Speciace - Speciation

Speciace je evoluční proces, kterým se populace vyvíjejí, aby se oddělily druh. Biolog Řečník F. Cook vytvořil termín v roce 1906 pro kladogeneze, rozdělení linií, na rozdíl od anageneze, phyletic evoluce v liniích.[1][2][3] Charles Darwin byl první, kdo popsal roli přírodní výběr ve své specifikaci ve své knize z roku 1859 O původu druhů.[4] Také identifikoval sexuální výběr jako pravděpodobný mechanismus, ale považoval to za problematické.
Existují čtyři geografické způsoby speciace v přírodě, založené na míře speciace populace jsou od sebe izolovaní: alopatrický, peripatrický, parapatrický, a sympatrický. Speciace může být také vyvolána uměle prostřednictvím chov zvířat, zemědělství, nebo laboratorní experimenty. Zda genetický drift je menší nebo hlavní přispěvatel do speciace je předmětem mnoha probíhajících diskusí.
Může dojít k rychlé sympatrické speciaci polyploidie, například zdvojnásobením počtu chromozomů; výsledkem jsou potomci, kteří jsou okamžitě reprodukčně izolovaný z mateřské populace. Nové druhy lze také vytvořit prostřednictvím hybridizace následuje, pokud je hybrid zvýhodněn přirozeným výběrem, reprodukční izolací.
Historické pozadí
Při řešení otázky původu druhů existují dvě klíčové otázky: (1) jaké jsou evoluční mechanismy speciace a (2) co vysvětluje oddělenost a individualitu druhů v biotě? Od doby Charlese Darwina se úsilí o pochopení podstaty druhů primárně zaměřilo na první aspekt a nyní se všeobecně uznává, že kritickým faktorem vzniku nových druhů je reprodukční izolace.[5] Dále se zaměříme na druhý aspekt původu druhů.
Darwinovo dilema: Proč existují druhy?
v O původu druhů (1859), Darwin interpretoval biologickou evoluci z hlediska přirozeného výběru, ale byl zmaten shlukováním organismů do druhů.[6] Kapitola 6 Darwinovy knihy má název „Obtížnosti teorie“. Při diskusi o těchto „obtížích“ si všiml „Zaprvé, proč, pokud druhy pocházejí z jiných druhů nečekaně jemnými gradacemi, neuvidíme všude nespočet přechodných forem? Proč není celá příroda zmatená místo toho, aby byl druh, jak vidíme je dobře definované? “ Toto dilema lze označit jako nepřítomnost nebo vzácnost přechodných odrůd v prostoru stanovišť.[7]
Další dilema,[8] související s první, je absence nebo vzácnost přechodných odrůd v čase. Darwin poukázal na to, že podle teorie přirozeného výběru „musely existovat nespočetné přechodné formy“, a přemýšlel, „proč je nenajdeme v nespočetných počtech v kůře Země?“ To, že jasně definované druhy ve skutečnosti v přírodě existují, a to jak v prostoru, tak v čase, znamená, že některé základní rysy přirozeného výběru fungují k vytváření a udržování druhů.[6]
Vliv sexuální reprodukce na formování druhů
Tvrdilo se, že řešení prvního Darwinova dilematu spočívá v tom, že překračování sexuální reprodukce má vlastní cenu vzácnosti.[9][10][11][12][13] Náklady na vzácnost vznikají následovně. Pokud se na gradientu zdrojů vyvíjí velké množství samostatných druhů, z nichž každý je skvěle přizpůsoben velmi úzkému pásmu na tomto gradientu, bude se každý druh nutně skládat z velmi malého počtu členů. Hledání partnera za těchto okolností může představovat potíže, když mnoho jedinců v sousedství patří k jiným druhům. Za těchto okolností, pokud se náhodně zvýší velikost populace jakéhokoli druhu (na úkor jednoho nebo druhého ze sousedních druhů, pokud je prostředí nasycené), bude to pro jeho členy okamžitě snazší najít sexuální partnery . Členové sousedních druhů, jejichž populace se zmenšila, mají větší potíže s hledáním kamarádů, a proto tvoří páry méně často než větší druhy. To má efekt sněhové koule, kdy velké druhy rostou na úkor menších a vzácnějších druhů a nakonec je vedou k zánik. Nakonec zbývá jen několik druhů, z nichž každý se výrazně liší od ostatních.[9][10][12] Náklady na raritu zahrnují nejen náklady na nalezení partnera, ale také nepřímé náklady, jako jsou náklady na komunikaci při hledání partnera při nízké hustotě populace.

Rarity s sebou přináší další náklady. Vzácné a neobvyklé vlastnosti jsou velmi zřídka výhodné. Ve většině případů označují (nemý ) mutace, což je téměř jisté, že bude škodlivé. Proto se sluší na sexuální stvoření, aby se zabránilo kamarádům se vzácnými nebo neobvyklými rysy (koinofilie ).[15][16] Sexuální populace proto rychle vylučují vzácné nebo periferní fenotypové rysy, čímž kanalizují celý vnější vzhled, jak je znázorněno na přiloženém obrázku Ledňáček říční, Ispidina picta. Tato uniformita všech dospělých členů sexuálního druhu stimulovala šíření polní vodítka na ptácích, savcích, plazech, hmyzu a mnoha dalších taxony, ve kterém lze druh popsat jedinou ilustrací (nebo dvěma, v případě sexuální dimorfismus ). Jakmile se populace stane stejně homogenním vzhledem, jaký je typický pro většinu druhů (a je to znázorněno na fotografii ledňáčka říčního), její členové se vyhnou páření s příslušníky jiných populací, které vypadají odlišně od sebe.[17] Vyhýbání se kamarádům vykazujícím vzácné a neobvyklé fenotypové znaky tedy nevyhnutelně vede k reprodukční izolaci, jedné z charakteristických znaků speciace.[18][19][20][21]
V kontrastním případě organismů to rozmnožovat nepohlavně neexistují žádné náklady na vzácnost; následkem toho jsou pouze jemné úpravy. Nepohlavní organismy tedy velmi často vykazují neustálé variace ve formě (často v mnoha různých směrech), které Darwin očekával, že způsobí evoluce, což vede k jejich klasifikaci na „druhy“ (přesněji morphospecies ) velmi obtížné.[9][15][16][22][23][24]
Režimy

V průběhu roku proběhly všechny formy přirozené speciace vývoj; přetrvávají však diskuse o relativní důležitosti každého mechanismu při řízení biologická rozmanitost.[25]
Jedním z příkladů přirozené speciace je rozmanitost třízubý stickleback, a námořní ryby, které po poslední ledové období, prošel specifikací do nového sladkovodní kolonie v izolovaných jezerech a potocích. Během odhadovaných 10 000 generací vykazují tyčinky strukturální rozdíly, které jsou větší než rozdíly mezi nimi rody ryb, včetně variací ve ploutvích, změn v počtu nebo velikosti jejich kostních desek, variabilní struktury čelistí a barevných rozdílů.[26]
Alopatrický
Během alopatrické (ze starořečtiny Allos, „ostatní“ + patrā, "vlast", speciace, populace se rozdělí na dvě geograficky izolované populace (např fragmentace stanovišť kvůli geografickým změnám, jako je horská formace ). Izolované populace pak podstoupí genotypové nebo fenotypový divergence jako: (a) jsou vystaveny rozdílům selektivní tlaky; b) nezávisle podléhají genetickému driftu; c) jiné mutace vznikají ve dvou populacích. Když se populace znovu dostanou do kontaktu, vyvinuly se tak, že jsou reprodukčně izolované a již nejsou schopné výměny geny. Ostrovní genetika je termín spojený s tendencí malých izolovaných genetických skupin produkovat neobvyklé vlastnosti. Mezi příklady patří ostrovní nanismus a radikální změny například u některých slavných ostrovních řetězců Komodo. The Galapágy jsou zvláště známí svým vlivem na Charlese Darwina. Během svých pěti týdnů to slyšel Galapágy želvy mohl být identifikován ostrovem a všiml si toho pěnkavy lišil se od jednoho ostrova k druhému, ale teprve o devět měsíců později si uvědomil, že takové skutečnosti mohou ukázat, že druhy jsou proměnlivé. Když se vrátil do Anglie, jeho spekulace o vývoji se prohloubily poté, co ho odborníci informovali, že se jedná o oddělené druhy, nejen o odrůdy, a skvěle také o tom, že ostatní odlišní ptáci Galapága jsou všechny druhy pěnkav. Ačkoli pěnkavy byly pro Darwina méně důležité, novější výzkum ukázal, že ptáci jsou nyní známí jako Darwinovy pěnkavy být klasickým případem adaptivního evolučního záření.[27]
Peripatric
V peripatrické speciaci, podformě alopatrické speciace, se tvoří nové druhy v izolovaných menších periferních populacích, které brání ve výměně genů s hlavní populací. Souvisí to s konceptem a zakladatelský efekt, protože malé populace často podstupují úzká místa. Genetický drift se často navrhuje hrát významnou roli v peripatrické speciaci.[28][29]
Případové studie zahrnují Mayrovo vyšetřování ptačí fauny;[30] australský pták Petroica vícebarevná;[31] a reprodukční izolace v populacích Drosophila s výhradou úzkých míst v populaci.[Citace je zapotřebí ]
Parapatric
V parapatrické speciaci existuje pouze částečné oddělení zón dvou odlišných populací, které poskytuje geografie; jednotlivci každého druhu mohou čas od času přijít do styku nebo překročit stanoviště, ale snížená zdatnost heterozygot vede k výběru chování nebo mechanismů, které jim brání křížení. Parapatrická speciace je modelována na kontinuální variaci v rámci „jediného“ propojeného stanoviště, které působí spíše jako zdroj přirozeného výběru, než na účinky izolace stanovišť produkované peripatrickou a alopatrickou speciací.[32]
Parapatrická speciace může být spojena s rozdílnou krajinnou závislostí výběr. I když existuje tok genů mezi dvěma populacemi může silná diferenciální selekce bránit asimilaci a nakonec se mohou vyvinout různé druhy.[33] Rozdíly v stanovištích mohou být při vývoji reprodukční izolace důležitější než doba izolace. Kavkazské skalní ještěrky Darevskia Rudis, D. valentini a D. portschinskii Všechno hybridizovat navzájem v jejich hybridní zóna; mezi nimi je však silnější hybridizace D. portschinskii a D. rudis, které se oddělily dříve, ale žijí na podobných stanovištích než mezi nimi D. valentini a dva další druhy, které se později oddělily, ale žijí v klimaticky odlišných stanovištích.[34]
Ekologové odkazují[je zapotřebí objasnění ] parapatrická a peripatrická speciace z hlediska ekologické výklenky. Aby byl nový druh úspěšný, musí být k dispozici mezera. Kroužkové druhy jako Larusi tvrdí se, že rackové ilustrují probíhající speciaci, i když situace může být složitější.[35] Tráva Anthoxanthum odoratum může začínat parapatrickou speciaci v oblastech kontaminace minami.[36]
Soucitný

Sympatická speciace je tvorba dvou nebo více potomků z jednoho druhu předků, kteří všichni zaujímají stejnou geografickou polohu.
Často citované příklady sympatrické speciace se vyskytují u hmyzu, který je závislý na různých hostitel rostliny ve stejné oblasti.[37][38]

Nejznámějším příkladem sympatrické speciace je příklad cichlidy z východní Afrika obývající Jezera Rift Valley, zejména Viktoriino jezero, Jezero Malawi a Jezero Tanganika. Popsaných druhů je přes 800 a podle odhadů by v této oblasti mohlo být i přes 1 600 druhů. Jejich vývoj je citován jako příklad obou přírodní a sexuální výběr.[39][40] Studie z roku 2008 naznačuje, že sympatrická speciace se vyskytla v Tennessee jeskynní mloci.[41] Sympatická speciace poháněná ekologickými faktory může také odpovídat za mimořádnou rozmanitost korýšů žijících v hlubinách Sibiře jezero Bajkal.[42]
Začínající speciace byla navržena jako zvláštní forma sympatrické speciace, při níž se malé skupiny jedinců postupně více izolují od rodové populace chováním přednostně jeden s druhým. Tento typ speciace by byl poháněn spojením různých výhod příbuzenské plemenitby, jako je vyjádření výhodných recesivních fenotypů, snížení rekombinační zátěže a snížení nákladů na sex.[43]

Hloh létá (Rhagoletis pomonella ), známý také jako červí moucha jablečná, podle všeho prochází sympatickou speciací.[44] Různé populace hlohu se živí různými plody. Zřetelná populace se v Severní Americe objevila v 19. století po nějaké době jablka Byl představen nepůvodní druh. Tato populace, která se živí jablky, se obvykle živí pouze jablky, a nikoli historicky preferovaným ovocem hlohy. Současná populace hlohového krmení se normálně neživí jablky. Některé důkazy, například těch šest ze třinácti alozym loci se liší, že hlohové mušky dozrávají později v sezóně a jejich zrání trvá déle než muškám jablečným; a že existuje jen málo důkazů o křížení (vědci zdokumentovali 4–6% míru hybridizace) naznačuje, že dochází k sympatrické speciaci.[45]
Metody výběru
Posílení

Výztuž, někdy označovaná jako Wallaceův efekt, je proces, kterým přirozený výběr zvyšuje reprodukční izolaci.[18] Může nastat po oddělení dvou populací stejného druhu a následném kontaktu. Pokud byla jejich reprodukční izolace úplná, pak se z nich již vyvinuli dva samostatné nekompatibilní druhy. Pokud je jejich reprodukční izolace neúplná, pak další páření mezi populacemi vyprodukuje hybridy, které mohou nebo nemusí být plodné. Pokud jsou hybridy neplodné nebo plodné, ale méně vhodné než jejich předkové, bude docházet k další reprodukční izolaci a v zásadě došlo k speciaci (např. Jako v koně a osli ).[46]
Důvodem je to, že pokud mají rodiče hybridních potomků každý přirozeně vybrané vlastnosti pro své vlastní určité prostředí, hybridní potomci budou mít vlastnosti obou, a proto by nezapadali ani do ekologického výklenku, ani do žádného z rodičů. Nízká zdatnost hybridů by způsobila upřednostňování výběru asortativní páření, který by řídil hybridizaci. Tomu se po evolučním biologovi někdy říká Wallaceův efekt Alfred Russel Wallace který na konci 19. století navrhl, že by to mohl být důležitý faktor speciace.[47]
Naopak, pokud jsou hybridní potomci vhodnější než jejich předkové, populace se spojí zpět do stejného druhu v oblasti, s níž jsou v kontaktu.
Pro parapatrickou a sympatrickou speciaci je nutná výztuž upřednostňující reprodukční izolaci. Bez posílení se geografická oblast kontaktu mezi různými formami stejného druhu, nazývaná jejich „hybridní zóna“, nevyvine do hranice mezi různými druhy. Hybridní zóny jsou oblasti, kde se setkávají a kříží rozdílné populace. Hybridní potomci jsou v těchto oblastech velmi běžní a jsou obvykle vytvářeni odlišnými druhy, které do nich přicházejí sekundární kontakt. Bez zesílení by tyto dva druhy byly nekontrolovatelné příbuzenské plemenitby. Posílení může být indukováno v experimentech s umělou selekcí, jak je popsáno níže.
Ekologický
Ekologický výběr je „interakce jednotlivců s jejich prostředím během získávání zdrojů“.[48] Přirozený výběr je neodmyslitelně zapojen do procesu speciace, přičemž „v rámci ekologické speciace populace v různých prostředích nebo populace využívající různé zdroje zažívají kontrastní tlaky přirozeného výběru na vlastnosti, které přímo nebo nepřímo způsobují vývoj reprodukční izolace“.[49] Existují důkazy o roli, kterou hraje ekologie v procesu speciace. Studie populací lipniček podporují ekologicky spojenou speciaci vznikající jako vedlejší produkt,[50] spolu s četnými studiemi paralelních speciací, kde se vyvíjí izolace mezi nezávislými populacemi druhů přizpůsobujících se kontrastním prostředím, než mezi nezávislými populacemi přizpůsobujícími se podobným prostředím.[51] K ekologické speciaci dochází u většiny důkazů, „... nashromážděných ze studií adaptace a izolace reprodukce shora dolů“.[51]
Sexuální výběr
To je široce oceňováno sexuální výběr mohl řídit speciaci v mnoha subtech, nezávisle na tom přírodní výběr.[52] Avšak výraz „speciace“ má v této souvislosti tendenci být používán ve dvou různých, nikoli však vzájemně se vylučujících smyslech. První a nejčastěji používaný význam odkazuje na „zrození“ nových druhů. To znamená, že rozdělení existujícího druhu na dva samostatné druhy nebo rozmnožování nového druhu z mateřského druhu, obojí poháněné biologickým „módním výstřelkem“ (preference pro rys nebo rysy v jednom nebo obou pohlaví, která nutně nemusí mít žádné adaptivní vlastnosti).[52][53][54][55] V druhém smyslu se „speciace“ vztahuje k rozšířené tendenci sexuálních tvorů seskupovat se do jasně definovaných druhů,[56][19] spíše než vytvářet kontinuum fenotypy jak v čase, tak v prostoru - což by byl zjevnější nebo logičtější důsledek přirozeného výběru. To skutečně uznal Darwine jako problematické a zahrnuty do jeho O původu druhů (1859), pod nadpisem „Problémy s teorií“.[6] Existuje několik návrhů, jak volba partnera může hrát významnou roli při řešení Darwinovo dilema.[19][9][15][16][17][57] Pokud dojde ke speciaci při absenci přirozeného výběru, lze jej označit jako nekologická speciace[58][59].
Umělá speciace


Nové druhy byly vytvořeny uživatelem chov zvířat, ale data a metody iniciace těchto druhů nejsou jasné. Domácí protějšek divokého předka se často může křížit a plodit plodné potomky, jako v případě domácích. dobytek, které lze považovat za stejný druh jako několik odrůd divočiny vůl, gaur, jaka atd., nebo domácí ovce které se mohou křížit s muflon.[60][61]
Nejlépe zdokumentované výtvory nových druhů v laboratoři byly provedeny na konci 80. let. William R. Rice a George W. Salt se chovali Drosophila melanogaster ovocné mušky pomocí bludiště se třemi různými možnostmi prostředí, jako je světlo / tma a vlhko / sucho. Každá generace byla umístěna do bludiště a skupiny much, které vyšly ze dvou z osmi východů, byly odděleny, aby se navzájem rozmnožovaly ve svých příslušných skupinách. Po třiceti pěti generacích byly obě skupiny a jejich potomci reprodukčně izolováni kvůli jejich silným preferencím stanovišť: pářili se pouze v oblastech, které upřednostňovali, a tak se nespárovali s muchami, které preferovaly ostatní oblasti.[62] Historie takových pokusů je popsána v Rice a Elen E. Hostert (1993).[63][64]Diane Dodd pomocí laboratorního experimentu ukázala, jak se může vyvinout reprodukční izolace Drosophila pseudoobscura ovocné mušky po několika generacích umístěním do různých médií, médií na bázi škrobu a maltózy.[65]

Doddův experiment bylo pro mnoho dalších snadné replikovat, včetně jiných druhů ovocných mušek a potravin.[66] Výzkum v roce 2005 ukázal, že tento rychlý vývoj reprodukční izolace může být ve skutečnosti pozůstatkem infekce Wolbachia bakterie.[67]
Alternativně jsou tato pozorování v souladu s představou, že sexuální stvoření se ze své podstaty zdráhají spojit s jednotlivci, jejichž vzhled nebo chování se liší od normy. Riziko, že takové odchylky jsou způsobeny dědičné maladaptations je velmi vysoká. Pokud je tedy sexuální stvoření, které není schopno předvídat budoucí směr přirozeného výběru, podmíněno vytvořením co nejschopnějších potomků, vyhne se kamarádům s neobvyklými zvyky nebo rysy.[68][69][15][16][17] Sexuální stvoření budou mít nevyhnutelně sklon seskupovat se do reprodukčně izolovaných druhů.[16]
Genetika
Bylo nalezeno několik speciačních genů. Obvykle zahrnují posilovací proces v pozdních fázích speciace. V roce 2008 byl hlášen speciační gen způsobující reprodukční izolaci.[70] Způsobuje to hybridní sterilitu mezi příbuznými poddruhy. Pořadí speciace tří skupin od společného předka může být nejasné nebo neznámé; sbírka tří takových druhů se označuje jako „trichotomie“.
Speciace prostřednictvím polyploidie

Polyploidie je mechanismus, který v systému způsobil mnoho rychlých speciačních událostí soucit protože potomci například tetraploidního x diploidního páření často vedou k triploidnímu sterilnímu potomstvu.[71] Avšak ne všechny polyploidy jsou reprodukčně izolovány z jejich rodičovských rostlin a tok genů se může stále vyskytovat například prostřednictvím triploidního hybridního x diploidního páření, které produkuje tetraploidy, nebo páření mezi meioticky nezredukovaný gamety z diploidů a gamety z tetraploidů (viz také hybridní speciace ).
Bylo navrženo, že mnoho stávajících rostlinných a většinových druhů zvířat prošlo během své evoluční historie událostí polyploidizace.[72][73] Reprodukce úspěšných druhů polyploidů je někdy bezpohlavní partenogeneze nebo apomixis, protože z neznámých důvodů je mnoho nepohlavních organismů polyploidních. Jsou známy vzácné případy polyploidních savců, které však nejčastěji vedou k prenatální smrti.
Hybridní speciace
Hybridizace mezi dvěma různými druhy někdy vede k odlišnosti fenotyp. Tento fenotyp může být také zdatnější než rodičovská linie a jako takový může přirozený výběr upřednostňovat tyto jedince. Nakonec, pokud je dosaženo reprodukční izolace, může to vést k samostatnému druhu. Dosažení reprodukční izolace mezi hybridy a jejich rodiči je obzvláště obtížné, a proto je hybridní speciace považována za extrémně vzácnou událost. The Mariana divoká se předpokládá, že vznikly hybridní speciací.
Hybridizace je důležitým prostředkem speciace v rostlinách, protože polyploidie (s více než dvěma kopiemi každého z nich chromozóm ) je snášenlivý u rostlin snadněji než u zvířat.[74][75] Polyploidie je u hybridů důležitá, protože umožňuje reprodukci, přičemž dvě různé sady chromozomů jsou během meiózy schopny spárovat se stejným partnerem.[73] Polyploidy mají také větší genetickou rozmanitost, což jim umožňuje vyhnout se inbrední deprese u malých populací.[76]
Hybridizace bez změny počtu chromozomů se nazývá homoploidní hybridní speciace. To je považováno za velmi vzácné, ale bylo uvedeno v Heliconius motýli[77] a slunečnice. Polyploidní speciace, která zahrnuje změny v počtu chromozomů, je častějším jevem, zejména u druhů rostlin.
Genová transpozice
Theodosius Dobžanský, který studoval ovocné mušky v raných dobách genetického výzkumu ve 30. letech 20. století, spekuloval, že části chromozomů, které přecházejí z jednoho místa na druhé, mohou způsobit rozdělení druhu na dva různé druhy. Zmapoval, jak je možné, aby se části chromozomů přemístily do genomu. Tyto mobilní sekce mohou u mezidruhových hybridů způsobit sterilitu, což může působit jako speciační tlak. Teoreticky byl jeho nápad zdravý, ale vědci dlouho diskutovali o tom, zda se to skutečně stalo v přírodě. Nakonec se ukázalo, že v přírodě se vyskytuje konkurenční teorie zahrnující postupné hromadění mutací tak často, že genetici do značné míry odmítli hypotézu pohyblivých genů.[78] Výzkum z roku 2006 však ukazuje, že skok genu z jednoho chromozomu do druhého může přispět k narození nových druhů.[79] Tím se potvrdí mechanismus reprodukční izolace, klíčová složka speciace.[80]
Sazby
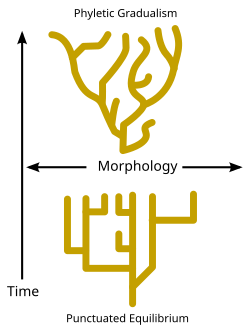
Diskutuje se o rychlosti, s jakou dochází ke speciačním událostem v geologickém čase. Zatímco někteří evoluční biologové tvrdí, že speciační události zůstaly v průběhu času relativně konstantní a postupné (známé jako „fyletický gradualismus“ - viz diagram), některé paleontologové jako Niles Eldredge a Stephen Jay Gould[81] tvrdí, že druhy obvykle zůstávají nezměněny po dlouhé časové úseky a že ke speciaci dochází pouze v relativně krátkých intervalech, což je pohled známý jako přerušovaná rovnováha. (Viz schéma a Darwinovo dilema.)
Interpunkční vývoj
Evoluce může být extrémně rychlá, jak ukazuje tvorba domestikovaných zvířat a rostlin ve velmi krátkém geologickém prostoru, který trvá jen několik desítek tisíc let. Kukuřice (Zea mays) například byl vytvořeno v Mexiko za pouhých několik tisíc let, počínaje zhruba před 7 000 až 12 000 lety.[82] To vyvolává otázku, proč je dlouhodobá míra vývoje mnohem pomalejší, než je teoreticky možné.[83][84][85][86]







Evoluce se vnucuje druhům nebo skupinám. U některých to není plánováno ani usilováno Lamarckista způsob.[87] Mutace, na nichž proces závisí, jsou náhodné události a kromě „tiché mutace „které neovlivňují funkčnost ani vzhled nosiče, jsou tedy obvykle nevýhodné a jejich šance prokázat se v budoucnu jako užitečná je mizivě malá. Proto by sice druh nebo skupina mohla mít prospěch z možnosti přizpůsobit se novému prostředí hromaděním široké škály genetických variací, je to na úkor Jednotlivci kteří musí nést tyto mutace, dokud malá, nepředvídatelná menšina z nich nakonec přispěje k takové adaptaci. To znamená, že schopnost vyvinout by vyžadovalo výběr skupiny, koncept zdiskreditovaný (například) George C. Williams,[88] John Maynard Smith[89] a Richard Dawkins[90][91][92][93] jako selektivně nevýhodné pro jednotlivce.
Řešení druhého Darwinova dilematu by tedy mohlo přijít následovně:
Pokud jsou sexuální jedinci znevýhodněni přenesením mutací na své potomky, vyhnou se mutovaným kamarádům se zvláštními nebo neobvyklými vlastnostmi.[69][15][16][57] Mutace, které ovlivňují vnější vzhled jejich nosičů, budou potom zřídka přeneseny na další a následující generace. Proto by byly zřídka testovány přirozeným výběrem. Evoluce je proto účinně zastavena nebo výrazně zpomalena. Jediné mutace, které se v populaci mohou akumulovat, jsou při tomto přerušovaném rovnovážném pohledu takové mutace, které nemají znatelný vliv na vnější vzhled a funkčnost jejich nositelů (tj. Jsou „tiché“ nebo „neutrální mutace, "které mohou být a jsou použity ke sledování příbuznost a věk populací a druhů.[15][94]) Tento argument naznačuje, že k evoluci může dojít pouze v případě, že nelze zabránit mutantním kamarádům v důsledku vážného nedostatku potenciálních kamarádů. K tomu pravděpodobně dojde v malé izolované komunity. Ty se vyskytují nejčastěji na malých ostrovech, v odlehlých údolích, jezerech, říčních systémech nebo jeskyních,[95] nebo v důsledku a masový zánik.[94] Za těchto okolností je nejen přísně omezen výběr kamarádů, ale úzká místa populace, účinky zakladatele, genetický drift a příbuzenská plemenitba způsobují rychlé, náhodné změny v genetickém složení izolované populace.[95] Kromě toho může hybridizace s příbuznými druhy chycenými ve stejném izolátu zavést další genetické změny. Pokud je izolovaná populace, jako je tato přežívá své genetické otřesy, a následně expanduje do neobsazeného výklenku nebo do výklenku, ve kterém má výhodu nad svými konkurenty, vznikne nový druh nebo poddruh. Z geologického hlediska to bude náhlá událost. Obnovení vyhýbání se mutantním kamarádům poté opět povede k evoluční stagnaci.[81][84]
Ve zjevném potvrzení tohoto přerušovaného rovnovážného pohledu na vývoj, fosilní záznam evoluční progrese typicky sestává z druhů, které se náhle objeví a nakonec zmizí, o stovky tisíc nebo miliony let později, bez jakékoli změny vnějšího vzhledu.[81][94][96] Graficky jsou tyto fosilní druhy reprezentovány čarami rovnoběžnými s časovou osou, jejichž délky ukazují, jak dlouho každý z nich existoval. Skutečnost, že čáry zůstávají rovnoběžné s časovou osou, ilustruje neměnný vzhled každého z fosilních druhů znázorněných na grafu. Během existence každého druhu se v náhodných intervalech objevují nové druhy, z nichž každý trvá také stovky tisíc let, než zmizí beze změny vzhledu. Přesnou příbuznost těchto souběžných druhů obecně nelze určit. To je znázorněno na schématu znázorňujícím distribuce homininových druhů v čase od homininy oddělené od linie, která vedla k vývoji našich nejbližších žijících příbuzných primátů, šimpanzů.[96]
Podobné evoluční časové linie viz například paleontologický seznam Africké dinosaury, Asijské dinosaury, Lampriformes a Amiiformes.
Viz také
Reference
- ^ Berlocher 1998, str.3
- ^ Cook, řečník F. (30. března 1906). "Faktory formování druhů". Věda. 23 (587): 506–507. Bibcode:1906Sci .... 23..506C. doi:10.1126 / science.23.587.506. PMID 17789700.
- ^ Cook, řečník F. (listopad 1908). „Evoluce bez izolace“. Americký přírodovědec. 42 (503): 727–731. doi:10.1086/279001.
- ^ Via, Sara (16. června 2009). „Přirozený výběr v akci během speciace“. PNAS. 106 (Suppl 1): 9939–9946. Bibcode:2009PNAS..106,9939V. doi:10.1073 / pnas.0901397106. PMC 2702801. PMID 19528641.
- ^ Mayr 1982, str. 273
- ^ A b C Darwin 1859
- ^ Sepkoski, David (2012). „1. Darwinovo dilema: paleontologie, fosilní záznam a evoluční teorie“. Přečtení fosilních záznamů: Růst paleobiologie jako evoluční disciplína. University of Chicago Press. str. 9–50. ISBN 978-0-226-74858-0.
Jednou z jeho největších obav bylo, že „neúplnost“ fosilního záznamu bude použita ke kritice jeho teorie: že zjevné „mezery“ ve fosilní posloupnosti lze uvést jako negativní důkaz, přinejmenším pro jeho návrh, aby všechny organismy sestoupili po minutách a postupných úpravách od společného předka.
- ^ Stower, Hannah (2013). „Řešení Darwinova dilema“. Genetika hodnocení přírody. 14 (747): 747. doi:10.1038 / nrg3614. S2CID 45302603.
Téměř simultánní výskyt nejmodernějších plánů těl zvířat při kambrické explozi naznačuje krátký interval rychlého fenotypového a genetického vývoje, o kterém se Darwin domníval, že je příliš rychlý na to, aby ho bylo možné vysvětlit přirozeným výběrem.
- ^ A b C d Bernstein, Harris; Byerly, Henry C .; Hopf, Frederic A .; et al. (21. prosince 1985). „Pohlaví a výskyt druhů“. Journal of Theoretical Biology. 117 (4): 665–690. doi:10.1016 / S0022-5193 (85) 80246-0. PMID 4094459.
- ^ A b Hopf, Frederic A .; Hopf, F. W. (únor 1985). "Role Alleeho efektu při balení druhů". Teoretická populační biologie. 27 (1): 27–50. doi:10.1016/0040-5809(85)90014-0.
- ^ Bernstein a Bernstein 1991
- ^ A b Michod 1995
- ^ Michod 1999
- ^ Hockey, Dean & Ryan 2005 176, 193
- ^ A b C d E F Koeslag, Johan H. (10. května 1990). „Koinophilia seskupuje sexuální tvory do druhů, podporuje stagnaci a stabilizuje sociální chování“. Journal of Theoretical Biology. 144 (1): 15–35. doi:10.1016 / s0022-5193 (05) 80297-8. ISSN 0022-5193. PMID 2200930.
- ^ A b C d E F Koeslag, Johan H. (21. prosince 1995). "Na motoru zrychlení". Journal of Theoretical Biology. 177 (4): 401–409. doi:10.1006 / jtbi.1995.0256. ISSN 0022-5193.
- ^ A b C Poelstra, Jelmer W .; Vijay, Nagarjun; Bossu, Christen M .; et al. (20. června 2014). „Genomická krajina, která je základem fenotypové integrity tváří v tvář toku genů u vran“. Věda. 344 (6190): 1410–1414. Bibcode:2014Sci ... 344.1410P. doi:10.1126 / science.1253226. PMID 24948738. S2CID 14431499.
Fenotypové rozdíly mezi Carrionem a vrány s kapucí napříč hybridizační zónou v Evropě pravděpodobně nebudou způsobeny asortativním krytím.
- Komentář Mazhuvancherry K. Unnikrishnana a H. S. Akhily - ^ A b Ridley, Marku. „Speciace - Jaká je role výztuže ve speciaci?“. Citováno 2015-09-07. Převzato z Vývoj (2004), 3. vydání (Malden, MA: Blackwell Publishing ), ISBN 978-1-4051-0345-9.
- ^ A b C Maynard Smith 1989, str. 275–280
- ^ Mayr 1988
- ^ Williams 1992, str. 118
- ^ Maynard Smith, John (Prosinec 1983). „Genetika stagnace a interpunkce“. Výroční přehled genetiky. 17: 11–25. doi:10.1146 / annurev.ge.17.120183.000303. PMID 6364957.
- ^ Clapham, Tutin a Warburg 1952
- ^ Udělit 1971
- ^ Baker, Jason M. (červen 2005). „Adaptivní speciace: Role přirozeného výběru v mechanismech geografické a negeografické speciace“ (PDF). Studie historie a filozofie vědy Část C: Studie historie a filozofie biologických a biomedicínských věd. 36 (2): 303–326. doi:10.1016 / j.shpsc.2005.03.005. PMID 19260194.
- ^ Kingsley, David M. (leden 2009). „Odhalení rozmanitosti: Od atomů k vlastnostem“. Scientific American. 300 (1): 52–59. doi:10.1038 / scientificamerican0109-52. PMID 19186749.
- ^ Sulloway, Frank J. (30. září 1982). „The Beagle sbírky Darwinových pěnkav (Geospizinae) ". Bulletin of the British Museum (Natural History), Zoology. 43 (2): 49–58.
- ^ Jerry A. Coyne; H. Allen Orr (2004). Speciace. Sinauer Associates. str. 105. ISBN 978-0-87893-091-3.
- ^ Lawson, Lucinda P .; Bates, John M .; Menegon, Michele; Loader, Simon P. (2015). „Divergence na okrajích: peripatrická izolace v montánním ostnatém throated žabím komplexu“. BMC Evoluční biologie. 15 (128): 128. doi:10.1186 / s12862-015-0384-3. PMC 4487588. PMID 26126573.
- ^ Mayr 1992, str. 21–53
- ^ Tokeshi, M. (1999). Koexistence druhů: ekologické a evoluční perspektivy. Oxford: Blackwell Science. str. 27. ISBN 0632061464. OCLC 47011551.
- ^ „Speciace: původ nových druhů | Naučte se vědu na scitable“. www.nature.com. Citováno 2020-02-16.
- ^ Endler 1977
- ^ Tarkhnishvili, David; Murtskhvaladze, Marine; Gavashelishvili, Alexander (srpen 2013). „Speciace u kavkazských ještěrek: klimatická odlišnost stanovišť je důležitější než doba izolace“. Biologický žurnál společnosti Linnean. 109 (4): 876–892. doi:10.1111 / bij.12092.
- ^ Liebers, Dorit; Knijff, Peter de; Helbig, Andreas J. (2004). „Komplex Racek stříbřitý není prstencový druh“. Proc Biol Sci. 271 (1542): 893–901. doi:10.1098 / rspb.2004.2679. PMC 1691675. PMID 15255043.
- ^ „Parapatrická speciace“. University of California Berkeley. Citováno 3. dubna 2017.
- ^ Feder, Jeffrey L .; Xianfa Xie; Rull, Juan; et al. (3. května 2005). „Mayr, Dobžanskij a Bush a složitost sympatrické speciace v Rhagoletis". PNAS. 102 (Suppl 1): 6573–6580. Bibcode:2005PNAS..102,6573F. doi:10.1073 / pnas.0502099102. PMC 1131876. PMID 15851672.
- ^ Berlocher, Stewart H .; Feder, Jeffrey L. (leden 2002). „Sympatická speciace u fytofágního hmyzu: posouváme se mimo kontroverze?“. Každoroční přezkum entomologie. 47: 773–815. doi:10.1146 / annurev.ento.47.091201.145312. PMID 11729091. S2CID 9677456.
- ^ Machado, Heather E .; Pollen, Alexander A .; Hofmann, Hans A .; et al. (Prosinec 2009). „Mezidruhové profilování genové exprese na základě komparativní genomové hybridizace: Přehled a nový přístup u afrických cichlídovitých ryb“. Integrativní a srovnávací biologie. 49 (6): 644–659. doi:10.1093 / icb / icp080. PMID 21665847.
- ^ Fanoušek, Shaohua; Elmer, Kathryn R .; Meyer, Axel (5. února 2012). „Genomika adaptace a speciace u ryb cichlíd: nedávné pokroky a analýzy v afrických a neotropických liniích“. Filozofické transakce královské společnosti B. 367 (1587): 385–394. doi:10.1098 / rstb.2011.0247. PMC 3233715. PMID 22201168.
- ^ Niemiller, Matthew L .; Fitzpatrick, Benjamin M .; Miller, Brian T. (květen 2008). „Nedávná divergence s tokem genů u mloků jeskynních v Tennessee (Plethodontidae: Gyrinophilus) odvozené z genových genealogií ". Molekulární ekologie. 17 (9): 2258–2275. doi:10.1111 / j.1365-294X.2008.03750.x. PMID 18410292. S2CID 20761880.
- ^ Martens, Koen (květen 1997). "Speciace ve starodávných jezerech". Trendy v ekologii a evoluci. 12 (5): 177–182. doi:10.1016 / S0169-5347 (97) 01039-2. PMID 21238028.
- ^ Joly, E. (9. prosince 2011). „Existence druhů spočívá na metastabilní rovnováze mezi příbuzenským plemenem a plemenitbou. Esej o úzkém vztahu mezi speciací, příbuzenským plemenem a recesivními mutacemi“. Biology Direct. 6: 62. doi:10.1186/1745-6150-6-62. PMC 3275546. PMID 22152499.
- ^ Feder, Jeffrey L .; Roethele, Joseph B .; Filchak, Kenneth; et al. (Březen 2003). „Důkazy o inverzním polymorfismu souvisejícím s tvorbou sympatrické hostitelské rasy u jablek červovitých, Rhagoletis pomonella". Genetika. 163 (3): 939–953. PMC 1462491. PMID 12663534. Citováno 2015-09-07.
- ^ Berlocher, Stewart H .; Bush, Guy L. (červen 1982). "Elektroforetická analýza fylogeneze Rhagoletis (Diptera: Tephritidae)". Systematická zoologie. 31 (2): 136–155. doi:10.2307/2413033. JSTOR 2413033.
- ^ Sætre, Glenn-Peter (2012). "Posílení". eLS. doi:10.1002 / 9780470015902.a0001754.pub3. ISBN 978-0470016176. Chybějící nebo prázdný
| název =(Pomoc) - ^ Ollerton, Jeff (září 2005). „Speciace: doba kvetení a Wallaceho efekt“ (PDF). Dědičnost. 95 (3): 181–182. doi:10.1038 / sj.hdy.6800718. PMID 16077739. S2CID 13300641. Archivovány od originál (PDF) dne 06.06.2007. Citováno 2015-09-07.
- ^ Howard D. Rundle a Patrik Nosil (2005), „Ekologická speciace“, Ekologie Dopisy, 8 (3): 336–352, doi:10.1111 / j.1461-0248.2004.00715.x
- ^ Dolph Schluter (2001), „Ekologie a původ druhů“, Trendy v ekologii a evoluci, 16 (7): 372–380, doi:10.1016 / S0169-5347 (01) 02198-X, PMID 11403870
- ^ Jeffrey S. McKinnon; et al. (2004), „Důkazy o roli ekologie ve speciaci“, Příroda, 429 (6989): 294–298, Bibcode:2004 Natur.429..294M, doi:10.1038 / nature02556, PMID 15152252, S2CID 2744267
- ^ A b Dolph Schluter (2009), „Důkazy pro ekologickou speciaci a její alternativu“, Věda, 326 (5915): 737–740, Bibcode:2009Sci ... 323..737S, doi:10.1126 / science.1160006, PMID 19197053, S2CID 307207
- ^ A b Panhuis, Tami M .; Butlin, Roger; Zuk, Marlene; et al. (Červenec 2001). "Sexual selection and speciation" (PDF). Trendy v ekologii a evoluci. 16 (7): 364–371. doi:10.1016/s0169-5347(01)02160-7. PMID 11403869.
- ^ Darwin, Charles; A. R. Wallace (1858). „O tendenci druhů vytvářet odrůdy a o udržování odrůd a druhů přirozenými způsoby výběru“ (PDF). Journal of the Proceedings of the Linnean Society of London. Zoologie. 3 (9): 46–50. doi:10.1111 / j.1096-3642.1858.tb02500.x.
- ^ Darwin, Charles (1859). "4". O původu druhů (1. vyd.). str. 89.
- ^ Eberhard, W. G. (1985). Sexuální výběr a genitálie zvířat. Harvard University Press, Cambridge, Massachusetts
- ^ Gould, Stephen Jay (1980). A Quahog is a Quahog. The Panda's thumb. More reflections in natural history. New York: W. W. Norton & Company. str.204–213. ISBN 978-0-393-30023-9.
- ^ A b Miller 2013, pp. 177, 395–396
- ^ Rundell, Rebecca J.; Price, Trevor D. (2009-07-01). "Adaptive radiation, nonadaptive radiation, ecological speciation and nonecological speciation". Trendy v ekologii a evoluci. 24 (7): 394–399. doi:10.1016/j.tree.2009.02.007. ISSN 0169-5347. PMID 19409647.
- ^ Czekanski-Moir, Jesse E .; Rundell, Rebecca J. (2019-05-01). "The Ecology of Nonecological Speciation and Nonadaptive Radiations". Trendy v ekologii a evoluci. 34 (5): 400–415. doi:10.1016/j.tree.2019.01.012. ISSN 0169-5347. PMID 30824193.
- ^ Nowak 1999
- ^ Hiendleder, Stefan; Kaupe, Bernhard; Wassmuth, Rudolf; et al. (May 7, 2002). "Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies". Sborník Královské společnosti B. 269 (1494): 893–904. doi:10.1098/rspb.2002.1975. PMC 1690972. PMID 12028771.
- ^ Rice, William R.; Salt, George W. (June 1988). „Speciace prostřednictvím rušivého výběru na preferenci stanoviště: experimentální důkazy“. Americký přírodovědec. 131 (6): 911–917. doi:10.1086/284831.
- ^ Rice, William R.; Hostert, Ellen E. (December 1993). "Laboratory Experiments on Speciation: What Have We Learned in 40 Years?". Vývoj. 47 (6): 1637–1653. doi:10.2307/2410209. JSTOR 2410209. PMID 28568007.
- ^ Gavrilets, Sergey (Říjen 2003). "Perspective: Models of Speciation: What Have We Learned in 40 Years?". Vývoj. 57 (10): 2197–2215. doi:10.1554/02-727. PMID 14628909. S2CID 198158082.
- ^ Dodd, Diane M. B. (September 1989). "Reproductive Isolation as a Consequence of Adaptive Divergence in Drosophila pseudoobscura". Vývoj. 43 (6): 1308–1311. doi:10.2307/2409365. JSTOR 2409365. PMID 28564510.
- ^ Kirkpatrick, Mark; Ravigné, Virginie (March 2002). „Speciace přirozeným a sexuálním výběrem: modely a experimenty“. Americký přírodovědec. 159 (S3): S22 – S35. doi:10.1086/338370. ISSN 0003-0147. PMID 18707367. S2CID 16516804.
- ^ Koukou, Katerina; Pavlikaki, Haris; Kilias, George; et al. (Leden 2006). "Influence of Antibiotic Treatment and Wolbachia Curing on Sexual Isolation Among Drosophila melanogaster Cage Populations". Vývoj. 60 (1): 87–96. doi:10.1554/05-374.1. PMID 16568634. S2CID 198153238.
- ^ Symons 1979
- ^ A b Langlois, Judith H.; Roggman, Lori A. (March 1990). "Attractive Faces Are Only Average". Psychologická věda. 1 (2): 115–121. doi:10.1111/j.1467-9280.1990.tb00079.x. S2CID 18557871.
- ^ Phadnis, Nitin; Orr, H. Allen (16. ledna 2009). "A Single Gene Causes Both Male Sterility and Segregation Distortion in Drosophila Hybrids". Věda. 323 (5912): 376–379. Bibcode:2009Sci...323..376P. doi:10.1126/science.1163934. PMC 2628965. PMID 19074311.
- ^ Ramsey, Justin; Schemske, Douglas W. (November 1998). "Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants". Výroční přehled ekologie a systematiky. 29: 467–501. doi:10.1146/annurev.ecolsys.29.1.467.
- ^ Otto, Sarah P.; Whitton, Jeannette (December 2000). "Polyploid Incidence and Evolution" (PDF). Výroční přehled genetiky. 34: 401–437. CiteSeerX 10.1.1.323.1059. doi:10.1146 / annurev.genet.34.1.401. PMID 11092833.
- ^ A b Comai, Luca (November 2005). "The advantages and disadvantages of being polyploid". Genetika hodnocení přírody. 6 (11): 836–846. doi:10.1038/nrg1711. PMID 16304599. S2CID 3329282.
- ^ Wendel, Jonathan F. (January 2000). "Genome evolution in polyploids". Molekulární biologie rostlin. 42 (1): 225–249. doi:10.1023/A:1006392424384. PMID 10688139. S2CID 14856314.
- ^ Sémon, Marie; Wolfe, Kenneth H. (December 2007). "Consequences of genome duplication". Aktuální názor na genetiku a vývoj. 17 (6): 505–512. doi:10.1016/j.gde.2007.09.007. PMID 18006297.
- ^ Soltis, Pamela S.; Soltis, Douglas E. (June 20, 2000). "The role of genetic and genomic attributes in the success of polyploids". PNAS. 97 (13): 7051–7057. Bibcode:2000PNAS...97.7051S. doi:10.1073/pnas.97.13.7051. PMC 34383. PMID 10860970.
- ^ Mavarez, Jesús; Salazar, Camilo A.; Bermingham, Eldredge; et al. (15. června 2006). "Speciation by hybridization in Heliconius motýli “. Příroda. 441 (7095): 868–871. Bibcode:2006Natur.441..868M. doi:10.1038/nature04738. PMID 16778888. S2CID 2457445.
- ^ Sherwood, Jonathan (September 8, 2006). "Genetic Surprise Confirms Neglected 70-Year-Old Evolutionary Theory" (Tisková zpráva). University of Rochester. Citováno 2015-09-10.
- ^ Masly, John P.; Jones, Corbin D.; Mohamed, A. F. Noor; et al. (8. září 2006). "Gene Transposition as a Cause of Hybrid Sterility in Drosophila". Věda. 313 (5792): 1448–1450. Bibcode:2006Sci...313.1448M. doi:10.1126/science.1128721. PMID 16960009. S2CID 23462115.
- ^ Minkel, J. R. (September 8, 2006). "Wandering Fly Gene Supports New Model of Speciation". Scientific American. Citováno 2015-09-11.
- ^ A b C Gould, Stephen Jay; Eldredge, Niles (Spring 1977). „Interpunkční rovnováhy: znovu zváženo tempo a způsob evoluce“ (PDF). Paleobiologie. 3 (2): 115–151. doi:10.1017 / s0094837300005224. JSTOR 2400177. Archivovány od originál (PDF) dne 2014-06-24. Citováno 2015-09-15.
- ^ Laws 2010, str. 210–215
- ^ Williams 1992, chpt. 9
- ^ A b Eldredge & Gould 1972, chpt. 5
- ^ Mayr 1954, str.157–180
- ^ Maynard Smith 1989, str. 281
- ^ Gould 1980, pt. 4, chpt. 18
- ^ Williams 1974
- ^ Maynard Smith, John (March 14, 1964). "Výběr skupiny a výběr příbuzného". Příroda. 201 (4924): 1145–1147. Bibcode:1964 Natur.201.1145S. doi:10.1038 / 201114aa. S2CID 4177102.
- ^ Dawkins 1995, chpt. 4
- ^ Dawkins, Richarde (Prosinec 1994). "Burying the Vehicle". Behaviorální a mozkové vědy. 17 (4): 616–617. doi:10.1017 / S0140525X00036207. ISSN 0140-525X. Archivovány od originál dne 2006-09-15. Citováno 2015-09-15. "Remarks on an earlier article by [Elliot] Sober [sic] a David Sloan Wilson, who made a more extended argument in their recent book Unto Others : The Evolution and Psychology of Unselfish Behavior"
- ^ Dennett, Daniel C. (Prosinec 1994). "E Pluribus Unum?". Behaviorální a mozkové vědy. 17 (4): 617–618. doi:10.1017/S0140525X00036219. Archivovány od originál dne 2007-12-27. "Commentary on Wilson & Sober: Group Selection."
- ^ Růžovější, Steven (18. června 2012). "The False Allure of Group Selection". edge.org. Edge Foundation, Inc. Citováno 2015-09-15.
- ^ A b C Campbell 1990, pp. 450–451, 487–490, 499–501
- ^ A b Ayala 1982, pp. 73–83, 182–190, 198–215
- ^ A b McCarthy & Rubidge 2005
Bibliografie
- Ayala, Francisco J. (1982). Population and Evolutionary Genetics. Benjamin/Cummings Series in the Life Sciences. Menlo Park, CA: Benjamin / Cummings Pub. Co. ISBN 978-0-8053-0315-5. LCCN 81021623. OCLC 8034790.
- Berlocher, Stewart H. (1998). "Origins: A Brief History of Research on Speciation". In Howard, Daniel J.; Berlocher, Stewart H. (eds.). Nekonečné formy: druhy a speciace. New York: Oxford University Press. ISBN 978-0-19-510901-6. LCCN 97031461. OCLC 37545522.
- Bernstein, Carol; Bernstein, Harris (1991). Stárnutí, sex a opravy DNA. San Diego, CA: Akademický tisk. ISBN 978-0-12-092860-6. LCCN 90014467. OCLC 22542921.
- Campbell, Neil A. (1990). Biologie (2. vyd.). Redwood City, CA: Benjamin / Cummings Pub. Co. ISBN 978-0-8053-1800-5. LCCN 89017952. OCLC 20352649.
- Clapham, Arthur Roy; Tutin, Thomas G.; Warburg, Edmund F. (1952). Flora z Britských ostrovů. Cambridge, Velká Británie: Cambridge University Press. LCCN 52008880. OCLC 1084058.
- Darwin, Charles (1859). O původu druhů pomocí přirozeného výběru nebo zachování příznivých ras v boji o život (1. vyd.). Londýn: John Murray. LCCN 06017473. OCLC 741260650. The book is available from Kompletní dílo Charlese Darwina online. Citováno 2015-09-12.
- Dawkins, Richarde (1995). River Out of Eden: A Darwinian Pohled na život. Série mistrů vědy. New York: Základní knihy. ISBN 978-0-465-01606-8. LCCN 94037146. OCLC 31376584.
- Eldredge, Niles; Gould, Stephen Jay (1972). "Punctuated Equilibria: An Alternative to Phyletic Gradualism". In Schopf, Thomas J. M. (ed.). Modely v paleobiologii. San Francisco, CA: Freeman Cooper & Co. ISBN 978-0-87735-325-6. LCCN 72078387. OCLC 572084. Přetištěno Eldredge 1985, pp. 193–223
- Eldredge, Niles (1985). Time Frames: The Rethinking of Darwinian Evolution and the Theory of Punctuated Equilibria. New York: Simon & Schuster. ISBN 978-0-671-49555-8. LCCN 84023632. OCLC 11443805.
- Endler, John A. (1977). Geographic Variation, Speciation, and Clines. Monografie v populační biologii. 10. Princeton, NJ: Princeton University Press. s. 1–246. ISBN 978-0-691-08187-8. LCCN 76045896. OCLC 2645720. PMID 409931.
- Gould, Stephen Jay (1980). Panda palec: Více odrazů v přírodní historii (1. vyd.). New York: W. W. Norton & Company. ISBN 978-0-393-01380-1. LCCN 80015952. OCLC 6331415.
- Grant, Verne (1971). Speciace rostlin. New York: Columbia University Press. ISBN 978-0-231-03208-7. LCCN 75125620. OCLC 139834.
- Hockey, Phil A. R.; Dean, W. Richard J.; Ryan, Peter G., eds. (2005). Roberts Birds z jižní Afriky (7. vydání). Cape Town, South Africa: Trustees of the J. Voelcker Bird Book Fund. ISBN 978-0-620-34053-3. LCCN 2006376728. OCLC 65978899.
- Zákony, Bill (2010). Padesát rostlin, které změnily běh dějin. Buffalo, NY: Firefly Books. ISBN 978-1-55407-798-4. LCCN 2011414731. OCLC 711609823.
- Maynard Smith, John (1989). Evoluční genetika. Oxford; New York: Oxford University Press. ISBN 978-0-19-854215-5. LCCN 88017041. OCLC 18069049.
- Mayr, Ernst (1954). "Change of Genetic Environment and Evolution". v Huxley, Julian; Hardy, Alister C.; Ford, Edmund B. (eds.). Evoluce jako proces. Londýn: Allen & Unwin. LCCN 54001781. OCLC 974739.
- Mayr, Ernst (1982). Růst biologického myšlení: rozmanitost, evoluce a dědičnost. Cambridge, Massachusetts: Belknap Press z Harvard University Press. ISBN 978-0-674-36445-5. LCCN 81013204. OCLC 7875904.
- Mayr, Ernst (1988). Směrem k nové filozofii biologie: Pozorování evolucionisty. Cambridge, Massachusetts: Belknap Press z Harvard University Press. ISBN 978-0-674-89665-9. LCCN 87031892. OCLC 17108004.
- Mayr, Ernst (1992). "Speciational Evolution or Punctuated Equilibrium". In Somit, Albert; Peterson, Steven A. (eds.). Dynamics of Evolution: The Punctuated Equilibrium Debate in the Natural and Social Sciences. Ithaca, NY: Cornell University Press. ISBN 978-0-8014-9763-6. LCCN 91055569. OCLC 24374091.
- McCarthy, Terence; Rubidge, Bruce (2005). The Story of Earth & Life: A Southern African Perspective on a 4.6-Billion-Year Journey. Cape Town, South Africa: Struik Publishers. ISBN 978-1-77007-148-3. LCCN 2006376206. OCLC 62098231.
- Michod, Richard E. (1995). Eros a evoluce: Přirozená filozofie sexu. Helix Books. Reading, MA: Addison-Wesley. ISBN 978-0-201-40754-9. LCCN 94013158. OCLC 30625193.
- Michod, Richard E. (1999). Darwinian Dynamics: Evolutionary Transitions in Fitness and Individuality. Princeton, NJ: Princeton University Press. ISBN 978-0-691-02699-2. LCCN 98004166. OCLC 38948118.
- Miller, William B., Jr. (2013). The Microcosm Within: Evolution and Extinction in the Hologenome. Boca Raton, FL: Universal-Publishers. ISBN 978-1-61233-277-2. LCCN 2013033832. OCLC 859168474.
- Nowak, Ronald M. (1999). Walkerovi savci světa (6. vydání). Baltimore, MD: Johns Hopkins University Press. ISBN 978-0-8018-5789-8. LCCN 98023686. OCLC 39045218.
- Symons, Donald (1979). Evoluce lidské sexuality. New York: Oxford University Press. ISBN 978-0-19-502535-4. LCCN 78023361. OCLC 4494283.
- Williams, George C. (1974) [Originally published 1966]. Adaptace a přirozený výběr: Kritika některého současného evolučního myšlení. Vědecká knihovna v Princetonu. Princeton, NJ: Princeton University Press. ISBN 978-0-691-02357-1. LCCN 65017164. OCLC 8500898.
- Williams, George C. (1992). Přirozený výběr: domény, úrovně a výzvy. Oxford Series in Ecology and Evolution. New York: Oxford University Press. ISBN 978-0-19-506933-4. LCCN 91038938. OCLC 228136567.
Další čtení
- Coyne, Jerry A.; Orr, H. Allen (2004). Speciace. Sunderlands, MA: Sinauer Associates. ISBN 978-0-87893-089-0. LCCN 2004009505. OCLC 55078441.
- Gavrilets, S. (2004). Fitness Landscapes and the Origin of Species. Princeton University Press. ISBN 978-0691119830.
- Grant, Verne (1981). Speciace rostlin (2. vyd.). New York: Columbia University Press. ISBN 978-0-231-05112-5. LCCN 81006159. OCLC 7552165.
- Marko, Peter B. (2008). "Allopatry". v Jørgensen, Sven Erik; Fath, Brian (eds.). Encyclopedia of Ecology. 1, A-C (1st ed.). Oxford, Velká Británie: Elsevier. str. 131–138. ISBN 978-0-444-52033-3. LCCN 2008923435. OCLC 173240026.
- Mayr, Ernst (1963). Druhy zvířat a evoluce. Cambridge, Massachusetts: Belknap Press z Harvard University Press. ISBN 978-0-674-03750-2. LCCN 63009552. OCLC 899044868.
- Schilthuizen, Menno (2001). Frogs, Flies, and Dandelions: The Making of Species. Oxford; New York: Oxford University Press. ISBN 978-0-19-850393-4. LCCN 2001270180. OCLC 46729094.
- Shapiro, J. B.; Leducq, J-B.; Mallet, J. (2016). "What is Speciation?". Genetika PLOS. 12 (3): e1005860. doi:10.1371/journal.pgen.1005860. PMC 4816541. PMID 27030977.
- White, Michael J. D. (1978). Modes of Speciation. A Series of Books in Biology. San Francisco, Kalifornie: W. H. Freeman and Company. ISBN 978-0-7167-0284-9. LCCN 77010955. OCLC 3203453.
externí odkazy
- Boxhorn, Joseph (September 1, 1995). "Observed Instances of Speciation". Archiv TalkOrigins. Houston, TX: The TalkOrigins Foundation, Inc.
- Hawks, John D. (February 9, 2005). "Speciation". John Hawks Weblog.
- "Speciation". University of California, Berkeley.
| Základní pojmy | ||
|---|---|---|
| Geographic modes | ||
| Isolating factors | ||
| Hybridizace | ||
| Speciation in taxa | ||

| Jevy |  | ||||
|---|---|---|---|---|---|
| Modely | |||||
| Příčiny | |||||
| Teorie a pojmy | |||||
| Události vyhynutí |
| ||||
| Vyhynulé druhy |
| ||||
| Organizace | |||||
| Viz také | |||||
| |||||