particular, the biomass of consumers (copepods, krill, shrimp, forage fish) is generally larger than the biomass of primary producers. This happens because the ocean's primary producers are mostly tiny phytoplankton which have r-stratég traits of growing and reproducing rapidly, so a small mass can have a fast rate of primary production. In contrast, many terrestrial primary producers, such as mature forests, have K-stratég traits of growing and reproducing slowly, so a much larger mass is needed to achieve the same rate of primary production. The rate of production divided by the average amount of biomass that achieves it is known as an organism's Production/Biomass (P/B) ratio.[123] Production is measured in terms of the amount of movement of mass or energy per area per unit of time. In contrast, the biomass measurement is in units of mass per unit area or volume. The P/B ratio utilizes inverse time units (example: 1/month). This ratio allows for an estimate of the amount of energy flow compared to the amount of biomass at a given trophic level, allowing for demarcations to be made between trophic levels. The P/B ratio most commonly decreases as trophic level and organismal size increases, with small, ephemeral organisms containing a higher P/B ratio than large, long-lasting ones.
Examples: The štětinatá borovice can live for thousands of years, and has a very low production/biomass ratio. Sinice Prochlorococcus lives for about 24 hours, and has a very high production/biomass ratio.


In oceans, most prvovýroba provádí řasy. This is a contrast to on land, where most primary production is performed by cévnatých rostlin.
Aquatic producers, such as planktonic algae or aquatic plants, lack the large accumulation of sekundární růst that exists in the woody trees of terrestrial ecosystems. However, they are able to reproduce quickly enough to support a larger biomass of grazers. This inverts the pyramid. Primary consumers have longer lifespans and slower growth rates that accumulates more biomass than the producers they consume. Phytoplankton live just a few days, whereas the zooplankton eating the phytoplankton live for several weeks and the fish eating the zooplankton live for several consecutive years.[125] Aquatic predators also tend to have a lower death rate than the smaller consumers, which contributes to the inverted pyramidal pattern. Population structure, migration rates, and environmental refuge for prey are other possible causes for pyramids with biomass inverted. Energy pyramids, however, will always have an upright pyramid shape if all sources of food energy are included, since this is dictated by the druhý zákon termodynamiky."[126][127]
Comparison of productivity in marine and terrestrial ecosystems[128] | |||
|---|---|---|---|
| Ekosystém | Čistá primární produktivita billion tonnes per year | Total plant biomass billion tonnes | Turnover time let |
Marine | 45–55 | 1–2 | 0.02–0.06 |
Pozemní | 55–70 | 600–1000 | 9–20 |
Anthropogenic effects

- Nadměrný rybolov
- Okyselení
Pteropods a křehké hvězdy společně tvoří základnu Arktidy potravinářské weby a oba jsou okyselením vážně poškozeni. Mušle Pteropods se rozpouštějí s rostoucím okyselením a křehké hvězdy ztrácejí svalovou hmotu při opětovném růstu příloh.[130] Vajíčka křehké hvězdy navíc zemřou během několika dní, když jsou vystavena očekávaným podmínkám vyplývajícím z okyselení Arktidy.[131] Okyselení hrozí zničením arktických potravinových sítí od základny. Arktické vody se rychle mění a jsou pokročilejší v procesu, kdy budou aragonitem nenasyceny.[132] Arktické potravinové sítě jsou považovány za jednoduché, což znamená, že v potravinovém řetězci existuje několik kroků od malých organismů po větší predátory. Například pteropody jsou „klíčovou kořistí mnoha vyšších predátorů - většího planktonu, ryb, mořských ptáků, velryb“.[133]


- Klimatická změna
"Our results show how future climate change can potentially weaken marine food webs through reduced energy flow to higher trophic levels and a shift towards a more detritus-based system, leading to food web simplification and altered producer–consumer dynamics, both of which have important implications for the structuring of benthic communities."[134][135]
"...increased temperatures reduce the vital flow of energy from the primary food producers at the bottom (e.g. algae), to intermediate consumers (herbivores), to predators at the top of marine food webs. Such disturbances in energy transfer can potentially lead to a decrease in food availability for top predators, which in turn, can lead to negative impacts for many marine species within these food webs... "Whilst climate change increased the productivity of plants, this was mainly due to an expansion of cyanobacteria (small blue-green algae)," said Mr Ullah. "This increased primary productivity does not support food webs, however, because these cyanobacteria are largely unpalatable and they are not consumed by herbivores. Understanding how ecosystems function under the effects of global warming is a challenge in ecological research. Most research on ocean warming involves simplified, short-term experiments based on only one or a few species."[135]
Viz také
Reference
- ^ US Department of Energy (2008) Cyklus uhlíku a biosekvestrace strana 81, Workshop report DOE / SC-108, US Department of Energy Office of Science.
- ^ Campbell, Mike (22. června 2011). „Role mořského planktonu při sekvestraci uhlíku“. EarthTimes. Citováno 22. srpna 2014.
- ^ Why should we care about the ocean? NOAA: National Ocean Service. Updated: 7 January 2020. Retrieved 1 March 2020.
- ^ Heinrichs, M.E., Mori, C. and Dlugosch, L. (2020) "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". In: Jungblut S., Liebich V., Bode-Dalby M. (Eds) YOUMARES 9-The Oceans: Our Research, Our Future , pages 279–297. Springer. doi:10.1007/978-3-030-20389-4_15.
- ^ Dunne, J.A., Williams, R.J. and Martinez, N.D. (2002) "Food-web structure and network theory: the role of connectance and size". Sborník Národní akademie věd, 99(20): 12917–12922. doi:10.1073 / pnas.192407699.
- ^ Dunne, J.A. (2006) "The network structure of food webs". In: M Pascual and J. A. Dunne (Eds.) Ecological networks: linking structure to dynamics in food webs, pages 27–86. ISBN 9780199775057.
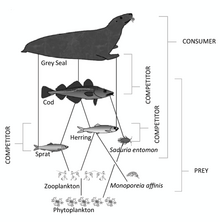
- ^ Karlson, A.M., Gorokhova, E., Gårdmark, A., Pekcan-Hekim, Z., Casini, M., Albertsson, J., Sundelin, B., Karlsson, O. and Bergström, L. (2020). „Propojení fyziologického stavu spotřebitele se strukturou potravinové sítě a hodnotou kořisti v Baltském moři“. Ambio, 49(2): 391–406. doi:10.1007 / s13280-019-01201-1
- ^ Odum, W. E.; Heald, E. J. (1975) "The detritus-based food web of an estuarine mangrove community". Pages 265–286 in L. E. Cronin, ed. Estuarine research. Sv. 1. Academic Press, New York.
- ^ Pimm, S.L .; Lawton, J. H. (1978). "Při krmení na více než jedné trofické úrovni". Příroda. 275 (5680): 542–544. doi:10.1038 / 275542a0. S2CID 4161183.
- ^ Pauly, D .; Palomares, M. L. (2005). „Rybaření sítí mořských potravin: je mnohem všudypřítomnější, než jsme si mysleli“ (PDF). Bulletin of Marine Science. 76 (2): 197–211. Archivovány od originál (PDF) dne 2013-05-14.
- ^ Cortés, E. (1999). "Standardized diet compositions and trophic levels of sharks". ICES J. Mar. Sci. 56 (5): 707–717. doi:10.1006/jmsc.1999.0489.
- ^ Pauly, D .; Trites, A.; Capuli, E.; Christensen, V. (1998). "Diet composition and trophic levels of marine mammals". ICES J. Mar. Sci. 55 (3): 467–481. doi:10.1006/jmsc.1997.0280.
- ^ Researchers calculate human trophic level for first time Phys.org . 3. prosince 2013.
- ^ Bonhommeau, S., Dubroca, L., Le Pape, O., Barde, J., Kaplan, D.M., Chassot, E. and Nieblas, A.E. (2013) "Eating up the world’s food web and the human trophic level". Sborník Národní akademie věd, 110(51): 20617–20620. doi:10.1073 / pnas.1305827110.
- ^ A b Chlorofyl NASA Earth Observatory. Zpřístupněno 30. listopadu 2019.
- ^ Kettler GC, Martiny AC, Huang K, Zucker J, Coleman ML, Rodrigue S, Chen F, Lapidus A, Ferriera S, Johnson J, Steglich C, Church GM, Richardson P, Chisholm SW (December 2007). "Patterns and implications of gene gain and loss in the evolution of Prochlorococcus". Genetika PLOS. 3 (12): e231. doi:10.1371/journal.pgen.0030231. PMC 2151091. PMID 18159947.
- ^ Nemiroff, R .; Bonnell, J., eds. (27. září 2006). "Earth from Saturn". Astronomický snímek dne. NASA.
- ^ Partensky F, Hess WR, Vaulot D (March 1999). "Prochlorococcus, a marine photosynthetic prokaryote of global significance". Recenze mikrobiologie a molekulární biologie. 63 (1): 106–27. doi:10.1128/MMBR.63.1.106-127.1999. PMC 98958. PMID 10066832.
- ^ „Nejdůležitější mikrob, o kterém jste nikdy neslyšeli“. npr.org.
- ^ Mann, D. G. (1999). "The species concept in diatoms". Phycologia. 38 (6): 437–495. doi:10.2216/i0031-8884-38-6-437.1.
- ^ Biologie koponožců Archivováno 01.01.2009 na Wayback Machine na Univerzita Carla von Ossietzkyho v Oldenburgu
- ^ A b C d Hays, G.C., Doyle, T.K. and Houghton, J.D. (2018) "A paradigm shift in the trophic importance of jellyfish?" Trendy v ekologii a evoluci, 33(11): 874-884. doi:10.1016/j.tree.2018.09.001
- ^ A b Hamilton, G. (2016) "The secret lives of jellyfish: long regarded as minor players in ocean ecology, jellyfish are actually important parts of the marine food web". Příroda, 531(7595): 432-435. doi:10.1038/531432a
- ^ Cardona, L., De Quevedo, I.Á., Borrell, A. and Aguilar, A. (2012) "Massive consumption of gelatinous plankton by Mediterranean apex predators". PLOS ONE, 7(3): e31329. doi:10.1371/journal.pone.0031329
- ^ Tiny Forage Fish At Bottom Of Marine Food Web Get New Protections Národní veřejné rádio, 7. dubna 2016.
- ^ Roman, J. & McCarthy, J.J. (2010). „Velrybí pumpa: mořští savci zvyšují primární produktivitu v pobřežní pánvi“. PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO ... 513255R. doi:10.1371 / journal.pone.0013255. PMC 2952594. PMID 20949007. e13255.CS1 maint: používá parametr autoři (odkaz)
- ^ Brown, Joshua E. (12. října 2010). „Velrybí hovínko pumpuje zdraví oceánu“. Věda denně. Citováno 18. srpna 2014.
- ^ Raina, J.B. (2018) "The life aquatic at the microscale". mSystémy, 3(2): e00150-17. doi:10.1128/mSystems.00150-17.
 Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0. - ^ Marco, D, ed. (2011). Metagenomics: Current Innovations and Future Trends. Caister Academic Press. ISBN 978-1-904455-87-5.
- ^ A b C Roslin, T. and Majaneva, S. (2016) "The use of DNA barcodes in food web construction—terrestrial and aquatic ecologists unite!". Genom, 59(9): 603–628. doi:10.1139/gen-2015-0229.
- ^ Wilhelm, Steven W .; Suttle, Curtis A. (1999). „Viry a cykly živin v moři“. BioScience. 49 (10): 781–788. doi:10.2307/1313569. ISSN 1525-3244. JSTOR 1313569.
- ^ Weinbauer, Markus G., et al. "Synergistic and antagonistic effects of viral lysis and protistan grazing on bacterial biomass, production and diversity." Mikrobiologie prostředí 9.3 (2007): 777-788.
- ^ Robinson, Carol, and Nagappa Ramaiah. "Microbial heterotrophic metabolic rates constrain the microbial carbon pump." The American Association for the Advancement of Science, 2011.
- ^ Van den Meersche, K., Middelburg, J.J., Soetaert, K., Van Rijswijk, P., Boschker, H.T. and Heip, C.H. (2004) "Carbon‐nitrogen coupling and algal‐bacterial interactions during an experimental bloom: Modeling a 13C tracer experiment". Limnologie a oceánografie, 49(3): 862–878. doi:10.4319/lo.2004.49.3.0862.
- ^ Käse L, Geuer JK. (2018) "Phytoplankton Responses to Marine Climate Change – An Introduction". In Jungblut S., Liebich V., Bode M. (Eds) YOUMARES 8–Oceans Across Boundaries: Learning from each other, pages 55–72, Springer. doi:10.1007/978-3-319-93284-2_5. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Heinrichs, M.E., Mori, C. and Dlugosch, L. (2020) "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". V: YOUMARES 9-The Oceans: Our Research, Our Future , pages 279–297. Springer. doi:10.1007/978-3-030-20389-4_15.
- ^ Duponchel, S. and Fischer, M.G. (2019) "Viva lavidaviruses! Five features of virophages that parasitize giant DNA viruses". PLoS pathogens, 15(3). doi:10.1371/journal.ppat.1007592. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Fischer, M. G.; Allen, M. J .; Wilson, W. H.; Suttle, C. A. (2010). "Giant virus with a remarkable complement of genes infects marine zooplankton" (PDF). Sborník Národní akademie věd. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073/pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Matthias G. Fischer; Michael J. Allen; William H. Wilson; Curtis A. Suttle (2010). "Giant virus with a remarkable complement of genes infects marine zooplankton" (PDF). Sborník Národní akademie věd. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073/pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Massana, Ramon; Javier Del Campo; Christian Dinter; Ruben Sommaruga (2007). "Crash of a population of the marine heterotrophic flagellate Cafeteria roenbergensis by viral infection". Mikrobiologie prostředí. 9 (11): 2660–2669. doi:10.1111/j.1462-2920.2007.01378.x. PMID 17922751. S2CID 30191542.
- ^ Fischer MG, Suttle CA (April 2011). "A virophage at the origin of large DNA transposons". Věda. 332 (6026): 231–4. Bibcode:2011Sci...332..231F. doi:10.1126/science.1199412. PMID 21385722. S2CID 206530677.
- ^ Fischer MG, Hackl (December 2016). "Host genome integration and giant virus-induced reactivation of the virophage mavirus". Příroda. 540 (7632): 288–91. Bibcode:2016Natur.540..288F. doi:10.1038/nature20593. PMID 27929021. S2CID 4458402.
- ^ Kagami, M., Miki, T. and Takimoto, G. (2014) "Mycoloop: chytrids in aquatic food webs". Frontiers in microbiology, 5: 166. doi:10.3389/fmicb.2014.00166. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Creative Commons Attribution 3.0 International License.
- ^ Amend, A., Burgaud, G., Cunliffe, M., Edgcomb, V.P., Ettinger, C.L., Gutiérrez, M.H., Heitman, J., Hom, E.F., Ianiri, G., Jones, A.C. and Kagami, M. (2019) "Fungi in the marine environment: Open questions and unsolved problems". MBio, 10(2): e01189-18. doi:10.1128/mBio.01189-18. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Gutierrez MH, Jara AM, Pantoja S (2016) "Fungal parasites infect marine diatoms in the upwelling ecosystem of the Humboldt current system off central Chile". Environ Microbiol, 18(5): 1646–1653. doi:10.1111/1462-2920.13257.
- ^ A b C Middelburg J.J. (2019) "The Return from Organic to Inorganic Carbon". V: Marine Carbon Biogeochemistry: A Primer for Earth System Scientists, pages 37–56, Springer. doi:10.1007/978-3-030-10822-9_3. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Legendre L, Rassoulzadegan F (1995) "Plankton and nutrient dynamics in marine waters". Ofélie, 41:153–172.
- ^ Pace ML, Cole JJ, Carpenter SR, Kitchell JF (1999) "Trophic cascades revealed in diverse ecosystems". Trendy Ecol Evol, 14: 483–488.
- ^ Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F (1983) "The ecological role of water-column microbes in the sea". Mar Ecol-Prog Ser, 10: 257–263.
- ^ Anderson TR and LeB Williams PJ (1998) "Modelling the seasonal cycle of dissolved organic carbonat station E1 in the English channel". Estuar Coast Shelf Sci, 46: 93–109.
- ^ Van den Meersche K, Middelburg JJ, Soetaert K, van Rijswijk P, Boschker HTS, Heip CHR (2004) "Carbon–nitrogen coupling and algal–bacterial interactions during an experimental bloom: modeling a 13C tracer experiment". Limnol Oceanogr, 49: 862–878.
- ^ Suttle CA (2005) "Viruses in the sea". Příroda, 437: 356–361.
- ^ Siegel, David A.; Buesseler, Ken O .; Behrenfeld, Michael J.; Benitez-Nelson, Claudia R .; Boss, Emmanuel; Brzezinski, Mark A.; Burd, Adrian; Carlson, Craig A .; d'Asaro, Eric A.; Doney, Scott C .; Perry, Mary J.; Stanley, Rachel H. R.; Steinberg, Deborah K. (2016). "Prediction of the Export and Fate of Global Ocean Net Primary Production: The EXPORTS Science Plan". Frontiers in Marine Science. 3. doi:10.3389/fmars.2016.00022. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Wang, F., Wu, Y., Chen, Z., Zhang, G., Zhang, J., Zheng, S. and Kattner, G. (2019) "Trophic interactions of mesopelagic fishes in the South China Sea illustrated by stable isotopes and fatty acids". Frontiers in Marine Science, 5: 522. doi:10.3389/fmars.2018.00522.
- ^ A b Tollefson, Jeff (27 February 2020) Enter the twilight zone: scientists dive into the oceans’ mysterious middle Zprávy o přírodě. doi:10.1038/d41586-020-00520-8.
- ^ A b Choy, C.A., Haddock, S.H. and Robison, B.H. (2017) "Deep pelagic food web structure as revealed by in situ feeding observations". Sborník Královské společnosti B: Biologické vědy, 284(1868): 20172116. doi:10.1098/rspb.2017.2116. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Kelly, T.B., Davison, P.C., Goericke, R., Landry, M.R., Ohman, M. and Stukel, M.R. (2019) "The importance of mesozooplankton diel vertical migration for sustaining a mesopelagic food web". Frontiers in Marine Science, 6: 508. doi:10.3389/fmars.2019.00508.
- ^ Změna klimatu v hlubokých oceánech může být do poloviny století sedmkrát rychlejší, uvádí se ve zprávě Opatrovník, 25. května 2020.
- ^ Brito-Morales, I., Schoeman, D.S., Molinos, J.G., Burrows, M.T., Klein, C.J., Arafeh-Dalmau, N., Kaschner, K., Garilao, C., Kesner-Reyes, K. a Richardson, A.J. (2020) „Rychlost klimatu odhaluje rostoucí vystavení hlubinné biologické rozmanitosti budoucímu oteplování“. Přírodní změna podnebí, str. 1-6. doi:10,5281 / zenodo.3596584.
- ^ Irigoien, X., Klevjer, T.A., Røstad, A., Martinez, U., Boyra, G., Acuña, J.L., Bode, A., Echevarria, F., Gonzalez-Gordillo, J.I., Hernandez-Leon, S. and Agusti, S. (2014) "Large mesopelagic fishes biomass and trophic efficiency in the open ocean". Příroda komunikace, 5: 3271. doi:10.1038 / ncomms4271
- ^ Fish biomass in the ocean is 10 times higher than estimated EurekAlert, 7. února 2014.
- ^ Choy, C.A., Wabnitz, C.C., Weijerman, M., Woodworth-Jefcoats, P.A. and Polovina, J.J. (2016) "Finding the way to the top: how the composition of oceanic mid-trophic micronekton groups determines apex predator biomass in the central North Pacific". Série pokroku v ekologii moří, 549: 9–25. doi:10.3354/meps11680.
- ^ A b C d E Kurata, N., Vella, K., Hamilton, B., Shivji, M., Soloviev, A., Matt, S., Tartar, A. and Perrie, W. (2016) "Surfactant-associated bacteria in the near-surface layer of the ocean". Nature: Scientific Reports, 6(1): 1–8. doi:10.1038/srep19123. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Ẑutić, V., Ćosović, B., Marčenko, E., Bihari, N. and Kršinić, F. (1981) "Surfactant production by marine phytoplankton". Marine Chemistry, 10(6): 505–520. doi:10.1016/0304-4203(81)90004-9.
- ^ Levin LA, Baco AR, Bowden DA, Colaco A, Cordes EE, Cunha MR, Demopoulos AWJ, Gobin J, Grupe BM, Le J, Metaxas A, Netburn AN, Rouse GW, Thurber AR, Tunnicliffe V, Van Dover CL, Vanreusel A and Watling L (2016). "Hydrothermal Vents and Methane Seeps: Rethinking the Sphere of Influence". Přední. Mar. Sci. 3:72. doi:10.3389/fmars.2016.00072
- ^ Portail, M., Olu, K., Dubois, S.F., Escobar-Briones, E., Gelinas, Y., Menot, L. and Sarrazin, J. (2016). "Food-web complexity in Guaymas Basin hydrothermal vents and cold seeps". PLOS ONE, 11(9): p.e0162263. doi:10.1371/journal.pone.0162263.
- ^ Bernardino AF, Levin LA, Thurber AR and Smith CR (2012). "Comparative Composition, Diversity and Trophic Ecology of Sediment Macrofauna at Vents, Seeps and Organic Falls". PLOS ONE, 7(4): e33515. pmid:22496753. doi:10.1371/journal.pone.0033515.
- ^ Portail M, Olu K, Escobar-Briones E, Caprais JC, Menot L, Waeles M, et al. (2015). "Comparative study of vent and seep macrofaunal communities in the Guaymas Basin". Biogeovědy. 12(18): 5455–79. doi:10.5194/bg-12-5455-2015.
- ^ A b Otero, X.L., De La Peña-Lastra, S., Pérez-Alberti, A., Ferreira, T.O. and Huerta-Diaz, M.A. (2018) "Seabird colonies as important global drivers in the nitrogen and phosphorus cycles". Příroda komunikace, 9(1): 1–8. doi:10.1038/s41467-017-02446-8. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Harris, P.T .; Macmillan-Lawler, M .; Rupp, J .; Baker, E.K. (2014). „Geomorfologie oceánů“. Mořská geologie. 352: 4–24. doi:10.1016 / j.margeo.2014.01.011.
- ^ Heymans, J.J., Coll, M., Libralato, S., Morissette, L. and Christensen, V. (2014). "Global patterns in ecological indicators of marine food webs: a modelling approach". PLOS ONE, 9(4). doi:10.1371/journal.pone.0095845.
- ^ Pranovi, F., Libralato, S., Raicevich, S., Granzotto, A., Pastres, R. and Giovanardi, O. (2003). "Mechanical clam dredging in Venice lagoon: ecosystem effects evaluated with a trophic mass-balance model". Mořská biologie, 143(2): 393–403. doi:10.1007/s00227-003-1072-1.
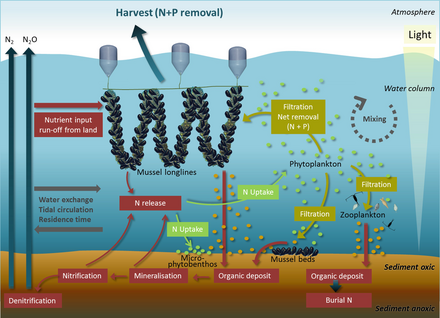
- ^ Petersen, J.K., Holmer, M., Termansen, M. and Hasler, B. (2019) "Nutrient extraction through bivalves". In: Smaal A., Ferreira J., Grant J., Petersen J., Strand Ø. (eds) Goods and Services of Marine Bivalves, pages 179–208. Springer. doi:10.1007/978-3-319-96776-9_10. ISBN 9783319967769
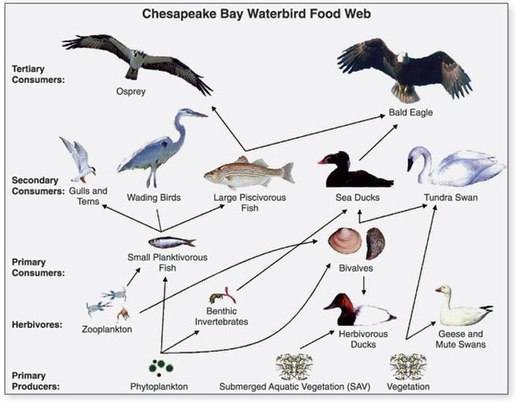
- ^ US Geological Survey (USGS). "Chapter 14: Changes in Food and Habitats of Waterbirds." Figure 14.1. Synthesis of U.S. Geological Survey Science for the Chesapeake Bay Ecosystem and Implications for Environmental Management. USGS Circular 1316.
 Tento článek včlení text z tohoto zdroje, který je v veřejná doména.
Tento článek včlení text z tohoto zdroje, který je v veřejná doména. - ^ Perry, M.C., Osenton, P.C., Wells-Berlin, A.M., and Kidwell, D.M., 2005, Food selection among Atlantic Coast sea ducks in relation to historic food habits, [abs.] in Perry, M.C., Second North American Sea Duck Conference, November 7–11, 2005, Annapolis, Maryland, Program and Abstracts, USGS Patuxent Wildlife Research Center, Maryland, 123 p. (p. 105).
- ^ Bowser, A.K., Diamond, A.W. and Addison, J.A. (2013) "From puffins to plankton: a DNA-based analysis of a seabird food chain in the northern Gulf of Maine". PLOS ONE, 8(12): e83152. doi:10.1371/journal.pone.0083152
- ^ Leray M, Meyer CP, Mills SC. (2015) „Metabarcodingová dietní analýza dravých ryb žijících v korálech demonstruje malý příspěvek korálových vzájemníků k jejich vysoce rozdělené, všeobecné stravě“. PeerJ, 3: e1047. doi:10,7717 / peerj.1047.
- ^ Stat, M., Huggett, M. J., Bernasconi, R., DiBattista, J. D., Berry, T. E., Newman, S. J., Harvey, E.S. a Bunce, M. (2017) „Ekosystémový biomonitoring s eDNA: metabarcoding přes strom života v tropickém mořském prostředí“. Vědecké zprávy, 7(1): 1–11. doi:10.1038 / s41598-017-12501-5.
- ^ Lobo, Eduardo A .; Heinrich, Carla Giselda; Schuch, Marilia; Wetzel, Carlos Eduardo; Ector, Luc (2016), Necchi JR, Orlando (ed.), „Diatoms as Bioindicators in Rivers“, Říční řasy, Springer International Publishing, s. 245–271, doi:10.1007/978-3-319-31984-1_11, ISBN 9783319319834
- ^ Stevenson, R. Jan; Pan, Yangdong; van Dam, Herman (2010), Smol, John P .; Stoermer, Eugene F. (eds.), "Posuzování podmínek prostředí v řekách a potokech s rozsivkami", Diatomy (2. vyd.), Cambridge University Press, str. 57–85, doi:10.1017 / cbo9780511763175.005, ISBN 9780511763175
- ^ Coll, M., Schmidt, A., Romanuk, T. a Lotze, H.K. (2011). „Struktura potravinových webů komunit mořských řas v různých prostorových měřítcích a dopadech člověka“. PLOS ONE, 6(7): e22591. doi:10.1371 / journal.pone.0022591. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ McCarthy, J.J., Canziani, O.F., Leary, N.A., Dokken, D.J. and White, K.S. (Eds.) (2001) Změna klimatu 2001: Dopady, přizpůsobení a zranitelnost: Příspěvek pracovní skupiny II ke třetí hodnotící zprávě Mezivládního panelu pro změnu klimatu Strana 807, Cambridge University Press. ISBN 9780521015004
- ^ Murphy, EJ, Cavanagh, RD, Drinkwater, KF, Grant, SM, Heymans, JJ, Hofmann, EE, Hunt Jr, GL a Johnston, NM (2016) „Porozumění struktuře a fungování polárních pelagických ekosystémů za účelem předpovědi dopadů změna". Sborník Královské společnosti B: Biologické vědy, 283(1844): 20161646. doi:10.1098 / rspb.2016.1646.
- ^ Dopady klimatu na ekosystémy: narušení potravinového webu EPA. Zpřístupněno 11. února 2020. Tento článek včlení text z tohoto zdroje, který je v veřejná doména.
- ^ „V Severním ledovém oceánu dochází k„ změně režimu “, říkají vědci“. phys.org. Citováno 16. srpna 2020.
- ^ Lewis, K. M .; Dijken, G. L. van; Arrigo, K. R. (10. července 2020). „Změny v koncentraci fytoplanktonu nyní vedou ke zvýšení primární produkce v Severním ledovém oceánu“. Věda. 369 (6500): 198–202. doi:10.1126 / science.aay8380. ISSN 0036-8075. PMID 32647002. S2CID 220433818. Citováno 16. srpna 2020.
- ^ Stoecker, D.K. a Lavrentyev, P.J. (2018). „Mixotrophic plankton in the polar sea: a pan-arctic review“. Frontiers in Marine Science, 5: 292. doi:10.3389 / fmars.2018.00292
- ^ Kilias, Estelle S .; Junges, Leandro; Šupraha, Luka; Leonard, Guy; Metfies, Katja; Richards, Thomas A. (2020). „Chytridové houby a jejich společný výskyt s rozsivkami korelují s táním mořského ledu v Severním ledovém oceánu“. Komunikační biologie. 3 (1): 183. doi:10.1038 / s42003-020-0891-7. PMC 7174370. PMID 32317738. S2CID 216033140. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Bender, S.J., Moran, D.M., McIlvin, M.R., Zheng, H., McCrow, J.P., Badger, J., DiTullio, G.R., Allen, A.E. a Saito, M.A. (2018) „Tvorba kolonií v Phaeocystis antarctica: propojení molekulárních mechanismů s biogeochemií železa ". Biogeovědy, 15(16): 4923–4942. doi:10.5194 / bg-15-4923-2018.
- ^ Pinkernell, S. a Beszteri, B. (2014) „Potenciální účinky změny klimatu na distribuční rozsah hlavního silikátového platinu v jižním oceánu“. Ekologie a evoluce, 4(16): 3147–3161. doi:10.1002 / ece3.1138
- ^ Cavan, EL, Belcher, A., Atkinson, A., Hill, SL, Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. a Steinberg, DK (2019) „Význam antarktického krilu v biogeochemických cyklech“. Příroda komunikace, 10(1): 1–13. doi:10.1038 / s41467-019-12668-7. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ A b Cordone, G., Marina, T.I., Salinas, V., Doyle, S.R., Saravia, L.A. and Momo, F.R. (2018). „Účinky ztráty makrořas v antarktickém mořském potravinářském webu: uplatnění prahů vyhynutí na studie potravinářských webů“. PeerJ, 6: e5531. doi:10,7717 / peerj.5531
- ^ Marina, T.I., Salinas, V., Cordone, G., Campana, G., Moreira, E., Deregibus, D., Torre, L., Sahade, R., Tatian, M., Oro, E.B. a De Troch, M. (2018). „Potravinová síť Potter Cove (Antarktida): složitost, struktura a funkce“. Estuarine, Coastal and Shelf Science, 200: 141–151. doi: 10.1016 / j.ecss.2017.10.015.
- ^ Koh, E.Y., Martin, A.R., McMinn, A. a Ryan, K.G. (2012) „Nedávné pokroky a budoucí perspektivy mikrobiální fototrofie v antarktickém mořském ledu“. Biologie, 1(3): 542-556. doi:10,3390 / biologie1030542.
- ^ Mikrobiální smyčka je zde znovu nakreslena a zkrácena z:
- Azam, F., Fenchel, T., Field, J.G., Gray, J.S., Meyer-Reil, L.A. a Thingstad, F. (1983) „Ekologická role mikrobů ve vodním sloupci v moři“. Řada pokroku mořské ekologie, 10(3): 257–263.
- Fenchel, T. (2008) „Mikrobiální smyčka - o 25 let později“. Journal of Experimental Marine Biology and Ecology, 366(1-2): 99-103. doi:10.1016 / j.jembe.2008.07.013.
- ^ Lamy, T., Koenigs, C., Holbrook, S.J., Miller, R.J., Stier, A.C. a Reed, D.C. (2020) „Druhy základů podporují stabilitu komunity zvyšováním rozmanitosti v obřím řase“. Ekologie, e02987. doi:10,1002 / ecy.2987.
- ^ Obří řasa dává mořským ekosystémům jižní Kalifornie silný základ, Národní vědecká nadace, 4. února 2020.
- ^ Angelini, Christine; Altieri, Andrew H .; et al. (Říjen 2011). „Interakce mezi základními druhy a jejich důsledky pro organizaci komunity, biologickou rozmanitost a ochranu přírody“. BioScience. 61 (10): 782–789. doi:10.1525 / bio.2011.61.10.8.
- ^ Ellison, Aaron M .; Bank, Michael S .; et al. (Listopad 2005). „Ztráta základových druhů: důsledky pro strukturu a dynamiku zalesněných ekosystémů“. Hranice v ekologii a životním prostředí. 3 (9): 479–486. doi:10.1890 / 1540-9295 (2005) 003 [0479: LOFSCF] 2.0.CO; 2.
- ^ Dayton, P. K. 1972. Směrem k pochopení odolnosti komunity a možných účinků obohacení bentosu v McMurdo Sound v Antarktidě. str. 81–96 ve sborníku kolokvia o problémech ochrany přírody Allen Press, Lawrence, Kansas.
- ^ Paine, R. T. (1995). „Konverzace o zdokonalení koncepce druhů lichoběžníkového zkreslení“. Biologie ochrany. 9 (4): 962–964. doi:10.1046 / j.1523-1739.1995.09040962.x.
- ^ Davic, Robert D. (2003). „Propojení klíčových druhů a funkčních skupin: nová operativní definice konceptu klíčových druhů“. Ekologie ochrany. Citováno 2011-02-03.
- ^ Paine, R. T. (1969). „Poznámka o trofické složitosti a stabilitě komunity“. Americký přírodovědec. 103 (929): 91–93. doi:10.1086/282586. JSTOR 2459472. S2CID 83780992.
- ^ „Keystone Species Hypothesis“. University of Washington. Archivovány od originál dne 10.01.2011. Citováno 2011-02-03.
- ^ Paine, R. T. (1966). "Složitost potravinového webu a druhová rozmanitost". Americký přírodovědec. 100 (910): 65–75. doi:10.1086/282400. JSTOR 2459379. S2CID 85265656.
- ^ Szpak, Paul; Orchard, Trevor J .; Salomon, Anne K .; Gröcke, Darren R. (2013). „Regionální ekologická variabilita a dopad obchodu s mořskými kožešinami na pobřežní ekosystémy na jihu Haida Gwaii (Britská Kolumbie, Kanada): důkazy ze stabilní izotopové analýzy kostního kolagenu skalních ryb (Sebastes spp.)“ Archeologické a antropologické vědy. 5 (2): 159–182. doi:10.1007 / s12520-013-0122-r. S2CID 84866250.
- ^ Cohn, J. P. (1998). „Porozumění vydřím mořským“. BioScience. 48 (3): 151–155. doi:10.2307/1313259. JSTOR 1313259.
- ^ A b C Millette, N.C., Grosse, J., Johnson, W.M., Jungbluth, M.J. a Suter, E.A. (2018). „Skryté na očích: Význam kryptických interakcí v mořském planktonu“. Limnologické a oceánografické dopisy, 3(4): 341–356. doi:10.1002 / lol2.10084. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
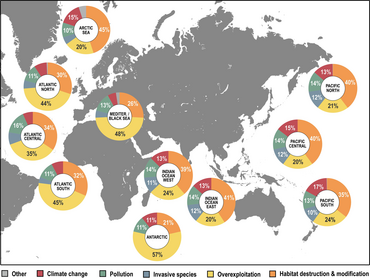
- ^ A b C Luypaert, T., Hagan, J.G., McCarthy, M.L. a Poti, M. (2020) „Status of Marine Biodiversity in the Anthropocene“. V: YOUMARES 9-The Oceans: Our Research, Our Future, strany 57-82, Springer. doi:10.1007/978-3-030-20389-4_4.
- ^ Estes JA, Tinker MT, Williams TM a kol. (1998) „Predace kosatek na mořských vydrách spojujících oceánské a pobřežní ekosystémy“. Věda, 282: 473–476. doi:10.1126 / science.282.5388.473.
- ^ Post, D. M. (1993). „Dlouhá a krátká délka potravinového řetězce“. Trendy v ekologii a evoluci. 17 (6): 269–277. doi:10.1016 / S0169-5347 (02) 02455-2.
- ^ Jerry Bobrow, Ph.D.; Stephen Fisher (2009). CliffsNotes CSET: Více předmětů (2. vyd.). John Wiley and Sons. str. 283. ISBN 978-0-470-45546-3.
- ^ Elton CS (1927) Ekologie zvířat. Publikováno 2001. University of Chicago Press.
- ^ Paine RT (1966). "Složitost potravinového webu a druhová rozmanitost". Americký přírodovědec. 100 (910): 65–75. doi:10.1086/282400. S2CID 85265656.
- ^ A b Května RM (2001) Stabilita a složitost v modelových ekosystémech Princeton University Press, dotisk vydání z roku 1973 s novou předmluvou. ISBN 978-0-691-08861-7.
- ^ A b Pimm SL (2002) Webové stránky o jídle University of Chicago Press, dotisk vydání z roku 1982 s novou předmluvou. ISBN 978-0-226-66832-1.
- ^ Kotta, J .; Wernberg, T .; Jänes, H .; Kotta, I .; Nurkse, K .; Pärnoja, M .; Orav-Kotta, H. (2018). „Nový krabový dravec způsobuje posun režimu mořského ekosystému“. Vědecké zprávy. 8 (1): 4956. doi:10.1038 / s41598-018-23282-w. PMC 5897427. PMID 29651152.
- ^ Megrey, Bernard a Werner, Francisco. „Hodnocení role regulace ekosystémů shora dolů vs. zdola nahoru z pohledu modelování“ (PDF).CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Frank, K. T .; Petrie, B .; Choi, J. S .; Leggett, W. C. (2005). „Trofické kaskády v dřívějším ekosystému ovládaném treskami“. Věda. 308 (5728): 1621–1623. doi:10.1126 / science.1113075. ISSN 0036-8075. PMID 15947186. S2CID 45088691.
- ^ Matsuzaki, Shin-Ichiro S .; Suzuki, Kenta; Kadoya, Taku; Nakagawa, Megumi; Takamura, Noriko (2018). „Vazby zdola nahoru mezi primární produkcí, zooplanktonem a rybami v mělkém hypereutrofním jezeře“. Ekologie. 99 (9): 2025–2036. doi:10,1002 / ecy.2414. PMID 29884987.
- ^ Lynam, Christopher Philip; Llope, Marcos; Möllmann, Christian; Helaouët, Pierre; Bayliss-Brown, Georgia Anne; Stenseth, Nils C. (únor 2017). „Trofická a environmentální kontrola v Severním moři“. Sborník Národní akademie věd. 114 (8): 1952–1957. doi:10.1073 / pnas.1621037114. PMC 5338359. PMID 28167770.
- ^ „Oldlist“. Výzkum prstenů na skalnatých horách. Citováno 8. ledna 2013.
- ^ Wetzel, Robert (2001). Limnologie: Jezerní a říční ekosystémy. San Diego: Academic Press. ISBN 9780127447605.
- ^ Bar-On, Y.M., Phillips, R. a Milo, R. (2018) „The biomass distribution on Earth“. Sborník Národní akademie věd, 115(25): 6506–6511. doi:10.1073 / pnas.1711842115.
- ^ Spellman, Frank R. (2008). The Science of Water: Concepts and Applications. CRC Press. str. 167. ISBN 978-1-4200-5544-3.
- ^ Odum, E. P .; Barrett, G. W. (2005). Základy ekologie (5. vydání). Brooks / Cole, součást Cengage Learning. ISBN 978-0-534-42066-6. Archivovány od originál dne 20.08.2011.
- ^ Wang, H .; Morrison, W .; Singh, A .; Weiss, H. (2009). „Modelování obrácených pyramid a útočišť z biomasy v ekosystémech“ (PDF). Ekologické modelování. 220 (11): 1376–1382. doi:10.1016 / j.ecolmodel.2009.03.005. Archivovány od originál (PDF) dne 07.10.2011.
- ^ Field, C.B., Behrenfeld, M.J., Randerson, J.T. a Falkowski, P. (1998) „Primární produkce biosféry: integrace suchozemských a oceánských složek“. Věda, 281(5374): 237–240. doi:10.1126 / science.281.5374.237.
- ^ Maureaud, A., Gascuel, D., Colléter, M., Palomares, M.L., Du Pontavice, H., Pauly, D. a Cheung, W.W. (2017) „Globální změna v trofickém fungování sítí mořských potravin“. PLOS ONE, 12(8): e0182826. doi:10.1371 / journal.pone.0182826
- ^ „Účinky acidifikace oceánů na mořské druhy a ekosystémy“. Zpráva. OCEANA. Citováno 13. října 2013.
- ^ „Komplexní studie okyselování v Severním ledovém oceánu“. Studie. CICERO. Archivovány od originál dne 10. prosince 2013. Citováno 14. listopadu 2013.
- ^ Lischka, S .; Büdenbender J .; Boxhammer T .; Riebesell U. (15. dubna 2011). „Dopad okyselování oceánů a zvýšených teplot na časné mladistvé pteropoda Limacina helicina: úhyn, degradace skořápky a růst skořápky“ (PDF). Zpráva. Biogeovědy. 919–932. Citováno 14. listopadu 2013.
- ^ „Antarktická mořská divočina je ohrožena, uvádí studie“. BBC Příroda. Citováno 13. října 2013.
- ^ Ullah, H., Nagelkerken, I., Goldenberg, S.U. a Fordham, D.A. (2018) „Změna klimatu by mohla vést ke kolapsu mořské potravní sítě prostřednictvím změněných trofických toků a šíření sinic“. Biologie PLoS, 16(1): e2003446. doi:10.1371 / journal.pbio.2003446
- ^ A b Změna klimatu vede ke kolapsu mořských potravinářských webů ScienceDaily. 9. ledna 2018.
- ^ IUCN (2018) Červený seznam ohrožených druhů IUCN: verze 2018-1