Cévní endoteliální růstový faktor - Vascular endothelial growth factor
Cévní endoteliální růstový faktor (VEGF), původně známý jako faktor vaskulární permeability (VPF),[1] je signální protein produkovaný buňkami, který stimuluje tvorbu krevních cév. Konkrétně VEGF je podskupina růstové faktory, růstový faktor odvozený z krevních destiček rodina cystinový uzel růstové faktory. Jsou důležitou signalizací bílkoviny podílí se na obou vaskulogeneze (dále jen de novo tvorba embrya oběhový systém ) a angiogeneze (růst krevních cév z již existující vaskulatury).
Je součástí systému, který obnovuje přívod kyslíku do tkání, když je krevní oběh nedostatečný, například v hypoxických podmínkách.[2] Sérová koncentrace VEGF je vysoká bronchiální astma a diabetes mellitus.[3]Normální funkcí VEGF je vytváření nových krevních cév během embryonální vývoj, nové krevní cévy po poranění, svaly po cvičení a nové cévy (kolaterální oběh ) obejít zablokované cévy. Může přispět k onemocnění. Pevné rakoviny nemohou růst nad omezenou velikost bez adekvátního přívodu krve; rakoviny, které mohou exprimovat VEGF, jsou schopné růst a metastázovat. Nadměrná exprese VEGF může způsobit vaskulární onemocnění v sítnice oka a dalších částí těla. Drogy jako aflibercept, bevacizumab, ranibizumab, a pegaptanib může inhibovat VEGF a kontrolovat nebo zpomalit tato onemocnění.
Dějiny
V roce 1970 Judah Folkman et al. popsal faktor vylučovaný nádory způsobujícími angiogenisis a nazval jej faktor angiogeneze nádoru.[4] V roce 1983 Senger et al. identifikoval a faktor vaskulární permeability vylučované nádory u morčat a křečků.[1] V roce 1989 Ferrara a Henzel popsali identický faktor v bovinních hypofyzárních folikulárních buňkách, které purifikovali, klonovali a pojmenovali VEGF. [5] Podobný alternativní sestřih VEGF objevil Tischer et al. v roce 1991.[6] V letech 1996 až 1997 Christinger a De Vos získali krystalovou strukturu VEGF, nejprve v rozlišení 2,5 Å a později v 1,9 Å.[7][8][9]
Fms podobná tyrosinkináza-1 (flt-1) byla Ferrarou prokázána jako VEGF receptor et al. v roce 1992.[10] The receptor kinázové inzertní domény (KDR) bylo prokázáno, že je VEGF receptorem Terman et al. v roce 1992 také.[11] V roce 1998 neuropilin 1 a neuropilin 2 Bylo prokázáno, že působí jako receptory VEGF.[12]
Klasifikace

U savců zahrnuje rodina VEGF pět členů: VEGF-A, růstový faktor placenty (PGF ), VEGF-B, VEGF-C a VEGF-D. Poslední členové byli objeveni po VEGF-A; před jejich objevem byl VEGF-A znám jako VEGF. Řada proteinů souvisejících s VEGF kódovaných viry (VEGF-E ) a v jedu některých hadů (VEGF-F ) byly také objeveny.
| Typ | Funkce |
|---|---|
| VEGF-A |
|
| VEGF-B | Embryonální angiogeneze (tkáň myokardu, přesněji řečeno)[13] |
| VEGF-C | Lymfangiogeneze[Citace je zapotřebí ] |
| VEGF-D | Nezbytné pro vývoj lymfatické vaskulatury obklopující plicní bronchioly[Citace je zapotřebí ] |
| PlGF | Důležité pro vaskulogenezi, také potřebné pro angiogenezi během ischemie, zánětu, hojení ran a rakoviny.[Citace je zapotřebí ] |
Aktivita VEGF-A, jak již název napovídá, byla studována hlavně na buňkách cév endotel, i když to má účinky na řadu dalších typů buněk (např. stimulace monocyt /makrofág migrace, neurony, rakovinné buňky, epiteliální buňky ledvin). In vitro bylo prokázáno, že VEGF-A stimuluje endoteliální buňky mitogeneze a migrace buněk. VEGF-A je také vazodilatátor a zvyšuje mikrovaskulární permeabilitu a původně byl označován jako faktor vaskulární permeability.
Izoformy

Existuje několik izoforem VEGF-A, které jsou výsledkem alternativní sestřih z mRNA od jednoho, 8-exon VEGFA gen. Ty jsou rozděleny do dvou skupin, na které se odkazuje podle jejich koncového místa sestřihu exonu (exonu 8): proximální místo sestřihu (označené VEGF)xxx) nebo místo distálního sestřihu (VEGFxxxb). Kromě toho alternativní sestřih exonu 6 a 7 mění jejich heparin - vazebná afinita a počet aminokyselin (u lidí: VEGF121, VEGF121b, VEGF145, VEGF165, VEGF165b, VEGF189, VEGF206; ortology hlodavců těchto proteinů obsahují o jednu aminokyselinu méně). Tyto domény mají důležité funkční důsledky pro varianty sestřihu VEGF, protože terminální místo sestřihu (exon 8) určuje, zda jsou proteiny pro-angiogenní (místo blízkého sestřihu, exprimované během angiogeneze) nebo antiangiogenní (místo distálního sestřihu, vyjádřené v normálním papírové kapesníky). Navíc zahrnutí nebo vyloučení exonů 6 a 7 zprostředkovává interakce s heparan sulfát proteoglykany (HSPG) a neuropilin ko-receptory na povrchu buněk, což zvyšuje jejich schopnost vázat se a aktivovat VEGF receptory (VEGFR).[14] Nedávno se ukázalo, že VEGF-C je důležitým induktorem neurogeneze v myší subventrikulární zóně, aniž by vyvolával angiogenní účinky.[15]
Mechanismus
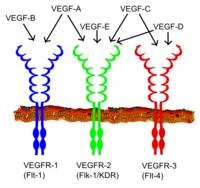
Všichni členové rodiny VEGF stimulují buněčné odpovědi vazbou na tyrosinkináza receptory ( VEGFR ) na povrchu buňky, což způsobí jejich dimerizaci a aktivaci skrz transfosforylace, i když na různá místa, časy a rozsahy. Receptory VEGF mají extracelulární část skládající se ze 7 domén podobných imunoglobulinům, jednu transmembránovou překlenovací oblast a intracelulární část obsahující štěpení tyrosinkináza doména. VEGF-A se váže na VEGFR-1 (Flt-1 ) a VEGFR-2 (KDR / Flk-1 ).[17] Zdá se, že VEGFR-2 zprostředkovává téměř všechny známé buněčné odpovědi na VEGF. Funkce VEGFR-1 je méně dobře definovaná, i když se předpokládá, že moduluje signalizaci VEGFR-2.[18] Další funkcí VEGFR-1 může být působit jako fiktivní / návnadový receptor, oddělovat VEGF od vazby VEGFR-2 (to se jeví jako zvláště důležité během vaskulogeneze v embryu). VEGF-C a VEGF-D, ale ne VEGF-A, jsou ligandy pro třetí receptor (VEGFR-3 / Flt4 ), který zprostředkovává lymfangiogeneze. Receptor (VEGFR3) je místem vazby hlavních ligandů (VEGFC a VEGFD), které zprostředkovávají trvalé působení a funkci ligandů na cílové buňky. Vaskulární endoteliální růstový faktor-C může stimulovat lymfangiogenezi (prostřednictvím VEGFR3) a angiogenezi prostřednictvím VEGFR2. Vaskulární endoteliální růstový faktor-R3 byl detekován v lymfatických endoteliálních buňkách u CL mnoha druhů, skotu, buvolů a primátů.[19]
Kromě vazby na VEGFR, VEGF se váže na receptorové komplexy skládající se z obou neuropiliny a VEGFR. Tento receptorový komplex zvýšil v VFF signální aktivitu endoteliální buňky (cévy ).[20][21] Neuropiliny (NRP) jsou pleitrofický receptory, a proto mohou jiné molekuly interferovat se signalizací komplexů receptorů NRP / VEGFR. Například třída 3 semafory soutěžit s VEGF165 pro vazbu NRP a mohl by tedy regulovat VEGF zprostředkovaný angiogeneze.[22]
Výraz
Produkce VEGF-A může být indukována v buňce, která nedostává dostatek kyslík.[17] Když má buňka nedostatek kyslíku, produkuje HIF, hypoxií indukovatelný faktor, transkripční faktor. HIF mimo jiné stimuluje uvolňování VEGF-A (včetně modulace erytropoézy). Cirkulující VEGF-A se poté váže na receptory VEGF na endoteliálních buňkách a spouští a tyrosinkináza cesta vedoucí k angiogenezi.[je zapotřebí objasnění ] Vyjádření angiopoetin-2 v nepřítomnosti VEGF vede k smrti endoteliálních buněk a vaskulární regresi.[23] Naopak německá studie hotová in vivo zjistili, že koncentrace VEGF skutečně poklesly po 25% snížení příjmu kyslíku po dobu 30 minut.[24] HIF1 alfa a HIF1 beta se neustále vyrábějí, ale HIF1 alfa je vysoce O2 labilní, takže za aerobních podmínek je degradován. Když se buňka stane hypoxickou, HIF1 alfa přetrvává a komplex HIF1alfa / beta stimuluje uvolňování VEGF. kombinované použití mikrovezikul a 5-FU mělo za následek zvýšenou chemosenzitivitu buněk karcinomu dlaždicových buněk více než použití samotného 5-FU nebo mikrovezikul. Kromě toho byla snížená regulace exprese genu VEGF spojena se sníženou expresí genu CD1[25]
Klinický význam
V nemoci
VEGF-A a odpovídající receptory jsou po traumatickém poranění rychle up-regulovány centrální nervový systém (CNS). VEGF-A je vysoce exprimován v akutních a subakutních stádiích poškození CNS, ale exprese proteinu v průběhu času klesá. Toto časové rozpětí exprese VEGF-A odpovídá endogennímu opětovná vaskularizace kapacita po zranění.[22] To by naznačovalo, že VEGF-A / VEGF165 může být použit jako cíl k podpoře angiogeneze po traumatickém poranění CNS. Existují však protichůdné vědecké zprávy o účincích léčby VEGF-A v modelech poranění CNS.[22]
VEGF-A je implikován se špatnou prognózou v rakovina prsu. Četné studie ukazují snížené celkové přežití a přežití bez onemocnění u těch nádorů, které nadměrně exprimují VEGF. Nadměrná exprese VEGF-A může být časným krokem v procesu metastáza, krok, který je zapojen do "angiogenního" přepínače. Ačkoli VEGF-A koreluje se špatným přežitím, jeho přesný mechanismus účinku při progresi nádorů zůstává nejasný[Citace je zapotřebí ].
VEGF-A je také vydáván v revmatoidní artritida v reakci na TNF-α, zvýšení propustnosti a otoku endotelu a také stimulace angiogeneze (tvorba kapilár)[Citace je zapotřebí ].
VEGF-A je také důležitý v diabetická retinopatie (DR). Mikrocirkulační problémy na sítnici lidí s cukrovka může způsobit ischemii sítnice, která má za následek uvolnění VEGF-A a změnu rovnováhy proangiogenního VEGFxxx izoformy nad normálně exprimovaným VEGFxxxb izoformy. VEGFxxx pak může způsobit tvorbu nových krevních cév v sítnici a jinde v oku, což ohlašuje změny, které mohou ohrozit zrak.
VEGF-A hraje roli v patologii nemoci mokré formy Věkem podmíněné makulární degenerace (AMD), což je hlavní příčina slepoty pro seniory v průmyslovém světě. Cévní patologie AMD sdílí určité podobnosti s diabetickou retinopatií, i když příčina onemocnění a typický zdroj neovaskularizace se u obou onemocnění liší.
Sérové hladiny VEGF-D jsou významně zvýšené u pacientů s angiosarkom.[26]
Po uvolnění může VEGF-A vyvolat několik odpovědí. Může to způsobit buňka přežít, přestěhovat se nebo dále rozlišovat. Proto je VEGF potenciálním cílem pro léčbu rakovina. První anti-VEGF lék, a monoklonální protilátka pojmenovaný bevacizumab, byl schválen v roce 2004. Přibližně 10–15% pacientů má prospěch z léčby bevacizumabem; biomarkery účinnosti bevacizumabu však dosud nejsou známy.
Současné studie ukazují, že VEGF nejsou jedinými promotory angiogeneze. Zejména, FGF2 a HGF jsou silné angiogenní faktory.
Bylo zjištěno, že pacienti trpící plicním emfyzémem mají snížené hladiny VEGF v plicních tepnách.
Bylo také prokázáno, že VEGF-D je nadměrně exprimován v lymfangioleiomyomatóza a v současné době se používá jako diagnostický biomarker při léčbě tohoto vzácného onemocnění [27].
V ledviny, zvýšená exprese VEGF-A v glomeruli přímo způsobuje glomerulární hypertrofii, která je spojena s proteinurií.[28]
Změny VEGF mohou předvídat časný nástup preeklampsie.[29]
Genové terapie pro refrakterní anginu pectoris stanovit expresi VEGF v epikardiálních buňkách k podpoře angiogeneze.[30]
Viz také
- Proteázy v angiogenezi
- Withaferin A, silný inhibitor angiogeneze
Reference
- ^ A b Senger, DR; Galli, SJ; Dvořák, AM; Perruzzi, CA; Harvey, VS; Dvořák, HF (25. února 1983). „Nádorové buňky vylučují faktor vaskulární permeability, který podporuje akumulaci ascitické tekutiny“. Věda. 219 (4587): 983–5. Bibcode:1983Sci ... 219..983S. doi:10.1126 / science.6823562. PMID 6823562.
- ^ Palmer, Biff F .; Clegg, Deborah J. (2014). "Detekce kyslíku a metabolická homeostáza". Molekulární a buněčná endokrinologie. 397 (1–2): 51–57. doi:10.1016 / j.mce.2014.08.001. PMID 25132648.
- ^ Cooper, Mark; Dimitria Vranes; Sherif Youssef; Steven A. Stacker; Alison J. Cox; Bishoy Rizkalla; David J. Casley; Leon A. Bach; Darren J. Kelly; Richard E. Gilbert (listopad 1999). „Zvýšená renální exprese vaskulárního endoteliálního růstového faktoru (VEGF) a jeho receptoru VEGFR-2 u experimentálního diabetu“ (PDF). Cukrovka. 48 (11): 2229–2239. doi:10.2337 / diabetes.48.11.2229. PMID 10535459. Citováno 6. listopadu 2013.
- ^ Folkman, J (1. února 1971). „Izolace nádorového faktoru odpovědného za angiogenezi“. Journal of Experimental Medicine. 133 (2): 275–288. doi:10.1084 / jem.133.2.275. PMC 2138906. PMID 4332371.
- ^ Ferrara, N; Henzel, WJ (15. června 1989). „Hypofyzární folikulární buňky vylučují nový růstový faktor vázající se na heparin specifický pro vaskulární endoteliální buňky“. Sdělení o biochemickém a biofyzikálním výzkumu. 161 (2): 851–8. doi:10.1016 / 0006-291x (89) 92678-8. PMID 2735925.
- ^ Tischer, E; Mitchell, R; Hartman, T; Silva, M; Gospodarowicz, D; Fiddes, JC; Abraham, JA (25. června 1991). „Lidský gen pro vaskulární endoteliální růstový faktor. Více proteinových forem je kódováno alternativním sestřihem exonu“. The Journal of Biological Chemistry. 266 (18): 11947–54. PMID 1711045.
- ^ Christinger, Hans W .; Muller, Yves A .; Berleau, Lea T .; Keyt, Bruce A .; Cunningham, Brian C .; Ferrara, Napoleone; de Vos, Abraham M. (listopad 1996). "Krystalizace receptorové vazebné domény vaskulárního endoteliálního růstového faktoru". Proteiny: struktura, funkce a genetika. 26 (3): 353–357. doi:10.1002 / (SICI) 1097-0134 (199611) 26: 3 <353 :: AID-PROT9> 3.0.CO; 2-E. PMID 8953654.
- ^ Muller, YA; Li, B; Christinger, HW; Wells, JA; Cunningham, BC; de Vos, AM (8. července 1997). „Vaskulární endoteliální růstový faktor: krystalová struktura a funkční mapování vazebného místa receptoru kinázové domény“. Sborník Národní akademie věd Spojených států amerických. 94 (14): 7192–7. Bibcode:1997PNAS ... 94,7192M. doi:10.1073 / pnas.94.14.7192. PMC 23789. PMID 9207067.
- ^ Muller, YA; Christinger, HW; Keyt, BA; de Vos, AM (15. října 1997). „Krystalová struktura vaskulárního endoteliálního růstového faktoru (VEGF) se upravila na rozlišení 1,93 A: flexibilita vícenásobných kopií a vazba na receptory“. Struktura. 5 (10): 1325–38. doi:10.1016 / s0969-2126 (97) 00284-0. PMID 9351807.
- ^ de Vries, C; Escobedo, JA; Ueno, H; Houck, K; Ferrara, N; Williams, LT (21. února 1992). "FMS-jako tyrosinkináza, receptor pro vaskulární endoteliální růstový faktor". Věda. 255 (5047): 989–91. Bibcode:1992Sci ... 255..989D. doi:10.1126 / science.1312256. PMID 1312256.
- ^ Terman, BI; Dougher-Vermazen, M; Carrion, ME; Dimitrov, D; Armellino, DC; Gospodarowicz, D; Böhlen, P (30. září 1992). "Identifikace KDR tyrosinkinázy jako receptoru pro růstový faktor vaskulárních endoteliálních buněk". Sdělení o biochemickém a biofyzikálním výzkumu. 187 (3): 1579–86. doi:10.1016 / 0006-291x (92) 90483-2. PMID 1417831.
- ^ Soker, S; Takashima, S; Miao, HQ; Neufeld, G; Klagsbrun, M (20. března 1998). „Neuropilin-1 je exprimován endoteliálními a nádorovými buňkami jako izoformně specifický receptor pro vaskulární endoteliální růstový faktor.“ Buňka. 92 (6): 735–45. doi:10.1016 / s0092-8674 (00) 81402-6. PMID 9529250.
- ^ Claesson-Welsh, L. (20. srpna 2008). „VEGF-B převzatý do našich srdcí: Specifický účinek VEGF-B na ischemii myokardu“. Arterioskleróza, trombóza a vaskulární biologie. 28 (9): 1575–1576. doi:10.1161 / ATVBAHA.108.170878. PMID 18716319.
- ^ Cébe Suarez, S; Pieren, M; Cariolato, L; Arn, S; Hoffmann, U; Bogucki, A; Manlius, C; Wood, J; Ballmer-Hofer, K (2006). „Varianta sestřihu VEGF-A defektní pro vazbu heparan sulfátu a neuropilinu-1 vykazuje oslabenou signalizaci prostřednictvím VEGFR-2“ (PDF). Buněčné a molekulární biologické vědy. 63 (17): 2067–2077. doi:10.1007 / s00018-006-6254-9. PMID 16909199.
- ^ Shin, Y. J .; Choi, J. S .; et al. (2010). „Indukce mRNA receptoru 3 vaskulárního endoteliálního růstového faktoru v gliových buňkách po fokální mozkové ischemii u potkanů“. J. Neuroimmunol. 229 (1–2): 81–90. doi:10.1016 / j.jneuroim.2010.07.008. PMID 20692049.
- ^ cancerpublications.com.
- ^ A b Holmes, Katherine; Roberts, Owain Ll; Thomas, Angharad M .; Kříž, Michael J. (2007). "Vaskulární endoteliální růstový faktor receptor-2: Struktura, funkce, intracelulární signalizace a terapeutická inhibice". Mobilní signalizace. 19 (10): 2003–12. doi:10.1016 / j.cellsig.2007.05.013. PMID 17658244.
- ^ Karkkainen, M.J .; Petrova, T.V. (2000). "Receptory vaskulárního endoteliálního růstového faktoru v regulaci angiogeneze a lymfangiogeneze". Onkogen. 19 (49): 5598–5605. doi:10.1038 / sj.onc.1203855. PMID 11114740.
- ^ Ali, Ibne; et al. (2013). „Exprese a lokalizace lokálně produkovaných růstových faktorů regulujících lymfangiogenezi v různých fázích estrálního cyklu v žlutém těle buvola“ (Bubalus bubalis). Theriogenologie. 81 (3): 428–436. doi:10.1016 / j.theriogenology.2013.10.017. PMID 24246422.
- ^ Soker, S .; Takashima, S .; Miao, H. Q .; Neufeld, G .; Klagsbrun, M. (1998). „Neuropilin-1 je exprimován endoteliálními a nádorovými buňkami jako izoformně specifický receptor pro vaskulární endoteliální růstový faktor.“ Buňka. 92 (6): 735–745. doi:10.1016 / s0092-8674 (00) 81402-6. ISSN 0092-8674. PMID 9529250.
- ^ Herzog, B; Pellet-Many, C; Britton, G; Hartzoulakis, B; Zachary, I. C. (2011). „Vazba VEGF na NRP1 je nezbytná pro VEGF stimulaci migrace endoteliálních buněk, tvorbu komplexu mezi NRP1 a VEGFR2 a signalizaci pomocí fosforylace FAK Tyr407“. Molekulární biologie buňky. 22 (15): 2766–2776. doi:10,1091 / mbc.E09-12-1061. ISSN 1939-4586. PMC 3145551. PMID 21653826.
- ^ A b C Mecollari, V; Nieuwenhuis, B; Verhaagen, J (2014). „Pohled na roli semaforinové signalizace třídy III při traumatu centrální nervové soustavy“. Hranice v buněčné neurovědě. 8: 328. doi:10.3389 / fncel.2014.00328. PMC 4209881. PMID 25386118.
- ^ Harmey, Judith (2004). VEGF a rakovina. Georgetown, Tex: Landes Bioscience / Eurekah.com New York, N.Y. Kluwer Academic / Plenum Publishers. ISBN 978-0-306-47988-5.
- ^ Oltmanns, K. M .; Gehring, H; Rudolf, S; Schultes, B; Hackenberg, C; Schweiger, U; Born, J; Fehm, H.L .; Peters, A (2006). „Akutní hypoxie snižuje plazmatickou koncentraci VEGF u zdravých lidí“. AJP: Endokrinologie a metabolismus. 290 (3): E434–9. doi:10.1152 / ajpendo.00508.2004. PMID 16219663.
- ^ Ghada A. Abd El Latif, Iman M. Aboushady a Dina Sabry Snížení exprese genů VEGF a cyklin D1 zvyšuje chemosenzitivitu buněk lidského karcinomu dlaždicových buněk na mikrovezikuly odvozené od 5-fluorouracilu a / nebo mezenchymálních kmenových buněk E.D.J. Sv. 65, 2, str. 1217-1228; 2019. DOI: 10.21608 / EDJ.2019.72197
- ^ Amo, Y .; Masuzawa, M .; Hamada, Y .; Katsuoka, K. (2004). "Sérové koncentrace vaskulárního endoteliálního růstového faktoru-D u pacientů s angiosarkomem". British Journal of Dermatology. 150 (1): 160–1. doi:10.1111 / j.1365-2133.2004.05751.x. PMID 14746640.
- ^ Young, L. R .; Inoue, Y .; McCormack, F. X. (2008). „Diagnostický potenciál séra VEGF-D pro lymfangioleiomyomatózu“. The New England Journal of Medicine. 358 (2): 199–200. doi:10.1056 / NEJMc0707517. PMC 3804557. PMID 18184970.
- ^ Liu, E .; Morimoto, M .; Kitajima, S .; Koike, T .; Yu, Y .; Shiiki, H .; Nagata, M .; Watanabe, T .; Fan, J. (2007). „Zvýšená exprese růstového faktoru vaskulárního endotelu v ledvinách vede k progresivnímu narušení glomerulárních funkcí“. Časopis Americké nefrologické společnosti. 18 (7): 2094–104. doi:10.1681 / ASN.2006010075. PMID 17554151.
- ^ Andraweera, P. H .; Dekker, G. A .; Roberts, C. T. (2012). „Rodina vaskulárních endoteliálních růstových faktorů v nepříznivých výsledcích těhotenství“. Aktualizace lidské reprodukce. 18 (4): 436–457. doi:10.1093 / humupd / dms011. PMID 22495259.
- ^ „Genová terapie pro refrakterní anginu pectoris“. Kontext genomu. 16. října 2019. Citováno 16. října 2019.
{kind=link}
Další čtení
- Bengoetxea H, Argandoña EG, Lafuente JV (2008). „Účinky vizuálního zážitku na expresi růstového faktoru vaskulárního endotelu během postnatálního vývoje zrakové kůry krysy“. Mozková kůra. 18 (7): 1630–39. doi:10.1093 / cercor / bhm190. PMC 2430152. PMID 17986606.
- Zan L, Wu H, Jiang J, Zhao S, Song Y, Teng G, Li H, Jia Y, Zhou M, Zhang X, Qi J, Wang J (2011). „Časový profil Src, SSeCKS a angiogenních faktorů po fokální mozkové ischemii: korelace s angiogenezí a mozkovým edémem“. Neurochem. Int. 58 (8): 872–9. doi:10.1016 / j.neuint.2011.02.014. PMC 3100427. PMID 21334414.
- Zan L, Zhang X, Xi Y, Wu H, Song Y, Teng G, Li H, Qi J, Wang J (2014). "Src reguluje angiogenní faktory a vaskulární permeabilitu po fokální mozkové ischemii-reperfúzi". Neurovědy. 262: 118–28. doi:10.1016 / j.neuroscience.2013.12.060. PMC 3943922. PMID 24412374.
- Wang J, Fu X, Jiang C, Yu L, Wang M, Han W, Liu L, Wang J (2014). „Transplantace mononukleárních buněk kostní dřeně podporuje terapeutickou angiogenezi prostřednictvím upregulace signální dráhy VEGF-VEGFR2 na modelu krysí vaskulární demence“. Chovat se. Brain Res. 265: 171–80. doi:10.1016 / j.bbr.2014.02.033. PMC 4000455. PMID 24589546.
- Ferrara N, Gerber HP (2002). "Role vaskulárního endoteliálního růstového faktoru v angiogenezi". Acta Haematol. 106 (4): 148–56. doi:10.1159/000046610. PMID 11815711.
- Orpana A, Salven P (2003). "Angiogenní a lymfangiogenní molekuly v hematologických malignitách". Leuku. Lymfom. 43 (2): 219–24. doi:10.1080/10428190290005964. PMID 11999550.
- Afuwape AO, Kiriakidis S, Paleolog EM (2003). „Role angiogenní molekuly VEGF v patogenezi revmatoidní artritidy“. Histol. Histopathol. 17 (3): 961–72. PMID 12168808.
- de Bont ES, Neefjes VM, Rosati S a kol. (2003). „Nová tvorba cév a aberantní signalizace VEGF / VEGFR u akutní leukémie: má to význam?“. Leuku. Lymfom. 43 (10): 1901–9. doi:10.1080/1042819021000015844. PMID 12481883.
- Ria R, Roccaro AM, Merchionne F a kol. (2003). "Vaskulární endoteliální růstový faktor a jeho receptory v mnohočetném myelomu". Leukémie. 17 (10): 1961–6. doi:10.1038 / sj.leu.2403076. PMID 14513045.
- Caldwell RB, Bartoli M, Behzadian MA a kol. (2004). „Vaskulární endoteliální růstový faktor a diabetická retinopatie: patofyziologické mechanismy a perspektivy léčby“. Diabetes Metab. Res. Rev. 19 (6): 442–55. doi:10,1002 / dmrr.415. PMID 14648803.
- Patan, Sybill (2004). "Vasculogenesis and Angiogenesis". Angiogeneze u mozkových nádorů. Léčba rakoviny a výzkum. 117. s. 3–32. doi:10.1007/978-1-4419-8871-3_1. ISBN 978-1-4613-4699-9. PMID 15015550.
- Machein, Marcia Regina; Plate, Karl Heinz (2004). "Role VEGF ve vývojové angiogenezi a v nádorové angiogenezi v mozku". Angiogeneze u mozkových nádorů. Léčba rakoviny a výzkum. 117. 191–218. doi:10.1007/978-1-4419-8871-3_13. ISBN 978-1-4613-4699-9. PMID 15015562.
- Eremina V, Quaggin SE (2004). "Role VEGF-A ve glomerulárním vývoji a funkci". Curr. Opin. Nephrol. Hyperteny. 13 (1): 9–15. doi:10.1097/00041552-200401000-00002. PMID 15090854.
- Storkebaum E, Lambrechts D, Carmeliet P (2004). „VEGF: kdysi považován za specifický angiogenní faktor, nyní zapojený do neuroprotekce“. BioEssays. 26 (9): 943–54. doi:10.1002 / bies.20092. PMID 15351965.
- Ribatti D (2005). „Rozhodující role faktoru vaskulární permeability / vaskulárního endoteliálního růstového faktoru v angiogenezi: historický přehled“. Br. J. Haematol. 128 (3): 303–9. doi:10.1111 / j.1365-2141.2004.05291.x. PMID 15667531.
- Loureiro RM, D'Amore PA (2005). "Transkripční regulace vaskulárního endoteliálního růstového faktoru u rakoviny". Cytokinový růstový faktor Rev. 16 (1): 77–89. doi:10.1016 / j.cytogfr.2005.01.005. PMID 15733833.
- Herbst RS, Onn A, Sandler A (2005). „Angiogeneze a rakovina plic: prognostické a terapeutické důsledky“. J. Clin. Oncol. 23 (14): 3243–56. doi:10.1200 / JCO.2005.18.853. PMID 15886312.
- Pufe T, Kurz B, Petersen W a kol. (2006). "Vliv biomechanických parametrů na expresi VEGF a endostatinu v kostním a kloubním systému". Ann. Anat. 187 (5–6): 461–72. doi:10.1016 / j.aanat.2005.06.008. PMID 16320826.
- Tong JP, Yao YF (2006). „Příspěvek VEGF a PEDF k choroidální angiogenezi: potřeba vyvážených výrazů“. Clin. Biochem. 39 (3): 267–76. doi:10.1016 / j.clinbiochem.2005.11.013. PMID 16409998.
- Lambrechts D, Carmeliet P (2007). „VEGF na neurovaskulárním rozhraní: terapeutické důsledky pro onemocnění motorických neuronů“. Biochim. Biophys. Acta. 1762 (11–12): 1109–21. doi:10.1016 / j.bbadis.2006.04.005. PMID 16784838.
- Matsumoto T, Mugishima H (2006). „Transdukce signálu přes receptory vaskulárního endoteliálního růstového faktoru (VEGF) a jejich role v aterogenezi“. J. Atheroscler. Tromb. 13 (3): 130–5. doi:10,5551 / jat.13.130. PMID 16835467.
- Bogaert E, Van Damme P, Van Den Bosch L, Robberecht W (2006). „Vaskulární endoteliální růstový faktor u amyotrofické laterální sklerózy a jiných neurodegenerativních onemocnění“. Svalový nerv. 34 (4): 391–405. doi:10,1002 / mus.20609. PMID 16856151.
- Mercurio AM, Lipscomb EA, Bachelder RE (2006). "Neaniogenní funkce VEGF u rakoviny prsu". Journal of Mammary Gland Biology and Neoplasia. 10 (4): 283–90. CiteSeerX 10.1.1.476.2778. doi:10.1007 / s10911-006-9001-9. PMID 16924371.
- Makinde T, Murphy RF, Agrawal DK (2007). "Imunomodulační role vaskulárního endoteliálního růstového faktoru a angiopoetinu-1 při remodelaci dýchacích cest". Curr. Mol. Med. 6 (8): 831–41. doi:10.2174/156652406779010795. PMID 17168735.
- Rini BI, Rathmell WK (2007). "Biologické aspekty a vazebné strategie vaskulárního endoteliálního růstového faktoru u karcinomu ledvinových buněk". Clin. Cancer Res. 13 (2 Pt 2): 741s – 746s. doi:10.1158 / 1078-0432.CCR-06-2110. PMID 17255303.
- Jiang C, Zuo F, Wang Y, Lu H, Yang Q, Wang J (2016). "Progesteron mění expresi VEGF a BDNF a podporuje neurogenezi po ischemické mozkové příhodě". Mol. Neurobiol. 54: 571–581. doi:10.1007 / s12035-015-9651-r. PMC 4938789. PMID 26746666.
- Rodgers LS, Lalani S, Hardy KM, Xiang X, Broka D, Antin PB, Camenisch TD (2006). „Depolymerizovaný hyaluronan indukuje vaskulární endoteliální růstový faktor, negativní regulátor vývojové transformace epitelu na mezenchymální“. Circ. Res. 99 (6): 583–9. doi:10.1161 / 01.RES.0000242561.95978.43. PMID 16931798.
- Qaum, T; Xu, Q; Joussen, AM; et al. (2001). „Rozpad bariéry krve a sítnice iniciované VEGF u časného diabetu“. Investujte Ophthalmol Vis Sci. 42 (10): 2408–2413. PMID 11527957.
externí odkazy
- Cévní + endoteliální + růst + faktory v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- Proteopedia Faktor faktoru vaskulárního endoteliálního růstu - struktura vaskulárního endoteliálního růstového faktoru v interaktivním 3D