Alela - Allele
An alela (Spojené království: /ˈ…liːl/, /əˈliːl/; NÁS: /əˈliːl/; moderní formace z řečtiny ἄλλος állos, "jiný")[1][2][3] je jednou ze dvou nebo více forem dané gen varianta.[4] Např. the ABO krevní skupina je řízen Gen ABO který má šest společných alel. Téměř každý živý člověk fenotyp pro gen ABO je nějaká kombinace jen těchto šesti alel.[5][6] Alela je jednou ze dvou nebo více verzí stejného genu v stejné místo na chromozóm. Může také odkazovat na různé varianty sekvencí pro několik stovek párů bází nebo více oblastí genomu, které kódují protein. Alely mohou přicházet v různých extrémních velikostech. Při nejnižší možné velikosti může být alela a polymorfismus jednoho nukleotidu (SNP).[7] Na horním konci to může být až několik tisíc párů bází dlouho.[8][9] Většina alel vede k malé nebo žádné pozorovatelné změně ve funkci proteinu, který gen kóduje.
Někdy však mohou mít různé alely za následek odlišné pozorovatelnosti fenotypové vlastnosti, například různé pigmentace. Pozoruhodným příkladem toho je Gregora Mendela objev, že bílá a fialová květinová barva v hrášek rostliny byly výsledkem vlastností „čisté linie“, tj. jednoho genu se dvěma alelami.
Všechno mnohobuněčné organismy mít dvě sady chromozomy v určitém okamžiku v jejich životní cyklus; to znamená, že jsou diploidní. V tomto případě mohou být chromozomy spárováno. Každý chromozom v páru obsahuje stejné geny ve stejném pořadí a umístění po celé délce chromozomu. Pro daný gen, pokud dva chromozomy obsahují stejnou alelu, jsou to oni i organismus homozygotní s ohledem na tento gen. Pokud jsou alely odlišné, jsou to oni i organismus heterozygotní s ohledem na tento gen.
Etymologie
Slovo „alela“ je krátká forma alelomorf („jiná forma“, slovo vytvořené britskými genetiky William Bateson a Edith Rebecca Saunders ),[10][11] který byl používán v počátcích roku 2006 genetika popsat variantní formy a gen detekováno jako odlišné fenotypy. Vyplývá to z řecký předpona ἀλληλο-, alelo-, což znamená „vzájemný“, „vzájemný“ nebo „navzájem“, což samo o sobě souvisí s řeckým přídavným jménem ἄλλος, Allos (příbuzný s latinský alius), což znamená „jiné“.
Alely, které vedou k dominantním nebo recesivním fenotypům
V mnoha případech lze popsat genotypové interakce mezi dvěma alelami v lokusu dominantní nebo recesivní, podle kterého ze dvou homozygotních fenotypů heterozygot nejvíce se podobá. Pokud je heterozygot nerozeznatelný od jednoho z homozygotů, vyjádřená alela je ta, která vede k „dominantnímu“ fenotypu,[12] a o druhé alele se říká, že je „recesivní“. Stupeň a model dominance se u jednotlivých lokusů liší. Tento typ interakce poprvé formálně popsal Gregor Mendel. Mnoho znaků však této jednoduché kategorizaci vzdoruje a fenotypy jsou modelovány pomocí společná dominance a polygenní dědičnost.
Termín "divoký typ „alela se někdy používá k popisu alely, o které se předpokládá, že přispívá k typickému fenotypovému charakteru, jak je vidět u„ divokých “populací organismů, jako jsou ovocné mušky (Drosophila melanogaster ). Taková alela „divokého typu“ byla historicky považována za vedoucí k dominantnímu (silnému - vždy vyjádřenému), běžnému a normálnímu fenotypu, na rozdíl od „mutant „alely, které vedou k recesivním, vzácným a často škodlivým fenotypům. Dříve se předpokládalo, že většina jedinců byla homozygotní pro alelu„ divokého typu “nanejvýš v genových lokusech a že jakákoli alternativní„ mutantní “alela byla nalezena v homozygotní formě v malá menšina „postižených“ jedinců, často jako genetické nemoci, a častěji v heterozygotní formě v „dopravci „pro mutovanou alelu. Nyní se oceňuje, že většina nebo všechny genové lokusy jsou vysoce polymorfní, s několika alelami, jejichž frekvence se liší od populace k populaci, a že velká genetická variace je skryta ve formě alel, které produkují zjevné fenotypové rozdíly.
Více alel




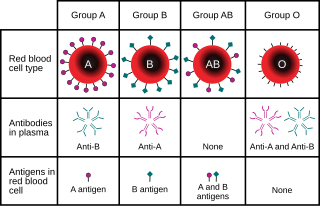
Populace nebo druh organizmů obvykle zahrnuje více alel v každém místě mezi různými jedinci. Alelická variace na místě je měřitelná jako počet alel (polymorfismus ) přítomný nebo podíl heterozygotů v populaci. A nulová alela je genová varianta, která postrádá normální funkci genu, protože buď není exprimována, nebo je exprimovaný protein neaktivní.
Například v genovém lokusu pro ABO krevní skupina uhlohydrát antigeny u lidí[13] klasická genetika rozeznává tři alely, IA, JáB, a i, které určují kompatibilitu krevní transfuze. Každý jedinec má jednu ze šesti možných genotypy (JáAJáA, JáAjá, jáBJáB, JáBjá, jáAJáB, a ii) které produkují jeden ze čtyř možných fenotypy: "Typ A" (vyrobeno I.AJáA homozygot a jáAi heterozygotní genotypy), "Typ B" (produkovaný I.BJáB homozygot a jáBi heterozygotní genotypy), „typ AB“ produkovaný I.AJáB heterozygotní genotyp a „typ O“ produkovaný ii homozygotním genotypem. (Nyní je známo, že každá z alel A, B a O je ve skutečnosti třída více alel s různými sekvencemi DNA, které produkují proteiny se stejnými vlastnostmi: na lokusu ABO je známo více než 70 alel.[14] Jednotlivcem s krví typu „A“ tedy může být heterozygot AO, homozygot AA nebo heterozygot AA se dvěma různými alelami „A“.)
Četnosti genotypu
Frekvenci alel v diploidní populaci lze použít k předpovědi frekvencí odpovídajících genotypů (viz Hardy-Weinbergův princip ). Pro jednoduchý model se dvěma alelami;


kde p je frekvence jedné alely a q je frekvence alternativní alely, která se nutně rovná jednotě. Pak, p2 je zlomek populace homozygotní pro první alelu, 2pq je zlomek heterozygotů a q2 je frakce homozygotní pro alternativní alelu. Pokud je první alela dominantní vůči druhé, pak zlomek populace, která bude vykazovat dominantní fenotyp, je p2 + 2pqa zlomek s recesivním fenotypem je q2.
Se třemi alelami:
- a


V případě více alel na diploidním místě je počet možných genotypů (G) s počtem alel (a) dán výrazem:

Alelická dominance u genetických poruch
Počet genetické poruchy jsou způsobeny, když jedinec zdědí dvě recesivní alely pro znak jednoho genu. Mezi recesivní genetické poruchy patří albinismus, cystická fibróza, galaktosémie, fenylketonurie (PKU) a Tay – Sachsova choroba. Jiné poruchy jsou také způsobeny recesivními alelami, ale protože genový lokus je umístěn na chromozomu X, takže muži mají pouze jednu kopii (tj. hemizygotní ), jsou častější u mužů než u žen. Mezi příklady patří červeno-zelená barvoslepost a syndrom křehkého X.
Jiné poruchy, jako např Huntingtonova choroba, nastanou, když jedinec zdědí pouze jednu dominantní alelu.
Epialleles
Zatímco dědičné vlastnosti jsou typicky studovány z hlediska genetických alel, epigenetický známky jako Methylace DNA mohou být zděděny ve specifických genomových oblastech u určitých druhů, což je proces nazývaný transgenerační epigenetická dědičnost. Termín epiallele se používá k rozlišení těchto dědičných značek od tradičních alel, které jsou definovány pomocí nukleotidová sekvence.[15] Specifická třída epiallele, metastabilní epiallely, byl objeven u myší a u lidí, pro které je charakteristické stochastické (pravděpodobnostní) vytvoření epigenetického stavu, který lze mitoticky zdědit.[16][17]
Viz také
Odkazy a poznámky
- ^ "Allele | Význam Allele od společnosti Lexico". Lexico slovníky | Angličtina. Citováno 7. ledna 2020.
- ^ "alela podstatné jméno - definice, obrázky, výslovnost a poznámky k použití". Oxford Advanced Learner's Dictionary. Citováno 29. října 2017.
- ^ „alela význam v anglickém slovníku Cambridge“. Dictionary.cambridge.org. Citováno 29. října 2017.
- ^ „Co jsou varianty, alely a haplotypy? | Lidská genetická variace“. Citováno 16. listopadu 2020.
- ^ Seltsam A, Hallensleben M, Kollmann A, Blasczyk R (říjen 2003). „Povaha rozmanitosti a diverzifikace v místě ABO“. Krev. 102 (8): 3035–42. doi:10.1182 / krev-2003-03-0955. PMID 12829588.
- ^ Ogasawara K, Bannai M, Saitou N, Yabe R, Nakata K, Takenaka M, Fujisawa K, Uchikawa M, Ishikawa Y, Juji T, Tokunaga K (červen 1996). „Rozsáhlý polymorfismus genu krevní skupiny ABO: tři hlavní linie alel pro běžné fenotypy ABO“. Genetika člověka. 97 (6): 777–83. doi:10.1007 / BF02346189. PMID 8641696.
- ^ Smigielski, Elizabeth M .; Sirotkin, Karl; Ward, Minghong; Sherry, Stephen T. (1. ledna 2000). „dbSNP: databáze jednonukleotidových polymorfismů“. Výzkum nukleových kyselin. 28 (1): 352–355. doi:10.1093 / nar / 28.1.352. ISSN 0305-1048. PMC 102496. PMID 10592272.
- ^ Elston, Robert; Satagopan, Jaya; Sun, Shuying (2012). "Genetická terminologie". Statistická genetika člověka. Metody v molekulární biologii (Clifton, N.J.). 850. s. 1–9. doi:10.1007/978-1-61779-555-8_1. ISBN 978-1-61779-554-1. ISSN 1064-3745. PMC 4450815. PMID 22307690.
- ^ „Jaký účinek mají varianty v kódujících oblastech?“. Vlak EMBL-EBI online. 2. května 2019. Citováno 14. listopadu 2019.
- ^ Craft, Jude (2013). „Geny a genetika: jazyk vědeckých objevů“. Geny a genetika. Oxfordský anglický slovník. Citováno 14. ledna 2016.
- ^ Bateson, W. a Saunders, E. R. (1902) „Fakta dědičnosti ve světle Mendelova objevu.“ Zprávy Evolučnímu výboru Královské společnosti, I. 125–160
- ^ Hartl, Daniel L .; Elizabeth W. Jones (2005). Esenciální genetika: Genomická perspektiva (4. vydání). Vydavatelé Jones & Bartlett. p. 600. ISBN 978-0-7637-3527-2.
- ^ Victor A. McKusick; Cassandra L. Kniffin; Paul J. Converse; Ada Hamosh (10. listopadu 2009). "ABO glykosyltransferáza; ABO". Online Mendelian Inheritance in Man. Národní lékařská knihovna. Archivováno z původního dne 24. září 2008. Citováno 24. března 2010.
- ^ Yip SP (leden 2002). "Variace sekvence na lidském ABO lokusu". Annals of Human Genetics. 66 (1): 1–27. doi:10.1017 / S0003480001008995. PMID 12014997.
- ^ Daxinger, Lucia; Whitelaw, Emma (31. ledna 2012). „Pochopení transgenerační epigenetické dědičnosti prostřednictvím gamet u savců“. Genetika hodnocení přírody. 13 (3): 153–62. doi:10.1038 / nrg3188. PMID 22290458.
- ^ Rakyan, Vardhman K; Blewitt, Marnie E; Druker, Riki; Preis, Jost I; Whitelaw, Emma (červenec 2002). "Metastabilní epiallely u savců". Trendy v genetice. 18 (7): 348–351. doi:10.1016 / S0168-9525 (02) 02709-9. PMID 12127774.
- ^ Waterland, RA; Dolinoy, DC; Lin, JR; Smith, CA; Shi, X; Tahiliani, KG (září 2006). "Mateřské methylové doplňky zvyšují metylaci DNA potomků u Axin Fused". Genesis. 44 (9): 401–6. doi:10.1002 / dvg.20230. PMID 16868943.