Alfa šroubovice - Alpha helix

The alfa šroubovice (α-šroubovice) je běžný motiv v sekundární struktura z bílkoviny a je pravá ruka -spirála konformace, ve které každá páteř N-H skupina Vodíkové vazby na páteř C = O skupina aminokyselina nachází tři nebo čtyři zbytky dříve podél proteinové sekvence.
Alfa šroubovici se také říká klasika Pauling – Corey – Branson α-šroubovice. Název 3.613-spirála se také používá pro tento typ šroubovice, což označuje průměrný počet zbytků na spirálovitý závit, přičemž 13 atomů je zapojeno do kruhu tvořeného vodíkovou vazbou.
Z typů lokální struktury v proteinech je α-šroubovice nejextrémnější a nejpředvídatelnější ze sekvence, stejně jako nejrozšířenější.

Objev


Na počátku 30. let William Astbury ukázal, že v EU došlo k drastickým změnám rentgen difrakce vláken vlhké vlny nebo vlasových vláken při výrazném protažení. Data naznačují, že nenapnutá vlákna měla navinutou molekulární strukturu s charakteristickým opakováním ≈5,1 ångströms (0.51 nanometry ).
Astbury původně navrhoval strukturu zauzleného řetězce vláken. Později se přidal k dalším výzkumníkům (zejména americkému chemikovi) Maurice Huggins ) tím, že navrhuje, aby:
- nenapnuté molekuly proteinu vytvořily spirálu (kterou nazval α-forma)
- protažení způsobilo, že se šroubovice odvinula a vytvořila prodloužený stav (který nazýval β-forma).
Ačkoli byly Astburyho modely těchto forem nesprávné ve svých detailech, byly v podstatě správné a odpovídaly moderním prvkům sekundární struktura, α-šroubovice a β-vlákno (Astburyho nomenklatura byla zachována), které byly vyvinuty Linus Pauling, Robert Corey a Herman Branson v roce 1951 (viz níže); ten papír ukazoval jak pravotočivé, tak levoruké šroubovice, i když v roce 1960 krystalová struktura myoglobinu[1] ukázal, že forma pro praváky je běžná. Hans Neurath byl první, kdo ukázal, že Astburyho modely nemohou být do detailu správné, protože šlo o střety atomů.[2] Neurathův dokument a data Astburyho se inspirovaly H. S. Taylor,[3] Maurice Huggins[4] a Bragg a spolupracovníky[5] navrhovat modely keratin které se poněkud podobají moderní α-šroubovici.
Dva klíčové vývojové trendy v modelování moderní α-šroubovice byly: správná geometrie vazby díky stanovení krystalové struktury z aminokyseliny a peptidy a Paulingova předpověď rovinný peptidové vazby; a jeho vzdání se předpokladu integrálního počtu reziduí na otáčku šroubovice. Rozhodující okamžik nastal brzy na jaře roku 1948, kdy Pauling nachladl a šel spát. Když se nudil, nakreslil na proužek papíru polypeptidový řetězec zhruba správných rozměrů, složil ho do šroubovice a dával pozor, aby udržoval rovinné peptidové vazby. Po několika pokusech vytvořil model s fyzikálně přijatelnými vodíkovými vazbami. Pauling poté spolupracoval s Coreym a Bransonem na potvrzení svého modelu před zveřejněním.[6] V roce 1954 získal Pauling svou první Nobelovu cenu „za výzkum podstaty chemické vazby a její aplikace při objasňování struktury komplexních látek“[7] (jako jsou proteiny), prominentně zahrnující strukturu α-šroubovice.
Struktura
Geometrie a vodíkové vazby
Aminokyseliny v α-šroubovici jsou uspořádány pravou rukou spirálovitý Struktura, kde každý aminokyselinový zbytek odpovídá otočení o 100 ° ve šroubovici (tj. šroubovice má 3,6 zbytku na otáčku) a translaci 1,5 Á (0,15 nm) podél spirálové osy. Dunitz[8] popisuje, jak Paulingův první článek na toto téma ve skutečnosti ukazuje levou spirálu, enantiomer skutečné struktury. Krátké kousky spirály pro leváky se někdy vyskytují s velkým obsahem achirálů glycin aminokyseliny, ale jsou nepříznivé pro ostatní normální, biologické L-aminokyseliny. Stoupání alfa-šroubovice (svislá vzdálenost mezi po sobě následujícími závity šroubovice) je 5,4 Å (0,54 nm), což je součin 1,5 a 3,6. Nejdůležitější je, že N-H skupina aminokyseliny tvoří a vodíková vazba s C = O skupina aminokyseliny čtyři zbytky dříve; toto se opakovalo i + 4 → i vodíková vazba je nejvýznamnější charakteristikou α-šroubovice. Oficiální mezinárodní nomenklatura[9][10] specifikuje dva způsoby definování α-helixů, pravidlo 6.2, pokud jde o opakování φ, ψ torzní úhly (viz níže) a pravidlo 6.3, pokud jde o kombinovaný vzor hřiště a vodíkových vazeb. Α-helixy lze identifikovat v proteinové struktuře pomocí několika výpočetních metod, z nichž jedna je DSSP (DefinovatSekundární struktura bílkovin).[11]
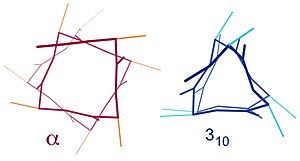
Podobné struktury zahrnují 310 spirála (i + 3 → i vodíkové vazby) a π-šroubovice (i + 5 → i vodíkové vazby). Α-helix lze popsat jako 3.613 spirála, protože i + 4 mezery přidávají do H-vázané smyčky další tři atomy ve srovnání s přísnější 310 helix, a v průměru je 3,6 aminokyselin zapojených do jednoho kruhu α-helixu. Dolní indexy označují počet atomů (včetně vodíku) v uzavřené smyčce tvořené vodíkovou vazbou.[12]

Zbytky v α-šroubovicích obvykle přijímají páteř (φ, ψ) vzepětí kolem (-60 °, -45 °), jak je znázorněno na obrázku vpravo. Obecněji řečeno, přijímají vzepětí úhly takové, že ψ úhel díry jednoho zbytku a φ vzepětí úhlu další součet zbytků zhruba -105 °. Důsledkem toho je, že α-šroubovicové vzepětí, obvykle spadá na diagonální pruh na Ramachandranův diagram (sklon -1), v rozmezí (-90 °, -15 °) až (-35 °, -70 °). Pro srovnání, součet dihedrálních úhlů pro 310 spirála je zhruba −75 °, zatímco u π-spirála je zhruba −130 °. Obecný vzorec pro úhel otočení Ω na zbytek jakékoli polypeptidové šroubovice s trans isomerů je dáno rovnicí[14][15]
- 3 cos Ω = 1 - 4 cos2 φ + ψ/2
Α-šroubovice je pevně zabalená; uvnitř šroubovice není téměř žádné volné místo. Aminokyselinové postranní řetězce jsou na vnější straně šroubovice a směřují zhruba „dolů“ (tj. Směrem k N-konci), jako větve vždyzeleného stromu (vánoční strom účinek). Tato směrovost se někdy používá v předběžných mapách elektronové hustoty s nízkým rozlišením k určení směru páteře proteinu.[16]
Stabilita
Helixy pozorované v proteinech se mohou pohybovat v rozmezí od čtyř do více než čtyřiceti zbytků, ale typická šroubovice obsahuje asi deset aminokyselin (asi tři otáčky). Obecně krátký polypeptidy nevykazují v roztoku mnoho α-helikální struktury, protože entropický náklady spojené se skládáním polypeptidového řetězce nejsou kompenzovány dostatečným množstvím stabilizačních interakcí. Obecně páteř Vodíkové vazby α-šroubovice jsou považovány za mírně slabší než ty, které se nacházejí v β-listy, a jsou snadno napadeny molekulami okolní vody. Ve více hydrofobních prostředích, jako je plazmatická membrána, nebo v přítomnosti pomocných rozpouštědel, jako je trifluorethanol (TFE) nebo izolované z rozpouštědla v plynné fázi,[17] oligopeptidy snadno přijmou stabilní a-helikální strukturu. Kromě toho mohou být do peptidů začleněny příčné vazby, aby se konformačně stabilizovaly šroubovité záhyby. Zesíťování stabilizuje spirálovitý stav entropickou destabilizací rozvinutého stavu a odstraněním entalpicky stabilizovaných „návnadových“ záhybů, které konkurují plně spirálovitému stavu.[18] Ukázalo se, že α-helixy jsou stabilnější, odolnější vůči mutacím a designovatelnější než β-řetězce v přírodních proteinech,[19] a také v umělých proteinech.[20]

Experimentální stanovení
Protože α-helix je definován svými vodíkovými vazbami a konformací páteře, nejpodrobnější experimentální důkaz pro α-helikální strukturu pochází z atomového rozlišení Rentgenová krystalografie například příklad zobrazený vpravo. Je zřejmé, že všechny páteřní karbonylové kyslíky směřují dolů (směrem k C-konci), ale mírně se rozprostírají a H-vazby jsou přibližně rovnoběžné s osou šroubovice. Proteinové struktury z NMR spektroskopie také dobře ukázat helixy s charakteristickými pozorováními jaderný Overhauserův efekt (NOE) vazby mezi atomy na sousedních šroubovicových závitech. V některých případech mohou být jednotlivé vodíkové vazby pozorovány přímo jako malá skalární vazba v NMR.
Existuje několik metod s nižším rozlišením pro přiřazení obecné spirálové struktury. The NMR chemické směny (zejména C.α, C.β a C ') a zbytkové dipolární spojky jsou často charakteristické pro helixy. Daleko UV (170–250 nm) kruhový dichroismus spektrum šroubovic je také idiosynkratické a vykazuje výrazné dvojnásobné minimum kolem 208 a 222 nm. Infračervený spektroskopie se používá jen zřídka, protože a-helikální spektrum se podobá spektru a náhodná cívka (i když je lze rozeznat např. výměna vodík-deuterium ). Nakonec kryo elektronová mikroskopie je nyní schopen rozeznat jednotlivé α-šroubovice v proteinu, ačkoli jejich přiřazení zbytkům je stále aktivní oblastí výzkumu.
Dlouhé homopolymery aminokyselin často tvoří šroubovice, pokud jsou rozpustné. Tak dlouhé izolované helixy lze detekovat i jinými metodami, jako je například dielektrická relaxace, tok dvojlom a měření difúzní konstanta. Přísněji řečeno, tyto metody detekují pouze charakteristiku prolovat (dlouhý doutníkový) hydrodynamický tvar spirály nebo její velké dipólový moment.
Sklony k aminokyselinám
Různé aminokyselinové sekvence mají různé sklony k vytváření a-helikální struktury. Methionin, alanin, leucin, glutamát, a lysin nenabité ("MALEK" v aminokyselina Jednopísmenové kódy), přičemž všechny mají obzvláště vysoké sklony k tvorbě šroubovice prolin a glycin mají špatné sklony k tvorbě šroubovice.[21] Prolin buď zlomí nebo zalomí šroubovici, protože nemůže darovat amida vodíková vazba (bez amidového vodíku), a také proto, že jeho postranní řetězec stericky interferuje s páteří předchozího tahu - uvnitř šroubovice to nutí ohyb asi 30 ° v ose šroubovice.[12] Prolin je však často považován za za prvé zbytku šroubovice, předpokládá se to kvůli její strukturní tuhosti. V opačném extrému glycin má také tendenci narušovat helixy, protože jeho vysoká konformační flexibilita činí entropicky nákladné přijmout relativně omezenou α-helikální strukturu.
Tabulka standardních sklonů aminokyselin k alfa-šroubovici
Odhadované rozdíly v energie zdarma, Δ (ΔG), odhadovaný v kcal / mol na zbytek v α-helikální konfiguraci, relativně k alaninu libovolně nastavený na nulu. Vyšší čísla (více pozitivních volných energií) jsou méně oblíbená. Jsou možné významné odchylky od těchto průměrných čísel v závislosti na totožnosti sousedních zbytků.
Rozdíly ve volné energii na zbytek[22] Aminokyselina 3-
dopis1-
dopisSpirálový trest kcal / mol kJ / mol Alanin Ala A 0.00 0.00 Arginin Arg R 0.21 0.88 Asparagin Asn N 0.65 2.72 Kyselina asparagová Asp D 0.69 2.89 Cystein Cys C 0.68 2.85 Kyselina glutamová Glu E 0.40 1.67 Glutamin Gln Q 0.39 1.63 Glycin Gly G 1.00 4.18 Histidin Jeho H 0.61 2.55 Isoleucin Ile Já 0.41 1.72 Leucin Leu L 0.21 0.88 Lysin Lys K. 0.26 1.09 Methionin Se setkal M 0.24 1.00 Fenylalanin Phe F 0.54 2.26 Prolin Pro P 3.16 13.22 Serine Ser S 0.50 2.09 Threonin Thr T 0.66 2.76 Tryptofan Trp Ž 0.49 2.05 Tyrosin Tyr Y 0.53 2.22 Valine Val PROTI 0.61 2.55
Dipólový moment
Spirála má overal dipólový moment vzhledem k agregačnímu účinku jednotlivých mikrodipolů z karbonyl skupiny peptidové vazby směřující podél osy šroubovice.[23] Účinky tohoto makrodipolu jsou předmětem určité kontroverze. α-šroubovice se často vyskytují s N-terminálním koncem vázaným záporně nabitou skupinou, někdy an aminokyselina postranní řetězec jako např glutamát nebo aspartát, nebo někdy fosfátový ion. Někteří považují makrodipol šroubovice za elektrostaticky interagující s takovými skupinami. Jiní mají pocit, že je to zavádějící, a je realističtější říci, že potenciál vodíkové vazby volných skupin NH na N-konci a-šroubovice lze uspokojit vodíkovou vazbou; toto lze také považovat za soubor interakcí mezi místními mikrodipoly, jako je C = O ··· H − N.[24][25]
Navinuté cívky
Šroubovité cívky α jsou vysoce stabilní formy, ve kterých se dvě nebo více šroubovic obtočí kolem sebe ve struktuře „supercoil“. Navinuté cívky obsahují vysoce charakteristickou vlastnost sekvenční motiv známý jako heptad opakovat, ve kterém se motiv opakuje každých sedm zbytků podél sekvence (aminokyselina zbytky, ne páry DNA bází). První a zejména čtvrtý zbytek (známý jako A a d pozice) jsou téměř vždy hydrofobní; čtvrtý zbytek je obvykle leucin - z toho vznikne název strukturální motiv volal a leucinový zip, což je typ stočené cívky. Tyto hydrofobní zbytky se shlukují ve vnitřku svazku šroubovice. Obecně platí, že pátý a sedmý zbytek ( E a G pozice) mají protichůdné náboje a tvoří solný most stabilizovaný pomocí elektrostatický interakce. Vláknité proteiny jako keratin nebo "stonky" z myosin nebo kinesin často přijímají struktury svinuté cívky, stejně jako několik dimerování bílkoviny. Pár stočených cívek - čtyřsvazek šroubovice - je velmi běžný strukturální motiv v proteinech. Například se vyskytuje u člověka růstový hormon a několik odrůd cytochrom. The Ropový protein, který podporuje replikaci plazmidu v bakteriích, je zajímavý případ, kdy jeden polypeptid tvoří svinutou cívku a dva monomery se skládají za vzniku svazku se čtyřmi šroubovicemi.
Uspořádání obličeje
Aminokyseliny, které tvoří konkrétní šroubovici, lze vynese na a spirálové kolo, reprezentace, která ilustruje orientace základních aminokyselin (viz článek pro leucinový zip pro takový diagram). Často v globulární proteiny, stejně jako ve specializovaných strukturách, jako jsou svinuté cívky a leucinové zipy, α-šroubovice bude vykazovat dvě „tváře“ - jednu obsahující převážně hydrofobní aminokyseliny orientované do vnitřku proteinu v hydrofobní jádro a jeden obsahující převážně polární aminokyseliny orientované na solventní -exponovaný povrch proteinu.
Ke změnám vazebné orientace dochází také u obličejově organizovaných oligopeptidů. Tento vzor je obzvláště běžný v antimikrobiální peptidy a bylo navrženo mnoho modelů, které popisují, jak to souvisí s jejich funkcí. Mnoho z nich má společné to, že hydrofobní povrch antimikrobiálního peptidu vytváří póry v plazmatické membráně po spojení s mastnými řetězci v jádru membrány.[26][27]
Větší sestavy

Myoglobin a hemoglobin, první dva proteiny, jejichž struktury byly vyřešeny rentgenem krystalografie, mají velmi podobné záhyby složené z přibližně 70% α-šroubovice, přičemž zbytek tvoří neopakující se oblasti nebo „smyčky“, které spojují šroubovice. Při klasifikaci proteinů podle jejich dominantního složení se Strukturální klasifikace proteinů databáze udržuje velkou kategorii speciálně pro proteiny all-α.
Hemoglobin má pak ještě větší měřítko kvartérní struktura, ve kterém je funkční molekula vázající kyslík tvořena čtyřmi podjednotkami.
Funkční role


Vazba DNA
α-Helices mají zvláštní význam v DNA vazebné motivy, včetně helix-turn-helix motivy, leucinový zip motivy a zinkový prst motivy. Důvodem je výhodný strukturální fakt, že průměr α-šroubovice je přibližně 12 Å (1,2 nm), včetně průměrné sady postranních řetězců, přibližně stejné jako šířka hlavní drážky ve formě B DNA, a také proto svinutá cívka (nebo leucinový zip) dimery šroubovic mohou snadno umístit pár interakčních povrchů, aby kontaktovaly druh symetrického opakování běžného v DNA s dvojitou spirálou.[28] Příkladem obou aspektů je transkripční faktor Max (viz obrázek vlevo), který používá spirálovitě stočenou cívku k dimerizaci, umístění další dvojice spirál pro interakci ve dvou po sobě jdoucích otočkách hlavní drážky DNA.
Překlenutí membrány
α-Helices jsou také nejběžnějším prvkem proteinové struktury, který prochází biologickými membránami (transmembránový protein ),[29] předpokládá se to, protože spirálovitá struktura může interně uspokojit všechny páteřní vodíkové vazby a nezanechává polární skupiny vystavené membráně, pokud jsou boční řetězce hydrofobní. Proteiny jsou někdy ukotveny jedinou spirálou překlenující membránu, někdy dvojicí a někdy svazkem spirál, většinou klasicky sestávajícím ze sedmi šroubovic uspořádaných nahoru a dolů v prstenci, například pro rhodopsiny (viz obrázek vpravo) nebo pro Receptory spojené s G proteinem (GPCR).
Mechanické vlastnosti
α-Helice pod axiální deformací v tahu, charakteristická podmínka zatížení, která se objevuje v mnoha vláknech a tkáních bohatých na alfa šroubovici, vede k charakteristickému třífázovému chování modulu tuhé-měkké-tuhé tangenty.[30] Fáze I odpovídá režimu malé deformace, během kterého se šroubovice homogenně protahuje, následovaná fází II, ve které se alfa-šroubovité otáčky zlomí zprostředkované prasknutím skupin H-vazeb. Fáze III je typicky spojena s protahováním kovalentní vazby s velkou deformací.
Dynamické vlastnosti
Alfa-helixy v bílkovinách mohou mít nízká frekvence akordeonový pohyb podle pozorování Ramanova spektroskopie[31] a analyzovány pomocí modelu kvazi-kontinua.[32][33] Helice nestabilizované terciárními interakcemi vykazují dynamické chování, které lze přičíst hlavně třepení šroubovice od konců.[34]
Přechod šroubovice – cívka
Homopolymery aminokyselin (např polylysin ) může při nízké teplotě převzít α-helikální strukturu, která se při vysokých teplotách „roztaví“. Tento přechod šroubovice – cívka kdysi se myslelo, že je analogický proteinu denaturace. The statistická mechanika tohoto přechodu lze modelovat pomocí elegantního přenosová matice metoda charakterizovaná dvěma parametry: sklon k zahájení šroubovice a sklon k prodloužení šroubovice.
V umění

Nejméně pět umělců se ve své práci výslovně zmínilo o šroubovici α: Julie Newdoll v malbě a Julian Voss-Andreae, Bathsheba Grossman, Byron Rubin a Mike Tyka v sochařství.
Umělkyně v San Francisku Julie Newdoll,[35] který vystudoval mikrobiologii s nezletilým v oboru umění, se specializuje na obrazy inspirované mikroskopickými obrazy a molekulami od roku 1990. Její obraz „Rise of the Alpha Helix“ (2003) představuje lidské postavy uspořádané do a šroubovického uspořádání. Podle umělce „květiny odrážejí různé typy postranních řetězců, které každá aminokyselina drží ve světě“.[35] Stejná metafora se odráží i ze strany vědce: „β listy nevykazují tuhou opakující se pravidelnost, ale plynou v půvabných křivkách kroucení, a dokonce i α-šroubovice je pravidelná spíše na způsob květního stonku, jehož větvící se uzly ukazují vliv prostředí, historie vývoje a vývoj každé části tak, aby odpovídala její vlastní idiosynkratické funkci. “[12]
Julian Voss-Andreae je německý sochař s tituly v experimentální fyzice a sochařství. Od roku 2001 Voss-Andreae vytváří „proteinové sochy“[36] založené na proteinové struktuře, přičemž α-šroubovice je jedním z jeho preferovaných objektů. Voss-Andreae vyrobil sochy šroubovice α-helix z různých materiálů včetně bambusu a celých stromů. Památník Voss-Andreae vytvořený v roce 2004 k oslavě památky Linus Pauling Objevitel α-šroubovice je vyroben z velkého ocelového nosníku přeskupeného ve struktuře α-šroubovice. 3 metry vysoká, jasně červená socha stojí před Paulingovým dětským domovem v Portland, Oregon.
Pásové diagramy α-šroubovice jsou prominentním prvkem v laserem leptaných krystalických plastikách proteinových struktur vytvořených umělcem Bathsheba Grossman, jako například z inzulín, hemoglobin, a DNA polymeráza.[37] Byron Rubin je bývalý krystalograf bílkovin, nyní profesionální sochař v oblasti proteinů, nukleových kyselin a molekul léčiv - z nichž mnohé obsahují α-helixy, jako například subtilisin, lidský růstový hormon, a fosfolipáza A2.[38]
Mike Tyka je výpočetní biochemik v University of Washington pracovat s David Baker. Tyka vyrábí sochy proteinových molekul od roku 2010 z mědi a oceli, včetně ubikvitin a a draslíkový kanál tetramer.[39]
Viz také
- 310 spirála
- Beta list
- Davydov soliton
- Skládací (chemie)
- Knoflíky do otvorů
- Šroubovice Pi
- Proteopedia Helices_in_Proteins
Reference
- ^ Kendrew JC, Dickerson RE, Strandberg BE, Hart RG, Davies DR, Phillips DC, Shore VC (únor 1960). „Struktura myoglobinu: trojrozměrná Fourierova syntéza při rozlišení 2 Å“. Příroda. 185 (4711): 422–7. Bibcode:1960Natur.185..422K. doi:10.1038 / 185422a0. PMID 18990802. S2CID 4167651.
- ^ Neurath H (1940). "Intramolekulární skládání polypeptidových řetězců ve vztahu k proteinové struktuře". Journal of Physical Chemistry. 44 (3): 296–305. doi:10.1021 / j150399a003.
- ^ Taylor HS (1942). "Velké molekuly prostřednictvím atomových brýlí". Sborník americké filozofické společnosti. 85 (1): 1–12. JSTOR 985121.
- ^ Huggins M (1943). "Struktura vláknitých proteinů". Chemické recenze. 32 (2): 195–218. doi:10.1021 / cr60102a002.
- ^ Bragg WL, Kendrew JC, Perutz MF (1950). "Konfigurace polypeptidového řetězce v krystalických proteinech". Sborník královské společnosti v Londýně. Řada A. Matematické a fyzikální vědy. 203 (1074): 321–?. Bibcode:1950RSPSA.203..321B. doi:10.1098 / rspa.1950.0142. S2CID 93804323.
- ^ Pauling L., Corey RB, Branson HR (Duben 1951). "Struktura proteinů; dvě vodíkově vázané šroubovicové konfigurace polypeptidového řetězce". Sborník Národní akademie věd Spojených států amerických. 37 (4): 205–11. Bibcode:1951PNAS ... 37..205P. doi:10.1073 / pnas.37.4.205. PMC 1063337. PMID 14816373.
- ^ „Nobelova cena za chemii 1954“.
- ^ Dunitz J. (2001). „Paulingova levoruká α-šroubovice“. Angewandte Chemie International Edition. 40 (22): 4167–4173. doi:10.1002 / 1521-3773 (20011119) 40:22 <4167 :: AID-ANIE4167> 3.0.CO; 2-Q. PMID 29712120.
- ^ Komise IUPAC-IUB pro biochemickou nomenklaturu (1970). "Zkratky a symboly pro popis konformace polypeptidových řetězců". Journal of Biological Chemistry. 245: 6489–6497.
- ^ „Polypeptidové konformace 1 a 2“. www.sbcs.qmul.ac.uk. Citováno 5. listopadu 2018.
- ^ Kabsch W, Sander C (prosinec 1983). "Slovník sekundární struktury bílkovin: rozpoznávání vzorů vodíkových vazeb a geometrických znaků". Biopolymery. 22 (12): 2577–637. doi:10,1002 / bip.360221211. PMID 6667333.
- ^ A b C Richardson JS (1981). "Anatomie a taxonomie proteinové struktury". Pokroky v chemii proteinů. 34: 167–339. doi:10.1016 / S0065-3233 (08) 60520-3. ISBN 9780120342341. PMID 7020376.
- ^ Lovell SC, Davis IW, Arendall WB, de Bakker PI, Word JM, Prisant MG, Richardson JS, Richardson DC (únor 2003). "Ověření struktury pomocí Calpha geometrie: odchylka phi, psi a Cbeta". Proteiny. 50 (3): 437–50. doi:10,1002 / prot.10286. PMID 12557186.
- ^ Dickerson RE, Geis I. (1969), Struktura a působení proteinů, Harper, New York
- ^ Zorko, Matjaž (2010). „Strukturální organizace proteinů“. V Langel, Ülo; Cravatt, Benjamin F.; Gräslund, Astrid; von Heijne, Gunnar; Land, Tiit; Niessen, Sherry; Zorko, Matjaž (eds.). Úvod do peptidů a proteinů. Boca Raton: CRC Press. 36–57. ISBN 9781439882047.
- ^ Terwilliger TC (březen 2010). „Rapid model building of alpha-helices in elektrone-density maps“. Acta Crystallographica oddíl D. 66 (Pt 3): 268–75. doi:10.1107 / S0907444910000314. PMC 2827347. PMID 20179338.
- ^ Hudgins RR, Jarrold MF (1999). „Tvorba spirály v neolvolovaných peptidech na bázi alaninu: spirálovité monomery a spirálovité dimery“. Journal of the American Chemical Society. 121 (14): 3494–3501. doi:10.1021 / ja983996a.
- ^ Kutchukian PS, Yang JS, Verdine GL, Shakhnovich EI (duben 2009). „All-atom model for stabilisation of alpha-helical structure in peptides by uhlovodíkové svorky“. Journal of the American Chemical Society. 131 (13): 4622–7. doi:10.1021 / ja805037p. PMC 2735086. PMID 19334772.
- ^ Abrusan G, Marsh JA (2016). „Alfa šroubovice jsou odolnější vůči mutacím než beta vlákna“. PLOS výpočetní biologie. 12 (12): e1005242. Bibcode:2016PLSCB..12E5242A. doi:10.1371 / journal.pcbi.1005242. PMC 5147804. PMID 27935949.
- ^ Rocklin GJ a kol. (2017). „Globální analýza skládání proteinů pomocí masivně paralelního designu, syntézy a testování“. Věda. 357 (6347): 168–175. Bibcode:2017Sci ... 357..168R. doi:10.1126 / science.aan0693. PMC 5568797. PMID 28706065.
- ^ Pace CN, Scholtz JM (červenec 1998). „Stupnice sklonu šroubovice založená na experimentálních studiích peptidů a proteinů“. Biofyzikální deník. 75 (1): 422–7. Bibcode:1998 BpJ .... 75..422N. doi:10.1016 / S0006-3495 (98) 77529-0. PMC 1299714. PMID 9649402.
- ^ Pace, C. Nick; Scholtz, J. Martin (1998). „Stupnice sklonu šroubovice založená na experimentálních studiích peptidů a proteinů“. Biofyzikální deník. 75. 422–427. Bibcode:1998 BpJ .... 75..422N. doi:10.1016 / s0006-3495 (98) 77529-0.
- ^ Hol WG, van Duijnen PT, Berendsen HJ (1978). "Dipól alfa šroubovice a vlastnosti proteinů". Příroda. 273 (5662): 443–446. Bibcode:1978Natur.273..443H. doi:10.1038 / 273443a0. PMID 661956. S2CID 4147335.
- ^ On JJ, Quiocho FA (říjen 1993). „Dominantní role místních dipólů při stabilizaci nekompenzovaných nábojů na síranu izolovaném v periplazmatickém aktivním transportním proteinu“. Věda o bílkovinách. 2 (10): 1643–7. doi:10.1002 / pro.5560021010. PMC 2142251. PMID 8251939.
- ^ Milner-White EJ (listopad 1997). „Částečný náboj atomu dusíku v peptidových vazbách“. Věda o bílkovinách. 6 (11): 2477–82. doi:10.1002 / pro.5560061125. PMC 2143592. PMID 9385654.
- ^ Kohn, Eric M .; Shirley, David J .; Arotsky, Lubov; Picciano, Angela M .; Ridgway, Zachary; Urban, Michael W .; Carone, Benjamin R .; Caputo, Gregory A. (02.02.2018). „Role kationtových postranních řetězců v antimikrobiální aktivitě C18G“. Molekuly. 23 (2): 329. doi:10,3390 / molekuly23020329. PMC 6017431. PMID 29401708.
- ^ Toke, Orsolya (2005). „Antimikrobiální peptidy: noví kandidáti v boji proti bakteriálním infekcím“. Biopolymery. 80 (6): 717–735. doi:10.1002 / bip.20286. ISSN 0006-3525. PMID 15880793.
- ^ Branden & Tooze, kapitola 10
- ^ Branden & Tooze, kapitola 12.
- ^ Ackbarow T, Chen X, Keten S, Buehler MJ (říjen 2007). „Hierarchie, více energetických bariér a robustnost určují lomovou mechaniku alfa-helikálních a beta-listových proteinových domén“. Sborník Národní akademie věd Spojených států amerických. 104 (42): 16410–5. Bibcode:2007PNAS..10416410A. doi:10.1073 / pnas.0705759104. PMC 2034213. PMID 17925444.
- ^ Painter PC, Mosher LE, Rhoads C (červenec 1982). "Nízkofrekvenční režimy v Ramanově spektru proteinů". Biopolymery. 21 (7): 1469–72. doi:10,1002 / bip.360210715. PMID 7115900.
- ^ Chou KC (prosinec 1983). „Identifikace nízkofrekvenčních režimů v proteinových molekulách“. The Biochemical Journal. 215 (3): 465–9. doi:10.1042 / bj2150465. PMC 1152424. PMID 6362659.
- ^ Chou KC (květen 1984). "Biologické funkce nízkofrekvenčních vibrací (fononů). III. Spirálové struktury a mikroprostředí". Biofyzikální deník. 45 (5): 881–9. Bibcode:1984BpJ .... 45..881C. doi:10.1016 / S0006-3495 (84) 84234-4. PMC 1434967. PMID 6428481.
- ^ Fierz B, Reiner A, Kiefhaber T (leden 2009). „Místní konformační dynamika v alfa šroubovicích měřená rychlým přenosem tripletů“. Sborník Národní akademie věd Spojených států amerických. 106 (4): 1057–62. Bibcode:2009PNAS..106.1057F. doi:10.1073 / pnas.0808581106. PMC 2633579. PMID 19131517.
- ^ A b „Julie Newdoll Vědecky inspirované umění, hudba, deskové hry“. www.brushwithscience.com. Citováno 2016-04-06.
- ^ Voss-Andreae J (2005). „Protein Sculptures: Life's Building Blocks Inspire Art“. Leonardo. 38: 41–45. doi:10.1162 / leon.2005.38.1.41. S2CID 57558522.
- ^ Grossman, Bathsheba. „About the Artist“. Socha Betsabé. Citováno 2016-04-06.
- ^ "O". moleculesculpture.com. Citováno 2016-04-06.
- ^ Tyka, Mike. "O". www.miketyka.com. Citováno 2016-04-06.
Další čtení
- Tooze J, Brändén C (1999). Úvod do proteinové struktury. New York: Garland Pub. ISBN 0-8153-2304-2..
- Eisenberg D (září 2003). „Objev alfa-šroubovice a beta-listu, hlavních strukturálních rysů proteinů“. Sborník Národní akademie věd Spojených států amerických. 100 (20): 11207–10. Bibcode:2003PNAS..10011207E. doi:10.1073 / pnas.2034522100. PMC 208735. PMID 12966187.
- Astbury WT, Woods HJ (1931). "Molekulární hmotnosti proteinů". Příroda. 127 (3209): 663–665. Bibcode:1931Natur.127..663A. doi:10.1038 / 127663b0. S2CID 4133226.
- Astbury WT, ulice A (1931). „Rentgenové studie struktur vlasů, vlny a příbuzných vláken. I. Obecně“. Trans. R. Soc. Lond. A230: 75–101. Bibcode:1932RSPTA.230 ... 75A. doi:10.1098 / rsta.1932.0003.
- Astbury WT (1933). "Některé problémy v rentgenové analýze struktury zvířecích chloupků a jiných proteinových vláken". Trans. Faraday Soc. 29 (140): 193–211. doi:10.1039 / tf9332900193.
- Astbury WT, Woods HJ (1934). „Rentgenové studie struktur vlasů, vlny a příbuzných vláken. II. Molekulární struktura a elastické vlastnosti vlasového keratinu“. Filozofické transakce Royal Society of London Series A. 232 (707–720): 333–394. Bibcode:1934RSPTA.232..333A. doi:10.1098 / rsta.1934.0010.
- Astbury WT, Sisson WA (1935). „Rentgenové studie struktur vlasů, vlny a příbuzných vláken. III. Konfigurace molekuly keratinu a její orientace v biologické buňce“. Sborník Královské společnosti. A150 (871): 533–551. Bibcode:1935RSPSA.150..533A. doi:10.1098 / rspa.1935.0121.
- Sugeta H, Miyazawa T (1967). "Obecná metoda pro výpočet spirálových parametrů polymerních řetězců z délek vazeb, úhlů vazeb a úhlů vnitřní rotace". Biopolymery. 5 (7): 673–679. doi:10,1002 / bip.1967.360050708.
- Wada A (1976). „Alfa-šroubovice jako elektrický makro-dipól“. Pokroky v biofyzice: 1–63. PMID 797240.
- Chothia C, Levitt M, Richardson D (říjen 1977). "Struktura proteinů: balení alfa-šroubovic a skládaných listů". Sborník Národní akademie věd Spojených států amerických. 74 (10): 4130–4. Bibcode:1977PNAS ... 74.4130C. doi:10.1073 / pnas.74.10.4130. PMC 431889. PMID 270659.
- Chothia C, Levitt M, Richardson D (leden 1981). "Helix to helix packing in protein". Journal of Molecular Biology. 145 (1): 215–50. doi:10.1016/0022-2836(81)90341-7. PMID 7265198.
- Hol WG (1985). „Role dipólu alfa-šroubovice ve funkci a struktuře proteinu“. Pokrok v biofyzice a molekulární biologii. 45 (3): 149–95. doi:10.1016 / 0079-6107 (85) 90001-X. PMID 3892583.
- Barlow DJ, Thornton JM (červen 1988). "Geometrie šroubovice v proteinech". Journal of Molecular Biology. 201 (3): 601–19. doi:10.1016/0022-2836(88)90641-9. PMID 3418712.
- Murzin AG, Finkelstein AV (prosinec 1988). "Obecná architektura alfa-šroubovicového globulu". Journal of Molecular Biology. 204 (3): 749–69. doi:10.1016 / 0022-2836 (88) 90366-X. PMID 3225849.
externí odkazy
- NetSurfP ver. 1.1 - Dostupnost povrchu bílkovin a předpovědi sekundární struktury
- Kalkulačka rotačního úhlu α-šroubovice
- Web umělkyně Julie Newdoll
- Web umělce Juliana Voss-Andreae
| Křivky |  | ||
|---|---|---|---|
| Helices |
| ||
| Spirály | |||