Respirační komplex I. - Respiratory complex I

Respirační komplex I., ES 7.1.1.2 (také známý jako NADH: ubichinon oxidoreduktáza, NADH dehydrogenáza typu I a mitochondriální komplex I.) je první velký proteinový komplex z dýchací řetězce mnoha organismů od bakterií po člověka. Katalyzuje přenos elektrony z NADH na koenzym Q10 (CoQ10) a translokuje protony přes vnitřní mitochondriální membrána u eukaryot nebo plazmatická membrána bakterií.
| Respirační komplex I. | |
|---|---|
| Identifikátory | |
| Symbol | Respirační komplex I. |
| OPM nadčeleď | 246 |
| OPM protein | 6g72 |
| Membranome | 255 |
| NADH: ubichinon reduktáza (H.+- přemístění). | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||||
| EC číslo | 1.6.5.3 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Tento enzym je nezbytný pro normální fungování buněk a mutace v jeho podjednotkách vedou k široké škále zděděných neuromuskulárních a metabolických poruch. Vady tohoto enzymu jsou zodpovědné za vývoj několika patologických procesů, jako jsou ischemie / reperfúze poškození (mrtvice a srdeční infarkt ), Parkinsonova choroba a další.
Funkce


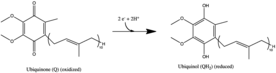
Komplex I je první enzym z mitochondriální elektronový transportní řetězec. V řetězci transportu elektronů existují tři enzymy transdukující energii - NADH: ubichinon oxidoreduktáza (komplex I), Koenzym Q - cytochrom c reduktáza (komplex III) a cytochrom c oxidáza (komplex IV).[1] Komplex I je největší a nejkomplikovanější enzym elektronového transportního řetězce.[2]
Reakce katalyzovaná komplexem I je:
- NADH + H+ + CoQ + 4H+v→ NAD+ + CoQH2 + 4H+ven
V tomto procesu komplex translokuje čtyři protony přes vnitřní membránu na molekulu oxidovaného NADH,[3][4][5] pomáhá budovat elektrochemický potenciál rozdíl používaný k výrobě ATP. Escherichia coli komplex I (NADH dehydrogenáza) je schopen translokace protonů stejným směrem k ustavenému Δψ, což ukazuje, že za testovaných podmínek je vazebným iontem H+.[6] Na+ byl pozorován transport v opačném směru, a přestože Na+ to nebylo nutné pro aktivity katalytického nebo protonového transportu, jeho přítomnost to zvýšila. H+ byl přemístěn Paracoccus denitrificans komplexní já, ale v tomto případě, H+ transport nebyl ovlivněn Na+a Na+ transport nebyl pozorován. Možná E-coli komplex I má dvě místa vazby energie (jedno Na+ nezávislý a druhý Na+závislé), jak bylo pozorováno u Rhodothermus marinus komplex I, zatímco vazební mechanismus P. denitrificans enzym je zcela Na+ nezávislý. Je také možné, že jiný transportér katalyzuje příjem Na+. Přenos energie komplexu I čerpáním protonů nemusí být výlučný pro R. marinus enzym. Pak+/ H+ antiportová aktivita se nejeví jako obecná vlastnost komplexu I.[6] Existence Na+- translokační aktivita komplexu I je stále otázkou.
Reakci lze zvrátit - označuje se jako NAD podporovaná aerobním sukcinátem+ redukce ubichinolem - v přítomnosti vysokého membránového potenciálu, ale přesný katalytický mechanismus zůstává neznámý. Hnací silou této reakce je potenciál přes membránu, který lze udržovat buď hydrolýzou ATP, nebo komplexy III a IV během oxidace sukcinátu.[7]
Komplex I může hrát roli při spouštění apoptóza.[8] Ve skutečnosti se ukázalo, že existuje korelace mezi mitochondriálními aktivitami a programovaná buněčná smrt (PCD) během vývoje somatického embrya.[9]
Komplex I není homologní Na+- rodina NADH dehydrogenázy (NDH) (TC # 3.D.1 ), člen Na+ transport Mrp superrodiny.
V důsledku oxidace dvou molekul NADH na NAD + mohou být komplexem IV v respiračním řetězci produkovány tři molekuly ATP.
Mechanismus
Celkový mechanismus
Všechny redoxní reakce probíhají v hydrofilní doméně komplexu I. NADH se zpočátku váže na komplex I a přenáší dva elektrony na flavin mononukleotid (FMN) protetická skupina enzymu, která vytváří FMNH2. Akceptor elektronů - isoalloxazinový kruh - FMN je totožný s akceptorem elektronů FAD. Elektrony jsou poté přeneseny přes FMN prostřednictvím řady shluků železa a síry (Fe-S),[10] a nakonec koenzym Q10 (ubichinon). Tento tok elektronů mění redoxní stav proteinu a vyvolává konformační změny proteinu, které mění pK. hodnoty ionizovatelného postranního řetězce a způsobí odčerpání čtyř vodíkových iontů z mitochondriální matrice.[11] Ubichinon (CoQ) přijímá dva elektrony, které mají být redukovány na ubichinol (CoQH2).[1]
Mechanismus přenosu elektronů
Navrhovaná cesta pro transport elektronů před redukcí ubichinonu je následující: NADH - FMN - N3 - N1b - N4 - N5 - N6a - N6b - N2 - Q, kde Nx je konvence značení pro klastry s obsahem železa a síry.[10] Vysoký redukční potenciál klastru N2 a relativní blízkost ostatních klastrů v řetězci umožňují efektivní přenos elektronů na velkou vzdálenost v proteinu (s přenosovými rychlostmi z NADH do klastru železo-síra N2 asi 100 μs).[12][13]
Rovnovážná dynamika komplexu I je primárně řízena redoxním cyklem chinonu. V podmínkách vysoké protonové hybné síly (a tedy i ubichinolem koncentrované směsi) běží enzym v opačném směru. Ubichinol se oxiduje na ubichinon a výsledné uvolněné protony snižují hybnou sílu protonu.[14]
Mechanismus translokace protonů
Spojení protonové translokace a transportu elektronů v komplexu I je v současné době navrhováno jako nepřímé (konformační změny s dlouhým dosahem) na rozdíl od přímé (redoxní meziprodukty ve vodíkových pumpách jako v heme skupiny komplexů III a IV ).[10] Architektura hydrofobní oblasti komplexu I ukazuje několik protonových transportérů, které jsou mechanicky propojeny. Tři centrální složky, o nichž se předpokládá, že přispívají k této konformační změně na velké vzdálenosti, jsou klastr N2 se sírou a sírou spojený s pH, redukce chinonu a transmembránové šroubovicové podjednotky ramene membrány. Transdukce konformačních změn k pohonu transmembránových transportérů spojených „ojnicí“ během redukce ubichinonu může představovat dva nebo tři ze čtyř protonů čerpaných na oxidovaný NADH. Zbývající proton musí být čerpán přímou vazbou na místo vázající ubichinon. Navrhuje se, aby přímé a nepřímé vazebné mechanismy odpovídaly za čerpání čtyř protonů.[15]
Blízkost klastru N2 k blízkému cysteinovému zbytku vede ke konformační změně po redukci blízkých šroubovic, což vede k malým, ale důležitým změnám v celkové proteinové konformaci.[16] Dále elektronová paramagnetická rezonance studie přenosu elektronů prokázaly, že většina energie, která se uvolní během následné redukce CoQ, je na konečné ubichinol krok formace od semichinon, poskytující důkazy o „jediném mrtvici“ H+ translokační mechanismus (tj. všechny čtyři protony se pohybují přes membránu současně).[14][17] Alternativní teorie navrhují „dvoutaktní mechanismus“, kde každý redukční krok (semichinon a ubichinol ) vede k úderu dvou protonů vstupujících do mezimembránového prostoru.[18][19]
Výsledný ubichinol lokalizovaný v membránové doméně interaguje se záporně nabitými zbytky v rameni membrány a stabilizuje konformační změny.[10] An antiporter mechanismus (Na+/ H+ swap) byl navržen s využitím důkazů o konzervovaných zbytcích Asp v rameni membrány.[20] Přítomnost zbytků Lys, Glu a His umožňuje protonové hradlování (protonace následovaná deprotonací přes membránu) poháněnou pKA zbytků.[10]
Složení a struktura
NADH: ubichinon oxidoreduktáza je největší z dýchacích komplexů. v savci, enzym obsahuje 44 samostatných ve vodě rozpustných proteinů periferní membrány, které jsou ukotveny k integrálním složkám membrány. Zvláštní funkční význam mají flavin protetická skupina (FMN) a osm klastry železo-síra (FeS). Ze 44 podjednotek je sedm kódováno mitochondriální genom.[21][22][23]
Struktura má tvar „L“ s dlouhou membránovou doménou (s přibližně 60 trans-membránovými šroubovicemi) a hydrofilní (nebo periferní) doménou, která zahrnuje všechna známá redoxní centra a vazebné místo NADH.[24] Všech třináct z E-coli proteiny, které obsahují NADH dehydrogenázu I, jsou kódovány v nuo operon a jsou homologní s podjednotkami I mitochondriálního komplexu. Každá antiporterová podjednotka NuoL / M / N obsahuje 14 konzervovaných šroubovic transmembrány (TM). Dva z nich jsou diskontinuální, ale podjednotka NuoL obsahuje 110 A dlouhou amfipatickou α-šroubovici, pokrývající celou délku domény. Podjednotka NuoL souvisí s Na+/ H+ antiportery z TC # 2.A.63.1.1 (PhaA a PhaD).
Tři ze konzervovaných podjednotek vázaných na membránu v NADH dehydrogenáze jsou navzájem příbuzné a antiportery Mrp sodný-proton. Strukturální analýza dvou prokaryotických komplexů Zjistil jsem, že každá ze tří podjednotek obsahuje čtrnáct transmembránových šroubovic, které se překrývají ve strukturním uspořádání: translokace tří protonů může být koordinována boční spirálou, která je spojuje.[25]
Komplex I obsahuje ubichinonovou vazebnou kapsu na rozhraní podjednotek 49 kDa a PSST. V blízkosti klastru železa a síry N2, navrhovaného okamžitého donoru elektronů pro ubichinon, představuje vysoce konzervovaný tyrosin kritický prvek místa redukce chinonu. Možná chinonová směnná cesta vede z klastru N2 k N-terminálnímu beta-listu 49-kDa podjednotky.[26] Všech 45 podjednotek hovězího NDHI bylo sekvenováno.[27][28] Každý komplex obsahuje nekovalentně vázaný FMN, koenzym Q a několik center železa a síry. Bakteriální NDH mají 8-9 center obsahujících železo a síru.
Použitá nedávná studie elektronová paramagnetická rezonance (EPR) spektra a dvojitá elektron-elektronová rezonance (DEER) k určení cesty přenosu elektronů komplexy železo-síra, které se nacházejí v hydrofilní doméně. Sedm z těchto klastrů tvoří řetězec od flavinu k místům vázajícím chinon; osmý klastr je umístěn na druhé straně flavinu a jeho funkce není známa. Výsledky EPR a DEER naznačují alternativní nebo „horskou dráhu“ potenciálního energetického profilu pro přenos elektronů mezi aktivními místy a podél klastrů železa a síry, což může optimalizovat rychlost pohybu elektronů a umožnit efektivní přeměnu energie v komplexu I.[29]
| # | Člověk /Hovězí podjednotka | Lidský protein | Popis bílkovin (UniProt ) | Pfam rodina s lidským proteinem | |
|---|---|---|---|---|---|
| Základní podjednotkyA | |||||
| 1 | NDUFS7 / PSST / NUKM | NDUS7_HUMAN | NADH dehydrogenáza [ubichinon] železo-sirný protein 7, mitochondriální ES 1.6.5.3 ES 1.6.99.3 | Pfam PF01058 | |
| 2 | NDUFS8 / TYKY / NUIM | NDUS8_HUMAN | NADH dehydrogenáza [ubichinon] protein železo-síra 8, mitochondriální ES 1.6.5.3 ES 1.6.99.3 | Pfam PF12838 | |
| 3 | NDUFV2 / 24kD / NUHMC | NDUV2_HUMAN | NADH dehydrogenáza [ubichinon] flavoprotein 2, mitochondriální ES 1.6.5.3 ES 1.6.99.3 | Pfam PF01257 | |
| 4 | NDUFS3 / 30kD / NUGM | NDUS3_HUMAN | NADH dehydrogenáza [ubichinon] železo-sirný protein 3, mitochondriální ES 1.6.5.3 ES 1.6.99.3 | Pfam PF00329 | |
| 5 | NDUFS2 / 49kD / NUCM | NDUS2_HUMAN | NADH dehydrogenáza [ubichinon] železo-sirný protein 2, mitochondriální ES 1.6.5.3 ES 1.6.99.3 | Pfam PF00346 | |
| 6 | NDUFV1 / 51kD / NUBM | NDUV1_HUMAN | NADH dehydrogenáza [ubichinon] flavoprotein 1, mitochondriální ES 1.6.5.3 ES 1.6.99.3 | Pfam PF01512 | |
| 7 | NDUFS1 / 75kD / NUAM | NDUS1_HUMAN | NADH-ubichinon oxidoreduktáza 75 kDa podjednotka, mitochondriální ES 1.6.5.3 ES 1.6.99.3 | Pfam PF00384 | |
| 8 | ND1 / NU1M | NU1M_HUMAN | Řetěz NADH-ubichinon oxidoreduktázy 1 ES 1.6.5.3 | Pfam PF00146 | |
| 9 | ND2 / NU2M | NU2M_HUMAN | Řetězec NADH-ubichinon oxidoreduktázy 2 ES 1.6.5.3 | Pfam PF00361, Pfam PF06444 | |
| 10 | ND3 / NU3M | NU3M_HUMAN | Řetězec NADH-ubichinon oxidoreduktázy 3 ES 1.6.5.3 | Pfam PF00507 | |
| 11 | ND4 / NU4M | NU4M_HUMAN | Řetězec NADH-ubichinon oxidoreduktázy 4 ES 1.6.5.3 | Pfam PF01059, Pfam PF00361 | |
| 12 | ND4L / NULM | NU4LM_HUMAN | NADH-ubichinon oxidoreduktázový řetězec 4L ES 1.6.5.3 | Pfam PF00420 | |
| 13 | ND5 / NU5M | NU5M_HUMAN | NADH-ubichinon oxidoreduktázový řetězec 5 ES 1.6.5.3 | Pfam PF00361, Pfam PF06455, Pfam PF00662 | |
| 14 | ND6 / NU6M | NU6M_HUMAN | Řetězec NADH-ubichinon oxidoreduktázy 6 ES 1.6.5.3 | Pfam PF00499 | |
| Základní doplňkové podjednotkyb | |||||
| 15 | NDUFS6 / 13A | NDUS6_HUMAN | NADH dehydrogenáza [ubichinon] železo-sirný protein 6, mitochondriální | Pfam PF10276 | |
| 16 | NDUFA12 / B17.2 | NDUAC_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplexní podjednotka 12 | Pfam PF05071 | |
| 17 | NDUFS4 / AQDQ | NDUS4_HUMAN | NADH dehydrogenáza [ubichinon] železo-sirný protein 4, mitochondriální | Pfam PF04800 | |
| 18 | NDUFA9 / 39kDa | NDUA9_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplex podjednotka 9, mitochondriální | Pfam PF01370 | |
| 19 | NDUFAB1 / ACPM | ACPM_HUMAN | Acylový nosný protein, mitochondriální | Pfam PF00550 | |
| 20 | NDUFA2 / B8 | NDUA2_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplexní podjednotka 2 | Pfam PF05047 | |
| 21 | NDUFA1 / MFWE | NDUA1_HUMAN | NADH dehydrogenáza [ubichinon] 1 podkomplex alfa podjednotky 1 | Pfam PF15879 | |
| 22 | NDUFB3 / B12 | NDUB3_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplexní podjednotka 3 | Pfam PF08122 | |
| 23 | NDUFA5 / AB13 | NDUA5_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplexní podjednotka 5 | Pfam PF04716 | |
| 24 | NDUFA6 / B14 | NDUA6_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplexní podjednotka 6 | Pfam PF05347 | |
| 25 | NDUFA11 / B14.7 | NDUAB_HUMAN | NADH dehydrogenáza [ubichinon] 1 subkomplexní podjednotka alfa 11 | Pfam PF02466 | |
| 26 | NDUFB11 / ESSS | NDUBB_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplex podjednotka 11, mitochondriální | Pfam PF10183 | |
| 27 | NDUFS5 / PFFD | NDUS5_HUMAN | NADH dehydrogenáza [ubichinon] protein železo-síra 5 | Pfam PF10200 | |
| 28 | NDUFB4 / B15 | NDUB4_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplexní podjednotka 4 | Pfam PF07225 | |
| 29 | NDUFA13 / A13 | NDUAD_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa subkomplexní podjednotka 13 | Pfam PF06212 | |
| 30 | NDUFB7 / B18 | NDUB7_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplexní podjednotka 7 | Pfam PF05676 | |
| 31 | NDUFA8 / PGIV | NDUA8_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplexní podjednotka 8 | Pfam PF06747 | |
| 32 | NDUFB9 / B22 | NDUB9_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplexní podjednotka 9 | Pfam PF05347 | |
| 33 | NDUFB10 / PDSW | NDUBA_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplexní podjednotka 10 | Pfam PF10249 | |
| 34 | NDUFB8 / ASHI | NDUB8_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta podkomplex podjednotka 8, mitochondriální | Pfam PF05821 | |
| 35 | NDUFC2 / B14.5B | NDUC2_HUMAN | NADH dehydrogenáza [ubichinon] 1 podjednotka C2 | Pfam PF06374 | |
| 36 | NDUFB2 / AGGG | NDUB2_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplex podjednotka 2, mitochondriální | Pfam PF14813 | |
| 37 | NDUFA7 / B14.5A | NDUA7_HUMAN | NADH dehydrogenáza [ubichinon] 1 subkomplexní podjednotka alfa 7 | Pfam PF07347 | |
| 38 | NDUFA3 / B9 | NDUA3_HUMAN | NADH dehydrogenáza [ubichinon] 1 subkomplexní podjednotka alfa 3 | Pfam PF14987 | |
| 39 | NDUFA4 / MLRQC | NDUA4_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplexní podjednotka 4 | Pfam PF06522 | |
| 40 | NDUFB5 / SGDH | NDUB5_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplex podjednotka 5, mitochondriální | Pfam PF09781 | |
| 41 | NDUFB1 / MNLL | NDUB1_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta podkomplex podjednotka 1 | Pfam PF08040 | |
| 42 | NDUFC1 / KFYI | NDUC1_HUMAN | NADH dehydrogenáza [ubichinon] 1 podjednotka C1, mitochondriální | Pfam PF15088 | |
| 43 | NDUFA10 / 42kD | NDUAA_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa subkomplexní podjednotka 10, mitochondriální | Pfam PF01712 | |
| 44 | NDUFA4L2 | NUA4L_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa podkomplex podjednotka 4 jako 2 | Pfam PF15880 | |
| 45 | NDUFV3 | NDUV3_HUMAN | NADH dehydrogenáza [ubichinon] flavoprotein 3, 10 kDa | - | |
| 46 | NDUFB6 | NDUB6_HUMAN | NADH dehydrogenáza [ubichinon] 1 beta subkomplexní podjednotka 6 | Pfam PF09782 | |
| Proteiny montážního faktoru[31] | |||||
| 47 | NDUFAF1C | CIA30_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa subkomplex, montážní faktor 1 | Pfam PF08547 | |
| 48 | NDUFAF2 | MIMIT_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa subkomplex, montážní faktor 2 | Pfam PF05071 | |
| 49 | NDUFAF3 | NDUF3_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa subkomplexní montážní faktor 3 | Pfam PF05071 | |
| 50 | NDUFAF4 | NDUF4_HUMAN | NADH dehydrogenáza [ubichinon] 1 alfa subkomplex, montážní faktor 4 | Pfam PF06784 | |
Poznámky:
- A Nalezeno u všech druhů kromě hub
- b Může nebo nemusí být přítomen u žádného druhu
- C Nalezeno v houbových druhů, jako jsou Schizosaccharomyces pombe
Inhibitory
Bullatacin (an acetogenin nalezen v Asimina triloba ovoce) je nejúčinnějším známým inhibitorem NADH dehydrogenázy (ubichinon) (IC50 = 1,2 nM, silnější než rotenon).[34] Nejznámější inhibitor komplexu I je rotenon (běžně používaný jako organický pesticid). Rotenon a rotenoidy jsou isoflavonoidy vyskytující se v několika rodech tropických rostlin, jako je Antonia (Loganiaceae ), Derris a Lonchocarpus (Faboideae, Fabaceae ). Již v 17. století se objevují zprávy o domorodých obyvatelích Francouzské Guyany, kteří používají ryby obsahující rotenon k rybolovu - kvůli jejichtyotoxickému účinku.[35] Rotenon se váže na ubichinon vazebné místo komplexu I i piericidin A, další silný inhibitor s blízkým strukturním homologem k ubichinonu.
Acetogeniny z Annonaceae jsou ještě účinnějšími inhibitory komplexu I. Zesíťují podjednotku ND2, což naznačuje, že ND2 je nezbytný pro vazbu chinonu.[36] Rolliniastatin-2, acetogenin, je prvním zjištěným komplexním inhibitorem I, který nesdílí stejné vazebné místo jako rotenon.[37]
Přes více než 50 let studia komplexu I nebyly nalezeny žádné inhibitory blokující tok elektronů uvnitř enzymu. Hydrofobní inhibitory jako rotenon nebo piericidin s největší pravděpodobností narušují přenos elektronů mezi koncovým FeS klastrem N2 a ubichinonem. Bylo prokázáno, že dlouhodobá systémová inhibice komplexu I rotenonem může vyvolat selektivní degeneraci dopaminergních neuronů.[38]
Komplex I je také blokován adenosindifosfát ribóza - reverzibilní kompetitivní inhibitor oxidace NADH - vazbou na enzym v místě vazby nukleotidů.[39] Jak hydrofilní NADH, tak hydrofobní ubichinonové analogy působí na začátku a na konci vnitřní transportní dráhy elektronů.
Antidiabetikum Metformin Bylo prokázáno, že indukuje mírnou a přechodnou inhibici mitochondriálního komplexu dýchacích řetězců I a zdá se, že tato inhibice hraje klíčovou roli v jeho mechanismu účinku.[40]
Byla zahrnuta inhibice komplexu I. hepatotoxicita například spojené s různými léky flutamid a nefazodon.[41]
Aktivní / neaktivní přechod
Katalytické vlastnosti eukaryotického komplexu I nejsou jednoduché. V jakékoli dané přípravě enzymu existují dvě katalyticky a strukturně odlišné formy: jedna je plně kompetentní, takzvaná „aktivní“ forma A a druhá je katalyticky tichá, spící, „neaktivní“ forma D. Po vystavení nečinného enzymu zvýšeným, ale fyziologickým teplotám (> 30 ° C) v nepřítomnosti substrátu se enzym přemění na D-formu. Tato forma je katalyticky nekompetentní, ale může být aktivována pomalou reakcí (k ~ 4 min−1) oxidace NADH s následnou redukcí ubichinonu. Po jednom nebo několika obratech se enzym aktivuje a může katalyzovat fyziologickou reakci NADH: ubichinon mnohem vyšší rychlostí (k ~ 104 min−1). V přítomnosti dvojmocných kationtů (Mg2+, Ca2+), nebo při alkalickém pH trvá aktivace mnohem déle.
Výška aktivační energie (270 kJ / mol) deaktivačního procesu naznačuje výskyt velkých konformačních změn v organizaci komplexu I. Jediným dosud pozorovaným konformačním rozdílem mezi těmito dvěma formami je však počet cystein zbytky vystavené na povrchu enzymu. Zpracování D-formy komplexu I sulfhydrylovými činidly N-ethylmaleimid nebo DTNB ireverzibilně blokuje kritický cysteinový zbytek (zbytky), čímž se ruší schopnost enzymu reagovat na aktivaci, čímž se ireverzibilně deaktivuje. A-forma komplexu I je necitlivá na sulfhydrylová činidla.
Bylo zjištěno, že tyto konformační změny mohou mít velmi důležitý fyziologický význam. Deaktivní, ale ne aktivní forma komplexu I byla náchylná k inhibici nitrosothioly a peroxynitrit.[42] Je pravděpodobné, že k přechodu z aktivní na neaktivní formu komplexu I dochází během patologických stavů, kdy je obrat enzymu omezen při fyziologických teplotách, například během hypoxie, nebo když tkáň oxid dusnatý: zvyšuje se poměr kyslíku (tj. metabolická hypoxie).[43]
Výroba superoxidu
Nedávná šetření naznačují, že komplex I je silným zdrojem reaktivní formy kyslíku.[44] Komplex mohu vyrobit superoxid (stejně jako peroxid vodíku ), alespoň dvěma různými cestami. Během dopředného přenosu elektronů se vytváří pouze velmi malé množství superoxidu (pravděpodobně méně než 0,1% celkového toku elektronů).[44][45]
Během přenosu reverzních elektronů může být komplex I nejdůležitějším místem produkce superoxidu v mitochondriích, přičemž asi 3-4% elektronů je odkloněno k tvorbě superoxidu.[46] Reverzní přenos elektronů, proces, při kterém elektrony ze snížené zásoby ubichinolu (dodávané společností sukcinát dehydrogenáza, glycerol-3-fosfátdehydrogenáza, flavoprotein přenášející elektrony nebo dihydroorotát dehydrogenáza v savčích mitochondriích) procházejí komplexem I ke snížení NAD+ k NADH, poháněn potenciálem vnitřní mitochondriální membrány elektrický potenciál. Ačkoli není přesně známo, za jakých patologických podmínek by došlo k přenosu reverzních elektronů in vivo, experimenty in vitro naznačují, že tento proces může být velmi účinným zdrojem superoxidu, když sukcinát koncentrace jsou vysoké a oxaloacetát nebo malát koncentrace jsou nízké.[47] K tomu může dojít během ischemie tkáně, kdy je blokován přísun kyslíku.[48]
Superoxid je reaktivní forma kyslíku, která přispívá k buněčnému oxidačnímu stresu a souvisí s neuromuskulárními chorobami a stárnutím.[49] NADH dehdyrogenáza produkuje superoxid přenosem jednoho elektronu z FMNH2 na kyslík (O.2). Zbytek radikálu flavinu je nestabilní a přenáší zbývající elektron do center železa a síry. Je to poměr NADH k NAD+ která určuje rychlost tvorby superoxidu.[50]
Patologie
Mutace v podjednotkách komplexu, které mohu způsobit mitochondriální nemoci, počítaje v to Leighův syndrom. Bodové mutace v různých komplexních I podjednotkách odvozených z mitochondriální DNA (mtDNA ) může také vést k Leberova dědičná optická neuropatie. Existují určité důkazy, že složité defekty I mohou hrát roli v etiologii Parkinsonova choroba Možná kvůli reaktivním kyslíkovým formám (komplex, který můžu mít rád komplex III, propouští elektrony na kyslík a tvoří vysoce toxické superoxid ).
Ačkoli přesná etiologie Parkinsonovy choroby není jasná, je pravděpodobné, že může hrát velkou roli mitochondriální dysfunkce, spolu s inhibicí proteazomu a toxiny z prostředí. Ve skutečnosti bylo prokázáno, že inhibice komplexu I způsobuje produkci peroxidů a snížení aktivity proteazomu, což může vést k Parkinsonově nemoci.[51] Navíc Esteves a kol. (2010) zjistili, že buněčné linie s Parkinsonovou chorobou vykazují zvýšený únik protonů v komplexu I, což způsobuje sníženou maximální respirační kapacitu.[52]
Nedávné studie zkoumaly další role aktivity komplexu I v mozku. Andreazza a kol. (2010) zjistili, že úroveň aktivity komplexu I byla významně snížena u pacientů s bipolární poruchou, ale ne u pacientů s depresí nebo schizofrenií. Zjistili, že pacienti s bipolární poruchou vykazovali zvýšenou oxidaci a nitraci bílkovin v prefrontální kůře. Tyto výsledky naznačují, že budoucí studie by se měly zaměřit na komplex I pro potenciální terapeutické studie pro bipolární poruchu.[53] Podobně Moran a kol. (2010) zjistili, že pacienti se závažným nedostatkem komplexu I vykazovali sníženou míru spotřeby kyslíku a pomalejší růst. Zjistili však, že mutace v různých genech v komplexu I vedou k různým fenotypům, čímž vysvětlují variace patofyziologických projevů deficitu komplexu I.[54]
Expozice pesticidům může také inhibovat komplex I a způsobit příznaky nemoci. Ukázalo se například, že chronická expozice nízkým hladinám dichlorvosu, organofosfátu používaného jako pesticid, způsobuje jaterní dysfunkci. K tomu dochází, protože dichlorvos mění úrovně aktivity komplexu I a II, což vede ke snížení aktivity přenosu mitochondriálních elektronů a ke snížení syntézy ATP.[55]
Geny
Následuje seznam lidských genů, které kódují složky komplexu I:
- NADH dehydrogenáza (ubichinon) 1 alfa subkomplex
- NDUFA1 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 1, 7,5 kDa
- NDUFA2 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 2, 8 kDa
- NDUFA3 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 3, 9 kDa
- NDUFA4 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 4, 9 kDa
- NDUFA4L - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 4-podobný
- NDUFA4L2 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 4 podobný 2
- NDUFA5 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 5, 13 kDa
- NDUFA6 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 6, 14 kDa
- NDUFA7 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 7, 14,5 kDa
- 8. NDUFA - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 8, 19 kDa
- NDUFA9 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 9, 39 kDa
- NDUFA10 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 10, 42 kDa
- NDUFA11 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 11, 14,7 kDa
- 12. NDUFA - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 12
- NDUFA13 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, 13
- NDUFAB1 - NADH dehydrogenáza (ubichinon) 1, alfa / beta subkomplex, 1, 8 kDa
- NDUFAF1 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, montážní faktor 1
- NDUFAF2 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, montážní faktor 2
- NDUFAF3 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, montážní faktor 3
- NDUFAF4 - NADH dehydrogenáza (ubichinon) 1 alfa subkomplex, montážní faktor 4
- NADH dehydrogenáza (ubichinon) 1 beta subkomplex
- NDUFB1 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 1, 7 kDa
- NDUFB2 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 2, 8 kDa
- NDUFB3 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 3, 12 kDa
- NDUFB4 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 4, 15 kDa
- NDUFB5 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 5, 16 kDa
- NDUFB6 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 6, 17 kDa
- NDUFB7 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 7, 18 kDa
- NDUFB8 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 8, 19 kDa
- NDUFB9 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 9, 22 kDa
- NDUFB10 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 10, 22 kDa
- NDUFB11 - NADH dehydrogenáza (ubichinon) 1 beta subkomplex, 11, 17,3 kDa
- NADH dehydrogenáza (ubichinon) 1, subkomplex neznámý
- NADH dehydrogenáza (ubichinon) Fe-S protein
- NDUFS1 - NADH dehydrogenáza (ubichinon) Fe-S protein 1, 75 kDa (NADH-koenzym Q reduktáza)
- NDUFS2 - NADH dehydrogenáza (ubichinon) Fe-S protein 2, 49 kDa (NADH-koenzym Q reduktáza)
- NDUFS3 - NADH dehydrogenáza (ubichinon) Fe-S protein 3, 30 kDa (NADH-koenzym Q reduktáza)
- NDUFS4 - NADH dehydrogenáza (ubichinon) Fe-S protein 4, 18 kDa (NADH-koenzym Q reduktáza)
- NDUFS5 - NADH dehydrogenáza (ubichinon) protein Fe-S 5, 15 kDa (NADH-koenzym Q reduktáza)
- NDUFS6 - NADH dehydrogenáza (ubichinon) Fe-S protein 6, 13 kDa (NADH-koenzym Q reduktáza)
- NDUFS7 - NADH dehydrogenáza (ubichinon) Fe-S protein 7, 20 kDa (NADH-koenzym Q reduktáza)
- 8. NDUFS - NADH dehydrogenáza (ubichinon) protein Fe-S 8, 23 kDa (NADH-koenzym Q reduktáza)
- Flavoprotein NADH dehydrogenázy (ubichinon) 1
- mitochondriálně kódovaná podjednotka NADH dehydrogenázy
- MT-ND1 - mitochondriálně kódovaná podjednotka NADH dehydrogenázy 1
- MT-ND2 - mitochondriálně kódovaná podjednotka NADH dehydrogenázy 2
- MT-ND3 - mitochondriálně kódovaná podjednotka NADH dehydrogenázy 3
- MT-ND4 - mitochondriálně kódovaná podjednotka NADH dehydrogenázy 4
- MT-ND4L - mitochondriálně kódovaná podjednotka NADH dehydrogenázy 4L
- MT-ND5 - mitochondriálně kódovaná podjednotka NADH dehydrogenázy 5
- MT-ND6 - mitochondriálně kódovaná podjednotka NADH dehydrogenázy 6
Reference
- ^ A b Berg, J; Tymoczko, J; L Stryer (2006). Biochemie (6. vydání). New York: WH Freeman & Company. 509–513.
- ^ Brandt U (2006). "Energeticky konvertující NADH: chinon oxidoreduktáza (komplex I)". Roční přehled biochemie. 75: 69–92. doi:10,1146 / annurev.biochem.75.103004.142539. PMID 16756485.
- ^ Wikström, M. (1984-04-24). "Dva protony jsou čerpány z mitochondriální matrice na elektron přenesený mezi NADH a ubichinonem." FEBS Dopisy. 169 (2): 300–304. doi:10.1016/0014-5793(84)80338-5. ISSN 0014-5793. PMID 6325245.
- ^ Galkin A, Dröse S, Brandt U (prosinec 2006). „Stechiometrii čerpání protonů čištěného mitochondriálního komplexu jsem rekonstituoval na proteoliposomy“. Biochim. Biophys. Acta. 1757 (12): 1575–81. doi:10.1016 / j.bbabio.2006.10.001. ISSN 0006-3002. PMID 17094937.
- ^ Galkin, A. S .; Grivennikova, V. G .; Vinogradov, A. D. (1999-05-21). „-> H + / 2e- stechiometrie v reakcích NADH-chinon reduktázy katalyzovaných submochondriálními částicemi hovězího srdce“. FEBS Dopisy. 451 (2): 157–161. doi:10.1016 / s0014-5793 (99) 00575-x. ISSN 0014-5793. PMID 10371157.
- ^ A b Batista AP, Pereira MM (březen 2011). „Vliv sodíku na transdukci energie komplexy I z Escherichia coli a Paracoccus denitrificans“. Biochimica et Biophysica Acta (BBA) - bioenergetika. 1807 (3): 286–92. doi:10.1016 / j.bbabio.2010.12.008. PMID 21172303.
- ^ Grivennikova VG, Kotlyar AB, Karliner JS, Cecchini G, Vinogradov AD (září 2007). „Redoxně závislá změna afinity nukleotidů k aktivnímu místu savčího komplexu I“. Biochemie. 46 (38): 10971–8. doi:10.1021 / bi7009822. PMC 2258335. PMID 17760425.
- ^ Chomova M, Racay P (březen 2010). „Mitochondriální komplex I v síti známých a neznámých faktů“. Obecná fyziologie a biofyzika. 29 (1): 3–11. doi:10,4149 / gpb_2010_01_3. PMID 20371875.
- ^ Petrussa E, Bertolini A, Casolo V, Krajnáková J, Macrì F, Vianello A (prosinec 2009). „Mitochondriální bioenergetika spojená s projevem programované buněčné smrti během somatické embryogeneze Abies alba“. Planta. 231 (1): 93–107. doi:10.1007 / s00425-009-1028-x. PMID 19834734.
- ^ A b C d E Sazanov LA (Červen 2015). „Obří molekulární protonová pumpa: struktura a mechanismus dýchacího komplexu I“. Recenze přírody. Molekulární buněčná biologie. 16 (6): 375–88. doi:10.1038 / nrm3997. PMID 25991374.
- ^ Donald J. Voet; Judith G. Voet; Charlotte W. Pratt (2008). „Kapitola 18, Mitochondriální syntéza ATP“. Principles of Biochemistry, 3. vydání. Wiley. str. 608. ISBN 978-0-470-23396-2.
- ^ Ohnishi, T (1998). „Klastry železo-síra / semichinony v komplexu I“. Biochim. Biophys. Acta. 1364 (2): 186–206. doi:10.1016 / s0005-2728 (98) 00027-9. PMID 9593887.
- ^ Bridges HR, Bill E, Hirst J (leden 2012). „Mössbauerova spektroskopie na respiračním komplexu I: soubor klastrů železo-síra v enzymu se sníženým obsahem NADH je částečně oxidován“. Biochemie. 51 (1): 149–58. doi:10.1021 / bi201644x. PMC 3254188. PMID 22122402.
- ^ A b Efremov RG, Sazanov LA (Říjen 2012). „Mechanismus vazby dýchacího komplexu I - strukturální a evoluční perspektiva“. Biochimica et Biophysica Acta (BBA) - bioenergetika. 1817 (10): 1785–95. doi:10.1016 / j.bbabio.2012.02.015. PMID 22386882.
- ^ Treberg JR, Quinlan CL, Brand MD (srpen 2011). „Důkazy pro dvě místa produkce superoxidů mitochondriální NADH-ubichinon oxidoreduktázou (komplex I)“. The Journal of Biological Chemistry. 286 (31): 27103–10. doi:10,1074 / jbc.M111.252502. PMC 3149303. PMID 21659507.
- ^ Berrisford JM, Sazanov LA (Říjen 2009). „Strukturální základ mechanismu dýchacího komplexu I“. The Journal of Biological Chemistry. 284 (43): 29773–83. doi:10.1074 / jbc.m109.032144. PMC 2785608. PMID 19635800.
- ^ Baranova EA, Morgan DJ, Sazanov LA (Srpen 2007). „Analýza jednotlivých částic potvrzuje distální umístění podjednotek NuoL a NuoM v komplexu I Escherichia coli.“ Journal of Structural Biology. 159 (2): 238–42. doi:10.1016 / j.jsb.2007.01.009. PMID 17360196.
- ^ Brandt U (říjen 2011). „Dvoustavový mechanismus stabilizace a změny pro proton-čerpající komplex I“. Biochimica et Biophysica Acta (BBA) - bioenergetika. 1807 (10): 1364–9. doi:10.1016 / j.bbabio.2011.04.006. PMID 21565159.
- ^ Zickermann V, Wirth C, Nasiri H, Siegmund K, Schwalbe H, Hunte C, Brandt U (leden 2015). "Strukturní biologie. Mechanický pohled z krystalové struktury mitochondriálního komplexu I" (PDF). Věda. 347 (6217): 44–9. doi:10.1126 / science.1259859. PMID 25554780.
- ^ Hunte C, Screpanti E, Venturi M, Rimon A, Padan E, Michel H (červen 2005). "Struktura antiporteru Na + / H + a poznatky o mechanismu působení a regulaci pomocí pH". Příroda. 435 (7046): 1197–202. doi:10.1038 / nature03692. PMID 15988517.
- ^ Voet, Judith G .; Voet, Donald (2004). Biochemie (3. vyd.). New York: J. Wiley & Sons. str.813 –826. ISBN 0-471-19350-X.
- ^ Carroll J, Fearnley IM, Skehel JM, Shannon RJ, Hirst J, Walker JE (říjen 2006). „Hovězí komplex I je komplex 45 různých podjednotek“. The Journal of Biological Chemistry. 281 (43): 32724–7. doi:10,1074 / jbc.M607135200. PMID 16950771.
- ^ Balsa E, Marco R, Perales-Clemente E, Szklarczyk R, Calvo E, Landázuri MO, Enríquez JA (září 2012). „NDUFA4 je podjednotka komplexu IV savčího elektronového transportního řetězce“. Buněčný metabolismus. 16 (3): 378–86. doi:10.1016 / j.cmet.2012.07.015. PMID 22902835.
- ^ Sazanov LA, Hinchliffe P (březen 2006). "Struktura hydrofilní domény dýchacího komplexu I z Thermus thermophilus". Věda. 311 (5766): 1430–6. doi:10.1126 / science.1123809. PMID 16469879.
- ^ Efremov RG, Baradaran R, Sazanov LA (Květen 2010). "Architektura dýchacího komplexu I". Příroda. 465 (7297): 441–5. doi:10.1038 / nature09066. PMID 20505720.
- ^ Tocilescu MA, Zickermann V, Zwicker K, Brandt U (prosinec 2010). "Vazba chinonu a redukce dýchacím komplexem I". Biochimica et Biophysica Acta (BBA) - bioenergetika. 1797 (12): 1883–90. doi:10.1016 / j.bbabio.2010.05.009. PMID 20493164.
- ^ Cardol P, Vanrobaeys F, Devreese B, Van Beeumen J, Matagne RF, Remacle C (říjen 2004). "Vyšší rostlinné podjednotkové složení mitochondriálního komplexu I z Chlamydomonas reinhardtii: 31 konzervovaných složek mezi eukaryoty". Biochimica et Biophysica Acta (BBA) - bioenergetika. 1658 (3): 212–24. doi:10.1016 / j.bbabio.2004.06.001. PMID 15450959.
- ^ Gabaldón T, Rainey D, Huynen MA (květen 2005). „Sledování vývoje velkého proteinového komplexu v eukaryotech, NADH: ubichinon oxidoreduktáza (komplex I)“. Journal of Molecular Biology. 348 (4): 857–70. doi:10.1016 / j.jmb.2005.02.067. PMID 15843018.
- ^ Roessler MM, King MS, Robinson AJ, Armstrong FA, Harmer J, Hirst J (únor 2010). „Přímé přiřazení EPR spekter strukturně definovaným shlukům železa a síry v komplexu I dvojitou elektron-elektronovou rezonancí“. Sborník Národní akademie věd Spojených států amerických. 107 (5): 1930–5. doi:10.1073 / pnas.0908050107. PMC 2808219. PMID 20133838.
- ^ Cardol P (listopad 2011). „Mitochondriální NADH: ubichinon oxidoreduktáza (komplex I) u eukaryot: vysoce konzervované složení podjednotky zvýrazněné těžbou proteinových databází“. Biochimica et Biophysica Acta (BBA) - bioenergetika. 1807 (11): 1390–7. doi:10.1016 / j.bbabio.2011.06.015. PMID 21749854.
- ^ Ogilvie I, Kennaway NG, Shoubridge EA (říjen 2005). „Molekulární chaperon pro sestavu mitochondriálního komplexu I je mutován v progresivní encefalopatii.“. The Journal of Clinical Investigation. 115 (10): 2784–92. doi:10,1172 / JCI26020. PMC 1236688. PMID 16200211.
- ^ Dunning CJ, McKenzie M, Sugiana C, Lazarou M, Silke J, Connelly A, Fletcher JM, Kirby DM, Thorburn DR, Ryan MT (červenec 2007). „Lidská CIA30 je zapojena do časného sestavení mitochondriálního komplexu I a mutací v jeho genu způsobujícím onemocnění“. Časopis EMBO. 26 (13): 3227–37. doi:10.1038 / sj.emboj.7601748. PMC 1914096. PMID 17557076.
- ^ Saada A, Vogel RO, Hoefs SJ, van den Brand MA, Wessels HJ, Willems PH, Venselaar H, Shaag A, Barghuti F, Reish O, Shohat M, Huynen MA, Smeitink JA, van den Heuvel LP, Nijtmans LG (červen 2009). „Mutace v NDUFAF3 (C3ORF60), kódující NDUFAF4 (C6ORF66) -interagující komplex I montážního proteinu, způsobují fatální neonatální mitochondriální onemocnění“. American Journal of Human Genetics. 84 (6): 718–27. doi:10.1016 / j.ajhg.2009.04.020. PMC 2694978. PMID 19463981.
- ^ Miyoshi H, Ohshima M, Shimada H, Akagi T, Iwamura H, McLaughlin JL (July 1998). "Essential structural factors of annonaceous acetogenins as potent inhibitors of mitochondrial complex I". Biochimica et Biophysica Acta (BBA) - bioenergetika. 1365 (3): 443–52. doi:10.1016/s0005-2728(98)00097-8. PMID 9711297.
- ^ Moretti C, Grenand P (September 1982). "[The "nivrées", or ichthyotoxic plants of French Guyana]". Journal of Ethnopharmacology (francouzsky). 6 (2): 139–60. doi:10.1016/0378-8741(82)90002-2. PMID 7132401.
- ^ Nakamaru-Ogiso E, Han H, Matsuno-Yagi A, Keinan E, Sinha SC, Yagi T, Ohnishi T (March 2010). "The ND2 subunit is labeled by a photoaffinity analogue of asimicin, a potent complex I inhibitor". FEBS Dopisy. 584 (5): 883–8. doi:10.1016/j.febslet.2010.01.004. PMC 2836797. PMID 20074573.
- ^ Degli Esposti M, Ghelli A, Ratta M, Cortes D, Estornell E (July 1994). "Natural substances (acetogenins) from the family Annonaceae are powerful inhibitors of mitochondrial NADH dehydrogenase (Complex I)". The Biochemical Journal. 301 ( Pt 1): 161–7. doi:10.1042/bj3010161. PMC 1137156. PMID 8037664.
- ^ Watabe M, Nakaki T (October 2008). "Mitochondrial complex I inhibitor rotenone inhibits and redistributes vesicular monoamine transporter 2 via nitration in human dopaminergic SH-SY5Y cells". Molekulární farmakologie. 74 (4): 933–40. doi:10.1124/mol.108.048546. PMID 18599602.
- ^ Zharova TV, Vinogradov AD (July 1997). "A competitive inhibition of the mitochondrial NADH-ubiquinone oxidoreductase (complex I) by ADP-ribose". Biochimica et Biophysica Acta (BBA) - bioenergetika. 1320 (3): 256–64. doi:10.1016/S0005-2728(97)00029-7. PMID 9230920.
- ^ Viollet B, Guigas B, Sanz Garcia N, Leclerc J, Foretz M, Andreelli F (March 2012). "Cellular and molecular mechanisms of metformin: an overview". Klinická věda. 122 (6): 253–70. doi:10.1042/CS20110386. PMC 3398862. PMID 22117616.
- ^ Nadanaciva, Sashi; Will, Yvonne (2011). "New Insights in Drug-Induced Mitochondrial Toxicity". Současný farmaceutický design. 17 (20): 2100–2112. doi:10.2174/138161211796904795. ISSN 1381-6128. PMID 21718246.
- ^ Galkin A, Moncada S (December 2007). "S-nitrosation of mitochondrial complex I depends on its structural conformation". The Journal of Biological Chemistry. 282 (52): 37448–53. doi:10.1074/jbc.M707543200. PMID 17956863.
- ^ Moncada S, Erusalimsky JD (March 2002). "Does nitric oxide modulate mitochondrial energy generation and apoptosis?". Recenze přírody. Molekulární buněčná biologie. 3 (3): 214–20. doi:10.1038/nrm762. PMID 11994742.
- ^ A b Murphy MP (January 2009). "How mitochondria produce reactive oxygen species". The Biochemical Journal. 417 (1): 1–13. doi:10.1042/BJ20081386. PMC 2605959. PMID 19061483.
- ^ Hansford RG, Hogue BA, Mildaziene V (February 1997). "Dependence of H2O2 formation by rat heart mitochondria on substrate availability and donor age". Journal of Bioenergetics and Biomembranes. 29 (1): 89–95. doi:10.1023/A:1022420007908. PMID 9067806.
- ^ Stepanova, Anna; Kahl, Anja; Konrad, Csaba; Ten, Vadim; Starkov, Anatoly S.; Galkin, Alexander (December 2017). "Reverse electron transfer results in a loss of flavin from mitochondrial complex I: Potential mechanism for brain ischemia reperfusion injury". Journal of Cerebral Blood Flow and Metabolism. 37 (12): 3649–3658. doi:10.1177/0271678X17730242. ISSN 1559-7016. PMC 5718331. PMID 28914132.
- ^ Muller FL, Liu Y, Abdul-Ghani MA, Lustgarten MS, Bhattacharya A, Jang YC, Van Remmen H (leden 2008). „Vysoká míra produkce superoxidu v mitochondriích kosterního svalstva dýchajících na komplexních substrátech vázaných na I i na komplex II“. The Biochemical Journal. 409 (2): 491–9. doi:10.1042 / BJ20071162. PMID 17916065.
- ^ Sahni, Prateek V.; Zhang, Jimmy; Sosunov, Sergey; Galkin, Alexander; Niatsetskaya, Zoya; Starkov, Anatoly; Brookes, Paul S.; Ten, Vadim S. (February 2018). "Krebs cycle metabolites and preferential succinate oxidation following neonatal hypoxic-ischemic brain injury in mice". Pediatrický výzkum. 83 (2): 491–497. doi:10.1038/pr.2017.277. ISSN 1530-0447. PMC 5866163. PMID 29211056.
- ^ Esterházy D, King MS, Yakovlev G, Hirst J (March 2008). "Production of reactive oxygen species by complex I (NADH:ubiquinone oxidoreductase) from Escherichia coli and comparison to the enzyme from mitochondria". Biochemie. 47 (12): 3964–71. doi:10.1021/bi702243b. PMID 18307315.
- ^ Kussmaul L, Hirst J (May 2006). "The mechanism of superoxide production by NADH:ubiquinone oxidoreductase (complex I) from bovine heart mitochondria". Sborník Národní akademie věd Spojených států amerických. 103 (20): 7607–12. doi:10.1073/pnas.0510977103. PMC 1472492. PMID 16682634.
- ^ Chou AP, Li S, Fitzmaurice AG, Bronstein JM (August 2010). "Mechanisms of rotenone-induced proteasome inhibition". Neurotoxikologie. 31 (4): 367–72. doi:10.1016/j.neuro.2010.04.006. PMC 2885979. PMID 20417232.
- ^ Esteves AR, Lu J, Rodova M, Onyango I, Lezi E, Dubinsky R, Lyons KE, Pahwa R, Burns JM, Cardoso SM, Swerdlow RH (May 2010). "Mitochondrial respiration and respiration-associated proteins in cell lines created through Parkinson's subject mitochondrial transfer". Journal of Neurochemistry. 113 (3): 674–82. doi:10.1111/j.1471-4159.2010.06631.x. PMID 20132468.
- ^ Andreazza AC, Shao L, Wang JF, Young LT (April 2010). "Mitochondrial complex I activity and oxidative damage to mitochondrial proteins in the prefrontal cortex of patients with bipolar disorder". Archiv obecné psychiatrie. 67 (4): 360–8. doi:10.1001/archgenpsychiatry.2010.22. PMID 20368511.
- ^ Morán M, Rivera H, Sánchez-Aragó M, Blázquez A, Merinero B, Ugalde C, Arenas J, Cuezva JM, Martín MA (May 2010). "Mitochondrial bioenergetics and dynamics interplay in complex I-deficient fibroblasts". Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 1802 (5): 443–53. doi:10.1016/j.bbadis.2010.02.001. PMID 20153825.
- ^ Binukumar BK, Bal A, Kandimalla R, Sunkaria A, Gill KD (April 2010). "Mitochondrial energy metabolism impairment and liver dysfunction following chronic exposure to dichlorvos". Toxikologie. 270 (2–3): 77–84. doi:10.1016/j.tox.2010.01.017. PMID 20132858.
externí odkazy
- IST Austria: Sazanov Group MRC MBU Sazanov group
- Interactive Molecular model of NADH dehydrogenase (Vyžaduje Zvonkohra MDL )
- Complex I homepage
- Electron+Transport+Complex+I v americké národní lékařské knihovně Lékařské předměty (Pletivo)
Do tato úprava, tento článek používá obsah z "3.D.1 The H+ or Na+-translocating NADH Dehydrogenase (NDH) Family", který je licencován způsobem, který umožňuje opětovné použití v rámci Creative Commons Attribution-ShareAlike 3.0 Unported License, ale ne pod GFDL. Je třeba dodržovat všechny příslušné podmínky.