Neurogeneze dospělých - Adult neurogenesis - Wikipedia

{kind=link}

Neurogeneze dospělých je proces, ve kterém neurony jsou generovány z nervové kmenové buňky u dospělých. Tento proces se liší od prenatální neurogeneze.
U většiny savců se nové neurony rodí v průběhu dospělosti ve dvou oblastech mozku:[3]
- The subgranulární zóna (SGZ), součást zubatý gyrus z hipokampus,[4][5] kde se rodí neurální kmenové buňky granulované buňky (podílí se na tvorbě paměti a učení).[Citace je zapotřebí ]
- The subventrikulární zóna (SVZ) boční komory, lze rozdělit do tří mikrodomén - laterální, dorzální a mediální).[6] Neurální kmenové buňky migrují do čichová žárovka skrz rostrální migrační proud kde se liší interneurony účast v čich. U lidí se však po narození vytvoří jen málo neuronů čichové cibulky.[7][8]
Větší pozornost byla věnována neurogenezi v zubatý gyrus než v striatum. v hlodavci mnoho novorozených zubatých gyrusových neuronů umírá krátce po narození,[4] ale řada z nich se funkčně integruje do okolní mozkové tkáně.[9][10][11] Počet neuronů narozených v lidském hipokampu zůstává kontroverzní; některé studie uvádějí, že u dospělých lidí se každý den přidá do hipokampu asi 700 nových neuronů,[12] zatímco jiné studie ukazují, že dospělá hipokampální neurogeneze u lidí neexistuje, nebo pokud ano, je na nedetekovatelné úrovni.[13] Role nových neuronů ve funkci mozku dospělého tak zůstává nejasná. Uvádí se, že neurogeneze dospělých hraje roli v učení a paměti, emocích, stresu, depresi, reakci na zranění a dalších podmínkách.[14]
Mechanismus
Dospělé nervové kmenové buňky
Neurální kmenové buňky (NSC) jsou samoobnovující se, multipotentní buňky, které generují hlavní fenotypy z nervový systém.
Přeprogramování počtu řádků (trans-diferenciace)
Objevující se důkazy naznačují, že neurální mikrovaskulární pericyty jsou podle pokynů rezidentních gliových buněk přeprogramovány na interneurony a obohacují místní neuronové mikroobvody.[15] Tato reakce je zesílena souběžným podáváním angiogeneze.
Modelové organismy neurogeneze
Planarian
Planarian jsou jedním z prvních modelové organismy zvyklý studovat regenerace s Pallas jako praotec planaristických studií. Planarian jsou klasický model bezobratlých, který se v posledních desetiletích používá ke zkoumání neurogeneze. Centrální nervový systém planaristy je jednoduchý, i když plně vytvořený se dvěma laloky umístěnými v hlavě a dvěma ventrální nervové šňůry. Tento model reprodukuje nepohlavně a produkuje kompletní a plně funkční nervový systém po dělení, což umožňuje důsledné vyšetření neurogeneze.
Axolotl
The axolotl je méně běžně používaný než jiné obratlovce, ale stále je klasickým modelem pro zkoumání regenerace a neurogeneze. Ačkoli si axolotl našel místo v biomedicínském výzkumu z hlediska regenerace končetin,[16][17] modelový organismus vykazoval silnou schopnost generovat nové neurony po poškození.[18][19] Axolotly přispěly jako přemosťovací organismus mezi bezobratlými a savci, protože tento druh má regenerační schopnost podstoupit úplnou neurogenezi a vytvořit širokou škálu neuronových populací, které se neomezují pouze na malý výklenek,[20] složitost a architektura jsou však v mnoha ohledech složité a analogické lidskému neurálnímu vývoji.
Zebrafish
Zebrafish byly dlouho klasikou vývojový kvůli jejich průhlednosti během organogeneze a byly těžce využívány v časné neurogenezi vývoje.[21][22]). Zebrafish vykazuje silnou neurogenerativní schopnost schopnou regenerovat různé tkáně a úplnou neuronální diverzitu (s výjimkou astrocyty, jak je ještě třeba identifikovat v mozku zebrafish) s pokračující neurogenezí po celou dobu života. V posledních desetiletích model upevnil svoji roli v regeneraci dospělých a neurogenezi po poškození.[23][24][25] Zebrafish, stejně jako axolotl, hrál klíčovou roli jako přemosťovací organismus mezi bezobratlými a savci. Zebrafish je rychle se rozvíjející organismus, jehož údržba je relativně levná a zároveň poskytuje pole snadnou genetickou manipulaci a komplexní nervový systém.
kuřátko
Ačkoli se ptáci používali především ke studiu raného embryonálního vývoje, v posledních desetiletích se vývoj vyvíjel kuřátko hraje klíčovou roli při vyšetřování neurogeneze a regenerace, protože mladá kuřata je v mladém věku schopna neuronového obratu, ale do dospělosti ztrácí neurogenerativní kapacitu.[26] Ztráta neuroregenerativní schopnosti během zrání umožnila vyšetřovatelům dále zkoumat genetické regulátory neurogeneze.
Hlodavci
Hlodavci, myši a krysy, byly nejvýznamnějším modelovým organismem od objevu moderních neuronů autorem Santiago Ramon y Cajal. Hlodavci mají velmi podobnou architekturu a složitý nervový systém s velmi malou regenerační schopností podobnou té u lidí. Z tohoto důvodu byly hlodavci hojně využívány předklinické testování. Hlodavci vykazují širokou škálu nervových obvodů odpovědných za složité chování, což je činí ideálními pro studium dendritického prořezávání a axonálního střihu.[27] I když organismus vytváří silný lidský analog, model má svá omezení, která nebyla nalezena v předchozích modelech: vyšší náklady na údržbu, nižší počty chovatelů a omezené neurogenerativní schopnosti.
Chobotnice
Také známý jako obyčejná chobotnice A, hlavonožce, tento organismus má složitý nervový systém, který demonstruje schopnost mozku produkovat nové buňky. V tomto případě a v porovnání s jinými taxony se tyto organismy přizpůsobují nepředvídatelnému prostředí pomocí nově vytvořených mozkových buněk.[28] Je to po krátkou dobu života (samice asi jeden rok), kdy divoké běžné chobotnice soustředí většinu své energie na páření a péči o potomky.[29][30] Nálezy naznačují, že chobotnice obecná má stejně jako jiné druhy s krátkým životem komplexní proliferaci hipokampu,[31][32] potřebné pro prostorovou / navigaci a krátkodobou a dlouhodobou paměť.[33][kruhový odkaz ]
Sledování neurogeneze
Vytvoření nové funkční neurony lze měřit několika způsoby,[34] shrnuty v následujících částech.
Značení DNA
Označené DNA může sledovat linii dělící se buňky a určit umístění jejích dceřiných buněk. A nukleová kyselina analog je vložen do genom buňky vytvářející neurony (například a gliová buňka nebo nervová kmenová buňka ).[35] Tymin analogy (3H) thymidin[36] a BrdU[37] jsou běžně používané DNA štítky a jsou používány pro radioaktivní značení a imunohistochemie resp.
Stanovení osudu pomocí markerů neuronové linie
DNA označení lze použít ve spojení s markery neuronové linie určit osud nových funkčních mozkové buňky. Za prvé, začleněny označené nukleotidy se používají k detekci populací nově rozdělené dceřiné buňky. Specifické typy buněk jsou poté určeny s jedinečnými rozdíly v jejich výraz z bílkoviny, které lze použít jako antigeny v imunotest. Například, NeuN / Fox3 a GFAP jsou antigeny běžně používané k detekci neurony, glia, a ependymální buňky. Ki67 je nejčastěji používaný antigen k detekci proliferace buněk. Nějaký antigeny lze použít k měření konkrétních kmenová buňka etapy. Například kmenové buňky vyžadují sox2 gen udržovat pluripotence a používá se k detekci trvalých koncentrací kmenových buněk v Tkáň CNS. The protein nestin je střední vlákno, což je nezbytné pro radiální růst axony, a proto se používá k detekci tvorby nových synapse.
Cre-Lox rekombinace
Některé studie genetického sledování využívají rekombinace cre-lox svázat a promotér do a reportér gen, jako lacZ nebo Gen GFP.[35][38] Tuto metodu lze použít pro dlouhodobou kvantifikaci buněčného dělení a značení, zatímco výše uvedené postupy jsou užitečné pouze pro krátkodobou kvantifikaci.
Virové vektory
V poslední době se stále častěji používá rekombinantní viry pro vložení genetická informace specifické pro kódování markery (obvykle protein fluorofory jako GFP ), které jsou pouze vyjádřený v buňky určitého druhu. Je vložen markerový gen po proudu a promotér, vedoucí k transkripce tohoto markeru pouze v buňkách obsahujících transkripční faktory které se vážou k tomuto promotoru. Například rekombinantní plazmid může obsahovat promotor pro doublecortin, protein exprimovaný převážně neurony, před sekvencí kódující GFP, čímž se vytvoří infikované buňky fluoreskují zelená po vystavení světlu v rozmezí od modré do ultrafialového záření[39] při odchodu mimo doublecortin buňky bez ovlivnění, i když obsahují plazmid. Mnoho buněk bude obsahovat více kopií plazmidu a samotného fluoroforu, což umožní přenos fluorescenčních vlastností podél infikované buňky linie.
Označením buňky, která vede k vzniku neuronů, jako je a nervové kmenové buňky nebo neurálních prekurzorových buněk lze sledovat tvorbu, proliferaci a dokonce migraci nově vytvořených neuronů.[40] Je však důležité si uvědomit, že zatímco plazmid je stabilní po dlouhou dobu, jeho proteinové produkty mohou mít vysoce variabilní půl života a jejich fluorescence se může snižovat a také se příliš zředit, aby byla viditelná v závislosti na počtu replikačních cyklů, které podstoupily, což činí tuto metodu užitečnější pro sledování podobných neurálních prekurzorů nebo neurálních kmenových buněk než samotných neuronů. Vložení genetický materiál přes a virový vektor má tendenci být sporadické a řídké vzhledem k celkovému počtu buněk v dané oblasti tkáň, což činí kvantifikaci buněčného dělení nepřesnou. Výše uvedená metoda však může poskytnout vysoce přesná data s ohledem na to, kdy buňka byla narozený stejně jako plné buňky morfologie.[41]
Metody pro inhibici neurogeneze
Mnoho studií analyzujících roli neurogeneze dospělých používá metodu inhibice buněčné proliferace ve specifických oblastech mozku, napodobující inhibici neurogeneze, ke sledování účinků na chování.[14]
Farmakologická inhibice
Farmakologická inhibice je široce používána v různých studiích, protože poskytuje mnoho výhod. To je obecně levné ve srovnání s jinými metodami, jako je ozařování, lze použít na různé druhy a nevyžaduje žádné invazivní postupy nebo operace pro subjekty.
Představuje však určité výzvy, protože tyto inhibitory nelze použít k inhibici proliferace ve specifických oblastech, což vede k nespecifickým účinkům ovlivňovaným jinými systémy. Aby se těmto účinkům zabránilo, je třeba více pracovat na stanovení optimálních dávek, aby se minimalizovaly účinky na systémy nesouvisející s neurogenezí.
Běžným farmakologickým inhibitorem pro neurogenezi dospělých je methylazoxymethanoacetát (MAM), chemoterapeutikum. Dalšími inhibitory buněčného dělení běžně používanými ve studiích jsou cytarabin a temozolomid.
Farmakogenetika
Další metodou používanou ke studiu účinků neurogeneze dospělých je použití farmakogenetických modelů. Tyto modely poskytují různé výhody z farmakologické cesty, protože umožňují větší specificitu zaměřením konkrétních prekurzorů neurogeneze a specifických promotorů kmenových buněk. Umožňuje také časovou specificitu s interakcí určitých léků. To je prospěšné při konkrétním pohledu na neurogenezi v dospělosti, po normálním vývoji jiných oblastí v mozku.
Thymidinkináza viru herpes simplex (HSV-TK) byla použita ve studiích ve spojení s antivirotiky k inhibici neurogeneze dospělých. Funguje tak, že cílí na kmenové buňky pomocí gliových fibrilárních kyselých proteinů a exprese nestinu. Tyto cílené kmenové buňky podléhají buněčné smrti místo buněčné proliferace, když jsou vystaveny antivirovým lékům.
Cre protein se také běžně používá při cílení na kmenové buňky, které po léčbě tamoxifenem podstoupí genové změny.
Ozáření
Ozařování je metoda, která umožňuje velmi specifickou inhibici neurogeneze dospělých. Může být zaměřen na mozek, aby nedošlo k ovlivnění jiných systémů a mělo nespecifické účinky. Může být dokonce použit k cílení na konkrétní oblasti mozku, což je důležité při určování toho, jak neurogeneze dospělých v různých oblastech mozku ovlivňuje chování.
Ozařování je však dražší než jiné metody a vyžaduje také velké vybavení vyškolenými jednotlivci.
Inhibice neurogeneze dospělých v hipokampu
Mnoho studií pozorovalo, jak inhibice dospělé neurogeneze u jiných savců, jako jsou krysy a myši, ovlivňují jejich chování.[14] Bylo prokázáno, že inhibice neurogeneze dospělých v hipokampu má různé účinky na učení a paměť, kondicionování a vyšetřovací chování.
Porucha podmíněnosti strachu byla pozorována ve studiích zahrnujících potkany s nedostatkem neurogeneze dospělých v hipokampu.[42] Inhibice neurogeneze dospělých v hipokampu byla také spojena se změnami chování při vyšetřovacích úkolech.[43] Krysy také vykazují snížené kontextualizované mrazivé chování v reakci na kontextualizovaný strach a zhoršení v učení prostorových umístění, když chybí neurogeneze dospělých.[44][45]
Účinky na oddělení vzorů
Změny v učení a paměti pozorované ve výše zmíněných studiích se považují za související s rolí neurogeneze dospělých v regulaci oddělování vzorů.[14] Oddělení vzorů je definováno jako „proces odstraňování nadbytečnosti z podobných vstupů, aby bylo možné oddělit události od sebe navzájem a omezit rušení a navíc může vytvořit více ortogonální, řídkou a kategorizovanou sadu výstupů.“[46]
Toto narušení oddělení vzorů by mohlo vysvětlit narušení viděné v jiných úlohách učení a paměti. Snížená schopnost snižovat rušení by mohla vést k větším obtížím při vytváření a uchovávání nových vzpomínek.,[14] ačkoli je obtížné rozlišovat mezi účinky neurogeneze na učení a oddělováním vzorců kvůli omezením ve výsledcích interpretace chování. “[47]
Studie ukazují, že krysy s inhibovanou dospělou neurogenezí prokazují potíže s diferenciací a učením kontextualizované úpravy strachu.[14] Krysy s blokovanou neurogenezí dospělých také vykazují zhoršené diferenciální zmrazení, když je zapotřebí rozlišovat mezi podobnými kontexty.[48] To také ovlivňuje jejich prostorové rozpoznávání v testech bludiště s radiálním ramenem, když jsou paže blíže k sobě, než když jsou dále od sebe.[49] Metaanalýza studií chování hodnotících účinek neurogeneze v různých testech separace vzorů ukázala konzistentní účinek ablace neurogeneze na výkon, i když v literatuře existují výjimky. "[50]
Účinky na inhibici chování
Inhibice chování je u potkanů a jiných zvířat důležitá při zastavení všeho, co právě dělají, aby bylo možné znovu posoudit situaci v reakci na hrozbu nebo cokoli jiného, co může vyžadovat jejich pozornost.[14]
Krysy s poškozenými hipokampy vykazují menší inhibici chování, jsou-li vystaveny hrozbám, jako je kočičí zápach.[51] Narušení normální buněčné proliferace a vývoje zubatého gyrusu u vyvíjejících se potkanů také zhoršuje jejich zmrazovací reakci, což je příklad inhibice chování, když je vystaven neznámému dospělému samci krysy.[52]
Toto zhoršení inhibice chování také souvisí s procesem učení a paměti, protože potlačování nesprávných odpovědí nebo chování vyžaduje schopnost inhibovat tuto reakci.[14]
Dopady
Role v učení
Funkční význam neurogeneze dospělých je nejistý,[53] ale existují určité důkazy, pro které je hipogenní neurogeneze dospělých důležitá učení se a Paměť.[54] Bylo navrženo několik mechanismů pro vztah mezi zvýšenou neurogenezí a zlepšeným poznáním, včetně výpočetních teorií, které ukazují, že nové neurony zvyšují kapacitu paměti,[55] snížit rušení mezi vzpomínkami,[56] nebo přidat informace o čase do vzpomínek.[57] Vzhledem k tomu, že rychlost neurogeneze se během dospělosti podstatně nemění, bylo navrženo toto jedinečné epizodické vzpomínky lze vytvořit jednoduchým spoléháním se na zvýšenou kapacitu mladých neuronů určitého věku založit stabilní nové synapse s vrstevníky představujícími jedinečné rysy události, která má být zapamatována [58] Experimenty zaměřené na ablaci neurogeneze se ukázaly neprůkaznými, ale několik studií navrhlo neurogenní závislost u některých typů učení,[59] a ostatní nevidí žádný účinek.[60] Studie prokázaly, že samotný akt učení je spojen se zvýšeným přežíváním neuronů.[61] Celková zjištění, že neurogeneze dospělých je důležitá pro jakýkoli druh učení, jsou však nejednoznačná.
Alzheimerova choroba
Některé studie naznačují, že snížená hipogenní neurogeneze může vést k rozvoji Alzheimerova choroba (INZERÁT).[62] Jiní přesto předpokládají, že pacienti s AD mají zvýšenou neurogenezi v oblasti CA1 Ammonova rohu (hlavní oblast AD hipokampální patologie), aby kompenzovali ztrátu neuronů.[63] Přesná povaha vztahu mezi neurogenezí a Alzheimerovou chorobou není známa, růstový faktor podobný inzulínu 1 stimulovaná neurogeneze produkuje velké změny v hipokampální plasticitě a zdá se, že se podílí na Alzheimerově patologii.[64] Allopregnanolon, a neurosteroid, pomáhá pokračující neurogenezi v mozku. Úrovně alopregnanolonu v mozku klesají v starý věk a Alzheimerova choroba.[65] Bylo prokázáno, že alopregnanolon zvrací narušení neurogeneze, aby zvrátil kognitivní deficity v model myši Alzheimerovy choroby.[66] Eph receptory a ephrin Bylo prokázáno, že signalizace reguluje neurogenezi dospělých v hipokampus a byly studovány jako potenciální cíle pro léčbu některých příznaků AD.[67] Molekuly spojené s patologií AD, včetně ApoE, PS1 a APLIKACE Bylo také zjištěno, že ovlivňují neurogenezi dospělých v hipokampu.[68]
Role ve schizofrenii
Studie naznačují, že lidé s schizofrenie mají snížený objem hipokampu, o kterém se předpokládá, že je způsoben snížením neurogeneze dospělých. Odpovídajícím způsobem může být tento jev základní příčinou mnoha příznaků onemocnění. Několik výzkumných prací dále zmiňovalo čtyři geny, protein vázající dystrobrevin 1 (DTNBP1), neuregulin 1 (NRG1), narušený u schizofrenie 1 (DISC1) a receptor neuregulinu 1 (ERBB4), protože jsou pravděpodobně odpovědné za tento deficit za normálních okolností regenerace neuronů.[69][70] Podobnosti mezi depresí a schizofrenií naznačují možnou biologickou souvislost mezi těmito dvěma nemocemi. Je však třeba provést další výzkum, aby byl tento vztah jasně prokázán.[71]
Neurogeneze dospělých a velká depresivní porucha
Výzkum naznačuje, že neurogeneze hipokampu dospělých je nepřímo úměrná depresi (MDD).[72] Neurogeneze je snížena v hipokampu na zvířecích modelech závažné depresivní poruchy a mnoha způsobech léčby této poruchy, včetně antidepresivum léky a elektrokonvulzivní terapie, zvyšují hipokampální neurogenezi. Předpokládá se, že snížená hipokampální neurogeneze u jedinců s depresivní poruchou může souviset s vysokou hladinou stresových hormonů nazývaných glukokortikoidy, které jsou také spojeny s poruchou. Hippocampus dává pokyny osa hypotalamus-hypofýza-nadledviny produkovat méně glukokortikoidů, když jsou hladiny glukokortikoidů vysoké. Nefunkční hipokampus by proto mohl vysvětlit chronicky vysoké hladiny glukokortikoidů u jedinců s velkou depresivní poruchou. Některé studie však ukázaly, že hipokampální neurogeneze není nižší u jedinců s velkou depresivní poruchou a že hladiny glukokortikoidů v krvi se nemění, když se hipokampální neurogeneze mění, takže asociace jsou stále nejisté.
Stres a deprese
Mnoho lidí nyní věří, že stres je nejdůležitějším faktorem pro nástup Deprese, kromě genetiky. Jak bylo diskutováno výše, hipokampální buňky jsou citlivé na stres, který může vést ke snížení neurogeneze. Tato oblast je častěji zvažována při zkoumání příčin a léčby deprese. Studie ukázaly, že odstranění nadledvina u potkanů způsobila zvýšenou neurogenezi v zubatý gyrus.[73] Nadledvina je zodpovědná za produkci kortizol v reakci na stresor, látku, která při své výrobě v chronickém množství způsobuje regulaci dolů serotonin receptory a potlačuje vznik neuronů.[74] Ve stejné studii bylo prokázáno, že podávání kortikosteronu normálním zvířatům potlačilo neurogenezi, opačný účinek.[73] Nejtypičtější skupinou antidepresiv podávaných na toto onemocnění jsou selektivní inhibitory zpětného vychytávání serotoninu (SSRI)[75] a jejich účinnost lze vysvětlit neurogenezí. V normálním mozku způsobuje zvýšení serotoninu potlačení hormon uvolňující kortikotropin (CRH) prostřednictvím připojení k hipokampu. Přímo působí na paraventrikulární jádro snížit uvolňování CRH a regulovat fungování noradrenalinu v locus coeruleus.[73] Vzhledem k tomu, že je CRH potlačován, je zvrácen také pokles neurogeneze, který je spojen se zvýšenými hladinami. To umožňuje produkci více mozkových buněk, zejména na receptoru 5-HT1a v zubatém gyrusu hipokampu, u kterého bylo prokázáno, že zlepšuje příznaky deprese. Obvykle trvá neuronům přibližně tři až šest týdnů, než dospějí,[76] což je přibližně stejné množství času, než se SSRI projeví. Tato korelace posiluje hypotézu, že SSRI působí prostřednictvím neurogeneze ke snížení příznaků deprese. Někteří neurologové vyjádřili skepsi, že neurogeneze je funkčně významná, vzhledem k tomu, že malý počet rodících se neuronů je skutečně integrován do existujících neurálních obvodů. Nedávná studie však použila ozařování rodících se hipokampálních neuronů u subhumánních primátů (NHP) k prokázání, že pro antidepresivní účinnost je nutná neurogeneze.[77]
Zdá se, že neurony narozené v dospělosti hrají roli v regulaci stres.[78][79] Studie spojily neurogenezi s prospěšnými účinky specifické antidepresiva, což naznačuje souvislost mezi sníženou hipokampální neurogenezí a depresí.[80][81] V průkopnické studii vědci prokázali, že behaviorální výhody podávání antidepresiv v myši je obrácen, když je zabráněno neurogenezi pomocí ozáření x techniky.[82] Ve skutečnosti jsou novorozené neurony vzrušivější než starší neurony kvůli rozdílné expresi GABA receptory.[83] Možným modelem proto je, že tyto neurony zvyšují roli hipokampu v mechanismu negativní zpětné vazby Osa HPA (fyziologický stres) a možná při potlačení amygdala (oblast mozku odpovědná za bázlivé reakce na podněty).[vágní ] Potlačení neurogeneze dospělých může skutečně vést ke zvýšené stresové reakci na ose HPA v mírně stresových situacích.[78] To je v souladu s četnými nálezy spojujícími činnosti uvolňující stres (učení, vystavení novému, ale neškodnému prostředí a cvičení) se zvýšenou úrovní neurogeneze, stejně jako pozorování, že zvířata vystavená fyziologickému stresu (kortizolu) nebo psychický stres (např. izolace) vykazují výrazně sníženou hladinu novorozených neuronů. Za podmínek chronického stresu zvyšuje elevace novorozených neuronů antidepresivy hipokampální závislost na stresové reakci; bez novorozených neuronů nejsou antidepresiva schopna obnovit regulaci stresové reakce a zotavení je nemožné.[79]
Některé studie předpokládají, že učení a paměť jsou spojeny s depresí a že to může podporovat neurogeneze neuroplasticity. Jedna studie navrhuje, že náladu lze regulovat na základní úrovni plasticitou, a tedy ne chemie. Účinky antidepresiv by tedy byly pouze druhotné ke změně plasticity.[84] Jiná studie však prokázala interakci mezi antidepresivy a plasticitou; Bylo prokázáno, že antidepresivum fluoxetin obnovuje plasticitu v mozku dospělého potkana.[85] Výsledky této studie naznačují, že antidepresivní terapie by ji mohla podporovat místo změn v plasticitě.
Účinky redukce spánku
Jedna studie spojila nedostatek spánku se snížením neurogeneze hipokampu u hlodavců. Navrhovaným mechanismem pro pozorovaný pokles byly zvýšené hladiny glukokortikoidy. Ukázalo se, že dva týdny nedostatek spánku působil jako inhibitor neurogeneze, který byl po návratu do normálního spánku zvrácen a dokonce se posunul k dočasnému zvýšení normální buněčné proliferace.[86] Přesněji řečeno, když jsou hladiny kortikosteronu zvýšené, deprivace spánku tento proces brzdí. Normální hladiny neurogeneze po chronické spánkové deprivaci se nicméně vrátí po 2 týdnech s dočasným zvýšením neurogeneze.[87]I když je to známo, přehlížena je poptávka po glukóze v krvi, která se projevuje během dočasných diabetických hypoglykemických stavů. Americká diabetická asociace mimo jiné dokumentuje pseudosilienci a agitovanost zjištěnou během dočasných hypoglykemických stavů. Je zapotřebí mnohem více klinické dokumentace, aby bylo možné kompetentně prokázat souvislost mezi sníženou hematologickou glukózou a neuronální aktivitou a náladou.
Možné použití při léčbě Parkinsonovy choroby
Parkinsonova choroba je neurodegenerativní porucha charakterizovaná progresivní ztrátou dopaminergní neurony v substantia nigra. Transplantace plodu dopaminergní prekurzorové buňky připravila cestu pro možnost buněčné substituční terapie, která by mohla zlepšit klinické příznaky u postižených pacientů.[88] V posledních letech poskytli vědci důkazy o existenci nervových kmenových buněk s potenciálem produkovat v mozku dospělých savců nové neurony, zejména dopaminergního fenotypu.[89][90][91] Experimentální vyčerpání dopaminu u hlodavců snižuje proliferaci prekurzorových buněk jak v subependymální zóně, tak v subgranulární zóně.[92] Proliferace je zcela obnovena selektivním agonistou receptorů podobných D2 (D2L).[92] Nervové kmenové buňky byly identifikovány v neurogenních oblastech mozku, kde neustále probíhá neurogeneze, ale také v jiných než neurogenních zónách, jako je střední mozek a striatum, kde se za normálních fyziologických podmínek nepředpokládá, že by k neurogenezi došlo.[88] Novější výzkum ukázal, že ve striatu ve skutečnosti existuje neurogeneze.[93]Podrobné porozumění faktorům ovlivňujícím dospělé nervové kmenové buňky in vivo může nakonec vést k elegantním buněčným terapiím pro neurodegenerativní poruchy, jako je Parkinsonova choroba, mobilizací autologních endogenních nervových kmenových buněk k nahrazení degenerovaných neuronů.[88]
Traumatické zranění mozku
Traumatické poranění mozku se liší v mechanismu poranění, vyvolávají tupé nebo pronikavé trauma vedoucí k a primární a sekundární zranění s excitotoxicita a relativně široká neuronová smrt. Vzhledem k ohromnému počtu traumatických poranění mozku v důsledku Válka proti teroru, bylo věnováno obrovské množství výzkumu k lepšímu pochopení patofyziologie traumatických poranění mozku i neuroprotektivní intervence a možné intervence vyvolávající regenerační neurogenezi. Hormonální intervence, jako je progesteron, estrogen a alopregnanolon, byly v posledních desetiletích intenzivně zkoumány jako možná neuroprotektivní činidla po traumatickém poranění mozku, aby se snížila zánětlivá odezva na neuronovou smrt.[94][95][96][97] U hlodavců, kterým chybí regenerační kapacita pro neurogenezi dospělých, aktivace kmenových buněk po podání a7 agonisty nikotinového acetylcholinového receptoru, PNU-282987, byl identifikován u poškozených sítnic následnou prací zkoumající aktivaci neurogeneze u savců po traumatickém poranění mozku.[98] V současné době neexistuje žádný lékařský zásah, který by prošel fáze III klinické studie pro použití u lidské populace.
Faktory ovlivňující
Změny ve stáří
Neurogeneze je v hipokampu starých zvířat podstatně snížena, což zvyšuje možnost, že může být spojena s poklesem funkce hipokampu souvisejícím s věkem. Například rychlost neurogeneze u starých zvířat předpovídá paměť.[99] Nově narozené buňky ve věku zvířat jsou však funkčně integrovány.[100] Vzhledem k tomu, že k neurogenezi dochází po celý život, lze očekávat, že se hipokampus bude v dospělosti neustále zvětšovat, a proto se u starých zvířat zvýší počet buněk granulí. To však není tento případ, což naznačuje, že proliferace je vyvážena smrtí buněk. Nejde tedy o přidání nových neuronů do hipokampu, které se zdá být spojeno s hipokampálními funkcemi, ale spíše o rychlost přeměny buněk granulí.[101]
Účinky cvičení
Vědci prokázali, že fyzická aktivita ve formě dobrovolného cvičení vede ke zvýšení počtu novorozených neuronů v hipokampu myší a potkanů.[102][103] Tyto a další studie ukázaly, že učení u obou druhů lze zlepšit tělesným cvičením.[104] Nedávný výzkum to ukázal neurotrofický faktor odvozený z mozku a růstový faktor podobný inzulínu 1 jsou klíčovými mediátory neurogeneze vyvolané cvičením.[103][105] Cvičení zvyšuje produkci BDNF, stejně jako podjednotku NR2B NMDA receptoru.[103] Cvičení zvyšuje absorpci IGF-1 z krve do různých oblastí mozku, včetně hipokampu. Kromě toho IGF-1 mění expresi c-fos v hipokampu. Když je IGF-1 blokovaný, cvičení již neindukuje neurogenezi.[105] Další výzkum ukázal, že cvičení myší, které neprodukovaly beta-endorfin, hormon zvyšující náladu, nemělo v neurogenezi žádnou změnu. Přesto myši, které tento hormon produkovaly, spolu s cvičením vykazovaly nárůst novorozených buněk a jejich míru přežití.[106]I když souvislost mezi cvičením zprostředkovanou neurogenezí a zlepšením učení zůstává nejasná, tato studie by mohla mít silné důsledky v oblastech stárnutí a / nebo Alzheimerova choroba.
Účinky kanabinoidů
Tato sekce potřebuje víc lékařské odkazy pro ověření nebo se příliš spoléhá na primární zdroje. (Listopad 2016) |
Některé studie ukázaly, že stimulace kanabinoidy vede k růstu nových nervových buněk v hipokampu z embryonálních i dospělých kmenových buněk. V roce 2005 klinická studie potkanů na univerzitě v Saskatchewanu prokázala regeneraci nervových buněk v hipokampu.[107] Studie ukázaly, že syntetická droga připomíná THC, hlavní psychoaktivní složka marihuany, poskytuje určitou ochranu proti zánět mozku, což by ve vyšším věku mohlo vést k lepší paměti. To je způsobeno receptory v systému, které mohou také ovlivnit produkci nových neuronů.[108]Studie zaměřená na Rutgers University nicméně prokázala, jak se po podání THC změnila synchronizace akčních potenciálů v hipokampu potkanů. Nedostatek synchronizace odpovídal zhoršenému výkonu ve standardním testu paměti.[109]Nedávné studie naznačují, že přírodní kanabinoid konopí, kanabidiol (CBD), zvyšuje neurogenezi dospělých, aniž by ovlivňoval učení. THC však narušilo učení a nemělo žádný vliv na neurogenezi.[110] Vyšší poměr CBD k THC ve vlasových analýzách uživatelů konopí koreluje s ochranou proti redukci šedé hmoty v pravém hipokampu.[111] Bylo také pozorováno, že CBD zmírňuje deficity ve vzpomínkách na prózu a visuoprostorovou asociativní paměť těch, kteří jsou v současné době pod vlivem konopí,[112][113] což znamená neuroprotektivní účinky proti silné expozici THC. Neurogeneze může hrát roli ve svých neuroprotektivních účincích, je však zapotřebí dalšího výzkumu.
Několik studií uvádí pozitivní souvislost mezi THC a hipokampální neurogenezí.[114][115] Někteří z nich hypotetizují dvoufázový efekt,[114] někteří z nich vyjadřují, že část negativních účinků lze připsat neuroadaptaci v důsledku expozice v určitém období života a že je možné ji zvrátit.[116]
Nařízení
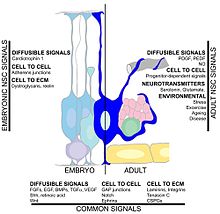
Na rychlost hipokampální neurogeneze může mít vliv mnoho faktorů. Cvičení a obohacené prostředí Bylo prokázáno, že podporují přežití neuronů a úspěšnou integraci novorozených buněk do existujícího hipokampu.[117][118][119][120] Dalším faktorem je centrální nervový systém zranění, protože k neurogenezi dochází po mozková ischemie,[121] epileptické záchvaty,[122] a bakteriální meningitida.[123] Na druhou stranu podmínky jako např chronický stres, virová infekce. a stárnutí can result in a decreased neuronal proliferation.[124][125][126][127]Circulating factors within the blood may reduce neurogenesis. In healthy aging humans, the plasma and cerebrospinal fluid levels of certain chemokines are elevated. In a mouse model, plasma levels of these chemokines correlate with reduced neurogenesis, suggesting that neurogenesis may be modulated by certain global age-dependent systemic changes. These chemokines include CCL11, CCL2 a CCL12, which are highly localized on mouse and human chromosomes, implicating a genetic locus in aging.[54] Another study implicated the cytokine, IL-1beta, which is produced by glia. That study found that blocking IL-1 could partially prevent the severe impairment of neurogenesis caused by a viral infection.[128]
Epigenetická regulace also plays a large role in neurogenesis. DNA methylation is critical in the fate-determination of adult neural stem cells in the subventricular zone for post-natal neurogenesis through the regulation of neuronic genes such as Dlx2, Neurog2, a Sp8. Mnoho mikroRNA such as miR-124 and miR-9 have been shown to influence cortical size and layering during development.[129]
Dějiny
Early neuroanatomists, including Santiago Ramón y Cajal, considered the nervous system fixed and incapable of regeneration. The first evidence of adult mammalian neurogenesis in the mozková kůra byl předložen Joseph Altman v roce 1962,[130] followed by a demonstration of adult neurogenesis in the dentate gyrus of the hippocampus in 1963.[131] In 1969, Joseph Altman discovered and named the rostral migratory stream as the source of adult generated granule cell neurons in the olfactory bulb.[132] Up until the 1980s, the scientific community ignored these findings despite use of the most direct method of demonstrating cell proliferation in the early studies, i.e. 3H-thymidine autoradiography. By that time, Shirley Bayer[133][134] (a Michael Kaplan ) again showed that adult neurogenesis exists in mammals (rats), and Nottebohm showed the same phenomenon in birds[135] sparking renewed interest in the topic. Studies in the 1990s[136][137] finally put research on adult neurogenesis into a mainstream pursuit. Also in the early 1990s hippocampal neurogenesis was demonstrated in non-human primates and humans.[138][139] More recently, neurogenesis in the cerebellum of adult rabbits has also been characterized.[140] Further, some authors (particularly Elizabeth Gould ) have suggested that adult neurogenesis may also occur in regions within the brain not generally associated with neurogenesis including the neokortex.[141][142][143] However, others[144] have questioned the vědecké důkazy of these findings, arguing that the new buňky may be of glial origin. Recent research has elucidated the regulatory effect of GABA on neural stem cells. GABA's well-known inhibitory effects across the brain also affect the local circuitry that triggers a stem cell to become dormant. They found that diazepam (Valium) has a similar effect.[145]
Viz také
Reference
- ^ Faiz M; Acarin L; Castellano B; Gonzalez B (2005). "Proliferation dynamics of germinative zone cells in the intact and excitotoxically lesioned postnatal rat brain". BMC Neuroscience. 6: 26. doi:10.1186/1471-2202-6-26. PMC 1087489. PMID 15826306.
- ^ Oomen CA; Girardi CE; Cahyadi R; et al. (2009). Baune, Bernhard (ed.). "Opposite effects of early maternal deprivation on neurogenesis in male versus female rats". PLOS ONE. 4 (1): e3675. Bibcode:2009PLoSO...4.3675O. doi:10.1371/journal.pone.0003675. PMC 2629844. PMID 19180242.
- ^ Ernst, A; Frisén, J (January 2015). "Adult neurogenesis in humans- common and unique traits in mammals". PLOS Biology. 13 (1): e1002045. doi:10.1371/journal.pbio.1002045. PMC 4306487. PMID 25621867.
- ^ A b Dayer, A.G.; Ford, A.A.; Cleaver, K.M.; Yassaee, M.; Cameron, H.A. (2003). "Short-term and long-term survival of new neurons in the rat dentate gyrus". The Journal of Comparative Neurology. 460 (4): 563–572. doi:10.1002/cne.10675. PMID 12717714. S2CID 7153599.
- ^ Vadodaria, Krishna C.; Gage, Fred H. (2014). "SnapShot: Adult Hippocampal Neurogenesis". Buňka. 156 (5): 1114–1114.e1. doi:10.1016/j.cell.2014.02.029. PMID 24581504.
- ^ Fiorelli, Roberto; Azim, Kasum; Fischer, Bruno; Raineteau, Olivier (2015-06-15). "Adding a spatial dimension to postnatal ventricular-subventricular zone neurogenesis". Rozvoj. 142 (12): 2109–2120. doi:10.1242/dev.119966. ISSN 1477-9129. PMID 26081572.
- ^ Ernst A, Alkass K, Bernard S, Salehpour M, Perl S, Tisdale J, Possnert G, Druid H, Frisén J (2014). "Neurogenesis in the striatum of the adult human brain". Buňka. 156 (5): 1072–83. doi:10.1016/j.cell.2014.01.044. PMID 24561062.
- ^ Bergmann, O.; Liebel, J.; Bernard, S.; Alkass, K.; Yeung, M.S.Y.; Steier, P.; Kutschera, W.; Johnson, L .; Landen, M.; Druid, H.; Spalding, K.L.; Frisen, J. (2012). "The age of olfactory bulb neurons in humans". Neuron. 74 (4): 634–639. doi:10.1016/j.neuron.2012.03.030. PMID 22632721.
- ^ Toni, N.; Teng, E.M.; Bushong, E.A.; Aimone, J.B.; Zhao, C .; Consiglio, A.; van Praag, H.; Martone, M.E.; Ellisman, M.H.; Gage, F.H. (2007). "Synapse formation on neurons born in the adult hippocampus". Přírodní neurovědy. 10 (6): 727–734. doi:10.1038/nn1908. PMID 17486101. S2CID 6796849.
- ^ Vivar, C.; Potter, M.C.; Choi, J .; Lee, J .; Stringer, T.P.; Callawy, E.M.; Gage, F.H.; Suh, H.; van Praag, H. (2012). "Monosynaptic inputs to new neurons in the dentate gyrus". Příroda komunikace. 3 (1038): 1107. Bibcode:2012NatCo...3E1107V. doi:10.1038/ncomms2101. PMC 4603575. PMID 23033083.
- ^ Toni, N.; Laplagne, D.A.; Zhao, C .; Lombardi, G .; Ribak, C.E.; Gage, F.H.; Schinder, A.F. (2008). "Neurons born in the adult dentate gyrus form functional synapses with target cells". Přírodní neurovědy. 11 (8): 901–907. doi:10.1038/nn.2156. PMC 2572641. PMID 18622400.
- ^ Spalding, Kirsty L.; Bergmann, Olaf; Alkass, Kanar; Bernard, Samuel; Salehpour, Mehran; Huttner, Hagen B.; Boström, Emil; Westerlund, Isabelle; Vial, Céline (2013-06-06). "Dynamics of hippocampal neurogenesis in adult humans". Buňka. 153 (6): 1219–1227. doi:10.1016/j.cell.2013.05.002. ISSN 1097-4172. PMC 4394608. PMID 23746839.
- ^ Sorrells, SF; Paredes, MF; Cebrian-Silla, A; Sandoval, K; Qi, D; Kelley, KW; James, D; Mayer, S; Chang, J; Auguste, KI; Chang, EF; Gutierrez, AJ; Kriegstein, AR; Mathern, GW; Oldham, MC; Huang, EJ; Garcia-Verdugo, JM; Yang, Z; Alvarez-Buylla, A (15 March 2018). "Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults". Příroda. 555 (7696): 377–381. Bibcode:2018Natur.555..377S. doi:10.1038/nature25975. PMC 6179355. PMID 29513649.
- ^ A b C d E F G h Cameron, Heather A.; Glover, Lucas R. (2015-01-03). "Adult Neurogenesis: Beyond Learning and Memory". Roční přehled psychologie. 66 (1): 53–81. doi:10.1146/annurev-psych-010814-015006. ISSN 0066-4308. PMC 5612417. PMID 25251485.
- ^ Farahani, Ramin M.; Rezaei‐Lotfi, Saba; Simonian, Mary; Xaymardan, Munira; Hunter, Neil (2019). "Neural microvascular pericytes contribute to human adult neurogenesis". Journal of Comparative Neurology. 527 (4): 780–796. doi:10.1002/cne.24565. ISSN 1096-9861. PMID 30471080. S2CID 53711787.
- ^ Carlson, B. M. (December 1975). "The effects of rotation and positional change of stump tissues upon morphogenesis of the regenerating axolotl limb". Vývojová biologie. 47 (2): 269–291. doi:10.1016/0012-1606(75)90282-1. hdl:1874/15361. ISSN 0012-1606. PMID 1204936.
- ^ Kragl, Martin; Knapp, Dunja; Nacu, Eugen; Khattak, Shahryar; Maden, Malcolm; Epperlein, Hans Henning; Tanaka, Elly M. (2009-07-02). "Cells keep a memory of their tissue origin during axolotl limb regeneration". Příroda. 460 (7251): 60–65. Bibcode:2009Natur.460...60K. doi:10.1038/nature08152. ISSN 1476-4687. PMID 19571878. S2CID 4316677.
- ^ Maden, Malcolm; Manwell, Laurie A.; Ormerod, Brandi K. (2013-01-17). "Proliferation zones in the axolotl brain and regeneration of the telencephalon". Neurální vývoj. 8: 1. doi:10.1186/1749-8104-8-1. ISSN 1749-8104. PMC 3554517. PMID 23327114.
- ^ Clarke, J. D.; Alexander, R.; Holder, N. (1988-06-17). "Regeneration of descending axons in the spinal cord of the axolotl". Neurovědy Dopisy. 89 (1): 1–6. doi:10.1016/0304-3940(88)90471-5. ISSN 0304-3940. PMID 3399135. S2CID 23650500.
- ^ Amamoto, Ryoji; Huerta, Violeta Gisselle Lopez; Takahashi, Emi; Dai, Guangping; Grant, Aaron K; Fu, Zhanyan; Arlotta, Paola (2016). "Adult axolotls can regenerate original neuronal diversity in response to brain injury". eLife. 5. doi:10.7554/eLife.13998. ISSN 2050-084X. PMC 4861602. PMID 27156560.
- ^ Zupanc, G. K. H. (2006-06-01). "Neurogenesis and neuronal regeneration in the adult fish brain". Journal of Comparative Physiology A. 192 (6): 649–670. doi:10.1007/s00359-006-0104-y. ISSN 0340-7594. PMID 16463148. S2CID 24063389.
- ^ Schmidt, Rebecca; Strähle, Uwe; Scholpp, Steffen (2013-02-21). "Neurogenesis in zebrafish – from embryo to adult". Neurální vývoj. 8: 3. doi:10.1186/1749-8104-8-3. ISSN 1749-8104. PMC 3598338. PMID 23433260.
- ^ Hentig, James T.; Byrd-Jacobs, Christine A. (2016-08-31). "Exposure to Zinc Sulfate Results in Differential Effects on Olfactory Sensory Neuron Subtypes in Adult Zebrafish". International Journal of Molecular Sciences. 17 (9): 1445. doi:10.3390/ijms17091445. ISSN 1422-0067. PMC 5037724. PMID 27589738.
- ^ Gorsuch, Ryne A.; Hyde, David R. (June 2014). "Regulation of Müller glial dependent neuronal regeneration in the damaged adult zebrafish retina". Experimental Eye Research. 123: 131–140. doi:10.1016/j.exer.2013.07.012. ISSN 1096-0007. PMC 3877724. PMID 23880528.
- ^ Kroehne, Volker; Freudenreich, Dorian; Hans, Stefan; Kaslin, Jan; Brand, Michael (November 2011). "Regeneration of the adult zebrafish brain from neurogenic radial glia-type progenitors". Rozvoj. 138 (22): 4831–4841. doi:10.1242/dev.072587. ISSN 1477-9129. PMID 22007133.
- ^ Fischer, Andy J. (March 2005). "Neural regeneration in the chick retina". Progress in Retinal and Eye Research. 24 (2): 161–182. doi:10.1016/j.preteyeres.2004.07.003. ISSN 1350-9462. PMID 15610972. S2CID 43652371.
- ^ Jones, Theresa A.; Schallert, Timothy (1992-05-22). "Overgrowth and pruning of dendrites in adult rats recovering from neocortical damage". Výzkum mozku. 581 (1): 156–160. doi:10.1016/0006-8993(92)90356-E. PMID 1498666. S2CID 34248397.
- ^ Di Cosmo, A. D., Bertapelle, C., Porcellini, A., & Polese, G. (2018). Magnitude Assessment of Adult Neurogenesis in the Octopus vulgaris Brain Using a Flow Cytometry-Based Technique. Frontiers in Physiology, 9. doi: 10.3389/fphys.2018.01050
- ^ Di Cosmo, A., and Polese, G. (2014). “Cephalopods meet neuroecology: the role of chemoreception in Octopus vulgaris reproductive behaviour,” in Neuroecology and Neuroethology in Molluscs – The Interface Between Behaviour and Environment, eds A. Di Cosmo and W. Winlow (New York, NY: NOVA Science Publisher), 117–132.
- ^ Polese G, Bertapelle C, Di Cosmo A (January 2015). "Role of olfaction in Octopus vulgaris reproduction". Gen. Comp. Endokrinol. 210: 55–62. doi:10.1016/j.ygcen.2014.10.006. PMID 25449183.
- ^ Amrein I, Lipp HP (February 2009). "Adult hippocampal neurogenesis of mammals: evolution and life history". Biol. Lett. 5 (1): 141–144. doi:10.1098/rsbl.2008.0511. PMC 2657751. PMID 18957357.
- ^ Amrein I, Isler K, Lipp HP (September 2011). "Comparing adult hippocampal neurogenesis in mammalian species and orders: influence of chronological age and life history stage". Eur. J. Neurosci. 34 (6): 978–87. doi:10.1111/j.1460-9568.2011.07804.x. PMID 21929629.
- ^ Hippocampus
- ^ Aimone J. B.; Li Y.; Lee S. W.; Clemenson G. D.; Deng W.; Gage F. H. (2014). "Regulation and Function of Adult Neurogenesis: From Genes to Cognition". Fyziologické recenze. 94 (4): 991–1026. doi:10.1152/physrev.00004.2014. PMC 4280160. PMID 25287858.
- ^ A b Ming Guo-Li, Song Hongjun (2011). "Adult Neurogenesis in the Mammalian Brain: Significant Answers and Significant Questions". Neuron. 70 (4): 687–702. doi:10.1016 / j.neuron.2011.05.001. PMC 3106107. PMID 21609825.
- ^ Altman J (1962). "Are new neurons formed in the brains of adult mammals?". Věda. 135 (3509): 1127–1128. Bibcode:1962Sci...135.1127A. doi:10.1126/science.135.3509.1127. PMID 13860748. S2CID 1606140.
- ^ Kuhn HG, Cooper-Kuhn CM (2007). "Bromodeoxyuridine and the detection of neurogenesis". Curr Pharmaceutical Biotechnol. 8 (3): 127–131. doi:10.2174/138920107780906531. PMID 17584085.
- ^ Imayoshi I, Sakamoto M, Ohtsuka T, Takao K, Miyakawa T, Yamaguchi M, Mori K, Ikeda T, Itohara S, Kageyama R (2008). "Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain". Nat Neurosci. 11 (10): 1153–1161. doi:10.1038/nn.2185. PMID 18758458. S2CID 11627647.
- ^ Prendergast, Franklyn G.; Mann, Kenneth G. (1978-08-22). "Chemical and physical properties of aequorin and the green fluorescent protein isolated from Aequorea forskalea". Biochemie. 17 (17): 3448–3453. doi:10.1021/bi00610a004. ISSN 0006-2960. PMID 28749.
- ^ Gonçalves, J. Tiago; Schafer, Simon T.; Gage, Fred H. (2016). "Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior". Buňka. 167 (4): 897–914. doi:10.1016/j.cell.2016.10.021. PMID 27814520.
- ^ Zhao C, Teng EM, Summers RG Jr, Ming GL, Gage FH (2006). "Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus". J Neurosci. 26 (1): 3–11. doi:10.1523/jneurosci.3648-05.2006. PMC 6674324. PMID 16399667.
- ^ Shors, Tracey J.; Miesegaes, George; Beylin, Anna; Zhao, Mingrui; Rydel, Tracy; Gould, Elizabeth (2001). "Neurogenesis in the adult is involved in the formation of trace memories". Příroda. 410 (6826): 372–376. doi:10.1038/35066584. PMID 11268214. S2CID 4430850.
- ^ Denny, Christine A .; Burghardt, Nesha S.; Schachter, Daniel M.; Hen, René; Drew, Michael R. (2012-05-01). "4- to 6-week-old adult-born hippocampal neurons influence novelty-evoked exploration and contextual fear conditioning". Hippocampus. 22 (5): 1188–1201. doi:10.1002/hipo.20964. ISSN 1098-1063. PMC 3193906. PMID 21739523.
- ^ Plack, C. J.; Oxenham, A. J.; Drga, V. (2006). "The Journal of Neurosci". The Journal of Neuroscience. 26 (34): 8767–8773. doi:10.1523/JNEUROSCI.1134-06.2006. PMC 1808348. PMID 16928865.
- ^ Drew, Michael R.; Denny, Christine A .; Hen, Rene (2010). "Arrest of adult hippocampal neurogenesis in mice impairs single- but not multiple-trial contextual fear conditioning". Behavioral Neuroscience. 124 (4): 446–454. doi:10.1037/a0020081. PMC 2925248. PMID 20695644.
- ^ Kesner, Raymond P. (2013-10-01). "An analysis of the dentate gyrus function". Behaviorální výzkum mozku. SI:Medial Temporal Lobe Memory Networks. 254: 1–7. doi:10.1016/j.bbr.2013.01.012. PMID 23348108. S2CID 5486456.
- ^ França, Thiago F.A.; Bitencourt, Alexandre M.; Maximilla, Naiana R.; Barros, Daniela M.; Monserrat, Jose M. (2017-07-20). "Hippocampal neurogenesis and pattern separation: a meta-analysis of behavioral data". Hippocampus. 9. 27 (9): 937–950. doi:10.1002/hipo.22746. PMID 28597491. S2CID 205914396.
- ^ Sahay, Amar; Scobie, Kimberly N.; Hill, Alexis S.; O'Carroll, Colin M.; Kheirbek, Mazen A .; Burghardt, Nesha S.; Fenton, André A.; Dranovsky, Alex; Hen, René (2011). "Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation". Příroda. 472 (7344): 466–470. Bibcode:2011Natur.472..466S. doi:10.1038/nature09817. PMC 3084370. PMID 21460835.
- ^ Clelland, C. D.; Choi, M.; Romberg, C.; Clemenson, G. D.; Fragniere, A.; Tyers, P.; Jessberger, S.; Saksida, L. M.; Barker, R. A. (2009-07-10). "A functional role for adult hippocampal neurogenesis in spatial pattern separation". Věda. 325 (5937): 210–213. Bibcode:2009Sci...325..210C. doi:10.1126/science.1173215. ISSN 1095-9203. PMC 2997634. PMID 19590004.
- ^ França, Thiago F.A.; Bitencourt, Alexandre M.; Maximilla, Naiana R.; Barros, Daniela M.; Monserrat, Jose M. (2017-07-20). "Hippocampal neurogenesis and pattern separation: a meta-analysis of behavioral data". Hippocampus. 9. 27 (9): 937–950. doi:10.1002/hipo.22746. PMID 28597491. S2CID 205914396.
- ^ Kheirbek, Mazen A .; Drew, Liam J.; Burghardt, Nesha S.; Costantini, Daniel O.; Tannenholz, Lindsay; Ahmari, Susanne E.; Zeng, Hongkui; Fenton, André A.; Hen, René (2013-03-06). "Differential control of learning and anxiety along the dorsoventral axis of the dentate gyrus". Neuron. 77 (5): 955–968. doi:10.1016/j.neuron.2012.12.038. ISSN 1097-4199. PMC 3595120. PMID 23473324.
- ^ Gould E, Cameron HA (1997). "Early NMDA receptor blockade impairs defensive behavior and increases cell proliferation in the dentate gyrus of developing rats". Chovat se. Neurosci. 111 (1): 49–56. doi:10.1037/0735-7044.111.1.49. PMID 9109623.
- ^ Kempermann G, Wiskott L, Gage FH (April 2004). "Functional significance of adult neurogenesis". Aktuální názor v neurobiologii. 14 (2): 186–91. doi:10.1016/j.conb.2004.03.001. PMID 15082323. S2CID 6241827.
- ^ A b G. Neves, G; S.F. Cooke; T.V. Bliss (2008). "Synaptic plasticity, memory and the hippocampus: A neural network approach to causality". Nature Reviews Neuroscience. 9 (1): 65–75. doi:10.1038/nrn2303. PMID 18094707. S2CID 33104731.
- ^ Becker S (2005). "A computational principle for hippocampal learning and neurogenesis". Hippocampus. 15 (6): 722–38. doi:10.1002/hipo.20095. PMID 15986407.
- ^ Wiskott L, Rasch MJ, Kempermann G (2006). "A functional hypothesis for adult hippocampal neurogenesis: avoidance of catastrophic interference in the dentate gyrus". Hippocampus. 16 (3): 329–43. CiteSeerX 10.1.1.408.8944. doi:10.1002/hipo.20167. PMID 16435309.
- ^ Aimone JB, Wiles J, Gage FH (June 2006). "Potential role for adult neurogenesis in the encoding of time in new memories". Nat. Neurosci. 9 (6): 723–7. doi:10.1038/nn1707. PMID 16732202. S2CID 10586519.
- ^ Kovács KA (září 2020). „Epizodické vzpomínky: Jak spolupracují hipokampus a přitažliví jezdci na okruhu?“. Frontiers in Systems Neuroscience. 14: 68. doi:10.3389 / fnsys.2020.559186.
- ^ Shors TJ, Townsend DA, Zhao M, Kozorovitskiy Y, Gould E (2002). "Neurogenesis may relate to some but not all types of hippocampal-dependent learning". Hippocampus. 12 (5): 578–84. doi:10.1002/hipo.10103. PMC 3289536. PMID 12440573.
- ^ Meshi D, Drew MR, Saxe M, et al. (Červen 2006). "Hippocampal neurogenesis is not required for behavioral effects of environmental enrichment". Nat. Neurosci. 9 (6): 729–31. doi:10.1038/nn1696. PMID 16648847. S2CID 11043203.
- ^ Gould, E .; Beylin, A.; Tanapat, P.; Reeves, A.; Shors, T. J. (1999). "Learning enhances adult neurogenesis in the hippocampal formation". Přírodní neurovědy. 2 (3): 260–265. doi:10.1038/6365. PMID 10195219. S2CID 12112156.
- ^ Donovan, M. H.; Yazdani, U; Norris, R. D.; Games, D; German, D. C.; Eisch, A. J. (2006). "Decreased adult hippocampal neurogenesis in the PDAPP mouse model of Alzheimer's disease". The Journal of Comparative Neurology. 495 (1): 70–83. doi:10.1002/cne.20840. PMID 16432899. S2CID 20559097.
- ^ Jin, K; Peel, A. L.; Mao, X. O.; Xie, L; Cottrell, B. A.; Henshall, D. C.; Greenberg, D. A. (2004). "Increased hippocampal neurogenesis in Alzheimer's disease". Sborník Národní akademie věd. 101 (1): 343–7. Bibcode:2004PNAS..101..343J. doi:10.1073/pnas.2634794100. PMC 314187. PMID 14660786.
- ^ Foster, P. P.; Rosenblatt, K. P.; Kuljiš, R. O. (2011). "Exercise-induced cognitive plasticity, implications for mild cognitive impairment and Alzheimer's disease". Hranice v neurologii. 2: 28. doi:10.3389/fneur.2011.00028. PMC 3092070. PMID 21602910.
- ^ Marx CE, Trost WT, Shampine LJ, et al. (Prosinec 2006). "The neurosteroid allopregnanolone is reduced in prefrontal cortex in Alzheimer's disease". Biol. Psychiatrie. 60 (12): 1287–94. doi:10.1016/j.biopsych.2006.06.017. PMID 16997284. S2CID 27813000.
- ^ Wang JM, Singh C, Liu L, Irwin RW, Chen S, Chung EJ, Thompson RF, Brinton RD (2010). "Allopregnanolone reverses neuron and cognitive deficits in a mouse model of Alzheimer's disease". Proc. Natl. Acad. Sci. USA. 107 (14): 6498–6503. Bibcode:2010PNAS..107.6498W. doi:10.1073/pnas.1001422107. PMC 2851948. PMID 20231471.
- ^ Cissé, M; Checler, F (2014). "Eph receptors: New players in Alzheimer's disease pathogenesis". Neurobiologie nemocí. 73C: 137–149. doi:10.1016/j.nbd.2014.08.028. PMID 25193466. S2CID 28922292.
- ^ Mu, Y; Gage, F. H. (2011). "Adult hippocampal neurogenesis and its role in Alzheimer's disease". Molekulární neurodegenerace. 6: 85. doi:10.1186/1750-1326-6-85. PMC 3261815. PMID 22192775.
- ^ LeStrat, Y (May 2009). "The role of genes involved in neuroplasticity and neurogenesis in the observation of a gene-environment interaction (GxE) in schizophrenia". Současná molekulární medicína. 9 (4): 506–18. doi:10.2174/156652409788167104. PMID 19519407.
- ^ Schreiber, Rudy; Newman-Tancredi, Adrian (April 2014). "Improving cognition in schizophrenia with antipsychotics that elicit neurogenesis through 5-HT1A receptor activation". Neurobiologie učení a paměti. 110: 72–80. doi:10.1016/j.nlm.2013.12.015. PMID 24423786. S2CID 28027825.
- ^ Reif, Andreas; Schmitt, Angelika; Fritzen, Sabrina; Lesch, Klaus-Peter (27 Apr 2007). "Neurogenesis and schizophrenia: dividing neurons in a divided mind?". European Archives of Psychiatry and Clinical Neuroscience. 257 (5): 290–299. doi:10.1007/s00406-007-0733-3. PMID 17468935. S2CID 28750292.
- ^ Numakawa, Tadahiro; Odaka, Haruki; Adachi, Naoki (2017). "Impact of glucocorticoid on neurogenesis". Výzkum regenerace nervů. 12 (7): 1028–1035. doi:10.4103/1673-5374.211174. PMC 5558474. PMID 28852377.
- ^ A b C Jacobs, B. L.; H. van Praag; F. H. Gage (2000). "Depression and the Birth and Death of Brain Cells". Americký vědec. 88 (4): 340. Bibcode:2000AmSci..88..340J. doi:10.1511/2000.4.340.
- ^ Kandel, E. R .; J. H. Schwartz & T. M. Jessell (2012-10-26). Principles of Neural Science (páté vydání). ISBN 978-0071390118.
- ^ National Institute of Mental Health (2010). "Mental Health Medications" (PDF).
- ^ Davies-Sala, M. G.; Espósito, M. S.; Piatti, V. C.; Mongiat, L. A.; Trinchero M. F. & Schinder A. F. (25 May 2011). "The Timing for Neuronal Maturation in the Adult Hippocampus Is Modulated by Local Network Activity". The Journal of Neuroscience. 31 (21): 7715–28. doi:10.1523/JNEUROSCI.1380-11.2011. PMC 3701257. PMID 21613484.
- ^ Perera TD, Dwork AJ, Keegan KA, Thirumangalakudi L, Lipira CM, Joyce N, Lange C, Higley JD, Rosoklija G, Hen R, Sackeim HA, Coplan JD (2011). "Necessity of hippocampal neurogenesis for the therapeutic action of antidepressants in adult nonhuman primates". PLOS ONE. 6 (4): e17600. Bibcode:2011PLoSO...617600P. doi:10.1371/journal.pone.0017600. PMC 3078107. PMID 21525974.
- ^ A b Schloesser RJ, Manji HK, Martinowich K (April 2009). "Suppression of adult neurogenesis leads to an increased hypothalamo-pituitary-adrenal axis response". NeuroReport. 20 (6): 553–7. doi:10.1097/WNR.0b013e3283293e59. PMC 2693911. PMID 19322118.
- ^ A b Surget A, Tanti A, Leonardo ED, et al. (Prosinec 2011). "Antidepressants recruit new neurons to improve stress response regulation". Molekulární psychiatrie. 16 (12): 1177–88. doi:10.1038/mp.2011.48. PMC 3223314. PMID 21537331.
- ^ Malberg JE, Eisch AJ, Nestler EJ, Duman RS (Prosinec 2000). „Chronická antidepresivní léčba zvyšuje neurogenezi v hipokampu dospělých potkanů“. J. Neurosci. 20 (24): 9104–10. doi:10.1523 / JNEUROSCI.20-24-09104.2000. PMC 6773038. PMID 11124987.
- ^ Manev H, Uz T, Smalheiser NR, Manev R (January 2001). "Antidepressants alter cell proliferation in the adult brain in vivo and in neural cultures in vitro". Eur J Pharmacol. 411 (1–2): 67–70. doi:10.1016/S0014-2999(00)00904-3. PMID 11137860.
- ^ Santarelli L, Saxe M, Gross C, et al. (Srpen 2003). "Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants". Věda. 301 (5634): 805–9. Bibcode:2003Sci...301..805S. doi:10.1126/science.1083328. PMID 12907793. S2CID 9699898.
- ^ Bradley, Joseph (2015). Addiction: From Suffering to Solution. Las Vegas, NV: Breaux Press International. p. 173. ISBN 978-0-9854418-0-7.
- ^ Castrén E (March 2005). "Is mood chemistry?". Nature Reviews Neuroscience. 6 (3): 241–6. doi:10.1038/nrn1629. PMID 15738959. S2CID 34523310.
- ^ Vetencourt, J. F. M.; Sale, A.; Viegi, A.; Baroncelli, L.; De Pasquale, R.; F. o'Leary, O.; Castren, E.; Maffei, L. (2008). "The Antidepressant Fluoxetine Restores Plasticity in the Adult Visual Cortex". Věda. 320 (5874): 385–8. Bibcode:2008Sci...320..385M. doi:10.1126/science.1150516. PMID 18420937. S2CID 39565465.
- ^ Mirescu C, Peters JD, Noiman L, Gould E (December 2006). "Sleep deprivation inhibits adult neurogenesis in the hippocampus by elevating glucocorticoids". Proc. Natl. Acad. Sci. USA. 103 (50): 19170–5. Bibcode:2006PNAS..10319170M. doi:10.1073/pnas.0608644103. PMC 1748194. PMID 17135354.
- ^ Mirescu C.; Peters J. D.; Noiman L.; Gould E. (2006). "Sleep deprivation inhibits adult neurogenesis in the hippocampus by elevating glucocorticoids". Sborník Národní akademie věd. 103 (50): 19170–19175. Bibcode:2006PNAS..10319170M. doi:10.1073/pnas.0608644103. PMC 1748194. PMID 17135354.
- ^ A b C Arias-Carrión O, Freundlieb N, Oertel WH, Höglinger GU (October 2007). "Adult neurogenesis and Parkinson's disease". Cíle léků na neurol disord CNS. 6 (5): 326–35. doi:10.2174/187152707783220875. PMID 18045161. Archivovány od originál on 2013-04-14.
- ^ Fallon J, Reid S, Kinyamu R, et al. (Prosinec 2000). "In vivo induction of massive proliferation, directed migration, and differentiation of neural cells in the adult mammalian brain". Proc. Natl. Acad. Sci. USA. 97 (26): 14686–91. Bibcode:2000PNAS...9714686F. doi:10.1073/pnas.97.26.14686. PMC 18979. PMID 11121069.
- ^ Arias-Carrión O, Verdugo-Díaz L, Feria-Velasco A, et al. (Říjen 2004). "Neurogenesis in the subventricular zone following transcranial magnetic field stimulation and nigrostriatal lesions". J Neurosci Res. 78 (1): 16–28. doi:10.1002/jnr.20235. PMID 15372495.
- ^ Arias-Carrión O, Hernández-López S, Ibañez-Sandoval O, Bargas J, Hernández-Cruz A, Drucker-Colín R (November 2006). "Neuronal precursors within the adult rat subventricular zone differentiate into dopaminergic neurons after substantia nigra lesion and chromaffin cell transplant". J Neurosci Res. 84 (7): 1425–37. doi:10.1002/jnr.21068. PMID 17006899.
- ^ A b Höglinger GU, Rizk P, Muriel MP, et al. (Červenec 2004). "Dopamine depletion impairs precursor cell proliferation in Parkinson disease". Nat. Neurosci. 7 (7): 726–35. doi:10.1038/nn1265. PMID 15195095. S2CID 952173.
- ^ Neurogenesis in the Striatum of the Adult Human Brain
- ^ Guo, Qingmin; Sayeed, Iqbal; Baronne, Lon M.; Hoffman, Stuart W.; Guennoun, Rachida; Stein, Donald G. (April 2006). "Progesterone administration modulates AQP4 expression and edema after traumatic brain injury in male rats". Experimentální neurologie. 198 (2): 469–478. doi:10.1016/j.expneurol.2005.12.013. ISSN 0014-4886. PMID 16445913. S2CID 26143265.
- ^ Petrone, Ashley B.; Gatson, Joshua W.; Simpkins, James W.; Reed, Miranda N. (2014-05-25). "Non-feminizing estrogens: a novel neuroprotective therapy". Molekulární a buněčná endokrinologie. 389 (1–2): 40–47. doi:10.1016/j.mce.2013.12.017. ISSN 1872-8057. PMC 4040321. PMID 24424441.
- ^ Han, S.; Zhao, B .; Pan, X .; Song, Z.; Liu, J .; Gong, Y.; Wang, M. (2015-12-03). "Estrogen receptor variant ER-α36 is involved in estrogen neuroprotection against oxidative toxicity". Neurovědy. 310: 224–241. doi:10.1016/j.neuroscience.2015.09.024. ISSN 1873-7544. PMID 26383254. S2CID 44289364.
- ^ Singh, Surjit; Hota, Debasish; Prakash, Ajay; Khanduja, Krishan L.; Arora, Sunil K.; Chakrabarti, Amitava (January 2010). "Allopregnanolone, the active metabolite of progesterone protects against neuronal damage in picrotoxin-induced seizure model in mice". Farmakologická biochemie a chování. 94 (3): 416–422. doi:10.1016/j.pbb.2009.10.003. ISSN 1873-5177. PMID 19840816. S2CID 45591868.
- ^ Webster, Mark K.; Cooley-Themm, Cynthia A.; Barnett, Joseph D.; Bach, Harrison B.; Vainner, Jessica M.; Webster, Sarah E.; Linn, Cindy L. (2017-03-27). "Evidence of BrdU-positive retinal neurons after application of an Alpha7 nicotinic acetylcholine receptor agonist". Neurovědy. 346: 437–446. doi:10.1016/j.neuroscience.2017.01.029. ISSN 1873-7544. PMC 5341387. PMID 28147247.
- ^ Drapeau, E.; Mayo, W.; Aurousseau, C.; Moal, M.L.; Piazza, P. & Abrous, D.N. (2003). "Spatial memory performances of aged rats in the water maze predict levels of hippocampal neurogenesis". PNAS. 100 (24): 14385–14390. Bibcode:2003PNAS..10014385D. doi:10.1073/pnas.2334169100. PMC 283601. PMID 14614143.
- ^ Marrone, D.F.; Ramirez-Amaya, V. & Barnes, C.A. (2012). "Neurons generated in senescence maintain capacity for functional integration". Hippocampus. 22 (5): 1134–1142. doi:10.1002/hipo.20959. PMC 3367380. PMID 21695743.
- ^ von Bohlen und Halbach O (2010). "Involvement of BDNF in age-dependent alterations in the hippocampus". Front Aging Neurosci. 2. doi:10.3389/fnagi.2010.00036. PMC 2952461. PMID 20941325.
- ^ Praag H, Christie BR, Sejnowski TJ, Gage FH (1999). "Running enhances neurogenesis, learning, and long-term potentiation in mice". Proc Natl Acad Sci U S A. 96 (23): 13427–31. Bibcode:1999PNAS...9613427V. doi:10.1073/pnas.96.23.13427. PMC 23964. PMID 10557337.
- ^ A b C Farmer J, Zhao X, van Praag H, Wodtke K, Gage FH, Christie BR (2004). "Effects of voluntary exercise on synaptic plasticity and gene expression in the dentate gyrus of adult male Sprague-Dawley rats in vivo". Neurovědy. 124 (1): 71–9. doi:10.1016/j.neuroscience.2003.09.029. PMID 14960340. S2CID 2718669.
- ^ van Praag H; Christie BR; Sejnowski TJ; Gage FH (Nov 1999). "Running enhances neurogenesis, learning, and long-term potentiation in mice". Proc Natl Acad Sci U S A. 96 (23): 13427–31. Bibcode:1999PNAS...9613427V. doi:10.1073/pnas.96.23.13427. PMC 23964. PMID 10557337.
- ^ A b Carro, E; Trejo, J. L.; Busiguina, S; Torres-Aleman, I (2001). "Circulating insulin-like growth factor I mediates the protective effects of physical exercise against brain insults of different etiology and anatomy". The Journal of Neuroscience. 21 (15): 5678–84. doi:10.1523/JNEUROSCI.21-15-05678.2001. PMC 6762673. PMID 11466439.
- ^ "Adult neurogenesis". Brain briefingy. Společnost pro neurovědy. Červen 2007. Citováno 2011-11-26.
- ^ Wen Jiang; Yun Zhang; Lan Xiao; Jamie Van Cleemput; Shao-Ping Ji; Guang Bai; Xia Zhang (2005-11-01). „Kanabinoidy podporují embryogenní a dospělou hipokampickou neurogenezi a vyvolávají anxiolytické a antidepresivní účinky“. Journal of Clinical Investigation. 115 (11): 3104–16. doi:10,1172 / JCI25509. PMC 1253627. PMID 16224541. Citováno 2011-03-02.
- ^ 11-19-2008 Ohio State study: Scientists are high on idea that marijuana reduces memory impairment. OSU.edu
- ^ 29. listopadu 2006. Study: Marijuana may affect neuron firing. United Press International
- ^ Wolf, Susanne A.; Bick-Sander, Anika; Fabel, Klaus; Leal-Galicia, Perla; Tauber, Svantje; Ramirez-Rodriguez, Gerardo; Müller, Anke; Melnik, Andre; Waltinger, Tim P.; Ullrich, Oliver; Kempermann, Gerd (2010). "Cannabinoid receptor CB1 mediates baseline and activity-induced survival of new neurons in adult hippocampal neurogenesis". Cell Communication and Signaling. 8 (1): 12. doi:10.1186/1478-811X-8-12. PMC 2898685. PMID 20565726.
- ^ Demirakca, T.; Sartorius, A.; Ende, G.; Meyer, N.; Welzel, H.; Skopp, G.; Mann, K.; Hermann, D. (2010). "Diminished gray matter in the hippocampus of cannabis users: Possible protective effects of cannabidiol". Závislost na drogách a alkoholu. 114 (2–3): 242–245. doi:10.1016/j.drugalcdep.2010.09.020. PMID 21050680.
- ^ Wright, M. J.; Vandewater, S. A.; Taffe, M. A. (2013). "Cannabidiol attenuates deficits of visuospatial associative memory induced by Δ9tetrahydrocannabinol". British Journal of Pharmacology. 170 (7): 1365–1373. doi:10.1111/bph.12199. PMC 3838683. PMID 23550724.
- ^ Morgan, C. J. A.; Schafer, G.; Freeman, T. P.; Curran, H. V. (2010). "Impact of cannabidiol on the acute memory and psychotomimetic effects of smoked cannabis: Naturalistic study". British Journal of Psychiatry. 197 (4): 285–290. doi:10.1192/bjp.bp.110.077503. PMID 20884951.
- ^ A b Calabrese, Edward J.; Rubio-Casillas, Alberto (May 2018). "Biphasic effects of THC in memory and cognition". European Journal of Clinical Investigation. 48 (5): e12920. doi:10.1111/eci.12920. PMID 29574698.
- ^ Suliman, Noor Azuin; Taib, Che Norma Mat; Moklas, Mohamad Aris Mohd; Basir, Rusliza (21 September 2017). "Delta-9-Tetrahydrocannabinol (∆9-THC) Induce Neurogenesis and Improve Cognitive Performances of Male Sprague Dawley Rats". Výzkum neurotoxicity. 33 (2): 402–411. doi:10.1007/s12640-017-9806-x. PMC 5766723. PMID 28933048.
- ^ Cuccurazzu, Bruna; Zamberletti, Erica; Nazzaro, Cristiano; Prini, Pamela; Trusel, Massimo; Grilli, Mariagrazia; Parolaro, Daniela; Tonini, Raffaella; Rubino, Tiziana (November 2018). "Adult Cellular Neuroadaptations Induced by Adolescent THC Exposure in Female Rats Are Rescued by Enhancing Anandamide Signaling". International Journal of Neuropsychopharmacology. 21 (11): 1014–1024. doi:10.1093/ijnp/pyy057. PMC 6209859. PMID 29982505.
- ^ Lazarov O, Robinson J, Tang YP, Hairston IS, Korade-Mirnics Z, Lee VM, Hersh LB, Sapolsky RM, Mirnics K, Sisodia SS (March 2005). "Environmental enrichment reduces Abeta levels and amyloid deposition in transgenic mice". Buňka. 120 (5): 701–13. doi:10.1016/j.cell.2005.01.015. PMID 15766532. S2CID 16756661.
- ^ Van Praag, H.; Shubert, T.; Zhao, C .; Gage, F. (2005). "Exercise enhances learning and hippocampal neurogenesis in aged mice". Journal of Neuroscience. 25 (38): 8680–8685. doi:10.1523/JNEUROSCI.1731-05.2005. PMC 1360197. PMID 16177036.
- ^ Van Praag, H.; Kempermann, G .; Gage, F. (1999). "Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus". Přírodní neurovědy. 2 (3): 266–270. doi:10.1038/6368. PMID 10195220. S2CID 7170664.
- ^ Bjørnebekk A, Mathé AA, Brené S (September 2005). "The antidepressant effect of running is associated with increased hippocampal cell proliferation". Int J Neuropsychopharmacol. 8 (3): 357–68. doi:10.1017/S1461145705005122. PMID 15769301.
- ^ Jin K, Wang X, Xie L, et al. (Srpen 2006). "Evidence for stroke-induced neurogenesis in the human brain". Proc. Natl. Acad. Sci. USA. 103 (35): 13198–202. Bibcode:2006PNAS..10313198J. doi:10.1073/pnas.0603512103. PMC 1559776. PMID 16924107.
- ^ Parent JM; Elliott, RC; Pleasure, SJ; Barbaro, NM; Lowenstein, DH (2006). "Aberrant seizure-induced neurogenesis in experimental temporal lobe epilepsy". Annals of Neurology. 59 (1): 81–91. doi:10.1002/ana.20699. hdl:2027.42/49280. PMID 16261566. S2CID 9655613.
- ^ Gerber J, Tauber SC, Armbrecht I, Schmidt H, Brück W, Nau R (2009). "Increased neuronal proliferation in human bacterial meningitis". Neurologie. 73 (13): 1026–32. doi:10.1212/WNL.0b013e3181b9c892. PMID 19786694. S2CID 26108905.
- ^ Sharma A, Valadi N, Miller AH, Pearce BD (2002). "Adult neurogenesis in the hippocampus is impaired following neonatal viral infection". Neurobiologie nemocí. 11 (2): 246–56. doi:10.1006/nbdi.2002.0531. PMID 12505418. S2CID 6405590.
- ^ Lee AL, Ogle WO, Sapolsky RM (April 2002). "Stress and depression: possible links to neuron death in the hippocampus". Bipolar Disord. 4 (2): 117–28. doi:10.1034/j.1399-5618.2002.01144.x. PMID 12071509.
- ^ Sheline YI, Gado MH, Kraemer HC (August 2003). "Untreated depression and hippocampal volume loss". Am J Psychiatry. 160 (8): 1516–8. doi:10.1176/appi.ajp.160.8.1516. PMID 12900317.
- ^ Jiang, W .; et al. (2005). „Kanabinoidy podporují embryogenní a dospělou hipokampickou neurogenezi a vyvolávají anxiolytické a antidepresivní účinky“. The Journal of Clinical Investigation. 115 (11): 3104–3116. doi:10,1172 / JCI25509. PMC 1253627. PMID 16224541.
- ^ Orr, Anna G.; Sharma, Anup; Binder, Nikolaus B.; Miller, Andrew H .; Pearce, Bradley D. (2010). "Interleukin-1 Mediates Long-Term Hippocampal Dentate Granule Cell Loss Following Postnatal Viral Infection". Journal of Molecular Neuroscience. 41 (1): 89–96. doi:10.1007/s12031-009-9293-5. PMID 19774496. S2CID 2427299.
- ^ Hu, X.L.; Wang, Y .; Shen, Q. (2012). "Epigenetic control on cell fate choice in neural stem cells". Protein & Cell. 3 (4): 278–290. doi:10.1007/s13238-012-2916-6. PMC 4729703. PMID 22549586.
- ^ Altman, J. (1962). "Are new neurons formed in the brains of adult mammals?". Věda. 135 (3509): 1127–1128. Bibcode:1962Sci...135.1127A. doi:10.1126/science.135.3509.1127. PMID 13860748. S2CID 1606140.
- ^ Altman, J. (1963). "Autoradiographic investigation of cell proliferation in the brains of rats and cats". Anatomický záznam. 145 (4): 573–591. doi:10.1002/ar.1091450409. PMID 14012334. S2CID 5340726.
- ^ Altman, J. (1969). "Autoradiographic and histological studies of postnatal neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb". The Journal of Comparative Neurology. 137 (4): 433–457. doi:10.1002/cne.901370404. PMID 5361244. S2CID 46728071.
- ^ Bayer, S. A.; Yackel, J. W.; Puri, P. S. (1982). "Neurons in the rat dentate gyrus granular layer substantially increase during juvenile and adult life". Věda. 216 (4548): 890–892. Bibcode:1982Sci...216..890B. doi:10.1126/science.7079742. PMID 7079742.
- ^ Bayer, S. A. (1982). "Changes in the total number of dentate granule cells in juvenile and adult rats: a correlated volumetric and 3H-thymidine autoradiographic study". Experimental Brain Research. Experimentelle Hirnforschung. Experimentation Cerebrale. 46 (3): 315–323. doi:10.1007/bf00238626. PMID 7095040. S2CID 18663323.
- ^ Goldman SA, Nottebohm F (April 1983). "Neuronal production, migration, and differentiation in a vocal control nucleus of the adult female canary brain". Proc. Natl. Acad. Sci. USA. 80 (8): 2390–4. Bibcode:1983PNAS...80.2390G. doi:10.1073/pnas.80.8.2390. PMC 393826. PMID 6572982.
- ^ Reynolds, B. A.; Weiss, S. (Mar 1992). "Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system". Věda. 255 (5052): 1707–1710. Bibcode:1992Sci...255.1707R. doi:10.1126/science.1553558. ISSN 0036-8075. PMID 1553558.
- ^ Gage, F. H.; Ray, J.; Fisher, L. J. (1995). "Isolation, Characterization, and use of Stem Cells from the CNS". Roční přehled neurovědy. 18: 159–92. doi:10.1146/annurev.ne.18.030195.001111. PMID 7605059.
- ^ Eriksson PS, Perfilieva E, Björk-Eriksson T, et al. (Listopad 1998). "Neurogenesis in the adult human hippocampus". Nat. Med. 4 (11): 1313–7. doi:10.1038/3305. PMID 9809557.
- ^ Gould, E .; Reeves; Fallah; Tanapat; Hrubý; Fuchs (1999). "Hippocampal neurogenesis in adult Old World primates". Sborník Národní akademie věd Spojených států amerických. 96 (9): 5263–5267. Bibcode:1999PNAS...96.5263G. doi:10.1073/pnas.96.9.5263. PMC 21852. PMID 10220454.
- ^ Ponti, G .; Peretto, B.; Bonfanti, L. (2008). Reh, Thomas A. (ed.). "Genesis of neuronal and glial progenitors in the cerebellar cortex of peripuberal and adult rabbits". PLOS ONE. 3 (6): e2366. Bibcode:2008PLoSO...3.2366P. doi:10.1371/journal.pone.0002366. PMC 2396292. PMID 18523645.
- ^ Gould, E .; Reeves, A. J.; Graziano, M. S.; Gross, C. G. (1999). "Neurogenesis in the neocortex of adult primates". Věda. 286 (5439): 548–552. doi:10.1126/science.286.5439.548. PMID 10521353.
- ^ Zhao, M.; Momma, S.; Delfani, K.; Carlen, M.; Cassidy, R. M.; Johansson, C. B.; Brismar, H.; Shupliakov, O.; Frisen, J.; Janson, A. (2003). "Evidence for neurogenesis in the adult mammalian substantia nigra". Sborník Národní akademie věd Spojených států amerických. 100 (13): 7925–7930. Bibcode:2003PNAS..100.7925Z. doi:10.1073/pnas.1131955100. PMC 164689. PMID 12792021.
- ^ Shankle; Rafii, M. S.; Landing, B. H.; Fallon, J. H. (1999). "Approximate doubling of numbers of neurons in postnatal human cerebral cortex and in 35 specific cytoarchitectural areas from birth to 72 months". Dětská a vývojová patologie. 2 (3): 244–259. doi:10.1007/s100249900120. PMID 10191348. S2CID 23546857.
- ^ Rakic P (February 2002). "Adult neurogenesis in mammals: an identity crisis". J. Neurosci. 22 (3): 614–8. doi:10.1523/JNEUROSCI.22-03-00614.2002. PMC 6758501. PMID 11826088.
- ^ Song, Juan; Zhong, Chun; Bonaguidi, Michael A.; Sun, Gerald J.; Hsu, Derek; Gu, Yan; Meletis, Konstantinos; Huang, Z. Josh; Ge, Shaoyu; Enikolopov, Grigori; Deisseroth, Karl; Luscher, Bernhard; Christian, Kimberly M.; Ming, Guo-li; Song, Hongjun (2012). "Neuronal circuitry mechanism regulating adult quiescent neural stem-cell fate decision". Příroda. 489 (7414): 150–154. Bibcode:2012Natur.489..150S. doi:10.1038/nature11306. PMC 3438284. PMID 22842902.
- Poznámky
- Aimone JB, Jessberger S, and Gage FH (2007) Adult Neurogenesis. Scholarpedia, p. 8739
- Gould E, Reeves AJ, Fallah M, Tanapat P, Gross CG, Fuchs E (April 1999). "Hippocampal neurogenesis in adult Old World primates". Proc. Natl. Acad. Sci. USA. 96 (9): 5263–7. Bibcode:1999PNAS...96.5263G. doi:10.1073/pnas.96.9.5263. PMC 21852. PMID 10220454.
- Gould E, Reeves AJ, Graziano MS, Gross CG (October 1999). "Neurogenesis in the neocortex of adult primates". Věda. 286 (5439): 548–52. doi:10.1126/science.286.5439.548. PMID 10521353.
- Gould E, Beylin A, Tanapat P, Reeves A, Shors TJ (March 1999). "Learning enhances adult neurogenesis in the hippocampal formation". Nat. Neurosci. 2 (3): 260–5. doi:10.1038/6365. PMID 10195219. S2CID 12112156.
- Moghadam KS, Chen A, Heathcote RD (August 2003). "Establishment of a ventral cell fate in the spinal cord". Dev. Dyn. 227 (4): 552–62. doi:10.1002/dvdy.10340. PMID 12889064.
- Rakic P (January 2002). "Neurogenesis in adult primate neocortex: an evaluation of the evidence". Nature Reviews Neuroscience. 3 (1): 65–71. doi:10.1038/nrn700. PMID 11823806. S2CID 15725675.
- Rolls, E.T & Treves, A. (1998). Neural Networks and Brain Function. Oxford: OUP. ISBN 0-19-852432-3.
- Santarelli L, Saxe M, Gross C, et al. (Srpen 2003). "Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants". Věda. 301 (5634): 805–9. Bibcode:2003Sci...301..805S. doi:10.1126/science.1083328. PMID 12907793. S2CID 9699898.
- Schloesser RJ, Manji HK, Martinowich K (April 2009). „Potlačení neurogeneze dospělých vede ke zvýšené reakci osy hypotalamo-hypofýza-nadledvina“. NeuroReport. 20 (6): 553–7. doi:10.1097 / WNR.0b013e3283293e59. PMC 2693911. PMID 19322118.
- Shankle WR, Rafii MS, Landing BH, Fallon JH (1999). „Přibližné zdvojnásobení počtu neuronů v postnatální lidské kůře a ve 35 konkrétních cytoarchitektonických oblastech od narození do 72 měsíců“. Dětská a vývojová patologie. 2 (3): 244–259. doi:10,1007 / s100249900120. PMID 10191348. S2CID 23546857.
- Zhao M, Momma S, Delfani K a kol. (Červen 2003). „Důkazy o neurogenezi u dospělých savců substantia nigra“. Proc. Natl. Acad. Sci. USA. 100 (13): 7925–30. Bibcode:2003PNAS..100.7925Z. doi:10.1073 / pnas.1131955100. PMC 164689. PMID 12792021.
- Vyhrazené vydání Filozofické transakce B na kmenových buňkách a opravě mozku. Některé články jsou volně dostupné.[trvalý mrtvý odkaz ]
externí odkazy
- Stručný úvod do neurogeneze z Wellesley College
- Komplexní web o neurogenezi z Lafayette College
- Časná literatura o neurogenezi dospělých
- Neurogeneze v mozku dospělých - Fred H. Gage a Henriette van Praag
- "Neurogeneze a Parkinsonova choroba"
- Článek Scholarpedia o neurogenezi dospělých
- „Trends in Neurosciences, 10. října 2001 (Michael S. Kaplan MD, PhD)
- New York Times: Studie zjistily, že mozkům rostou nové buňky
- Michael Spectre: Přehodnocení mozku - Jak písničky kanárků narušily základní vědecký princip
- Experiment neurogeneze - Série článků o neurogenezi dospělých lidí
- Seed magazine: The Reinvention of the Self - Historické pozadí v oblasti neurogeneze a důsledky tohoto výzkumu
- BBC Radio 4: The Experience Experience - Použij to nebo to zahoď
- PBS: Změna mysli - Vypěstujte si vlastní mozek
- Lobes of Steel: Zdá se, že aerobní cvičení podporuje neurogenezi, New York Times, 19. srpna 2007.
 Média související s neurogeneze na Wikimedia Commons
Média související s neurogeneze na Wikimedia Commons