Mořští bezobratlí - Marine invertebrates

Mořští bezobratlí jsou bezobratlých kteří žijí v mořská stanoviště. Bezobratlý je všeobecný termín to zahrnuje vše zvířata kromě obratlovců členové strunat kmen. Bezobratlí chybí páteř a některé se vyvinuly a skořápka nebo tvrdý exoskeleton. Stejně jako na zemi i ve vzduchu mají mořští bezobratlí širokou škálu plány těla a byly rozděleny do více než 30 phyla. Tvoří většinu makroskopického života v oceánech.
Vývoj
| Část série přehledů o |
| mořský život |
|---|
Nejdříve zvířata byli mořští bezobratlých, to znamená, obratlovců přišel později. Zvířata jsou mnohobuňečný eukaryoty,[poznámka 1] a liší se od rostlin, řas a hub tím, že chybí buněčné stěny.[1] Mořští bezobratlí jsou zvířata, která obývají a námořní prostředí kromě členů obratlovců strunat kmen; bezobratlým chybí a páteř. Některé se vyvinuly a skořápka nebo tvrdý exoskeleton.
Nejčasnější zvířata mohou patřit do rodu Dickinsonia,[2] Před 571 miliony až 541 miliony let.[3] Individuální Dickinsonia typicky připomínají oboustranně symetrický žebrovaný ovál. Stále rostly, dokud nebyly pokryty sedimenty nebo jinak zabity,[4] a strávili většinu svého života s těly pevně ukotvenými v sedimentu.[5] Jejich taxonomické spříznění jsou v současné době neznámé, ale jejich způsob růstu je shodný s a bilaterální afinita.[6]
Na rozdíl od Dickinsonia, nejdříve široce přijímané zvířecí fosilie jsou velmi moderní cnidarians (skupina, která zahrnuje Medúza, mořské sasanky a Hydra ), možná z okolí 580 Ma[7] The Ediacara biota, který vzkvétal posledních 40 milionů let před začátkem EU Kambrijský,[8] byli prvními zvířaty o délce více než jen pár centimetrů. Jako Dickinsonia, mnoho z nich bylo plochých s „prošívaným“ vzhledem a vypadalo to tak divně, že existoval návrh klasifikovat je jako samostatné království, Vendozoa.[9] Jiné však byly interpretovány již dříve měkkýši (Kimberella[10][11]), ostnokožci (Arkarua[12]), a členovci (Spriggina,[13] Parvancorina[14]). O klasifikaci těchto vzorků stále existuje debata, zejména proto, že diagnostické vlastnosti, které umožňují taxonomům klasifikovat novější organismy, jako jsou podobnosti s živými organismy, v Ediacarans obecně chybí. Zdá se však, že o tom není pochyb Kimberella byl alespoň a triploblastický bilaterální zvíře, jinými slovy, zvíře podstatně složitější než cnidariáni.[15]
The malá šupinatá fauna jsou velmi smíšenou sbírkou fosilií nalezených mezi pozdním Ediacaranem a Střední kambrie období. Nejdříve, Cloudina, vykazuje známky úspěšné obrany proti predaci a může označovat začátek evoluční závod ve zbrojení. Některé drobné rané kambrické mušle téměř jistě patřily měkkýšům, zatímco majitelé některých „pancéřových desek“ Halkieria a Microdictyon, byly nakonec identifikovány, když byly v kambriu nalezeny úplnější vzorky lagerstätten která zachovala zvířata s měkkým tělem.[16]

Dickinsonia může být nejranějším zvířetem. Objevují se ve fosilním záznamu před 571 až 541 miliony let.

Kimberella, raný měkkýš důležitý pro pochopení Kambrijská exploze. Bezobratlí jsou seskupeni do různých phyla (plány těla ).

Opabinie, zaniklý kmenová skupina členovec se objevil v Střední kambrie[17]:124–136



V 70. letech se již vedla debata o tom, zda vznik moderní fyly byl „výbušný“ nebo postupný, ale skrytý nedostatkem Precambrian fosilie zvířat.[16] Opětovná analýza fosilií z Burgess Shale lagerstätte zvýšil zájem o tuto problematiku, když odhalila zvířata, jako např Opabinie, který se nehodil do žádného známého kmen. V té době to byly interpretovány jako důkaz, že moderní phyla se velmi rychle vyvinula v kambrijské explozi a že „divné divy“ Burgess Shale ukázaly, že raná kambrie byla jedinečným experimentálním obdobím vývoje zvířat.[17] Pozdější objevy podobných zvířat a vývoj nových teoretických přístupů vedly k závěru, že mnoho „divných divů“ byly evoluční „tety“ nebo „bratranci“ moderních skupin[18]—Například to Opabinie byl členem lobopody, což je skupina, která zahrnuje předky členovců a která mohla úzce souviset s moderní dobou tardigrades.[19] Přesto stále existuje mnoho debat o tom, zda kambrijská exploze byla skutečně výbušná, a pokud ano, jak a proč k ní došlo a proč se zdá být v historii zvířat jedinečná.[20]
Klasifikace

Bezobratlí jsou seskupeni do různých phyla. Neformálně lze phylu považovat za způsob seskupování organismů podle jejich tělesný plán.[21][22]:33 Plán těla odkazuje na plán, který popisuje tvar nebo morfologie organismu, jako je jeho symetrie, segmentace a jeho dispozice přílohy. Myšlenka tělesných plánů vznikla obratlovců, které byly seskupeny do jednoho kmene. Plán těla obratlovců je však pouze jedním z mnoha a bezobratlí se skládají z mnoha plánů phyly nebo těla. Na historii objevování tělních plánů lze pohlížet jako na pohyb od světonázoru zaměřeného na obratlovce, až po vidění obratlovců jako jednoho tělesného plánu mezi mnoha. Mezi průkopníky zoologové „Linné identifikoval dva plány těla mimo obratlovce; Cuvier identifikoval tři; a Haeckel měl čtyři, stejně jako Protista s osmi dalšími, celkem dvanáct. Pro srovnání, počet kmenů uznaných moderními zoology je stoupl na 35.[22]
Historicky byly plány těla považovány za plány, které se rychle vyvinuly během Kambrijská exploze,[23] ale jemnější chápání evoluce zvířat naznačuje postupný vývoj tělesných plánů v raných fázích Paleozoikum a za.[24] Obecněji lze kmen definovat dvěma způsoby: jak je popsáno výše, jako skupina organismů s určitým stupněm morfologické nebo vývojové podobnosti ( fenetický definice), nebo skupina organismů s určitým stupněm evoluční příbuznosti (dále jen fylogenetické definice).[24]
Stejně jako na zemi i ve vzduchu tvoří bezobratlí velkou většinu veškerého makroskopického života, protože obratlovci tvoří podkmen jednoho z více než 30 známých živočišných kmenů, takže tento výraz nemá téměř žádný význam taxonomické účel. Mořský život bezobratlých zahrnuje následující skupiny, z nichž některé jsou kmeny:


- Acoela, mezi nejprimitivnější bilaterální zvířata;
- Annelida, (mnohoštětinatci a moře pijavice );
- Brachiopoda, mořští živočichové, kteří mají na horním a dolním povrchu tvrdé „chlopně“ (mušle);
- Bryozoa, známé také jako mechová zvířata nebo mořské rohože;
- Chaetognatha, běžně známí jako červi šípu, jsou kmenem dravých mořských červů, kteří jsou hlavní složkou planktonu;
- Cephalochordata zastoupené v moderních oceánech lancelety (také známý jako Amphioxus);
- Cnidaria, jako Medúza, mořské sasanky, a korály;
- Korýš, počítaje v to humři, kraby, krevety, rak, barnacles, krabi poustevníka, mantis krevety, a copepods;
- Ctenophora, známá také jako hřebenová želé, největší zvířata, která plavou pomocí řasinek;
- Ostnokožci, počítaje v to mořské hvězdy, křehké hvězdy, mořští ježci, pískové dolary, mořské okurky, krinoidy, a mořské sedmikrásky;
- Echiura, také známí jako lžíční červi;
- Gnathostomulids, štíhlé až nitkovité červy, s průhledným tělem, které obývá písek a bahno pod mělkými pobřežními vodami;
- Gastrotricha, často nazývané chlupaté zády, se nacházejí převážně intersticiálně mezi částicemi sedimentu;
- Hemichordata, zahrnuje žalud červi, osamělé červí organizmy;
- Kamptozoa, sedací vodní živočichové ve tvaru poháru, s relativně dlouhými stonky a „korunou“ pevných chapadel, nazývanou také Entoprocta;
- Kinorhyncha segmentovaná zvířata bez končetin, rozšířená v bahně nebo písku ve všech hloubkách, nazývaná také bahenní draci;
- Loricifera, velmi malá až mikroskopická mořská zvířata žijící v sedimentech objevená až v roce 1983;
- Měkkýš, počítaje v to měkkýši, oliheň, chobotnice, surmovky, Nautilus, sépie obecná, nudibranchs, lastury, mořské šneky, Aplacophora, Caudofoveata, Monoplacophora, Polyplacophora, a Scaphopoda;
- Myzostomida, taxonomická skupina malých mořských červů, kteří parazitují na krinoidech nebo „mořských liliích“;
- Nemertinea, známé také jako „pásoví červi“ nebo „proboscis červi“;
- Orthonectida, malý kmen špatně známých parazitů mořských bezobratlých, kteří patří mezi nejjednodušší z mnohobuněčných organismů;
- Phoronida, kmen mořských živočichů, kteří filtrují krmení s lophophore ("korunou" chapadel) a staví svislé chitinové trubice na podporu a ochranu jejich měkkých těl;
- Placozoa, malá, zploštělá, mnohobuněčná zvířata s průměrem přibližně 1 milimetr a nejjednodušší strukturou. Nemají pravidelný obrys, i když spodní povrch je poněkud konkávní a horní povrch je vždy zploštělý;
- Porifera (houby), mnohobuněčné organismy, které mají těla plná pórů a kanálů umožňujících cirkulaci vody;
- Priapulida nebo červi penisu jsou kmenem mořských červů, kteří žijí v mořském bahně. Oni jsou jmenováni pro jejich rozšiřitelnou ostnatou proboscis, která u některých druhů může mít tvar jako tvar lidského penisu;
- Pycnogonida, také nazývaní mořští pavouci, nesouvisí s pavouky, ani s pavouky, kterým se podobají;
- Sipunculida, také nazývaná arašídové červy, je skupina obsahující 144–320 druhů (odhady se liší) dvoustranně symetrických, nesegmentovaných mořských červů;
- Tunicata, známé také jako mořské stříkance nebo vepřové maso, jsou krmítka filtrů připojená ke skalám nebo podobně vhodným povrchům na dně oceánu;
- Nějaký ploché červy tříd Turbellaria a Monogenea;
- Xenoturbella, rod bilaterálních zvířat, který obsahuje pouze dva druhy podobné mořským červům;
- Xiphosura, zahrnuje velké množství vyhynulých linií a pouze čtyři nedávné druhy z čeledi Limulidae, mezi něž patří i krabi podkovy.
Členovci celkem asi 1113 000 popsaných existujících druhů, měkkýši asi 85 000 a strunatci asi 52 000.[25]
Mořské houby

Houby jsou zvířata kmen Porifera (moderní latina pro nesoucí póry [26]). Jsou to mnohobuněčné organismy, které mají těla plná pórů a kanálů umožňujících cirkulaci vody skrz ně, skládající se z želé mesohyl vložené mezi dvě tenké vrstvy buňky. Mají nespecializované buňky, které mohou transformovat do jiných typů a které v tomto procesu často migrují mezi hlavními buněčnými vrstvami a mezohylem. Houby nemají nervový, zažívací nebo oběhové systémy. Místo toho se většina spoléhá na udržování stálého průtoku vody tělem, aby získala jídlo a kyslík a odstranila odpady.
Houby jsou podobné ostatním zvířatům v tom, že jsou mnohobuňečný, heterotrofní, nedostatek buněčné stěny a vyrábět spermie. Na rozdíl od jiných zvířat jim chybí pravda papírové kapesníky a orgány, a nemají žádné symetrie těla. Tvary jejich těl jsou přizpůsobeny pro maximální účinnost proudění vody centrální dutinou, kde ukládá živiny, a odchází otvorem zvaným osculum. Mnoho hub má vnitřní kostry spongin a / nebo spikuly z uhličitan vápenatý nebo oxid křemičitý. Všechny houby jsou přisedlý vodní živočichové. I když existují sladkovodní druhy, velká většina jsou mořské (slané) druhy, od přílivových zón do hloubek přesahujících 8 800 m (5,5 mil).
Zatímco většina z přibližně 5 000–10 000 známých druhů se živí bakterie a další částice potravy ve vodě, někteří hostitelé fotosyntéza mikroorganismy jako endosymbionty a tato aliance často produkují více jídla a kyslíku, než kolik spotřebují. Stalo se několik druhů hub, které žijí v prostředí chudém na potraviny masožravci že kořist hlavně na malé korýši.[27]

Houba biologická rozmanitost. Na této fotografii jsou čtyři druhy hub.

Květinový koš Venuše v hloubce 2572 metrů





Linné omylem identifikoval houby jako rostliny v pořadí Řasy.[28] Po dlouhou dobu byly houby přiřazeny k samostatnému subkultuře, Parazoa (význam vedle zvířat).[29] Nyní jsou klasifikovány jako paraphyletic kmen ze kterých se vyvinula vyšší zvířata.[30]
Mořští cnidarians

Cnidarians (Řek pro kopřiva) se vyznačují přítomností bodavé buňky, specializované buňky, které používají hlavně k zachycení kořisti. Cnidarians patří korály, mořské sasanky, Medúza a hydrozoans. Tvoří a kmen obsahující přes 10 000[31] druh z zvířata nacházejí se výhradně ve vodním (hlavně mořském) prostředí. Jejich těla se skládají z mezoglea, neživá rosolovitá látka, vložená mezi dvě vrstvy epitel které jsou většinou jedno buňka tlustá. Mají dvě základní tělesné formy: plavání medusae a přisedlý polypy, z nichž oba jsou radiálně symetrické s ústy obklopenými chapadly, která nesou cnidocyty. Obě formy mají jednu otvor a tělesná dutina, která se používá k trávení a dýchání.
Fosilní cnidarians byly nalezeny ve skalách vytvořených kolem 580 před miliony let. Fosílie cnidariánů, kteří se nestaví mineralizovaný struktury jsou vzácné. Vědci si v současné době myslí, že jsou to obyvatelé ctenofory a bilaterians jsou užší příbuzní vápenaté houby než jsou ostatní houby, a to anthozoans jsou evoluční „tety“ nebo „sestry“ jiných cnidariánů a jsou nejblíže příbuzným bilateriánům.
Cnidarians jsou nejjednodušší zvířata, ve kterých jsou buňky organizovány do tkání.[32] The hvězdice mořská sasanka se používá jako modelový organismus ve výzkumu.[33] Je snadné se o něj starat v laboratoři a protokol byl vyvinut, který může denně přinést velké množství embryí.[34] Existuje značná míra podobnosti v zachování genové sekvence a složitosti mezi mořskou sasankou a obratlovci.[34] Zejména sasanky jsou přítomny také geny zabývající se tvorbou hlavy u obratlovců.[35][36]

Mořské sasanky jsou běžné v tidepools

Jejich chapadla bodat a paralyzovat malé ryby

Pokud se ostrov potopí pod mořem, může růst korálů držet krok s rostoucí vodou a tvořit atol





Lví hříva medúzy, největší známý Medúza[37]

The bioluminiscence mořský ořech má přechodný konečník[38] který se tvoří pouze tehdy, když potřebuje vyprázdnit

Turritopsis dohrnii dosahuje biologická nesmrtelnost přenosem jeho buněk zpět do dětství [39][40]




Mořští červi
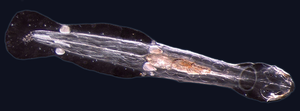
Červi (Stará angličtina pro had) mají obvykle dlouhá válcovitá trubkovitá tělesa a č končetiny. Mořští červi se u některých mořských polychaete červů liší velikostí od mikroskopické po délku přes 1 metr (3,3 ft) (štětinatí červi )[41] a až 58 metrů (190 stop) pro mořského nemerteanského červa (bootlace červ ).[42] Některé mořské červy zabírají malou škálu parazitický výklenky, žijící uvnitř těl jiných zvířat, zatímco jiné žijí volněji v mořském prostředí nebo hrabáním se v podzemí.
Různé skupiny mořských červů jsou příbuzné jen vzdáleně, takže se vyskytují v několika různých phyla tak jako Annelida (segmentovaní červi), Chaetognatha (šipkoví červi), Hemichordata, a Phoronida (červi podkovy). Mnoho z těchto červů má speciální chapadla používaná k výměně kyslíku a oxidu uhličitého a mohou být také použita k reprodukci. Někteří mořští červi jsou trubkové červy, tak jako obří trubkový červ který žije ve vodách blízko pod vodou sopky a vydrží teploty až 90 stupňů Celsia.
Platyhelminthes (ploché červy) tvoří další kmen červů, který zahrnuje třídu Cestoda parazitických tasemnic. Mořská tasemnice Polygonoporus giganticus, nalezený ve střevech spermie velryby, může dorůst na více než 30 m (100 ft).[43][44]
Nematody (škrkavky) tvoří další kmen červů s tubulárními trávicí systémy a otvor na obou koncích.[45][46] Bylo popsáno více než 25 000 druhů hlístic,[47][48] z nichž více než polovina je parazitických. Odhaduje se, že další milion zůstane nepopsaný.[49] Jsou všudypřítomné v mořských, sladkovodních a suchozemských prostředích, kde často převyšují počet ostatních zvířat v počtu jedinců i druhů. Nacházejí se v každé části Země litosféra, z vrcholu hor na dno oceánské příkopy.[50] Podle počtu představují 90% všech zvířat na internetu dno oceánu.[51] Jejich početní dominance, často přesahující milion jedinců na metr čtvereční a tvoří asi 80% všech jednotlivých zvířat na Zemi, jejich rozmanitost životních cyklů a jejich přítomnost na různých trofických úrovních ukazují na důležitou roli v mnoha ekosystémech.[52]

Obří trubkoví červi shluknout se hydrotermální průduchy

Lamellibrachia luymes, a studený prosakovat trubkovitý, žije více než 250 let.

Nematody jsou všudypřítomné pseudocoelomates které mohou parazitovat na mořských rostlinách a zvířatech.

Bloodworms se obvykle nacházejí na dně mělkých mořských vod

Bobbit červi jsou přepadení dravci kteří žijí na mořském dně





Ostnokožci

Ostnokožci (Řek pro ostnatá kůže) je kmen, který obsahuje pouze mořské bezobratlé. Dospělí jsou rozeznatelní podle jejich radiální symetrie (obvykle pětibodové) a zahrnout mořská hvězdice, mořští ježci, pískové dolary, a mořské okurky, stejně jako mořské lilie.[53] Ostnokožci se nacházejí v každé hloubce oceánu, od přílivová zóna do hlubinná zóna. Kmen obsahuje asi 7 000 živých druh,[54] což z něj činí druhé největší seskupení deuterostomy (superkmen), po strunatci (které zahrnují obratlovců, jako ptactvo, Ryby, savci, a plazi ).
Ostnokožci jsou mezi zvířaty jedineční v tom, že mají bilaterální symetrii v larválním stádiu, ale pětinásobnou symetrii (pentamerismus, speciální typ radiální symetrie) jako dospělí.[55]
Ostnokožci jsou důležití biologicky i geologicky. Biologicky existuje několik dalších skupin tak hojných v biotická poušť z hluboké moře, stejně jako mělčí oceány. Většina ostnokožců je schopna regenerovat tkáně, orgány, končetiny a rozmnožování nepohlavně; v některých případech mohou podstoupit úplnou regeneraci z jedné končetiny. Geologicky je hodnota ostnokožců v jejich zkostnatělý kostry, které významně přispívají k mnoha vápenec formace a mohou poskytnout cenné stopy o geologickém prostředí. Byly nejpoužívanějšími druhy v regenerativním výzkumu v 19. a 20. století. Někteří vědci dále tvrdí, že za ozáření ostnokožců bylo odpovědné záření Mesozoic Marine Revolution.

Ostnokožce doslovně znamená „ostnatá kůže“, protože toto vodní meloun mořský ježek ilustruje

Mořské okurky filtrování krmiva na plankton a nerozpuštěné látky

Benthopelagické mořské okurky může zvednout z mořského dna a cestovat až 1 000 m (3 300 ft) po vodním sloupci

The okrová mořská hvězda byl první lichoběžníkového predátora být studován. Omezují mušle což může přemoci přílivové komunity.[56]

Barvitý mořské lilie v mělkých vodách





Kromě těžko klasifikovatelného Arkarua (A Precambrian zvíře s ostnokožcovitou pentamerní radiální symetrií), první definitivní členové kmene se objevili blízko začátku Kambrijský.
Mořští měkkýši

Měkkýši (Latinsky pro měkký) tvoří a kmen s asi 85 000 existující uznáno druh.[57] Podle počtu druhů jsou největší námořní kmen obsahující asi 23% všech jmenovaných mořských organismy.[58] Měkkýši mají rozmanitější formy než jiné kmeny bezobratlých. Jsou velmi různorodé, nejen co do velikosti a velikosti anatomický struktury, ale také v chování a v prostředí. Většina druhů stále žije v oceánech, od mořských pobřeží až po hlubinná zóna, ale některé tvoří a významná část sladkovodní fauny a pozemské ekosystémy.
Kmen měkkýšů je rozdělen na 9 nebo 10 taxonomické třídy, z nichž dva jsou vyhynulý. Mezi tyto třídy patří plži, mlži a hlavonožci, stejně jako další méně známé, ale výrazné třídy. Plži s ochrannými skořápkami se označují jako šneci (mořské šneky ), zatímco plži bez ochranných obalů se označují jako slimáci (mořští slimáci ). Plži jsou zdaleka nejpočetnějšími měkkýši z hlediska klasifikovaných druhů, což představuje 80% z celkového počtu.[25] Mlži zahrnout škeble, ústřice, srdcovky, mušle, lastury a mnoho dalších rodiny. Existuje asi 8 000 druhů mořských mlžů (včetně brakická voda a ústí druhů) a asi 1 200 sladkovodních druhů. Hlavonožce zahrnout chobotnice, oliheň a sépie obecná. Vyskytují se ve všech oceánech a neurologicky jsou nejpokročilejší z bezobratlých.[59] Bylo identifikováno asi 800 živých druhů mořských hlavonožců,[60] a podle odhadů vyhynulo 11 000 lidí taxony byly popsány.[61] Neexistují žádní plně sladkovodní hlavonožci.[62]

Kolosální chobotnice, největší ze všech bezobratlých [63]

The nautilus je žijící fosilie od doby, kdy se vyvinula před 500 miliony let jako jedna z prvních, se nezměnilo hlavonožci.[64][65][66]

Marine plži jsou mořské šneky nebo mořští slimáci. Tento nudibranch je mořský slimák.

Mořský šnek Syrinx aruanus má největší skořápku ze všech živých plžů

Měkkýši mají obvykle oči. Hraničící s okrajem pláště a lastura, a škeble měkkýši, může být více než 100 jednoduché oči.

Slávka obecná, další mlž





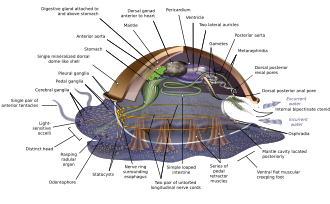
Měkkýši mají tak rozmanité tvary, že mnoho učebnic zakládá svůj popis anatomie měkkýšů na zobecněném nebo hypotetický rodový měkkýš. Tento zobecněný měkkýš je nesegmentovaný a bilaterálně symetrické se spodní stranou sestávající z jediného svalu chodidlo.[67][68]:484–628 Kromě toho má tři další klíčové funkce. Za prvé, má svalnatý plášť zvaný a plášť pokrývající jeho vnitřnosti a obsahující významnou dutinu používanou k dýchání a vylučování. A skořápka vylučovaný pláštěm pokrývá horní povrch.[68] Zadruhé (kromě mlžů) má chraplavý jazyk zvaný a radula používá se ke krmení. Za třetí, má nervový systém včetně komplexního trávicího systému používajícího mikroskopické, svalové vlasy zvané řasy vyzařovat sliz. Zobecněný měkkýš má dva spárované nervové šňůry (tři v mlžech). The mozek U druhů, které ho mají, obklopuje jícen. Většina měkkýšů má oči a všechny mají senzory detekující chemikálie, vibrace a dotek. Nejjednodušší typ měkkýšů rozmnožovací systém spoléhá na vnější hnojení, ale vyskytují se složitější variace. Všechny produkty vejce, ze kterého mohou vzejít trochofor larvy, složitější veliger larvy nebo miniaturní dospělí. Vyobrazení je spíše podobné modernímu monoplakoforany a někteří naznačují, že se může podobat velmi časným měkkýšům.[67]:284–291[67]:298–300[69][70]
Existují dobré důkazy o výskytu mořských plžů, hlavonožci a mlži v Kambrijský doba 541 až 485,4 před miliony let. Evoluční historie obou měkkýšů se však vynořila z předků Lophotrochozoa a jejich diverzifikace na známé bydlení a fosilní Formy jsou stále předmětem intenzivní debaty mezi vědci.
Mořští členovci

Členovci (Řek pro kloubové nohy) mít exoskeleton (externí kostra ), a segmentované tělo a spojené přílohy (spárované přílohy). Tvoří a kmen který zahrnuje hmyz, pavoukovci, myriapods, a korýši. Členovci se vyznačují spojenými končetinami a pokožka vyroben z chitin, často mineralizované uhličitan vápenatý. Členovci tělesný plán skládá se z segmenty, každý s párem přílohy. Tuhá kutikula brzdí růst, takže ji členovci pravidelně nahrazují línání. Jejich všestrannost jim umožnila stát se druhově nejbohatšími členy ze všech ekologické cechy ve většině prostředí.
Velikost mořských členovců je mikroskopická korýš Stygotantulus do Krab japonský. Primární vnitřní dutina členovců je a hemocoel, který pojme jejich vnitřní orgány, a prostřednictvím kterého jejich hemolymfa - analog krev - cirkuluje; oni mají otevřené oběhové systémy. Stejně jako jejich exteriéry jsou vnitřní orgány členovců obecně postaveny z opakovaných segmentů. Jejich nervový systém je „žebříkový“, se spárovanými břišní nervové šňůry prochází všemi segmenty a tvoří spárované ganglia v každém segmentu. Jejich hlavy jsou tvořeny fúzí různého počtu segmentů a jejich mozky jsou tvořeny fúzí ganglií těchto segmentů a obklopují jícen. The respirační a vyměšovací systémy členovců se liší v závislosti na jejich prostředí i na subphylum ke kterému patří.
Jejich vize závisí na různých kombinacích složené oči a pigmentová jáma ocelli: u většiny druhů ocelli dokáže detekovat pouze směr, ze kterého světlo přichází, a složené oči jsou hlavním zdrojem informací, ale hlavní oči pavouci jsou ocelli, kteří mohou vytvářet obrazy a v několika případech se mohou otáčet, aby sledovali kořist. Členovci mají také širokou škálu chemických a mechanických senzorů, většinou založených na modifikacích mnoha štítek (štětiny), které vyčnívají přes jejich kůžičky. Metody rozmnožování a vývoje členovců jsou rozmanité; všechny druhy suchozemských druhů vnitřní hnojení, ale často se jedná o nepřímý přenos spermií pomocí přívěsku nebo země, nikoli přímou injekcí. Mořští druhy kladou vajíčka a používají buď vnitřní, nebo vnější hnojení. Mláďata členovců se liší od miniaturních dospělých až po housenky, které postrádají kloubové končetiny a nakonec podstoupí celkem metamorfóza produkovat formu pro dospělé.

Krab podkovy, a žijící fosilie členovec z doby před 450 miliony let

- Korýši

Mnoho korýšů je velmi malých, jako je tento malý amphipod a tvoří významnou část oceánu zooplankton

The pruhované čistší krevety je korýš běžný v tropech.

The Tasmánský obří krab je dlouhověký a pomalu rostoucí, takže je náchylný k nadměrnému rybolovu.[72]

The Krab japonský má nejdelší rozpětí nohou ze všech členovců.




Evoluční předky členovců sahají do Kambrijský doba. Skupina je obecně považována za monofyletický a mnoho analýz podporuje umístění členovců s cyklonuralisté (nebo jejich složkové subtypy) v superkmenu Ecdysozoa. Celkově však bazální vztahy Metazoa ještě nejsou dobře vyřešeny. Stejně tak se stále aktivně diskutuje o vztazích mezi různými skupinami členovců.
Ostatní phyla
- Tardigrade, Lobopodia, (Onychophora )
- Non-craniate (bez obratlovců) strunatci: Cephalochordate, Tunicata a Haikouella. Tito bezobratlí jsou blízcí příbuzní obratlovců.
- Non-craniate strunatci jsou blízcí příbuzní obratlovců

The kopí, malá průsvitná ryba Cephalochordate, je nejbližší žijící bezobratlý příbuzný obratlovců.[73][74]

Zářivky mořských stříkaček, Rhopalaea crassa. Pláštěnci mohou poskytnout vodítko pro předky obratlovců (a tedy lidí).[75]

Salp řetěz
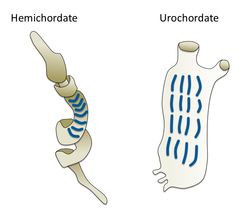
Gill štěrbiny v červu žaludu (vlevo) a pláštěnce (vpravo)




Minerály z mořské vody
Existuje řada mořských bezobratlých, kteří používají minerály přítomné v moři v tak malém množství, že byly nezjistitelné až do příchodu spektroskopie. Vanadium je koncentrován některými pláštěnci pro použití v jejich krevních buňkách na úroveň deset milionůkrát vyšší než okolní mořská voda. Podobně se koncentrují i další pláštěnci niob a tantal.[67]:947 Humři používají měď v jejich respiračním pigmentu hemocyanin, i když podíl tohoto kovu v mořské vodě je nepatrný.[67]:638 Ačkoli jsou tyto prvky v oceánu přítomny v obrovském množství, jejich těžba člověkem není ekonomická.[76]
Viz také
- Skleněné mořské tvory
- mořský život
- mořská biologie
- Mořští obratlovci
- Seznam mořských akvarijních druhů bezobratlých
- Wiwaxia
Poznámky
- ^ Myxozoa byly považovány za výjimku, ale nyní jsou považovány za silně modifikované členy Cnidaria. Jímenez-Guri, Eva; Philippe, Hervé; Okamura, Beth; Holland, Peter W. H. (6. července 2007). "Buddenbrockia Je červ Cnidarian “. Věda. 317 (5834): 116–118. Bibcode:2007Sci ... 317..116J. doi:10.1126 / science.1142024. ISSN 0036-8075. PMID 17615357. S2CID 5170702.CS1 maint: ref = harv (odkaz)
Reference
- ^ Davidson, Michael W. (26. května 2005). "Struktura zvířecích buněk". Molekulární výrazy. Tallahassee, FL: Florida State University. Citováno 2008-09-03.
- ^ Vogel, Gretchen (2018-09-20). „Tato fosilie je podle molekul tuku uchovaných po dobu půl miliardy let jedním z prvních zvířat na světě“. Věda. AAAS. Citováno 21. září 2018.
- ^ Bobrovskiy, Ilya (2018). „Starověké steroidy zakládají fosilii Ediacaran Dickinsonia jako jedno z prvních zvířat“. Věda. 361 (6408): 1246–1249. Bibcode:2018Sci ... 361.1246B. doi:10.1126 / science.aat7228. PMID 30237355.
- ^ Retallack, G.J. (2007). „Růst, rozpad a zhutnění pohřebiště Dickinsonia, ikonické fosílie Ediacaran“ (PDF). Alcheringa: Australasian Journal of paleontologie. 31 (3): 215–240. doi:10.1080/03115510701484705. S2CID 17181699.
- ^ Sperling, Erik; Vinther, Jakob; Pisani, Davide; Peterson, Kevin (2008). „Placozoanská afinita k Dickinsonia a vývoj režimů krmení pozdních prekambrických metazoanů “ (PDF). V Cusack, M; Owen, A; Clark, N (eds.). Program s abstrakty. Výroční zasedání paleontologické asociace. 52. Glasgow, Velká Británie. p. 81.
- ^ Gold, D. A .; Runnegar, B .; Gehling, J. G .; Jacobs, D. K. (2015). „Rekonstrukce ontogeneze předků podporuje bilaterální afinitu k Dickinsonii“. Evoluce a vývoj. 17 (6): 315–397. doi:10.1111 / ede.12168. PMID 26492825.
- ^ Jun-Yuan Chen; Oliveri, Paola; Feng Gao; et al. (1. srpna 2002). „Život prekambrických zvířat: Pravděpodobné vývojové a dospělé formy Cnidarian z jihozápadní Číny“ (PDF). Vývojová biologie. 248 (1): 182–196. doi:10.1006 / dbio.2002.0714. ISSN 0012-1606. PMID 12142030. Archivovány od originál (PDF) 26. května 2013. Citováno 2015-02-04.CS1 maint: ref = harv (odkaz)
- ^ Grazhdankin, Dima (červen 2004). "Distribuční vzorce v Ediacaranské biotě: facie versus biogeografie a evoluce". Paleobiologie. 30 (2): 203–221. doi:10.1666 / 0094-8373 (2004) 030 <0203: PODITE> 2.0.CO; 2. ISSN 0094-8373.CS1 maint: ref = harv (odkaz)
- ^ Seilacher, Adolf (Srpen 1992). „Vendobionta a Psammocorallia: ztracené stavby předkambrijské evoluce“. Časopis geologické společnosti. 149 (4): 607–613. Bibcode:1992JGSoc.149..607S. doi:10.1144 / gsjgs.149.4.0607. ISSN 0016-7649. S2CID 128681462. Citováno 2015-02-04.CS1 maint: ref = harv (odkaz)
- ^ Martin, Mark W .; Grazhdankin, Dmitriy V .; Bowring, Samuel A .; et al. (5. května 2000). „Věk neoproterozoických bilaterálních fosílií těla a stop, Bílé moře, Rusko: důsledky pro vývoj metazoanů“. Věda. 288 (5467): 841–845. Bibcode:2000 sci ... 288..841 mil. doi:10.1126 / science.288.5467.841. ISSN 0036-8075. PMID 10797002. S2CID 1019572.CS1 maint: ref = harv (odkaz)
- ^ Fedonkin, Michail A .; Wagoner, Benjamin M. (28. srpna 1997). „Pozdní prekambrická fosilie Kimberella je měkkýšovitý bilaterální organismus “. Příroda. 388 (6645): 868–871. Bibcode:1997 Natur.388..868F. doi:10.1038/42242. ISSN 0028-0836. S2CID 4395089.CS1 maint: ref = harv (odkaz)
- ^ Mooi, bohatý; David, Bruno (prosinec 1998). „Evoluce v bizarním kmeni: Homologie prvních ostnokožců“ (PDF). Americký zoolog. 38 (6): 965–974. doi:10.1093 / icb / 38.6.965. ISSN 1540-7063. Citováno 2015-02-05.CS1 maint: ref = harv (odkaz)
- ^ McMenamin, Mark A. S. (Září 2003). Spriggina je trilobitoidní ekdyzozoán. Geoscience Horizons Seattle 2003. Abstrakty s programy. 35. Boulder, CO: Geologická společnost Ameriky. p. 105. OCLC 249088612. Citováno 2007-11-24.CS1 maint: ref = harv (odkaz) Příspěvek č. 40-2 představený na výročním zasedání geologické společnosti v Americe v Seattlu v roce 2003 (2. – 5. Listopadu 2003) 2. listopadu 2003 na Washington State Convention Center.
- ^ Jih-Pai Lin; Gon, Samuel M., III; Gehling, James G .; et al. (2006). "A Parvancorina- jako členovec z kambriu v jižní Číně “. Historická biologie: Mezinárodní žurnál paleobiologie. 18 (1): 33–45. doi:10.1080/08912960500508689. ISSN 1029-2381. S2CID 85821717.
- ^ Butterfield, Nicholas J. (prosinec 2006). „Zaháknutí některých„ červů “ze skupiny kmenů: fosilní lophotrochozoany v měděné břidlici.“ BioEssays. 28 (12): 1161–1166. doi:10.1002 / bies.20507. ISSN 0265-9247. PMID 17120226. S2CID 29130876.CS1 maint: ref = harv (odkaz)
- ^ A b Bengtson 2004, s. 67–78
- ^ A b Gould, Stephen Jay (1990) Wonderful Life: The Burgess Shale and the Nature of History W. W. Norton. ISBN 9780393307009.
- ^ Budd, Graham E. (Únor 2003). „Kambrijský fosilní záznam a původ kmene“ (PDF). Integrativní a srovnávací biologie. 43 (1): 157–165. doi:10.1093 / icb / 43.1.157. ISSN 1557-7023. PMID 21680420. Citováno 2015-02-06.CS1 maint: ref = harv (odkaz)
- ^ Budd, Graham E. (březen 1996). „Morfologie Opabinia regalis a rekonstrukce skupiny kmenů členovců“. Lethaia. 29 (1): 1–14. doi:10.1111 / j.1502-3931.1996.tb01831.x. ISSN 0024-1164.CS1 maint: ref = harv (odkaz)
- ^ Marshall, Charles R. (květen 2006). „Vysvětlení kambrijské„ exploze “zvířat“. Výroční přehled o Zemi a planetárních vědách. 34: 355–384. Bibcode:2006AREPS..34..355M. doi:10.1146 / annurev.earth.33.031504.103001. ISSN 1545-4495. S2CID 85623607.CS1 maint: ref = harv (odkaz)
- ^ Valentine, James W. (2004). O původu Phyly. Chicago: University Of Chicago Press. p. 7. ISBN 978-0-226-84548-7.
Klasifikace organismů v hierarchických systémech se používaly v sedmnáctém a osmnáctém století. Organismy byly obvykle seskupeny podle jejich morfologických podobností, jak je vnímají tito raní pracovníci, a tyto skupiny byly poté seskupeny podle jejich podobností atd., Aby vytvořily hierarchii.
CS1 maint: ref = harv (odkaz) - ^ A b Valentine, James W (2004-06-18). O původu Phyly. ISBN 9780226845487.
- ^ Erwin, Douglas; Valentine, James; Jablonski, David (1997). „Nedávné fosilní nálezy a nové poznatky o vývoji zvířat poskytují nové pohledy na hádanku exploze zvířat během raného kambriu“. Americký vědec (Březen duben).
- ^ A b Budd, G. E.; Jensen, S. (květen 2000). „Kritické přehodnocení fosilních záznamů bilaterální phyly“. Biologické recenze. 75 (2): 253–295. doi:10.1111 / j.1469-185X.1999.tb00046.x. PMID 10881389. S2CID 39772232.
- ^ A b Ponder, W.F .; Lindberg, D.R., eds. (2008). Fylogeneze a evoluce měkkýšů. Berkeley: University of California Press. p. 481. ISBN 978-0-520-25092-5.
- ^ Porifera (n.) Online slovník etymologie. Vyvolány 18 August je 2016.
- ^ Vacelet & Duport 2004, str. 179–190.
- ^ „Spongia Linnaeus, 1759“. Světový registr mořských druhů. Citováno 18. července 2012.
- ^ Rowland, S. M. & Stephens, T. (2001). "Archaeocyatha: Historie fylogenetické interpretace". Journal of Paleontology. 75 (6): 1065–1078. doi:10.1666 / 0022-3360 (2001) 075 <1065: AAHOPI> 2.0.CO; 2. JSTOR 1307076.
- ^ Sperling, E. A .; Pisani, D .; Peterson, K. J. (1. ledna 2007). „Poriferanská parafylie a její důsledky pro prekambrickou paleobiologii“ (PDF). Geologická společnost, Londýn, speciální publikace. 286 (1): 355–368. Bibcode:2007GSLSP.286..355S. doi:10.1144 / SP286.25. S2CID 34175521. Archivovány od originál (PDF) 20. prosince 2009. Citováno 22. srpna 2012.
- ^ Zhang, Z.-Q. (2011). „Biodiverzita zvířat: Úvod do vyšší klasifikace a taxonomického bohatství“ (PDF). Zootaxa. 3148: 7–12. doi:10.11646 / zootaxa.3148.1.3.
- ^ "Nematostella vectensis v1.0 ". Portál genomu. University of California. Citováno 2014-01-19.
- ^ "Nematostella". Nematostella.org. Citováno 2014-01-18.
- ^ A b Genikhovich, G. & U. Technau (2009). „Hvězdná mořská sasanka Nematostella vectensis: Anthozoanský modelový organismus pro studium srovnávací genomiky a funkční evoluční vývojové biologie ". Cold Spring Harbor Protocols. 2009 (9): pdb.emo129. doi:10.1101 / pdb.emo129. PMID 20147257.
- ^ „Odkud pochází naše hlava? Bezmozká mořská sasanka vrhá nové světlo na evoluční původ hlavy“. Věda denně. 2013-02-12. Citováno 2014-01-18.
- ^ Sinigaglia, C .; et al. (2013). „Bilaterální gen pro modelování hlavy six3 / 6 řídí vývoj aborální domény u cnidariana“. PLOS Biology. 11 (2): e1001488. doi:10.1371 / journal.pbio.1001488. PMC 3586664. PMID 23483856.
- ^ Karleskint G, Richard Turner R a, James Small J (2012) Úvod do mořské biologie Cengage Learning, vydání 4, strana 445. ISBN 9781133364467.
- ^ Michael Le Page (březen 2019). „Zvíře s konečníkem, které přichází a odchází, může odhalit, jak se naše vyvinulo“. Nový vědec.
- ^ Bavestrello, Giorgio; Christian Sommer; Michele Sarà (1992). "Obousměrná přeměna v Turritopsis nutricula (Hydrozoa)". Scientia Marina. 56 (2–3): 137–140.
- ^ Piraino, Stefano; F. Boero; B. Aeschbach; V. Schmid (1996). „Zvrácení životního cyklu: medúzy transformující se na polypy a buněčná transdiferenciace v Turritopsis nutricula (Cnidaria, Hydrozoa)“. Biologický bulletin. 190 (3): 302–312. doi:10.2307/1543022. JSTOR 1543022. PMID 29227703. S2CID 3956265.
- ^ „Cornwall - Příroda - Červ Superstar“. BBC.
- ^ Mark Carwardine (1995) Guinnessova kniha záznamů o zvířatech. Guinness Publishing. p. 232.
- ^ „Trvalí paraziti“. Časopis Time. 1957-04-08.
- ^ Hargis, William J. (1985). „Parazitologie a patologie mořských organismů světového oceánu“. NOAA Tech. Rep.
- ^ „Klasifikace parazitů zvířat“.
- ^ García, Lynne. „Klasifikace lidských parazitů, vektorů a podobných organismů“ (PDF). UCLA Medical Center Los Angeles, Kalifornie.
- ^ Hodda, M (2011). „Phylum Nematoda Cobb, 1932. In: Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness“. Zootaxa. 3148: 63–95. doi:10.11646 / zootaxa.3148.1.11.
- ^ Zhang, Z (2013). „Biodiverzita zvířat: Aktualizace klasifikace a rozmanitosti v roce 2013. In: Zhang, Z.-Q. (Ed.) Animal Biodiversity: An Outline of High-level Classification and Survey of Taxonomic Richness (Addenda 2013)“. Zootaxa. 3703 (1): 5–11. doi:10.11646 / zootaxa.3703.1.3. S2CID 85252974.
- ^ Lambshead PJD (1993). „Nejnovější vývoj v oblasti výzkumu mořské bentické biologické rozmanitosti“. Oceanis. 19 (6): 5–24.
- ^ Borgonie G, García-Moyano A, Litthauer D, Bert W, Bester A, van Heerden E, Möller C, Erasmus M, Onstott TC (červen 2011). „Nematoda z pozemského hlubokého podzemí Jižní Afriky“. Příroda. 474 (7349): 79–82. Bibcode:2011Natur.474 ... 79B. doi:10.1038 / nature09974. hdl:1854 / LU-1269676. PMID 21637257. S2CID 4399763.
- ^ Danovaro R, Gambi C, Dell'Anno A, Corinaldesi C, Fraschetti S, Vanreusel A, Vincx M, Gooday AJ (leden 2008). „Exponenciální pokles fungování hlubinných ekosystémů spojený se ztrátou bentické biologické rozmanitosti“. Curr. Biol. 18 (1): 1–8. doi:10.1016 / j.cub.2007.11.056. PMID 18164201. S2CID 15272791. Shrnutí ležel – EurekAlert!.
- ^ Platt HM (1994). "úvodní slovo". In Lorenzen S, Lorenzen SA (eds.). Fylogenetická systematika volně žijících hlístic. London: The Ray Society. ISBN 978-0-903874-22-9.
- ^ "Sea Lily". Science Encyclopedia. Citováno 5. zář 2014.
- ^ „Web pro rozmanitost zvířat - Echinodermata“. Muzeum zoologie University of Michigan. Citováno 26. srpna 2012.
- ^ Fox, Richarde. "Asterias forbesi". Anatomie bezobratlých online. Lander University. Citováno 14. června 2014.
- ^ Holsinger, K. (2005). Keystone druhy. Citováno 10. května 2010 z „Archivovaná kopie“. Archivovány od originál dne 30. 6. 2010. Citováno 2010-05-12.CS1 maint: archivovaná kopie jako titul (odkaz)
- ^ Chapman, A.D. (2009). Numbers of Living Species in Australia and the World, 2. vydání. Australská studie biologických zdrojů, Canberra. Citováno 12. ledna 2010. ISBN 978-0-642-56860-1 (tištěné); ISBN 978-0-642-56861-8 (online).
- ^ Hancock, Rebecca (2008). „Uznáváme výzkum měkkýšů“. Australské muzeum. Archivovány od originál dne 2009-05-30. Citováno 2009-03-09.
- ^ Barnes, R.S.K, Calow P, Olive, P.J.W., Golding DW, Spicer JI (2001). Bezobratlí, syntéza (3. vyd.). UK: Blackwell Science.
- ^ „Vítejte na CephBase“. CephBase. Citováno 29. ledna 2016.
- ^ Wilbur, Karl M .; Clarke, M.R .; Trueman, E.R., eds. (1985), Měkkýši, 12. Paleontologie a neontologie hlavonožců, New York: Academic Press, ISBN 0-12-728702-7
- ^ „Existují nějaké sladkovodní hlavonožci?“. 16. ledna 2013.
- ^ Black, Richard (26. dubna 2008). „Kolosální chobotnice z mrazáku“. BBC novinky.
- ^ Ewen Callaway (2. června 2008). „Simple-Minded Nautilus Shows Flash of Memory“. Nový vědec. Citováno 7. března 2012.
- ^ Kathryn Phillips (15. června 2008). „Živé fosilní vzpomínky“. Journal of Experimental Biology. 211 (12): iii. doi:10.1242 / jeb.020370. S2CID 84279320.
- ^ Robyn Crook & Jennifer Basil (2008). „Dvoufázová paměťová křivka v komorovém nautilu, Nautilus pompilius L. (Cephalopoda: Nautiloidea) ". The Journal of Experimental Biology. 211 (12): 1992–1998. doi:10.1242 / jeb.018531. PMID 18515730. S2CID 6305526.
- ^ A b C d E F Ruppert, Edward E .; Fox, Richard, S .; Barnes, Robert D. (2004). Zoologie bezobratlých, 7. vydání. Cengage Learning. ISBN 978-81-315-0104-7.
- ^ A b Hayward, PJ (1996). Příručka pro mořské fauny severozápadní Evropy. Oxford University Press. ISBN 978-0-19-854055-7.
- ^ Giribet, G .; Okusu, A, A .; Lindgren, A.R., A.R .; Huff, S.W., S. W .; Schrödl, M, M .; Nishiguchi, M. K., M. K. (květen 2006). „Důkazy o kladu složeném z měkkýšů se sériově se opakujícími strukturami: monoplakoforany jsou příbuzné chitonům“. Sborník Národní akademie věd Spojených států amerických. 103 (20): 7723–7728. Bibcode:2006PNAS..103,7723G. doi:10.1073 / pnas.0602578103. PMC 1472512. PMID 16675549.
- ^ Healy, J.M. (2001). „Měkkýš“. V Anderson, D.T. (ed.). Zoologie bezobratlých (2. vyd.). Oxford University Press. str. 120–171. ISBN 978-0-19-551368-4.
- ^ Práce, ano, my. „Zjistit více o First Life - David Attenborough's First Life“. Archivovány od originál dne 26.01.2011.
- ^ D. R. Currie; T. M. Ward (2009). Krab jižní australský (Pseudocarcinus gigas) Rybářství (PDF). Jižní australský institut pro výzkum a vývoj. Zpráva o hodnocení rybolovu pro PIRSA. Archivovány od originál (PDF) dne 28. března 2012. Citováno 9. prosince 2013.
- ^ Gewin, V (2005). „Funkční genomika zesiluje biologickou zápletku“. PLOS Biology. 3 (6): e219. doi:10.1371 / journal.pbio.0030219. PMC 1149496. PMID 15941356.
- ^ Lanceletový (amphioxus) genom a původ obratlovců Ars Technica, 19. června 2008.
- ^ Lemaire, P (2011). "Evoluční křižovatka ve vývojové biologii: pláštěnci". Rozvoj. 138 (11): 2143–2152. doi:10,1242 / dev.048975. PMID 21558365. S2CID 40452112.
- ^ Carson, Rachel (1997). Moře kolem nás. Oxfordské brožované výtisky. 190–191. ISBN 978-0195069976.

