Cytochrom P450 - Cytochrome P450 - Wikipedia
| Cytochrom P450 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
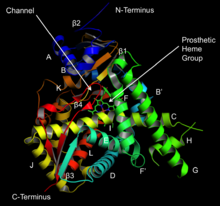 Struktura lanosterol 14α-demethylázy (CYP51 ) | |||||||||
| Identifikátory | |||||||||
| Symbol | p450 | ||||||||
| Pfam | PF00067 | ||||||||
| InterPro | IPR001128 | ||||||||
| STRÁNKA | PDOC00081 | ||||||||
| SCOP2 | 2cpp / Rozsah / SUPFAM | ||||||||
| OPM nadčeleď | 39 | ||||||||
| OPM protein | 2 bdm | ||||||||
| Membranome | 265 | ||||||||
| |||||||||
Cytochromy P450 (CYP) plocha nadčeleď z enzymy obsahující heme jako kofaktor že funkce jako monooxygenázy.[1][2][3] U savců tyto proteiny oxidují steroidy, mastné kyseliny, a xenobiotika, a jsou důležité pro odbavení různých sloučenin, stejně jako pro syntézu a rozklad hormonů. V rostlinách jsou tyto proteiny důležité pro biosyntézu obranné sloučeniny, mastné kyseliny a hormony.[2]
Enzymy CYP byly identifikovány ve všech království života: zvířata, rostliny, houby, protistů, bakterie, a archaea, stejně jako v viry.[4] Nejsou však všudypřítomné; například nebyly nalezeny v Escherichia coli.[3][5] Od roku 2018[Aktualizace], je známo více než 300 000 odlišných proteinů CYP.[6][7]
CYP jsou obecně terminální oxidáza enzymy v elektronový přenos řetězy, obecně klasifikované jako Systémy obsahující P450. Termín "P450" je odvozen od spektrofotometrické vrchol na vlnová délka z absorpční maximum enzymu (450nm ) když je v snížena stát a komplex s kysličník uhelnatý. Většina CYP vyžaduje, aby jeden nebo více proteinových partnerů bylo dodáno elektrony snížit žehlička (a nakonec molekulární kyslík ).
Nomenklatura
Geny kódující enzymy CYP a samotné enzymy jsou označeny kořenový symbol CYP pro nadčeleď, následované číslem označujícím genová rodina, velké písmeno označující podrodinu a další číslice pro jednotlivý gen. Konvence je kurzíva název, když se odkazuje na gen. Například, CYP2E1 je gen, který kóduje enzym CYP2E1 —Jeden z enzymů zapojených do paracetamol (acetaminofen) metabolismus. The CYP nomenklatura je oficiální konvence pojmenování, i když občas CYP450 nebo CYP450 se používá synonymně. Některé názvy genů nebo enzymů pro CYP se však mohou od této nomenklatury lišit, což znamená katalytickou aktivitu a název sloučeniny použité jako substrát. Mezi příklady patří CYP5A1, tromboxan A2 syntáza, zkráceno na TBXAS1 (ThromBÓXznovu A2 Synthase 1), a CYP51A1 lanosterol 14-α-demethyláza, někdy neoficiálně zkrácená na LDM podle svého substrátu (Lanosterol) a aktivita (DEMethylace).[8]
Současné pokyny pro nomenklaturu naznačují, že členové nových rodin CYP sdílejí alespoň 40% aminokyselina identity, zatímco členové podskupin musí sdílet alespoň 55% aminokyselinovou identitu. Existují výbory pro nomenklaturu, které přiřazují a sledují názvy obou základních genů (Domovská stránka cytochromu P450 ) a alela jména (Výbor pro nomenklaturu alel CYP ).[9][10]
Klasifikace
Na základě povahy proteinů pro přenos elektronů lze CYP rozdělit do několika skupin:[11]
- Mikrozomální systémy P450
- ve kterém jsou přenášeny elektrony NADPH přes cytochrom P450 reduktáza (různě CPR, POR nebo CYPOR). Cytochrom b5 (cyb5) může také přispět ke snížení výkonu tohoto systému poté, co byl snížen o cytochrom b5 reduktáza (CYB5R).
- Mitochondriální systémy P450
- které zaměstnávají adrenodoxin reduktáza a adrenodoxin k přenosu elektronů z NADPH na P450.
- Bakteriální systémy P450
- které zaměstnávají a ferredoxin reduktáza a a ferredoxin k přenosu elektronů na P450.
- CYB5R / cyb5/ P450 systémy, ve kterém oba elektrony vyžadované CYP pocházejí z cytochromu b5.
- Systémy FMN / Fd / P450
- původně nalezen v Rhodococcus druh, ve kterém a FMN -obsahující doménu reduktáza je fúzován s CYP.
- Pouze P450
- systémy, které nevyžadují externí redukční výkon. Pozoruhodné patří tromboxan syntáza (CYP5), prostacyklin syntáza (CYP8) a CYP74A (allenoxid syntáza ).
Nejběžnější reakcí katalyzovanou cytochromy P450 je a monooxygenáza reakce, např. vložení jednoho atomu kyslíku do alifatické polohy organického substrátu (RH), zatímco druhý atom kyslíku je snížena zalít:
RH + O2 + NADPH + H+ → ROH + H2O + NADP+
Mnoho hydroxylace reakce (vložení hydroxyl skupiny) používají enzymy CYP.
Mechanismus
Struktura
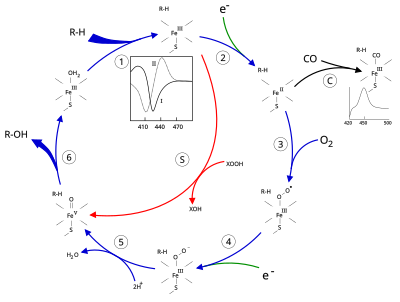
Aktivní místo cytochromu P450 obsahuje a heme - železné centrum. Železo je připoutáno k proteinu pomocí a cystein thiolát ligand. Tento cystein a několik přilehlých zbytků je vysoce konzervováno u známých CYP a má formální formu STRÁNKA podpisový konsensuální vzor [FW] - [SGNH] - x - [GD] - {F} - [RKHPT] - {P} - C - [LIVMFAP] - [GAD].[12] Kvůli velké škále reakcí katalyzovaných CYP se aktivity a vlastnosti mnoha CYP liší v mnoha aspektech.[13] Katalytický cyklus P450 obecně probíhá následovně:
Katalytický cyklus
- Substrát se váže v blízkosti heme skupina, na straně protilehlé k axiálnímu thiolátu. Vazba substrátu vyvolává změnu v konformaci aktivního místa, často vytlačuje molekulu vody z distální axiální koordinační polohy hemového železa,[14] a změna stavu hemového železa z low-spin na high-spin.[15]
- Vazba substrátu indukuje přenos elektronů z NAD (P) H prostřednictvím cytochrom P450 reduktáza nebo jiný přidružený reduktáza.[16]
- Molekulární kyslík se váže na výsledné železné centrum hemu v distální axiální koordinační poloze, přičemž nejprve dává a dioxygenový adukt ne na rozdíl od oxy-myoglobinu.
- Druhý elektron je přenesen z obou cytochrom P450 reduktáza, ferredoxiny nebo cytochrom b5, snížení Fe-O2 adukt, který dává krátkodobý peroxo stav.
- Peroxoskupina vytvořená v kroku 4 je dvakrát rychle protonována, přičemž uvolní jednu molekulu vody a vytvoří vysoce reaktivní druhy označované jako Sloučenina P450 1 (nebo jen sloučenina I). Tento vysoce reaktivní meziprodukt byl izolován v roce 2010,[17] Sloučenina P450 1 je oxo (IV) železo trajekt ) druhy s dalším oxidačním ekvivalentem přemístěn přes porfyrin a thiolátové ligandy. Důkazy pro alternativní perferryl železo (V) -oxo [14] chybí.[17]
- V závislosti na použitém substrátu a enzymu mohou enzymy P450 katalyzovat jakoukoli z celé řady reakcí. Na tomto obrázku je znázorněna hypotetická hydroxylace. Po uvolnění produktu z aktivního místa se enzym vrací do původního stavu, přičemž molekula vody se vrací a zaujímá distální koordinační polohu jádra železa.

- Alternativní cesta pro mono-okysličení je přes „peroxidový zkrat“ (cesta „S“ na obrázku). Tato cesta zahrnuje oxidaci komplexu železitý-substrát s donory atomu kyslíku, jako jsou peroxidy a chlornany.[18] Na obrázku je znázorněn hypotetický peroxid „XOOH“.
Spektroskopie
Vazba substrátu se odráží ve spektrálních vlastnostech enzymu se zvýšením absorbance při 390 nm a snížením při 420 nm. To lze měřit rozdílovou spektroskopií a označuje se jako rozdílové spektrum „typu I“ (viz vložený graf na obrázku). Některé substráty způsobují dosud nejasné procesy ve spektrálních vlastnostech, spektrum „reverzního typu I“. Inhibitory a určité substráty, které se vážou přímo na hemové železo, vytvářejí rozdílné spektrum typu II, s maximem při 430 nm a minimem při 390 nm (viz vložený graf na obrázku). Pokud nejsou k dispozici žádné redukční ekvivalenty, může tento komplex zůstat stabilní, což umožňuje určit stupeň vazby z měření absorbance in vitro[18]C: Pokud se oxid uhelnatý (CO) váže na redukovanou P450, je katalytický cyklus přerušen. Tato reakce poskytuje klasické rozdílové spektrum CO s maximem při 450 nm.
P450 u lidí
Lidské CYP jsou primárně proteiny spojené s membránou[19] nachází se buď ve vnitřní membráně mitochondrie nebo v endoplazmatické retikulum buněk. CYP metabolizují tisíce endogenní a exogenní Chemikálie. Některé CYP metabolizují pouze jeden (nebo jen velmi málo) substrátů, jako např CYP19 (aromatáza ), zatímco jiné mohou více metabolizovat substráty. Obě tyto charakteristiky odpovídají jejich ústřednímu významu v lék. Enzymy cytochromu P450 jsou přítomny ve většině tkání těla a hrají v nich důležitou roli hormon syntéza a rozpad (včetně estrogen a testosteron syntéza a metabolismus), cholesterol syntéza a Vitamín D metabolismus. Enzymy cytochromu P450 také fungují při metabolizaci potenciálně toxických sloučenin, včetně léky a produkty endogenního metabolismu jako např bilirubin, hlavně v játra.
The Projekt lidského genomu identifikovala 57 lidských genů kódujících různé enzymy cytochromu P450.[20]
Metabolismus drog

CYP jsou hlavní enzymy podílející se na metabolismus léků, což představuje asi 75% celkového metabolismu.[22] Většina léků podléhá deaktivaci pomocí CYP, a to buď přímo, nebo zprostředkovaně vylučování z těla. Mnoho látek také je bioaktivováno pomocí CYP za vzniku jejich aktivních sloučenin, jako je protidestičkový lék klopidogrel.
Léková interakce
Mnoho léků může zvyšovat nebo snižovat aktivitu různých izoenzymů CYP buď indukcí biosyntézy izozymu (indukce enzymů ) nebo přímou inhibicí aktivity CYP (inhibice enzymů ). Klasický příklad zahrnuje antiepileptika, jako Fenytoin, který indukuje CYP1A2, CYP2C9, CYP2C19, a CYP3A4.
Účinky na aktivitu izozymu CYP jsou hlavním zdrojem nežádoucích účinků lékové interakce, protože změny aktivity enzymu CYP mohou ovlivnit metabolismus a odbavení různých drog. Například pokud jeden lék inhibuje metabolismus jiného léku zprostředkovaný CYP, může se druhý lék hromadit v těle na toxické hladiny. Tyto lékové interakce proto mohou vyžadovat úpravu dávkování nebo výběr léků, které neinteragují se systémem CYP. Tyto lékové interakce je zvláště důležité vzít v úvahu při užívání léků zásadního významu pro pacienta, léků s významným účinkem vedlejší efekty nebo léky s úzkým terapeutický index, ale jakýkoli lék může podléhat změněné plazmatické koncentraci v důsledku změněného metabolismu léčiva.
Mnoho substrátů pro CYP3A4 jsou léky s úzkým terapeutickým indexem, jako např amiodaron[23] nebo karbamazepin.[24] Protože tyto léky jsou metabolizovány CYP3A4, průměrné plazmatické hladiny těchto léků se může zvýšit kvůli inhibici enzymů nebo snížit kvůli indukci enzymů.
Interakce jiných látek
Přirozeně se vyskytující sloučeniny mohou také indukovat nebo inhibovat aktivitu CYP. Například, bioaktivní sloučeniny nalezené v grepový džus a některé další ovocné šťávy, včetně bergamotin, dihydroxybergamottin, a paradicin-A Bylo zjištěno, že inhibují metabolismus CYP3A4 zprostředkovaný některé léky, což vede ke zvýšení biologická dostupnost a tedy silná možnost předávkování.[25] Kvůli tomuto riziku se obvykle doporučuje vyhnout se grapefruitovému džusu a čerstvým grapefruitům úplně, když užíváte drogy.[26]
Další příklady:
- Třezalka tečkovaná, běžný bylinný lék indukuje CYP3A4, ale také inhibuje CYP1A1, CYP1B1.[27][28]
- Kouření tabáku indukuje CYP1A2 (příkladem jsou substráty CYP1A2 klozapin, olanzapin, a fluvoxamin )[29]
- Při relativně vysokých koncentracích hvězdné ovoce Bylo také prokázáno, že šťáva inhibuje CYP2A6 a další CYP.[30] Řeřicha je také známým inhibitorem cytochromu P450 CYP2E1, což může vést ke změně metabolismu léků u jednotlivců užívajících určité léky (např. chlorzoxazon ).[31]
- Tributylcín Bylo zjištěno, že inhibuje funkci cytochromu P450, což vede k maskulinizaci měkkýšů.[32]
- Goldenseal, se svými dvěma významnými alkaloidy berberine a hydrastin Bylo prokázáno, že mění enzymatické aktivity markeru P450 (zahrnující CYP2C9, CYP2D6 a CYP3A4).[33]
Další specifické funkce CYP
Steroidní hormony
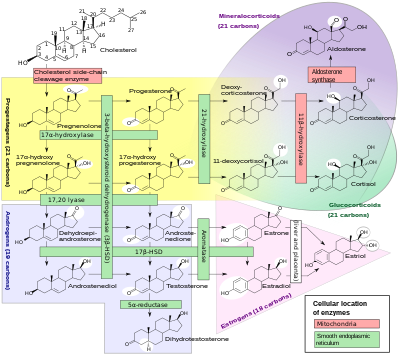
Podskupina enzymů cytochromu P450 hraje důležitou roli při syntéze steroidní hormony (steroidogeneze ) podle nadledviny, pohlavní žlázy a periferní tkáň:
- CYP11A1 (také známý jako P450scc nebo P450c11a1) v nadledvinách mitochondrie ovlivňuje „aktivitu dříve známou jako 20,22-desmoláza“ (steroid 20α-hydroxyláza, steroid 22-hydroxyláza, cholesterol postranní řetěz rozštěpení).
- CYP11B1 (kódující protein P450c11β) nalezený v vnitřní mitochondriální membrána z kůra nadledvin má steroid 11β-hydroxylázu, steroid 18-hydroxyláza a aktivity steroidní 18-methyloxidázy.
- CYP11B2 (kódující protein P450c11AS), nalezený pouze v mitochondriích nadledvin zona glomerulosa, má aktivity steroidní 11p-hydroxylázy, steroidní 18-hydroxylázy a steroidní 18-methyloxidázy.
- CYP17A1, v endoplazmatickém retikulu kůry nadledvin má aktivity steroidů 17α-hydroxylázy a 17,20-lyázy.
- CYP21A2 (P450c21) ve vedení kůry nadledvin 21-hydroxyláza aktivita.
- CYP19A (P450arom, aromatáza ) v endoplazmatické retikulum z pohlavní žlázy, mozek, tuková tkáň a jinde katalyzuje aromatizaci androgeny na estrogeny.
Polynenasycené mastné kyseliny a eikosanoidy
Určité enzymy cytochromu P450 mají zásadní význam pro metabolismus polynenasycené mastné kyseliny (PUFA) na biologicky aktivní, mezibuněčný buněčná signalizace molekuly (eikosanoidy ) a / nebo metabolizovat biologicky aktivní metabolity PUFA na méně aktivní nebo neaktivní produkty. Tyto CYP mají cytochrom P450 omega hydroxyláza a / nebo epoxygenáza aktivita enzymu.
- CYP1A1, CYP1A2, a CYP2E1 metabolizují endogenní PUFA na signální molekuly: metabolizují kyselina arachidonová (tj. AA) na kyselinu 19-hydroxyeikosatetraenovou (tj. 19-HETE; viz 20-hydroxyeikosatetraenová kyselina ); kyselina eikosapentaenová (tj. EPA) do epoxyeikosatetraenové kyseliny (tj. EEQ); a kyselina dokosahexaenová (tj. DHA) do epoxydokosapentaenové kyseliny (tj. EDP).
- CYP2C8, CYP2C9, CYP2C18, CYP2C19, a CYP2J2 metabolizují endogenní PUFA na signální molekuly: metabolizují AA na epoxyeikosatetraenové kyseliny (tj. EET); EPA až EEQ; a DHA EDP.
- CYP2S1 metabolizuje PUFA na signální molekuly: metabolizuje AA na EET a EPA na EEQ.
- CYP3A4 metabolizuje AA na signální molekuly EET.
- CYP4A11 metabolizuje endogenní PUFA na signální molekuly: metabolizuje AA na 20-HETE a EET; také hydroxyláty DHA na 22-hydroxy-DHA (tj. 12-HDHA).
- CYP4F2, CYP4F3A a CYP4F3B (viz CYP4F3 u posledních dvou CYP) metabolizují PUFA na signální molekuly: metabolizují AA na 20-HETE. Rovněž metabolizují EPA na kyselinu 19-hydroxyeikosapentaenovou (19-HEPE) a kyselinu 20-hydroxyeikosapentaenovou (20-HEPE) a metabolizují DHA na 22-HDA. Rovněž deaktivují nebo snižují aktivitu signálních molekul: metabolizují se leukotrien B4 (LTB4) až 20-hydroxy-LTB4, Kyselina 5-hydroxyeikosatetraenová (5-HETE) až 5,20-diHETE, Kyselina 5-oxo-eikosatetraenová (5-oxo-ETE) až 5-oxo, 20-hydroxy-ETE, Kyselina 12-hydroxyeikosatetraenová (12-HETE) až 12,20-diHETE, EET až 20-hydroxy-EET a lipoxiny na 20-hydroxy produkty.
- CYP4F8 a CYP4F12 metabolizují PUFA na signální molekuly: metabolizují EPA na EEQ a DHA na EDP. Rovněž metabolizují AA na kyselinu 18-hydroxyeikosatetraenovou (18-HETE) a 19-HETE.
- CYP4F11 inaktivuje nebo snižuje aktivitu signálních molekul: metabolizuje LTB4 na 20-hydroxy-LTB4, (5-HETE) na 5,20-diHETE, (5-oxo-ETE) na 5-oxo, 20-hydroxy-ETE, ( 12-HETE) až 12,20-diHETE, EET až 20-hydroxy-EET a lipoxiny na 20-hydroxy produkty.
- CYP4F22 ω-hydroxyláty extrémně dlouhé "mastné kyseliny s velmi dlouhým řetězcem „, tj. mastné kyseliny, které mají 28 nebo více uhlíků. ω-hydroxylace těchto speciálních mastných kyselin je zásadní pro vytvoření a udržení funkce vodní bariéry pokožky; autozomálně recesivní inaktivující mutace CYP4F22 jsou spojeny Lamelová ichtyóza podtyp Vrozený ichtyosiformní erytrodém u lidí.[35]
Rodiny CYP u lidí
Lidé mají 57 genů a více než 59 pseudogeny rozdělena mezi 18 rodin genů cytochromu P450 a 43 podrodin.[36] Toto je souhrn genů a proteinů, které kódují. Podrobné informace najdete na domovské stránce Výboru pro názvosloví cytochromu P450.[20]
| Rodina | Funkce | Členové | Geny | pseudogeny |
| CYP1 | droga a steroid (zejména estrogen ) metabolismus, benzo [A] pyren toxifikace (formování (+) - benzo [A] pyren-7,8-dihydrodiol-9,10-epoxid ) | 3 podrodiny, 3 geny, 1 pseudogen | CYP1A1, CYP1A2, CYP1B1 | CYP1D1P |
| CYP2 | droga a steroid metabolismus | 13 podrodin, 16 genů, 16 pseudogeny | CYP2A6, CYP2A7, CYP2A13, CYP2B6, CYP2C8, CYP2C9, CYP2C18, CYP2C19, CYP2D6, CYP2E1, CYP2F1, CYP2J2, CYP2R1, CYP2S1, CYP2U1, CYP2W1 | Příliš mnoho na seznam |
| CYP3 | droga a steroid (počítaje v to testosteron ) metabolismus | 1 podčeleď, 4 geny, 4 pseudogeny | CYP3A4, CYP3A5, CYP3A7, CYP3A43 | CYP3A51P, CYP3A52P, CYP3A54P, CYP3A137P |
| CYP4 | kyselina arachidonová nebo metabolismus mastných kyselin | 6 podskupin, 12 genů, 10 pseudogeny | CYP4A11, CYP4A22, CYP4B1, CYP4F2, CYP4F3, CYP4F8, CYP4F11, CYP4F12, CYP4F22, CYP4V2, CYP4X1, CYP4Z1 | Příliš mnoho na seznam |
| CYP5 | tromboxan A2 syntáza | 1 podčeleď, 1 gen | CYP5A1 | |
| CYP7 | žlučová kyselina biosyntéza 7-alfa hydroxylázy steroidního jádra | 2 podrodiny, 2 geny | CYP7A1, CYP7B1 | |
| CYP8 | pestrý | 2 podrodiny, 2 geny | CYP8A1 (prostacyklin syntáza), CYP8B1 (biosyntéza žlučových kyselin) | |
| CYP11 | steroid biosyntéza | 2 podrodiny, 3 geny | CYP11A1, CYP11B1, CYP11B2 | |
| CYP17 | steroid biosyntéza, 17-alfa hydroxyláza | 1 podčeleď, 1 gen | CYP17A1 | |
| CYP19 | steroid biosyntéza: aromatáza syntetizuje estrogen | 1 podčeleď, 1 gen | CYP19A1 | |
| CYP20 | neznámá funkce | 1 podčeleď, 1 gen | CYP20A1 | |
| CYP21 | steroid biosyntéza | 1 podrodiny, 1 gen, 1 pseudogen | CYP21A2 | CYP21A1P |
| CYP24 | Vitamín D degradace | 1 podčeleď, 1 gen | CYP24A1 | |
| CYP26 | kyselina retinová hydroxyláza | 3 podrodiny, 3 geny | CYP26A1, CYP26B1, CYP26C1 | |
| CYP27 | pestrý | 3 podrodiny, 3 geny | CYP27A1 (žlučová kyselina biosyntéza), CYP27B1 (Vitamín D3 1-alfa hydroxyláza, aktivuje vitamin D.3), CYP27C1 (neznámá funkce) | |
| CYP39 | 7-alfa hydroxylace 24-hydroxycholesterolu | 1 podčeleď, 1 gen | CYP39A1 | |
| CYP46 | cholesterol 24-hydroxyláza | 1 podčeleď, 1 gen, 1 pseudogen | CYP46A1 | CYP46A4P |
| CYP51 | cholesterol biosyntéza | 1 podčeleď, 1 gen, 3 pseudogeny | CYP51A1 (lanosterol 14-alfa demethyláza) | CYP51P1, CYP51P2, CYP51P3 |
P450 u jiných druhů
Zvířata
Mnoho zvířat má tolik nebo více genů CYP než lidé. Hlášená čísla se pohybují od 35 genů v houbě Amphimedon queenslandica na 235 genů v hlavonožci Branchiostoma floridae.[37] Myši mít geny pro 101 CYP a mořští ježci mají ještě více (možná až 120 genů).[38]U většiny enzymů CYP se předpokládá, že mají aktivitu monooxygenázy, jako je tomu u většiny savčích CYP, které byly zkoumány (kromě např. CYP19 a CYP5 ). Gen a sekvenování genomu je daleko předstihující biochemické charakterizace enzymatické funkce, ačkoli mnoho genů s blízkými homologie Byly nalezeny enzymy CYP se známou funkcí, které poskytují vodítko k jejich funkčnosti.
Třídy CYP, které se nejčastěji zkoumají u zvířat jiných než lidských, jsou ty, kterých se účastní rozvoj (např., kyselina retinová nebo hormon metabolismus) nebo se podílejí na metabolismu toxických sloučenin (např heterocyklické aminy nebo polyaromatické uhlovodíky ). Často existují rozdíly v genová regulace nebo funkce enzymu CYP u příbuzných zvířat, což vysvětluje pozorované rozdíly v citlivosti na toxické sloučeniny (např. neschopnost psů metabolizovat xanthiny, jako je kofein). Některé léky podléhají metabolismu u obou druhů prostřednictvím různých enzymů, což vede k různým metabolitům, zatímco jiné léky jsou metabolizovány u jednoho druhu, ale vylučovány beze změny u jiného druhu. Z tohoto důvodu není reakce jednoho druhu na látku spolehlivým ukazatelem účinků látky na člověka. Druh Sonoran Desert Drosophila, který používá regulovanou expresi CYP28A1 gen pro detoxikaci kaktusové hniloby je Drosophila mettleri. Mouchy tohoto druhu přizpůsobily upregulaci tohoto genu v důsledku expozice vysokým hladinám alkaloidů v hostitelských rostlinách.
CYP byly rozsáhle zkoumány v myši, krysy, psy a už méně zebrafish, aby se usnadnilo jejich používání modelové organismy v objev drog a toxikologie. Nedávno byly také objeveny CYP u ptačích druhů, zejména krůt, které se mohou ukázat jako užitečný model pro výzkum rakoviny u lidí.[39] CYP1A5 a CYP3A37 u krůt bylo zjištěno, že jsou velmi podobné člověku CYP1A2 a CYP3A4 z hlediska jejich kinetických vlastností i metabolismu aflatoxinu B1.[40]
CYP byly také intenzivně studovány v hmyz, často pochopit odolnost vůči pesticidům. Například, CYP6G1 je spojena s insekticidní rezistencí v DDT -odolný Drosophila melanogaster[41] a CYP6M2 v komářích malárie vektor Anopheles gambiae je schopen přímé metabolizace pyrethroidy.[42]
Mikrobiální
Mikrobiální cytochromy P450 jsou často rozpustné enzymy a podílejí se na různých metabolických procesech. V bakteriích je distribuce P450 velmi variabilní, přičemž mnoho bakterií nemá identifikované P450 (např. E. coli). Některé bakterie, převážně aktinomycety, mají četné P450 (např.[43][44]). Dosud identifikované osoby se obecně účastní buď biotransformace xenobiotických sloučenin (např. CYP105A1 z Streptomyces griseolus metabolizuje herbicidy sulfonylmočoviny na méně toxické deriváty,[45]) nebo jsou součástí specializovaných biosyntetických drah metabolitů (např. CYP170B1 katalyzuje produkci sesquiterpenoidu albaflavenonu v Streptomyces albus[46]). Ačkoli dosud nebylo prokázáno, že by P450 byl v mikrobu nezbytný, Rodina CYP105 je vysoce konzervativní a má zástupce ve všech streptomycete genom dosud sekvenován.[47] Vzhledem k rozpustnosti bakteriálních enzymů P450 jsou obecně považovány za jednodušší než s eukaryotickými P450 vázanými na membránu. To v kombinaci s pozoruhodnou chemií, kterou katalyzují, vedlo k mnoha studiím využívajícím heterologně exprimované proteiny in vitro. Několik studií zkoumalo, co P450 dělají in vivo, co jsou přírodní substráty a jak P450 přispívají k přežití bakterií v přírodním prostředí. Zde jsou uvedeny tři příklady, které významně přispěly ke strukturálním a mechanickým studiím, ale mnoho různých rodiny existují.
- Cytochrom P450 cam (CYP101A1) původně z Pseudomonas putida byl použit jako model pro mnoho cytochromů P450 a byl první strukturou trojrozměrného proteinu cytochromu P450 řešenou rentgenovou krystalografií. Tento enzym je součástí katalytického cyklu kafr-hydroxylace, který se skládá ze dvou kroků přenosu elektronů z putidaredoxin, proteinový kofaktor obsahující klastr 2Fe-2S.
- Cytochrom P450 eryF (CYP107A1) původně z bakterie aktinomycety Saccharopolyspora erythraea je zodpovědný za biosyntézu antibiotikum erythromycin C6-hydroxylací makrolidu 6-deoxyerythronolidu B.
- Cytochrom P450 BM3 (CYP102A1) z půdní bakterie Bacillus megaterium katalyzuje NADPH-dependentní hydroxylaci několika mastné kyseliny s dlouhým řetězcem na pozicích ω – 1 až ω – 3. Na rozdíl od téměř všech ostatních známých CYP (kromě CYP505A1, cytochromu P450 foxy) představuje přirozený fúzní protein mezi doménou CYP a elektronovým kofaktorem. BM3 je tedy potenciálně velmi užitečný v biotechnologických aplikacích.[48][49]
- Cytochrom P450 119 (CYP119A1 ) izolovaný od termofilní archea Sulfolobus solfataricus [50] byl použit v různých mechanistických studiích.[17] Protože se termofilní enzymy vyvinuly tak, aby fungovaly při vysokých teplotách, mají tendenci fungovat pomaleji při pokojové teplotě (pokud vůbec), a jsou tedy vynikajícími mechanickými modely.
Houby
Běžně používané azol antifungální léky třídy fungují inhibicí plísní cytochrom P450 14α-demethyláza. Tím se přeruší převod lanosterol na ergosterol, součást buněčné membrány hub. (To je užitečné pouze proto, že lidská P450 má jinou citlivost; takhle tato třída antimykotika práce.)[51]
Probíhá významný výzkum houbových P450, jako je řada hub patogenní lidem (např Candida droždí a Aspergillus ) a rostlinám.
Cunninghamella elegans je kandidátem pro použití jako model metabolismu léčiv u savců.
Rostliny
Rostlinné cytochromy P450 jsou zapojeny do široké škály biosyntetických reakcí a zaměřují se na rozmanitou škálu biomolekul. Tyto reakce vedou k různým mastné kyseliny konjugáty, rostlinné hormony, sekundární metabolity, ligniny a řadu obranných sloučenin.[52] Anotace rostlinného genomu naznačují, že geny cytochromu P450 tvoří až 1% rostlinných genů. Počet a rozmanitost genů P450 je částečně zodpovědný za množství bioaktivních sloučenin.[53]
Cytochrom P450 aromatická O-demethyláza, který je vyroben ze dvou odlišných promiskuitních částí: proteinu cytochromu P450 (GcoA) a tří doménové reduktázy, je významný svou schopností převádět lignin, aromatický biopolymer běžný ve stěnách rostlinných buněk, na obnovitelné uhlíkové řetězce v katabolické sadě reakcí . Stručně řečeno, je to zprostředkovatel kritického kroku při konverzi ligninu.
P450 v biotechnologii
Pozoruhodná reaktivita a promiskuita substrátu u P450 dlouho přitahovala pozornost chemiků.[54] Nedávný pokrok při realizaci potenciálu použití P450 k obtížným oxidacím zahrnoval: (i) eliminaci potřeby přírodních kofaktorů jejich nahrazením levnými molekulami obsahujícími peroxid;[55] ii) zkoumání kompatibility P450 s organickými rozpouštědly,[56] a (iii) použití malých nechirálních pomocných látek k předvídatelně přímé oxidaci P450.[Citace je zapotřebí ]
Podskupiny InterPro
Tato část může vyžadovat vyčištění setkat se s Wikipedií standardy kvality. Specifický problém je: nefunkční odkazy; fragmentovaný odstavec. (Září 2016) (Zjistěte, jak a kdy odstranit tuto zprávu šablony) |
InterPro podrodiny:
- Cytochrom P450, třída B. InterPro: IPR002397
- Cytochrom P450, mitochondriální InterPro: IPR002399
- Cytochrom P450, třída E, skupina I. InterPro: IPR002401
- Cytochrom P450, třída E, skupina II InterPro: IPR002402
- Cytochrom P450, třída E, skupina IV InterPro: IPR002403
- Aromatáza
Klozapin, imipramin, paracetamol, fenacetin Heterocyklické arylaminy Neindukovatelné a 5–10% CYP1A2 deficientoxidují v hemovém metabolismu uroporfyrinogen na uroporfyrin (CYP1A2), ale mohou mít další neobjevené endogenní substráty. jsou indukovatelné některými polycyklickými uhlovodíky, z nichž některé se nacházejí v cigaretovém kouři a zuhelnatělém jídle.
Tyto enzymy jsou zajímavé, protože v testech mohou aktivovat sloučeniny na karcinogeny. Vysoké hladiny CYP1A2 byly spojeny se zvýšeným rizikem rakoviny tlustého střeva. Protože enzym 1A2 může být indukován kouřením cigaret, spojuje to kouření s rakovinou tlustého střeva.[57]
Viz také
Reference
- ^ Gonzalez FJ, Gelboin HV (listopad 1992). „Lidské cytochromy P450: evoluce a cDNA-řízená exprese“. Perspektivy zdraví a životního prostředí. 98: 81–5. doi:10,1289 / ehp. 929881. PMC 1519618. PMID 1486867.
- ^ A b „Cytochrom P450“. InterPro.
- ^ A b Danielson PB (prosinec 2002). „Nadčeleď cytochromu P450: biochemie, evoluce a metabolismus léčiv u lidí“. Současný metabolismus drog. 3 (6): 561–97. doi:10.2174/1389200023337054. PMID 12369887.
- ^ Lamb DC, Lei L, Warrilow AG, Lepesheva GI, Mullins JG, Waterman MR, Kelly SL (srpen 2009). „První virově kódovaný cytochrom p450“. Journal of Virology. 83 (16): 8266–9. doi:10.1128 / JVI.00289-09. PMC 2715754. PMID 19515774.
- ^ Sigel R, Sigel A, Sigel H (2007). Všudypřítomné role proteinů cytochromu P450: kovové ionty v biologických vědách. New York: Wiley. ISBN 978-0-470-01672-5.
- ^ Nelson, David R. (leden 2018). „Rozmanitost cytochromu P450 ve stromu života“. Biochimica et Biophysica Acta (BBA) - bílkoviny a proteomika. 1866 (1): 141–154. doi:10.1016 / j.bbapap.2017.05.003. PMC 5681887. PMID 28502748.
- ^ Nelson D (2009). „Domovská stránka cytochromu P450“. Lidská genomika. University of Tennessee. 4 (1): 59–65. doi:10.1186/1479-7364-4-1-59. PMC 3500189. PMID 19951895. Citováno 2014-11-13.
- ^ „Prohlížeč sekvencí NCBI“. Citováno 2007-11-19.
- ^ Nelson, DR (říjen 2009). „Domovská stránka cytochromu p450“. Lidská genomika. 4 (1): 59–65. doi:10.1186/1479-7364-4-1-59. PMC 3500189. PMID 19951895.
- ^ Nelson, David R. (leden 2011). "Pokrok ve sledování evolučních cest cytochromu P450". Biochimica et Biophysica Acta (BBA) - bílkoviny a proteomika. 1814 (1): 14–18. doi:10.1016 / j.bbapap.2010.08.008. PMID 20736090.
- ^ Hanukoglu I (1996). „Proteiny přenosu elektronů systémů cytochromu P450“ (PDF). Adv. Mol. Cell Biol. Pokroky v molekulární a buněčné biologii. 14: 29–55. doi:10.1016 / S1569-2558 (08) 60339-2. ISBN 9780762301133.
- ^ [1]STRÁNKA konsensuální vzor pro P450
- ^ Srinivasan, Bharath (10. 10. 2020). „Explicitní léčba non Michaelis-Menten a atypická kinetika při včasném objevování drog“. dx.doi.org. Citováno 2020-11-09.
- ^ A b Meunier B, de Visser SP, Shaik S (září 2004). "Mechanismus oxidačních reakcí katalyzovaných enzymy cytochromu p450". Chemické recenze. 104 (9): 3947–80. doi:10,1021 / cr020443g. PMID 15352783. S2CID 33927145.
- ^ Poulos TL, Finzel BC, Howard AJ (červen 1987). "Krystalová struktura cytochromu P450cam s vysokým rozlišením". Journal of Molecular Biology. 195 (3): 687–700. doi:10.1016/0022-2836(87)90190-2. PMID 3656428.
- ^ Sligar SG, Cinti DL, Gibson GG, Schenkman JB (říjen 1979). "Kontrola stavu otáčení redoxního potenciálu jaterního cytochromu P450". Sdělení o biochemickém a biofyzikálním výzkumu. 90 (3): 925–32. doi:10.1016 / 0006-291X (79) 91916-8. PMID 228675.
- ^ A b C Rittle J, Green MT (listopad 2010). "Sloučenina I cytochromu P450: kinetika zachycení, charakterizace a aktivace CH vazby". Věda. 330 (6006): 933–7. Bibcode:2010Sci ... 330..933R. doi:10.1126 / science.1193478. PMID 21071661. S2CID 206528205.
- ^ A b Ortiz de Montellano, Paul R .; Paul R. Ortiz de Montellano (2005). Cytochrom P450: struktura, mechanismus a biochemie (3. vyd.). New York: Kluwer Academic / Plenum Publishers. ISBN 978-0-306-48324-0.
- ^ Berka K, Hendrychová T, Anzenbacher P, Otyepka M (říjen 2011). „Pozice membrány ibuprofenu souhlasí s doporučeným vstupem do přístupové cesty k aktivnímu místu cytochromu P450 2C9“. The Journal of Physical Chemistry A. 115 (41): 11248–55. Bibcode:2011JPCA..11511248B. doi:10.1021 / jp204488j. PMC 3257864. PMID 21744854.
- ^ A b „Tabulka P450“.
- ^ doctorfungus> Antifungální lékové interakce Archivováno 01.08.2012 v Archiv. Dnes Ředitel obsahu: Russell E. Lewis, Pharm.D. Citováno 23. ledna 2010
- ^ Guengerich FP (leden 2008). "Cytochrom p450 a chemická toxikologie". Chemický výzkum v toxikologii. 21 (1): 70–83. doi:10.1021 / tx700079z. PMID 18052394. S2CID 17548932. (Metabolismus je v této souvislosti chemická modifikace nebo degradace léčiv.)
- ^ Zahno, A; Brecht, K; Morand, R; Maseneni, S; Török, M; Lindinger, PW; Krähenbühl, S (1. února 2011). „Úloha CYP3A4 v toxicitě spojené s amiodaronem na buňky HepG2“. Biochem Pharmacol. 81 (3): 432–41. doi:10.1016 / j.bcp.2010.11.002. PMID 21070748.
- ^ „Karbamazepin: Sledujte mnoho potenciálních lékových interakcí“. Lékárna Times.
- ^ Bailey DG, Dresser GK (2004). "Interakce mezi grapefruitovým džusem a kardiovaskulárními léky". American Journal of Cardiovascular Drugs. 4 (5): 281–97. doi:10.2165/00129784-200404050-00002. PMID 15449971. S2CID 11525439.
- ^ Zeratsky K (11.06.2008). „Grapefruitový džus: Může to způsobit lékové interakce?“. Zeptejte se odborníka na potraviny a výživu. MayoClinic.com. Citováno 2009-02-09.
- ^ Chaudhary A, Willett KL (leden 2006). „Inhibice lidských cytochromových enzymů CYP 1 flavonoidy třezalky tečkované“. Toxikologie. 217 (2–3): 194–205. doi:10.1016 / j.tox.2005.09.010. PMID 16271822.
- ^ Strandell J, Neil A, Carlin G (únor 2004). „Přístup k hodnocení in vitro potenciálu pro inhibici enzymů cytochromu P450 z bylin a jiných přírodních léčiv“. Fytomedicin. 11 (2–3): 98–104. doi:10.1078/0944-7113-00379. PMID 15070158.
- ^ Kroon LA (září 2007). "Lékové interakce s kouřením". American Journal of Health-System Pharmacy. 64 (18): 1917–21. doi:10.2146 / ajhp060414. PMID 17823102. S2CID 5397510.
- ^ Zhang JW, Liu Y, Cheng J, Li W, Ma H, Liu HT, Sun J, Wang LM, He YQ, Wang Y, Wang ZT, Yang L (2007). „Inhibice lidského jaterního cytochromu P450 šťávou z hvězdného ovoce“. Journal of Pharmacy & Pharmaceutical Sciences. 10 (4): 496–503. doi:10.18433 / j30593. PMID 18261370.
- ^ Leclercq I, Desager JP, Horsmans Y (srpen 1998). „Inhibice metabolismu chlorzoxazonu, klinická sonda pro CYP2E1, jediným požitím řeřichy“. Klinická farmakologie a terapeutika. 64 (2): 144–9. doi:10.1016 / S0009-9236 (98) 90147-3. PMID 9728894. S2CID 43863786.
- ^ Walmsley, Simon. „Znečištění tributylcinem v globálním měřítku. Přehled relevantního a nedávného výzkumu: dopady a problémy“ (PDF). WWF UK.
- ^ Chatterjee P, Franklin MR (listopad 2003). „Inhibice lidského cytochromu p450 a tvorba metabolicko-meziproduktového komplexu extraktem z Goldenseal a jeho methylendioxyfenylovými složkami“. Metabolismus a dispozice léků. 31 (11): 1391–7. doi:10.1124 / dmd.31.11.1391. PMID 14570772. S2CID 2967171.
- ^ Häggström, Mikael; Richfield, David (2014). „Schéma cest lidské steroidogeneze“. WikiJournal of Medicine. 1 (1). doi:10.15347 / wjm / 2014.005. ISSN 2002-4436.
- ^ Sugiura K, Akiyama M (červenec 2015). „Aktualizace autosomálně recesivní vrozené ichtyózy: analýza mRNA pomocí vzorků vlasů je mocným nástrojem pro genetickou diagnostiku“. Journal of Dermatological Science. 79 (1): 4–9. doi:10.1016 / j.jdermsci.2015.04.009. PMID 25982146.
- ^ Nelson D (2003). Cytochromy P450 u lidí. Citováno 9. května 2005.
- ^ Nelson DR, Goldstone JV, Stegeman JJ (únor 2013). „Místo geneze cytochromu P450: vznik a vývoj zvířecích cytochromů P450“. Filozofické transakce Královské společnosti v Londýně. Série B, Biologické vědy. 368 (1612): 20120474. doi:10.1098 / rstb.2012.0474. PMC 3538424. PMID 23297357.
- ^ Goldstone JV, Hamdoun A, Cole BJ, Howard-Ashby M, Nebert DW, Scally M, Dean M, Epel D, Hahn ME, Stegeman JJ (prosinec 2006). „Chemická obrana: geny pro snímání a reakci na životní prostředí v genomu Strongylocentrotus purpuratus“. Vývojová biologie. 300 (1): 366–84. doi:10.1016 / j.ydbio.2006.08.066. PMC 3166225. PMID 17097629.
- ^ Rawal S, Kim JE, Coulombe R (prosinec 2010). „Aflatoxin B1 u drůbeže: toxikologie, metabolismus a prevence“. Výzkum ve veterinární vědě. 89 (3): 325–31. doi:10.1016 / j.rvsc.2010.04.011. PMID 20462619.
- ^ Rawal S, Coulombe RA (srpen 2011). „Metabolismus aflatoxinu B1 v mikrosomech krůtých jater: relativní role cytochromů P450 1A5 a 3A37“. Toxikologie a aplikovaná farmakologie. 254 (3): 349–54. doi:10.1016 / j.taap.2011.05.010. PMID 21616088.
- ^ McCart C, Ffrench-Constant RH (červen 2008). "Disekce na gen cytochromu P450 spojený s rezistencí na insekticidy Cyp6g1". Věda o ochraně proti škůdcům. 64 (6): 639–45. doi:10.1002 / ps.1567. PMID 18338338.
- ^ Ismail, Hanafy; O'Neill, Paul; Hong, David; Finn, Robert; Henderson, Colin; Wright, Aaron; Cravatt, Benjamin; Hemingway, Janet; Paine, Mark (3. prosince 2013). „Sondy založené na aktivitě pyrethroidů pro profilování aktivit cytochromu P450 spojených s interakcemi insekticidů“. PNAS. 110 (49): 19766–19771. Bibcode:2013PNAS..11019766I. doi:10.1073 / pnas.1320185110. PMC 3856776. PMID 24248381.
- ^ McLean KJ, Clift D, Lewis DG, Sabri M, Balding PR, Sutcliffe MJ, Leys D, Munro AW (květen 2006). "Převaha P450 v genomu Mycobacterium tuberculosis". Trendy v mikrobiologii. 14 (5): 220–8. doi:10.1016 / j.tim.2006.03.002. PMID 16581251.
- ^ Ikeda H, Ishikawa J, Hanamoto A, Shinose M, Kikuchi H, Shiba T, Sakaki Y, Hattori M, Omura S (květen 2003). "Kompletní sekvence genomu a komparativní analýza průmyslového mikroorganismu Streptomyces avermitilis". Přírodní biotechnologie. 21 (5): 526–31. doi:10.1038 / nbt820. PMID 12692562.
- ^ Leto, O'Keefe (1988). "Identifikace konstitutivních a herbicidem indukovatelných cytochromů P-450 v Streptomyces griseolus". Arch Microbiol. 149 (5): 406–12. doi:10.1007 / BF00425579. S2CID 35526991.
- ^ Moody SC, Zhao B, Lei L, Nelson DR, Mullins JG, Waterman MR, Kelly SL, Lamb DC (květen 2012). „Zkoumání zachování biosyntetické dráhy albaflavenonu a bifunkčnosti CYP170 ve streptomycetech“. FEBS Journal. 279 (9): 1640–9. doi:10.1111 / j.1742-4658.2011.08447.x. PMID 22151149.
- ^ Moody SC, Loveridge EJ (prosinec 2014). „Struktury, funkce a role CYP105 v zajímavé rodině enzymů ve Streptomyces“. Journal of Applied Microbiology. 117 (6): 1549–63. doi:10.1111 / jam.12662. PMC 4265290. PMID 25294646.
- ^ Narhi LO, Fulco AJ (červen 1986). „Charakterizace katalyticky soběstačné monooxygenázy cytochromu P-450 vyvolané barbituráty v Bacillus megaterium 119 000 daltonů“. The Journal of Biological Chemistry. 261 (16): 7160–9. PMID 3086309.
- ^ Girvan HM, Waltham TN, Neeli R, Collins HF, McLean KJ, Scrutton NS, Leys D, Munro AW (prosinec 2006). „Flavocytochrom P450 BM3 a původ fúzních druhů CYP102“. Transakce s biochemickou společností. 34 (Pt 6): 1173–7. doi:10.1042 / BST0341173. PMID 17073779.
- ^ Wright RL, Harris K, Solow B, White RH, Kennelly PJ (duben 1996). "Klonování potenciálního cytochromu P450 z archaeonu Sulfolobus solfataricus". FEBS Dopisy. 384 (3): 235–9. doi:10.1016/0014-5793(96)00322-5. PMID 8617361. S2CID 19579406.
- ^ Vanden Bossche H, Marichal P, Gorrens J, Coene MC (září 1990). "Biochemický základ pro aktivitu a selektivitu perorálních antifungálních léků". British Journal of Clinical Practice. Doplněk. 71: 41–6. PMID 2091733.
- ^ Schuler MA, Werck-Reichhart D (01.01.2003). "Funkční genomika P450". Roční přehled biologie rostlin. 54 (1): 629–67. doi:10.1146 / annurev.arplant.54.031902.134840. PMID 14503006.
- ^ Mizutani M, Sato F (březen 2011). "Neobvyklé reakce P450 v sekundárním metabolismu rostlin". Archivy biochemie a biofyziky. Mechanismy katalýzy P450. 507 (1): 194–203. doi:10.1016 / j.abb.2010.09.026. PMID 20920462.
- ^ Chefson A, Auclair K (říjen 2006). "Pokrok směrem ke snazšímu používání enzymů P450". Molekulární biosystémy. 2 (10): 462–9. doi:10.1039 / b607001a. PMID 17216026.
- ^ Chefson A, Zhao J, Auclair K (červen 2006). „Nahrazení přírodních kofaktorů vybranými dárci peroxidu vodíku nebo organickými peroxidy vede ke zlepšení aktivity pro CYP3A4 a CYP2D6.“ ChemBioChem. 7 (6): 916–9. doi:10.1002 / cbic.200600006. PMID 16671126. S2CID 39329433.
- ^ Chefson A, Auclair K (červenec 2007). „Aktivita CYP3A4 v přítomnosti organických kosolventů, iontových kapalin nebo organických rozpouštědel nemísitelných s vodou“. ChemBioChem. 8 (10): 1189–97. doi:10.1002 / cbic.200700128. PMID 17526062. S2CID 11845235.
- ^ Petros WP, Younis IR, Ford JN, Weed SA (říjen 2012). „Účinky kouření tabáku a nikotinu na léčbu rakoviny“. Farmakoterapie. 32 (10): 920–31. doi:10.1002 / j.1875-9114.2012.01117. PMC 3499669. PMID 23033231.
Další čtení
- Gelboin HV, Krausz K (březen 2006). „Monoklonální protilátky a multifunkční cytochrom P450: metabolismus léčiv jako paradigma“. Journal of Clinical Pharmacology. 46 (3): 353–72. doi:10.1177/0091270005285200. PMID 16490812.
- Gelboin HV, Krausz KW, Gonzalez FJ, Yang TJ (listopad 1999). „Inhibiční monoklonální protilátky proti lidským enzymům cytochromu P450: nová cesta k objevu léčiv“ (PDF). Trendy ve farmakologických vědách. 20 (11): 432–8. doi:10.1016 / S0165-6147 (99) 01382-6. PMID 10542439.
- „Metabolismus léků a karcinogenů zprostředkovaných cytochromem P450 pomocí monoklonálních protilátek“. home.ccr.cancer.gov. Citováno 2018-04-02.
- Krausz KW, Goldfarb I, Buters JT, Yang TJ, Gonzalez FJ, Gelboin HV (listopad 2001). „Monoklonální protilátky specifické a inhibiční vůči lidským cytochromům P450 2C8, 2C9 a 2C19“. Metabolismus a dispozice léků. 29 (11): 1410–23. PMID 11602516.
- Gonzalez FJ, Gelboin HV (1994). „Role lidských cytochromů P450 při metabolické aktivaci chemických karcinogenů a toxinů“. Recenze metabolismu drog. 26 (1–2): 165–83. doi:10.3109/03602539409029789. PMID 8082563.
externí odkazy
- Sigaroudi A, Vollbrecht H (2019). „tabulka farmakokinetických interakcí“. Sigaroudi & Vollbrecht.
- Preissner S (2010). "Databáze cytochromu P450". Výzkum nukleových kyselin.
- Degtyarenko K (01.01.2009). "Adresář systémů obsahujících P450". Mezinárodní centrum pro genetické inženýrství a biotechnologie. Archivovány od originál dne 2016-07-16. Citováno 2009-02-10.
- Estabrook RW (prosinec 2003). „Vášeň pro P450 (vzpomínky na ranou historii výzkumu cytochromu P450)“. Metabolismus a dispozice léků. 31 (12): 1461–73. doi:10.1124 / dmd.31.12.1461. PMID 14625342.
- Flockhart DA (2007). „Tabulka lékových interakcí cytochromu P450“. Indiana University - Purdue University Indianapolis. Citováno 2009-02-10.
- Sim SC (2008-09-04). „Výbor pro nomenklaturu alel lidského cytochromu P450 (CYP)“. Karolinska Institutet. Citováno 2009-02-10.
- Performance of P450 inhibition Studies The performance of in vitro cytochrome P450 inhibition studies including analysis of the data.