Neurální záhyb - Neural fold
| Neurální záhyb | |
|---|---|
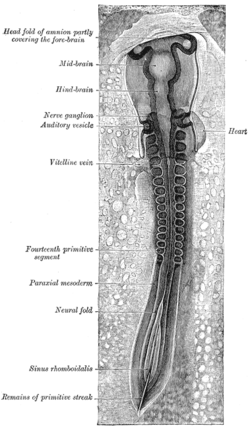 Kuřecí embryo třicet tři hodin inkubace, při pohledu z dorzálního hlediska. 30x. (Neurální záhyb označený vlevo dole, třetí zdola.) | |
| Detaily | |
| Carnegie fáze | 9 |
| Předchůdce | neurální deska |
| Dává povstat | neurální trubice |
| Identifikátory | |
| latinský | plica neuralis |
| TE | E5.13.1.0.1.0.2 |
| Anatomická terminologie | |
The nervový záhyb je struktura, která vzniká během neurulace v embryonálním vývoji obou ptactvo a savci mezi jinými organismy.[1][2] Tato struktura je spojena s primární neurulace, což znamená, že se tvoří spojením vrstev tkáně, spíše než shlukováním a následným vydutím jednotlivých buněk (známých jako sekundární neurulace ). U lidí jsou nervové záhyby zodpovědné za vznik přední konec neurální trubice. Nervové záhyby jsou odvozeny od neurální deska, předběžná struktura skládající se z protáhlé ektoderm buňky. Záhyby vznikají buňky neurální lišty, jakož i vytvoření formace neurální trubice.[1][3]
Rozvoj
U embrya vzniká tvorba nervových záhybů z oblasti, kde neurální deska a okolí ektoderm konvergovat. Tato oblast embrya je vytvořena po gastrulace a sestává z epiteliální tkáně. Zde se epiteliální buňky prodlužují pomocí mikrotubul polymerace, zvýšení jejich výšky. Miniatura níže ukazuje tento proces, stejně jako následnou tvorbu buněk neurální lišty a neurální trubice, které vznikají spojením nervových záhybů.[4]
Skládací

Tvorba nervového záhybu je zahájena uvolněním vápník z buněk. Uvolněný vápník interaguje s bílkovinami, které mohou modifikovat aktin vlákna ve vnější epiteliální tkáni nebo ektodermu, aby se vyvolaly dynamické pohyby buněk nezbytné k vytvoření záhybu.[6] Tyto buňky drží pohromadě kadheriny (konkrétně E a N-kadherin), typy mezibuněčného vazebného proteinu. Když se buňky na vrcholech nervových záhybů dostanou do vzájemné blízkosti, je to afinita k podobným kadherin molekuly (N-kadheriny), které umožňují těmto buňkám vzájemně se vázat. Když tedy prekurzorové buňky neurální trubice začnou exprimovat N-kadherin místo E-kadherinu, způsobí to vytvoření a oddělení neurální trubice od ektodermu a usazení uvnitř embrya.[1] Když se buňky nespojí způsobem, který není součástí normálního vývoje, je to vážné nemoci může dojít.
Přehled procesu
Proces skládání začíná, když se buňky v centrální oblasti nervové ploténky, buňky středního kloubového bodu, váží na notochord pod nimi. Tím se vytvoří centrální bod ukotvení pro proces skládání a následně se vytvoří nervová drážka. Jak se nervové záhyby stále rozšiřují, tvoří se dorzolaterální body závěsu, které umožňují záhybům zakřivit se do trubkovité struktury. Když se vrcholy záhybů (známé jako oblasti neurální lišty) dotýkají, splývají a evolvují a vytvářejí neurální trubici pod nově vytvořenou epidermální vrstvou.[7]
Mechanismus
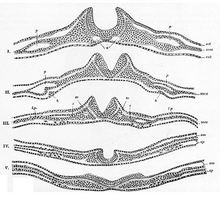
Molekulární mechanismus tohoto procesu spočívá v expresi a represi kostní morfogenetické proteiny (BMP). BMP jsou široká rodina proteinů, které plní mnoho funkcí v průběhu růstu embrya, včetně stimulace růstu chrupavky a kostí. Aby se umožnil růst prekurzorových nervových tkání, na rozdíl od prekurzorových tkání nebo chrupavek je exprese BMP snížena v nervové ploténce, konkrétně podél střední linie, kde se brzy vytvoří nervová drážka. Proteiny produkované z geny Člun ze dřeva a Chordin inhibují tyto BMP a následně umožňují geny neurálního závazku SOX, k vyjádření. Tyto geny kódují transkripční faktory, které mění genomovou expresi těchto buněk a podporují je cestou nervových buněk.[8]Tento proces inhibice BMP umožňuje ukotvení buněk mediálního kloubového bodu, což poskytuje nervovým záhybům základ nezbytný pro složení a uzavření. Člun ze dřeva a Chordin mají další role v procesu neurulace, včetně stimulace buněk neurální lišty k emigraci z nově vytvořené neurální trubice.[9][10] The Sonic ježek Gen také hraje roli při oslabování exprese BMP, formování mediálního kloubového bodu při inhibici tvorby dorzolaterálních kloubových bodů a při zajišťování správného uzavření nervových záhybů.[11]Předchordální destička, notochord a non-neurální ektoderm jsou považovány za důležité indukční tkáně, které uvolňují tyto chemické signály, aby spustily skládání nervové destičky.[8]
Konečná adheze konvergujících nervových záhybů je způsobena několika různými typy mezibuněčných vazebných proteinů. Kadheriny a jejich molekuly CAM receptoru jsou například přítomny ve dvou typech v nervové prekurzorové tkáni: E-kadherin udržuje buňky nervové ploténky a okolní ektoderm se navzájem adherují, zatímco N-kadherin dělá totéž pro buňky nervový záhyb. Pouze buňky exprimující stejný druh kadherinu se mohou navzájem vázat; protože vrcholy nervových záhybů oba exprimují N-kadherin, jsou schopné sloučit se do souvislého listu buněk. Stejně tak je to tato snížená afinita mezi buňkami exprimujícími různé typy kadherinu, která umožňuje oddělit prekurzorové buňky neurální trubice od ektodermu a vytvořit tak neurální trubici uvnitř embrya a pravou epidermis vnějšku.[1]Další sadou molekul zapojených do slučování nervových záhybů jsou molekuly efrinu a jejich Eph receptory, které se drží podobným způsobem jako molekuly kadherinu diskutované výše.[8]
Derivační struktury
Sloučení nervových záhybů vede k mnoha strukturám, včetně: neurální trubice (předchůdce centrální nervový systém ), buňky neurální lišty (které vedou k rozmanitosti různých mezenchymální buňky) a do pravdy epidermální vrstva.[1] Nervový záhyb je nesmírně důležitá struktura v tom, že tento mechanismus je potřebný k produkci těchto různých druhů buněk na správných místech.
Klinický význam

Existuje mnoho potenciálních onemocnění, která mohou vzniknout nesprávnou adhezí nebo sloučením nervových záhybů. Během skládání jsou otvory, které jsou vytvořeny na lebeční a ocasní oblasti se nazývají lebeční a kaudální neuropóry.[12] Pokud se kaudální neuropor neuzavře, došlo k onemocnění spina bifida může nastat, že spodní část míchy zůstane odkrytá. Tento stav lze často zjistit během prenatální vyšetření a být léčeni před narozením, i když v závažnějších případech může jedinec zvládnout tento stav po zbytek svého života.[13] V závislosti na závažnosti a postižené oblasti mohou jednotlivci pociťovat řadu příznaků, včetně různé motorické funkce a pohyblivosti, kontroly močového měchýře a / nebo sexuální funkce.[14]
Pokud je porucha místo toho v lebečním neuroporu, anencefálie dojde. V tomto stavu je mozková tkáň přímo vystavena působení plodová voda, a následně degradován.[15] Pokud se celá neurální trubice neuzavře, stav se označuje jako craniorachischisis.
Viz také
Reference
- ^ A b C d E Gilbert, Scott F. (2010). Vývojová biologie (9. vydání). Sunderland, Massachusetts: Sinauer Associates. ISBN 978-0878933846.[stránka potřebná ]
- ^ Colas JF, Schoenwolf GC (červen 2001). "Směrem k buněčnému a molekulárnímu porozumění neurulaci". Dynamika vývoje. 221 (2): 117–45. doi:10.1002 / dvdy.1144. PMID 11376482.
- ^ Yamaguchi Y, Miura M (září 2013). „Jak formovat a uzavřít mozek: pohled na mechanismus uzavření lebeční neurální trubice u savců“. Buněčné a molekulární biologické vědy. 70 (17): 3171–86. doi:10.1007 / s00018-012-1227-7. PMC 3742426. PMID 23242429.
- ^ Lawson A, Anderson H, Schoenwolf GC (únor 2001). "Buněčné mechanismy tvorby nervových záhybů a morfogeneze u kuřecího embrya". Anatomický záznam. 262 (2): 153–68. doi:10.1002 / 1097-0185 (20010201) 262: 2 <153 :: AID-AR1021> 3.0.CO; 2-W. PMID 11169910.
- ^ "File: Embryonic Development CNS.gif". Wikimedia Commons. 2012-04-04. Citováno 1. dubna 2013.
- ^ Ferreira MC, Hilfer SR (říjen 1993). "Regulace vápníku při tvorbě nervových záhybů: vizualizace aktinového cytoskeletu v živých kuřecích embryích". Vývojová biologie. 159 (2): 427–40. doi:10.1006 / dbio.1993.1253. PMID 8405669.
- ^ Rocky S. Tuan; Cecilia W. Lo, vyd. (2000). "15". Protokoly vývojové biologie, svazek 136. Humama. str.125–134. ISBN 9781592590650. Citováno 1. dubna 2013.
- ^ A b C Khong, Hrsg. Jean W. Keeling; Hrsg. T. Yee (2007). Fetální a neonatální patologie (4. vydání). Godalming: Springer London. 702–704. ISBN 978-1846285240.
- ^ Anderson RM, Stottmann RW, Choi M, Klingensmith J (září 2006). „Antagonisté morfogenetického proteinu endogenní kosti regulují tvorbu a přežití nervového hřebenu savců“. Dynamika vývoje. 235 (9): 2507–20. doi:10.1002 / dvdy.20891. PMC 6626635. PMID 16894609.
- ^ Stottmann RW, Berrong M, Matta K, Choi M, Klingensmith J (červenec 2006). „Antagonista BMP Noggin podporuje kraniální a spinální neurulaci odlišnými mechanismy“. Vývojová biologie. 295 (2): 647–63. doi:10.1016 / j.ydbio.2006.03.051. PMC 3001110. PMID 16712836.
- ^ Kirillova I, Novikova I, Augé J a kol. (Květen 2000). "Exprese genu sonického ježka v lidských embryích s defekty neurální trubice". Teratologie. 61 (5): 347–54. doi:10.1002 / (SICI) 1096-9926 (200005) 61: 5 <347 :: AID-TERA6> 3.0.CO; 2- #. PMID 10777830.
- ^ Gilbert, SF (2000). „12: Formace neurální trubice“. Vývojová biologie (6. vydání). Sunderland, MA: Sinauer Associates. ISBN 978-0-87893-243-6. Citováno 30. listopadu 2011.
- ^ "Spina Bifida". Citováno 1. dubna 2013.
- ^ "SB a páteř". Další informace o Spina Bifida. Archivovány od originál dne 23. dubna 2013. Citováno 1. dubna 2013.
- ^ „7.2: Trilaminární zárodečný disk (3. týden)“. Lidská embryologie: embryogeneze. Citováno 22. března 2013.
{kind=link}
externí odkazy
- YouTube video o embryonální kuřátkové neurulaci
- Anatomy of the Human Body's The Neural Tube and Groove od Henry Gray
| Kontrolní úřad |
|---|