Vývojová zaujatost - Developmental bias
v evoluční biologie, vývojová zaujatost odkazuje na produkci proti nebo vůči určitým ontogenetické trajektorie které v konečném důsledku ovlivňují směr a výsledek evoluční změny ovlivňováním rychlostí, velikostí, směrů a limitů vlastnost vývoj.[1][2] Historicky byl tento termín synonymizován vývojovým omezením,[1][3][4] posledně jmenovaný je však v poslední době interpretován jako odkaz pouze na negativní roli vývoje v evoluci.[5]
Role embrya

V moderní evoluční biologii je myšlenka vývojové předpojatosti zakotvena v proudu myšlení zvaném Strukturalismus (také „internalizmus“ nebo „variační strukturalismus“) zaměřený na roli organismu jako a kauzální síla evoluční změny.[6] Ve strukturalistickém pohledu je fenotypová evoluce výsledkem působení přírodní výběr na dříve „filtrované“ variace v průběhu roku ontogeneze.[7][8] To kontrastuje s Funkcionalista (také „adaptační“, „panselekcionistický“ nebo „externalistický“) pohled, ve kterém fenotypická evoluce vyplývá pouze z interakce mezi deterministickým působením přirozeného výběru a variací způsobenou mutací.[3][7]
Odůvodnění role organismu, konkrétněji embrya, jako příčinné síly v evoluci a pro existenci zkreslení je následující: Tradiční, neodarwinovský, přístup k vysvětlení procesu za evoluční změnou je přirozený výběr působící na dědičnou variaci způsobenou geneticky mutace.[9] Přirozený výběr však působí fenotypy a mutace sama o sobě neprodukuje fenotypové variace, existuje tedy koncepční mezera, pokud jde o souvislost mezi mutací a potenciální změnou fenotypu.[6] Aby mutace mohla snadno změnit fenotyp, a byla tedy viditelná pro přirozený výběr, musí upravit ontogenetickou trajektorii, proces označovaný jako vývojové přeprogramování.[10] Některé druhy přeprogramování se vyskytnou s větší pravděpodobností než jiné vzhledem k povaze mapy genotyp-fenotyp, která určuje sklon systému měnit se v určitém směru,[8][11] tedy vytvoření zaujatosti. Jinými slovy, základní architektura vývojových systémů ovlivňuje druhy možných fenotypových výsledků.
Vývojové zkreslení se však může vyvíjet přirozeným výběrem a oba procesy současně ovlivňují fenotypový vývoj. Například vývojové zkreslení může ovlivnit rychlost nebo cestu k adaptivnímu vrcholu (fenotyp s vysokou kondicí),[5] a naopak, silná směrová selekce může modifikovat vývojové zkreslení, aby se zvýšila fenotypová variace ve směru selekce.[12] Oba myšlenkové proudy se tedy vzájemně doplňují a je třeba je kombinovat, abychom lépe porozuměli procesům, které jsou základem evoluční změny.[6]

Typy zkreslení
- Vývojové omezení
- Vývojová omezení odkazují na omezení fenotypové variability (nebo absence variací) způsobená inherentní strukturou a dynamikou vývojového systému.[1] Omezení jsou zkreslení proti určitou ontogenetickou trajektorii a následně se předpokládá, že omezuje adaptivní evoluci.[12][13]
- Vývojový pohon
- Vývojový pohon se týká přirozených přirozených tendencí organismů a jejich ontogenetických trajektorií ke změně v určitém směru (tj. Zkreslení vůči určitá ontogenetická trajektorie).[14][5][6] Předpokládá se, že tento typ předpětí usnadňuje adaptivní vývoj sladěním fenotypové variability se směrem výběru.[15][12]
Distribuce fenotypových variací
Morphospace

Morfosfát je kvantitativní reprezentace fenotypů ve vícerozměrném prostoru, kde každá dimenze odpovídá znaku. Fenotyp každého organismu nebo druhu je pak reprezentován jako bod v tomto prostoru, který shrnuje kombinaci hodnot nebo stavů u každého konkrétního znaku.[16] Tento přístup se používá ke studiu vývoje realizovaných fenotypů ve srovnání s těmi, které jsou teoreticky možné, ale neexistují.[16][17]
Nerandomizovaná (anizotropní) distribuce fenotypových variací
Jedním z hlavních cílů je popsat a pochopit hybné síly distribuce fenotypových variací v přírodě evoluční biologie.[2] Jedním ze způsobů, jak studovat distribuci fenotypových variací, je zobrazení objemu morfosfátu obsazeného množinou organismů nebo druhů. Teoreticky může existovat přirozený proces, který generuje téměř rovnoměrně (kvazi stochasticky) distribuovaný vzor fenotypů v morfosfátu, pokud jde o to, že nové nezbytné druhy mají tendenci zaujímat bod v morfosfátu, který je blízký bodu jeho fylogenetických příbuzných.[18] Nyní se však všeobecně uznává, že organismy nejsou rovnoměrně distribuovány podél morfosfátu, tj. Izotropní variace, ale místo toho jsou distribuovány nepravidelně, tj. Anizotropní variace.[17][19] Jinými slovy, existuje nesoulad mezi zjevnými (nebo teoretickými) možnými fenotypy a jejich skutečnou přístupností.[17]

Některé fenotypy jsou tedy nepřístupné (nebo nemožné) kvůli základní architektuře vývojové trajektorie, zatímco jiné jsou přístupné (nebo možné).[20] Z možných fenotypů jsou však některé „snadnější“ nebo pravděpodobnější než jiné.[8][19] Například fenotyp, jako je klasická postava draka (tj. Obří tvor podobný plazům se dvěma páry končetin a předním párem křídel), může být nemožné protože u obratlovců jsou přední končetiny a přední pár křídel homologní znaky (např. ptáci a netopýři), a proto se vzájemně vylučují. Na druhou stranu, pokud jsou možné dva fenotypy (a stejně vhodné), ale jedna forma přeprogramování vyžaduje pouze jednu mutaci, zatímco druhá vyžaduje dvě nebo více, bude pravděpodobnější, že dojde k prvnímu (za předpokladu, že genetické mutace nastanou náhodně).[8]
Důležitý rozdíl mezi strukturalismem a funkcionalismem se týká především interpretace příčin prázdných oblastí v morfosféře (tj. Neexistujících fenotypů): Podle funkcionalistického pohledu odpovídají prázdné prostory fenotypům, které jsou jednak ontogeneticky možné, jednak stejně pravděpodobné, ale jsou eliminovány přirozeným výběrem kvůli jejich nízké hodnotě zdatnost.[20] Naproti tomu z pohledu strukturalismu prázdné prostory odpovídají ontogeneticky nemožným nebo nepravděpodobným fenotypům,[3][20] tedy implikující zkreslení u typů fenotypů, které lze vytvořit za předpokladu stejného množství variací (genetické mutace) v obou modelech.[6][8]
Klasické příklady anizotropních variací

V klasickém přírodním příkladu zkreslení se ukázalo, že v přírodě byla realizována jen malá část všech možných tvarů ulit šneků a skutečné druhy byly spíše omezeny na diskrétní oblasti skořápky-morfosfátu, než aby byly kontinuálně distribuovány.[21] V dalším přirozeném příkladu se ukázalo, že stonožky žijící v půdě mají enormní rozdíly v počtu párů nohou, přičemž nejnižší je 27 a nejvyšší 191 párů; neexistují však žádné druhy se sudým počtem párů nohou, což naznačuje, že buď jsou tyto fenotypy během vývoje nějak omezeny, nebo že existuje vývojový pohon na lichá čísla.[22] Naopak, vývojové abnormality (nebo teratologie ) byly také použity jako klasický příklad k pochopení logiky mechanismů, které produkují variace.[23] Například u široké škály zvířat, od ryb až po člověka, jsou dvouhlavé organismy mnohem častější než tříhlavé organismy; obdobně by siamská dvojčata teoreticky mohla „fúzovat“ jakoukoli oblastí v těle, ale fúze se vyskytuje častěji v břišní oblasti.[7][23] Tento trend byl označován jako transpecifický paralelismus, což naznačuje existenci hlubokých historických pravidel upravujících expresi abnormálních forem u vzdáleně příbuzných druhů.[7]
Předpjaté fenotypy I: Kontinuální variace
Vývojová integrace a P-matice
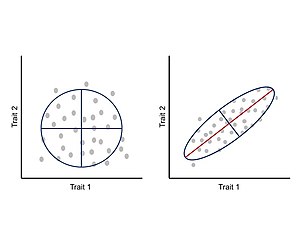
Bylo navrženo, že integrace nebo kovariace mezi vlastnostmi během vývoje omezí fenotypovou evoluci na určité oblasti morfosfátu a omezí adaptivní evoluci.[24] Tyto allometrický změny jsou v přírodě velmi rozšířené a mohou odpovídat za širokou škálu realizovaných tvarosloví a následných ekologických a fyziologických změn.[25][26] V rámci tohoto přístupu je fenotyp považován za integrovaný systém, kde se každý znak vyvíjí a vyvíjí ve shodě s ostatními znaky, a proto změna jednoho znaku ovlivňuje vzájemně působící části korelovaným způsobem.[24][27] Korelace mezi vlastnostmi je důsledkem architektury mapy genotyp-fenotyp, zejména mapy pleiotropní účinky základních genů.[11] Tuto korelovanou změnu mezi vlastnostmi lze měřit a analyzovat pomocí fenotypové variance-kovarianční matice (P-matice), která shrnuje dimenze fenotypové variability a hlavní variační osu.[24]
Kvantitativní genetika a G-matice
Kvantitativní genetika je statistický rámec zabývající se hlavně modelováním vývoje spojitých znaků.[9] V tomto rámci by korelace mezi vlastnostmi mohla být výsledkem dvou procesů: 1) přirozený výběr působící současně na několik vlastností zajišťujících, že jsou zděděny společně (tj. vazebná nerovnováha ),[28] nebo 2) přirozený výběr působící na jeden znak způsobující korelovanou změnu ostatních znaků v důsledku pleiotropních účinků genů.[11] Pro množinu vlastností je rovnicí, která popisuje rozptyl mezi vlastnostmi, vícerozměrný chovatelská rovnice Δz = β x G, kde Δz je vektor rozdílů ve zvláštních prostředcích, β je vektor výběrových koeficientů a G je matice aditivní genetická variance a kovarianci mezi vlastnostmi.[29][30] Okamžitá schopnost populace reagovat na selekci je tedy určena G-maticí, ve které je rozptyl funkcí stálé genetické variace a kovariance vyplývá z pleiotropie a vazebné nerovnováhy.[30][31] Ačkoli G-matice je jedním z nejdůležitějších parametrů ke studiu vyvíjitelnost,[12] mutační matice (M-matice), známá také jako distribuce mutačních účinků, se ukázala být stejně důležitá.[31] Matice M popisuje potenciální účinky nových mutací na stávající genetické odchylky a kovariance a tyto účinky budou záviset na epistatických a pleiotropních interakcích podkladových genů.[12][31][32] Jinými slovy, M-matice určuje G-matici, a tedy reakci na výběr populace.[31] Podobně jako u P-matice popisuje G-matice hlavní variační osu.
Cesty nejmenšího odporu

Obecným důsledkem P-matic a G-matic je, že evoluce bude mít tendenci následovat „cestu nejmenšího odporu“. Jinými slovy, je-li hlavní osa variace vyrovnána se směrem výběru, kovariace (genetická nebo fenotypová) usnadní rychlost adaptivního vývoje; je-li však hlavní osa variace kolmá na směr výběru, kovariace omezí rychlost adaptivního vývoje.[2][12][24] Obecně platí, že pro populaci pod vlivem jediného optima kondice je míra morfologické divergence (od rodového k novému fenotypu nebo mezi dvojicemi druh ) je nepřímo úměrný úhlu tvořenému hlavní variační osou a směru výběru, což způsobuje zakřivenou trajektorii skrz morfosfát.[33]
Z P-matice pro sadu znaků lze extrahovat dvě široce důležité míry sklonů k variacím: 1) Odpovědnost: schopnost vývojového systému měnit se jakýmkoli směrem a 2) Vyvinutelnost: schopnost vývojového systému měnit se směrem přirozeného výběru.[24] V druhém případě je hlavní osa fenotypové variace zarovnána se směrem výběru. Podobně z G-matice je nejdůležitějším parametrem, který popisuje sklon k variacím, olovo vlastní vektor G (napřmax), který popisuje směr největšího aditivní genetická variance pro soubor souvislých znaků v populacích.[31][33] U populace procházející směrovou selekcí, gmax ovlivní hlavní směr trajektorie.[33]
Předpjaté fenotypy II: Vlastnosti genových regulačních sítí
Hierarchie a optimální pleiotropie

GRN jsou modulární, vícevrstvé a polohierarchicky systémy genů a jejich produktů: každý transkripční faktor poskytuje více vstupů do jiných genů, vytváří komplexní pole interakcí,[35] a informace týkající se načasování, místa a množství genové exprese obecně plynou z několika kontrolních genů na vysoké úrovni přes více intermediálních genů do periferních genových baterií, které nakonec určují osud každé buňky.[19][35] Tento typ architektury naznačuje, že řídicí geny na vysoké úrovni mají tendenci být více pleiotropní ovlivňující více následných genů, zatímco střední a periferní geny mají tendenci mít mírné až nízké pleiotropní účinky.[19][35]
Obecně se očekává, že nově vzniklé mutace s vyšší dominancí a méně pleiotropními a epistatický účinky budou pravděpodobněji cílem evoluce,[36] hierarchická architektura vývojových cest tak může ovlivňovat genetický základ evolučních změn. Například se očekává, že geny v GRN s „optimálně pleiotropními“ účinky, tj. Geny, které mají nejrozšířenější účinek na vybraný znak, ale jen málo účinků na jiné znaky, akumulují vyšší podíl mutací, které způsobují evoluční změnu.[37] Tyto strategicky umístěné geny mají potenciál filtrovat náhodné genetické variace a přeložit je na nerandně funkčně integrované fenotypy, což umožňuje efektivní přístup k adaptivním variantám výběru,[12] a tak v těchto genech může být koncentrována řada mutací přispívajících k fenotypové evoluci.[36][38]
Neutrální sítě
Perspektiva mapy genotyp-fenotyp stanoví, že způsob, jakým lze mapovat genotypové variace na fenotypové variace, je zásadní pro schopnost systému vyvíjet se.[11] Prevalence neutrální mutace v přírodě znamená, že biologické systémy mají více genotypy než fenotypy,[39] a důsledkem tohoto vztahu „mnoho k několika“ mezi genotypem a fenotypem je existence neutrální sítě.[6][40] Ve vývoji jsou neutrální sítě shluky GRN, které se liší pouze v jedné interakci mezi dvěma uzly (např. Nahrazení transkripce potlačením) a přesto produkují stejný fenotypový výsledek.[6][12] V tomto smyslu lze jednotlivý fenotyp v populaci zmapovat na několik ekvivalentních GRN, které společně tvoří neutrální síť. Naopak GRN, která se liší v jedné interakci a způsobuje odlišný fenotyp, je považována za nenulovou.[6] Vzhledem k této architektuře bude pravděpodobnost mutace z jednoho fenotypu na jiný záviset na počtu neutrálních sousedů ve srovnání s neneutrálními sousedy pro konkrétní GRN,[6][12] a tedy fenotypová změna bude ovlivněna polohou GRN v síti a bude zaujatá směrem ke změnám, které vyžadují několik mutací, aby se dosáhlo sousedícího neutrálního GRN.[12][40]
Viz také
- Speciace
- John Maynard Smith
- Pere Alberch
- Stephen Jay Gould
- David M. Raup
- Stevan J. Arnold
- Günter P. Wagner
- Dolph Schluter
- David L. Stern
Reference
- ^ A b C Maynard Smith, J .; Burian, R .; Kauffman, S .; Alberch, P .; Campbell, J .; Goodwin, B .; Lande, R .; Raup, D .; Wolpert, L. (1985). "Vývojová omezení a evoluce". Čtvrtletní přehled biologie. 60 (3): 265–287. doi:10.1086/414425.
- ^ A b C Arthur, Wallace (2004). „Vliv vývoje na směr vývoje: směrem ke konsensu 21. století“. Evoluce a vývoj. 6 (4): 282–288. doi:10.1111 / j.1525-142x.2004.04033.x. ISSN 1520-541X. PMID 15230968.
- ^ A b C Gould, S. J .; Lewontin, R. C. (1979). „Spandrely San Marco a panglosovské paradigma: kritika adaptačního programu“. Proc. R. Soc. Lond. B. 205 (1161): 581–598. Bibcode:1979RSPSB.205..581G. doi:10.1098 / rspb.1979.0086. ISSN 0080-4649. PMID 42062.
- ^ Gould, Stephen Jay (1989). „Vývojové omezení v Cerionu, s poznámkami o definici a interpretaci omezení v evoluci“. Vývoj. 43 (3): 516–539. doi:10.2307/2409056. JSTOR 2409056. PMID 28568388.
- ^ A b C Arthur, Wallace (2001). "Vývojový pohon: důležitý determinant směru fenotypového vývoje". Evoluce a vývoj. 3 (4): 271–278. doi:10.1046 / j.1525-142x.2001.003004271.x. ISSN 1520-541X. PMID 11478524.
- ^ A b C d E F G h i P., WAGNER, GUNTER (2014). HOMOLOGIE, GENY A EVOLUČNÍ INOVACE. PRINCETON UNIVERSITY PRES. ISBN 978-0691180670. OCLC 1005108561.
- ^ A b C d Alberch, Pere (1989). „Logika příšer: Důkazy vnitřních omezení ve vývoji a vývoji“. Geobios. 22: 21–57. doi:10.1016 / s0016-6995 (89) 80006-3. ISSN 0016-6995.
- ^ A b C d E Arthur, Wallace (2004). Předpjatá embrya a evoluce. Cambridge: Cambridge University Press. doi:10.1017 / cbo9780511606830. ISBN 9780511606830.
- ^ A b Zimmer, Carl .; Emlen D .; Perkins, Alison EH (2013). Evoluce: smysl života. Greenwood Village, CO: Roberts. ISBN 9781319202590. OCLC 1051973071.
- ^ Arthur, Wallace (2000). „Koncept vývojového přeprogramování a hledání inkluzivní teorie evolučních mechanismů“. Evoluce a vývoj. 2 (1): 49–57. doi:10.1046 / j.1525-142x.2000.00028.x. ISSN 1520-541X. PMID 11256417.
- ^ A b C d Wagner, Günter P .; Altenberg, Lee (1996). „Perspektiva: Složité adaptace a vývoj evolučnosti“. Vývoj. 50 (3): 967–976. doi:10.1111 / j.1558-5646.1996.tb02339.x. ISSN 0014-3820. PMID 28565291.
- ^ A b C d E F G h i j Uller, Tobias; Moczek, Armin P .; Watson, Richard A .; Brakefield, Paul M .; Laland, Kevin N. (2018). „Vývojová zaujatost a evoluce: perspektiva regulační sítě“. Genetika. 209 (4): 949–966. doi:10.1534 / genetika.118.300995. ISSN 0016-6731. PMC 6063245. PMID 30049818.
- ^ Drost, Hajk-Georg; Janitza, Philipp; Grosse, Ivo; Quint, Marcel (2017). „Porovnání vývojových přesýpacích hodin napříč královstvími“. Aktuální názor na genetiku a vývoj. 45: 69–75. doi:10.1016 / j.gde.2017.03.003. PMID 28347942.
- ^ Altenberg, L. (1995). „Růst genomu a vývoj mapy genotyp-fenotyp“. In Banzhaf, W .; Eeckman, F. H. (eds.). Evoluce a biopočítání: výpočetní modely evoluce. Berlín: Springer. str.205 –259. ISBN 978-3-540-49176-7.
- ^ Altenberg, L. (2005). „Modularita ve vývoji: několik otázek na nízké úrovni“. In Callebaut, W .; Rasskin-Gutman, D .; Herbert A. Simon (eds.). Modularita: Pochopení vývoje a vývoje přirozených komplexních systémů. Cambridge, MA: MIT Press. str.99 –128. ISBN 978-0-262-03326-8.
- ^ A b Chartier, Marion; Jabbour, Florian; Gerber, Sylvain; Mitteroecker, Philipp; Sauquet, Hervé; von Balthazar, Maria; Staedler, Yannick; Crane, Peter R .; Schönenberger, Jürg (2014). „Květinový morfosfát - moderní komparativní přístup ke studiu vývoje krytosemenných rostlin“. Nový fytolog. 204 (4): 841–853. doi:10,1111 / nph.12969. ISSN 0028-646X. PMC 5526441. PMID 25539005.
- ^ A b C Gerber, Sylvain (2014). „Ne všechny silnice lze použít: vývoj indukuje anizotropní dostupnost v morfosforu“. Evoluce a vývoj. 16 (6): 373–381. doi:10.1111 / ede.12098. ISSN 1520-541X. PMID 25212955.
- ^ Kemp, T.S. (2016). Původ vyšších taxonů: paleobiologické, vývojové a ekologické perspektivy. Oxford University Press. doi:10.1093 / acprof: oso / 9780199691883.001.0001. ISBN 9780199691883.
- ^ A b C d Jablonski, D. (2017). „Přístupy k makroevoluci: 1. Obecné koncepty a původ variací“. Evoluční biologie. 44 (4): 427–450. doi:10.1007 / s11692-017-9420-0. ISSN 0071-3260. PMC 5661017. PMID 29142333.
- ^ A b C Olson, M.E. (2012). „Vývojová renesance v adaptacionismu“. Trendy v ekologii a evoluci. 27 (5): 278–287. doi:10.1016 / j.tree.2011.12.005. ISSN 0169-5347. PMID 22326724.
- ^ Raup, D.M. (1966). "Geometrická analýza navíjení skořepiny: Obecné problémy". J. Paleontol.: 1178–1190.
- ^ Arthur, W (2002). „Interakce mezi vývojovým zkreslením a přirozeným výběrem: od segmentů stonožky po obecnou hypotézu“. Dědičnost. 89 (4): 239–246. doi:10.1038 / sj.hdy.6800139. ISSN 0018-067X. PMID 12242638.
- ^ A b Blumberg, M.S. (2009). Zrůdy přírody Co nám anomálie říkají o vývoji a vývoji. Oxford University Press, USA. ISBN 9780199750641. OCLC 1058406207.
- ^ A b C d E Goswami, A .; Smaers, J. B .; Soligo, C .; Polly, P. D. (2014-08-19). „Makroevoluční důsledky fenotypové integrace: od vývoje k hlubokému času“. Phil. Trans. R. Soc. B. 369 (1649): 20130254. doi:10.1098 / rstb.2013.0254. ISSN 0962-8436. PMC 4084539. PMID 25002699.
- ^ Gould, S.J. (1966). "Alometrie a velikost v ontogenezi a fylogenezi". Biol. Rev. 41 (4): 587–640. doi:10.1111 / j.1469-185X.1966.tb01624.x. PMID 5342162.
- ^ Emlen, Douglas J. (2001-02-23). „Náklady a diverzifikace přehnaných zvířecích struktur“ (PDF). Věda. 291 (5508): 1534–1536. Bibcode:2001Sci ... 291.1534E. doi:10.1126 / science.1056607. ISSN 0036-8075. PMID 11222856.
- ^ Pigliucci, M (2003). „Fenotypová integrace: studium ekologie a evoluce komplexních fenotypů“. Ekologie Dopisy. 6 (3): 265–272. doi:10.1046 / j.1461-0248.2003.00428.x. ISSN 1461-023X.
- ^ Lande, Russell; Arnold, Stevan J. (1983). "Měření výběru u korelovaných znaků". Vývoj. 37 (6): 1210–1226. doi:10.1111 / j.1558-5646.1983.tb00236.x. JSTOR 2408842. PMID 28556011.
- ^ Arnold, S.J. (1992). "Omezení fenotypového vývoje". Americký přírodovědec. 140: S85 – S107. doi:10.1086/285398. PMID 19426028.
- ^ A b Steppan, Scott J .; Patrick C. Phillips; David Houle (2002). "Srovnávací kvantitativní genetika: vývoj G matice". Trendy v ekologii a evoluci. 17 (7): 320–327. doi:10.1016 / S0169-5347 (02) 02505-3. ISSN 0169-5347.
- ^ A b C d E Jones, Adam G .; Arnold, Stevan J .; Bürger, Reinhard (2007). „Mutační matice a vývoj evoluční schopnosti“. Vývoj. 61 (4): 727–745. doi:10.1111 / j.1558-5646.2007.00071.x. ISSN 0014-3820. PMID 17439608.
- ^ Cheverud, James M. (1984). "Kvantitativní genetika a vývojová omezení vývoje podle výběru". Journal of Theoretical Biology. 110 (2): 155–171. doi:10.1016 / s0022-5193 (84) 80050-8. ISSN 0022-5193. PMID 6492829.
- ^ A b C Schluter, Dolph (1996). "Adaptivní záření podél genetických linií nejmenšího odporu". Vývoj. 50 (5): 1766–1774. doi:10.2307/2410734. JSTOR 2410734. PMID 28565589.
- ^ Hoekstra, HE (05.07.06). "Genetika, vývoj a vývoj adaptivní pigmentace u obratlovců". Dědičnost. 97 (3): 222–234. doi:10.1038 / sj.hdy.6800861. ISSN 0018-067X. PMID 16823403.
- ^ A b C Erwin, Douglas H .; Davidson, Eric H. (2009). „Vývoj hierarchických regulačních sítí genů“ (PDF). Genetika hodnocení přírody. 10 (2): 141–148. doi:10.1038 / nrg2499. ISSN 1471-0056. PMID 19139764.
- ^ A b Stern, D.L. (2011). Evoluce, vývoj a předvídatelný genom. Greenwood Village, Colorado: Roberts and Company Publishers. ISBN 978-1936221011. OCLC 762460688.
- ^ Kopp, A. (2009). „Metamodely a fylogenetická replikace: Systematický přístup k vývoji vývojových cest“. Vývoj. 63 (11): 2771–2789. doi:10.1111 / j.1558-5646.2009.00761.x. PMID 19545263.
- ^ Stern, D.L .; Orgogozo, V. (2008). „Místa evoluce: Jak předvídatelná je genetická evoluce?“. Vývoj. 62 (9): 2155–2177. doi:10.1111 / j.1558-5646.2008.00450.x. ISSN 0014-3820. PMC 2613234. PMID 18616572.
- ^ Schuster, Peter; Fontana, Walter; Stadler, Peter F .; Hofacker, Ivo L. (1994). "Od sekvencí k tvarům a zpět: případová studie sekundárních struktur RNA". Proc. R. Soc. Lond. B. 255 (1344): 279–284. Bibcode:1994RSPSB.255..279S. doi:10.1098 / rspb.1994.0040. ISSN 0962-8452. PMID 7517565.
- ^ A b Wagner, Andreas (2011). „Sítě genotypu osvětlují evoluční omezení“ (PDF). Trendy v ekologii a evoluci. 26 (11): 577–584. doi:10.1016 / j.tree.2011.07.001. ISSN 0169-5347. PMID 21840080.
Další čtení
- Ontogeneze a fylogeneze (Gould, 1977)
- Předpjatá embrya a evoluce (Arthur, 2004)
- Evoluce: Vývojový přístup (Arthur, 2010)
- Homologie, geny a evoluční inovace (Wagner, 2014)
- Evoluce, vývoj a předvídatelný genom (Stern, 2011)