Arabidopsis thaliana - Arabidopsis thaliana
| Arabidopsis thaliana | |
|---|---|
 | |
| Vědecká klasifikace | |
| Království: | Plantae |
| Clade: | Tracheofyty |
| Clade: | Krytosemenné rostliny |
| Clade: | Eudicots |
| Clade: | Rosidy |
| Objednat: | Brassicales |
| Rodina: | Brassicaceae |
| Rod: | Arabidopsis |
| Druh: | A. thaliana |
| Binomické jméno | |
| Arabidopsis thaliana | |
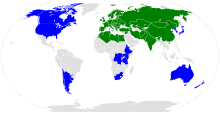 | |
Rozsah Arabidopsis thaliana.
| |
| Synonyma[1] | |
Arabis thaliana | |
Arabidopsis thaliana, thale řeřicha, myš-ušní řeřicha nebo arabidopsis, je malý kvetoucí rostlina původem z Eurasie a Afrika.[2][3][4][5][6][7] A. thaliana je považován za plevel; nachází se u silnic a v narušené zemi.
Zima roční s relativně krátkým životním cyklem, A. thaliana je populární modelový organismus v biologie rostlin a genetika. Pro komplexní mnohobuněčný eukaryot, A. thaliana má relativně malý genom přibližně 135 megabázové páry (Mbp).[8] Byla to první rostlina, která má svůj genom sekvenován, a je oblíbeným nástrojem pro pochopení molekulární biologie mnoha rostlinných znaků, včetně květ vývoj a snímání světla.
Popis

Arabidopsis thaliana je roční (zřídka dvouletý ) rostlina, obvykle dorůstající do výšky 20–25 cm.[6] The listy tvoří růžici na spodní části rostliny, s několika listy také na kvetoucí stonku. Bazální listy jsou zelené až mírně purpurové barvy, 1,5–5 cm dlouhé a 2–10 mm široké, s celkovým až hrubě pilovitým okrajem; stonkové listy jsou menší a neloupané, obvykle s celým okrajem. Listy jsou pokryty malými jednobuněčnými chloupky trichomy. The květiny jsou 3 mm v průměru, uspořádané do a chocholík; jejich struktura je typická Brassicaceae. The ovoce je siliqua 5–20 mm dlouhý, obsahující 20–30 semena.[9][10][11][12] Kořeny mají jednoduchou strukturu, s jediným primárním kořenem, který roste svisle dolů a později vytváří menší boční kořeny. Tyto kořeny tvoří interakce s rhizosféra bakterie jako např Bacillus megaterium.[13]

A. thaliana může dokončit celý svůj životní cyklus za šest týdnů. Centrální stonka, která produkuje květiny, roste asi po třech týdnech a květy se přirozeně opylují. V laboratoři A. thaliana mohou být pěstovány v Petriho miskách, květináčích nebo hydroponii, pod zářivkami nebo ve skleníku.[14]
Taxonomie
Rostlina byla poprvé popsána v roce 1577 v Pohoří Harz podle Johannes Thal (1542–1583), lékař z Nordhausen, Thüringen, Německo, kdo to nazval Pilosella siliquosa. V roce 1753 Carl Linné přejmenoval rostlinu Arabis thaliana na počest Thal. V roce 1842 německý botanik Gustav Heynhold postavil nový rod Arabidopsis a umístil rostlinu do tohoto rodu. Jméno rodu, Arabidopsis, pochází z řecký, což znamená „připomínající Arabové "(rod, do kterého jej Linnaeus původně umístil).
Tisíce přirozených inbredních přírůstků A. thaliana byly shromážděny z celé jeho přirozené a zavedené oblasti.[15] Tyto přístupy vykazují značné genetické a fenotypové variace, které lze použít ke studiu adaptace tohoto druhu na různá prostředí.[15]
Rozšíření a stanoviště
A. thaliana je původem z Evropa, Asie, Afrika a lidská pozorování naznačují, že jeho geografické rozložení je poměrně kontinuální od Středomoří na Skandinávie a Španělsko na Řecko.[16] Rovněž se zdá, že je původem z tropických alpských ekosystémů v Africe a možná v Jižní Africe.[17][18] Bylo zavedeno a naturalizováno po celém světě,[19] včetně Severní Ameriky ca. 17. století.[20]
A. thaliana snadno roste a často je průkopníkem ve skalnatých, písčitých a vápenitých půdách. Obecně se považuje za plevel, a to kvůli jeho rozšířené distribuci v zemědělských oblastech, na silnicích, železničních tratích, na odpadních pozemcích a jiných narušených stanovištích,[19][21] ale kvůli své omezené konkurenční schopnosti a malé velikosti není kategorizován jako škodlivý plevel.[22] Jako většina druhů Brassicaceae, A. thaliana je jedlý lidé jako salát nebo vařený, ale nemá rád široké použití jako jarní zelenina.[23]
Použití jako modelový organismus
Botanici a biologové začali zkoumat A. thaliana počátkem 20. století a první systematický popis mutantů byl proveden kolem roku 1945.[24] A. thaliana je nyní široce používán pro studium rostlinné vědy, počítaje v to genetika, vývoj, populační genetika a vývoj rostlin.[25][26][27] Ačkoli A. thaliana má malý přímý význam pro zemědělství, má několik zvláštností, které z něj činí užitečný model pro pochopení genetické, buněčné a molekulární biologie kvetoucích rostlin.

První mutant v A. thaliana byl dokumentován v roce 1873 autorem Alexander Braun, popisující a dvojitý květ fenotyp (mutovaný gen byl pravděpodobně Agamous, klonován a charakterizován v roce 1990).[28] Avšak ne až do roku 1943 Friedrich Laibach (který publikoval číslo chromozomu v roce 1907) navrhuje A. thaliana jako modelový organismus.[29] Jeho studentka Erna Reinholzová zveřejnila svoji diplomovou práci dne A. thaliana v roce 1945, popisující první sbírku A. thaliana mutanty, které generovali pomocí rentgen mutageneze. Laibach pokračoval ve svých důležitých příspěvcích pro A. thaliana výzkum shromážděním velkého počtu přístupů (často sporně označovaných jako „ekotypy '). S pomocí Alberta Kranze byly tyto organizovány do velké sbírky 750 přírodních přírůstků A. thaliana z celého světa.
V padesátých a šedesátých letech John Langridge a George Rédei hrála důležitou roli při založení A. thaliana jako užitečný organismus pro biologické laboratorní experimenty. Rédei napsal několik odborných recenzí, které pomohly představit model vědecké komunitě. Začátek A. thaliana výzkumná komunita se datuje k vydání zpravodaje s názvem Arabidopsis Informační služba (AIS), založená v roce 1964. První mezinárodní Arabidopsis Konference se konala v roce 1965 v Göttingen, Německo.
V 80. letech A. thaliana se začaly široce používat ve výzkumných laboratořích po celém světě. Byl to jeden z několika kandidátů, kteří zahrnovali kukuřice, petunie, a tabák.[29] Poslední dva byly atraktivní, protože byly snadno transformovatelné pomocí tehdejších současných technologií, zatímco kukuřice byla zavedeným genetickým modelem biologie rostlin. Rok 1986 byl pro rok 2006 průlomovým A. thaliana jako modelový závod, ve kterém T-DNA -zprostředkovaný proměna a první naklonovaný A. thaliana gen byl popsán.[30][31]
Genomika

Jaderný genom
Jeho malá velikost genom a skutečnost, že je diploidní, dělá Arabidopsis thaliana užitečné pro genetické mapování a sekvenování - s přibližně 157 mega párů bází[34] a pět chromozomy, A. thaliana má jeden z nejmenších genomů mezi rostlinami.[8] Dlouho se myslelo, že má nejmenší genom ze všech kvetoucích rostlin,[35] ale tento titul je nyní považován za patřící rostlinám rodu Genlisea, objednat Lamiales, s Genlisea tuberosa, masožravá rostlina vykazující velikost genomu přibližně 61 Mbp.[36] Jednalo se o první rostlinný genom, který byl sekvenován, dokončený v roce 2000 Arabidopsis Genome Initiative.[37] Nejaktuálnější verze A. thaliana genom je udržován Arabidopsis Information Resource (TAIR).[38] Hodně práce bylo uděláno pro přiřazení funkcí k jeho 27 000 geny a 35 000 proteinů, které kódují.[39] Postgenomický výzkum, jako je metabolomika, také poskytl užitečné poznatky o metabolismu tohoto druhu a o tom, jak narušení životního prostředí[40] může ovlivnit metabolické procesy.[41]
Genom chloroplastů
Plastom z Arabidopsis thaliana je molekula DNA dlouhá 154 478 párů bází,[32] velikost typicky se vyskytující u většiny kvetoucích rostlin (viz seznam sekvenovaných plastomů ). Zahrnuje 136 genů kódujících malé podjednotkové ribozomální proteiny (rps, žlutě: viz obrázek), velké podjednotky ribozomální proteiny (rpl, oranžová), hypotetické proteiny s otevřeným čtecím rámcem chloroplastů (ycf, citron), bílkoviny účastnící se fotosyntetických reakcí (zelená) nebo jiných funkcí (červená), ribozomální RNA (rrn, modrá) a přenosové RNA (trn, Černá).[33]
Mitochondriální genom
Mitochondriální genom Arabidopsis thaliana je 367 808 párů bází dlouhý a obsahuje 57 genů.[42] V regionu existuje mnoho opakovaných oblastí Arabidopsis mitochondriální genom. Největší opakování rekombinovat pravidelně a izomerizovat genom.[43] Jako většina rostlinných mitochondriálních genomů, i Arabidopsis mitochondriální genom existuje jako komplexní uspořádání překrývajících se rozvětvených a lineárních molekul in vivo.[44]
Genetika
Genetická transformace z A. thaliana je rutina, využití Agrobacterium tumefaciens převést DNA do genomu rostliny. Současný protokol, nazývaný „květinový dip“, zahrnuje jednoduché ponoření květin do roztoku obsahujícího Agrobacterium nesoucí požadovaný plazmid a detergent.[45][46] Tato metoda se vyhne potřebě tkáňová kultura nebo regenerace rostlin.
The A. thaliana sbírky genových knockoutů jsou jedinečným zdrojem pro biologii rostlin, který umožňuje dostupnost vysoce výkonné transformace a financování genomických zdrojů. Místo inzerce T-DNA bylo určeno pro více než 300 000 nezávislých transgenních linií, přičemž informace a semena jsou přístupná online Databáze T-DNA. Prostřednictvím těchto sbírek jsou inzerční mutanti k dispozici pro většinu genů v A. thaliana.
Charakterizované přístupy a mutantní linie A. thaliana sloužit jako experimentální materiál v laboratorních studiích. Nejčastěji používané čáry pozadí jsou Lehm (Landsberg erecta) a Col nebo Columbia.[47] Další základní linie méně často citované ve vědecké literatuře jsou Ws, nebo Wassilewskija, C24, Cvi nebo Kapverdské ostrovy, Nossen atd. (Viz např.[48]) Byly získány a charakterizovány sady úzce souvisejících přístupů s názvem Col-0, Col-1 atd.; obecně jsou mutantní linie dostupné prostřednictvím zásobních center, z nichž nejznámější jsou Nottingham Arabidopsis Stock Center-NASC[47] a Arabidopsis Biological Resource Center-ABRC v Ohiu, USA.[49]Přistoupení Col-0 vybral Rédei z (neozářené) populace semen označených jako „Landsberg“, kterou obdržel od Laibacha.[50] Columbia (pojmenovaný podle umístění Rédeiho bývalé instituce, University of Missouri v Columbia ) byl referenční přístup seřazený v Arabidopsis Genome Initiative. Později (Landsberg erecta) linii vybral Rédei (kvůli svému nízkému vzrůstu) z populace Landsbergů, kterou mutagenizoval pomocí rentgenových paprsků. Jako Lehm sbírka mutantů je odvozena z této počáteční linie, Lehm-0 neodpovídá přístupům Landsberg, které označily La-0, La-1 atd.
Tvorba trichomů je iniciována proteinem GLABROUS1. Vyřazení příslušného genu vedou k lysý rostliny. Tento fenotyp již byl použit v editace genů experimenty a může být zajímavý jako vizuální značka pro výzkum rostlin ke zdokonalení metod úpravy genů, jako např CRISPR / Cas9.[51][52]
Non-Mendelian dědičnost diskuse
V roce 2005 vědci z Purdue University navrhl to A. thaliana vlastnil alternativu k dříve známým mechanismům Oprava DNA, což vytváří neobvyklý vzor dědictví. Pozorovaný jev však byl (reverze mutovaných kopií HORKÁ HLAVA Gen do stavu divokého typu) byl později navržen jako artefakt, protože mutanti vykazují zvýšené křížení v důsledku fúze orgánů.[53][54][55]
Životní cyklus
Malá velikost rostliny a rychlý životní cyklus jsou také výhodné pro výzkum. Specializuje se jako jarní pomíjivost, bylo použito k nalezení několika laboratorních kmenů, které trvají asi šest týdnů od klíčení až po zralé semeno. Malá velikost rostliny je vhodná pro pěstování na malém prostoru a produkuje mnoho semen. Samoopalovací povaha této rostliny dále pomáhá genetickým experimentům. Také jako samostatná rostlina může vyprodukovat několik tisíc semen; každé z výše uvedených kritérií vede k A. thaliana být oceňován jako genetický modelový organismus.
Rozvoj
Vývoj květin
A. thaliana byl rozsáhle studován jako model pro vývoj květin. Vyvíjející se květina má čtyři základní orgány: sepals, okvětní lístky, tyčinky, a carpels (které se dále formují pestíky ). Tyto orgány jsou uspořádány do řady přeslenů: čtyři sepaly na vnějším přeslenu, následované čtyřmi okvětními lístky uvnitř tohoto, šest tyčinek a centrální oblast carpel. Homeotický mutace v A. thaliana mít za následek změnu jednoho orgánu na jiný - v případě agamous mutace, například, tyčinky se stanou okvětními lístky a carpels jsou nahrazeny novou květinou, což má za následek rekurzivně opakovaný vzor sepal-petal-petal.

Pozorování homeotických mutací vedla k formulaci ABC model vývoje květin podle E. Coen a E. Meyerowitz.[56] Podle tohoto modelu jsou geny pro identitu květinových orgánů rozděleny do tří tříd: geny třídy A (které ovlivňují sepaly a okvětní lístky), geny třídy B (které ovlivňují okvětní lístky a tyčinky) a geny třídy C (které ovlivňují tyčinky a plodolistky). Tyto geny kódují transkripční faktory které v průběhu vývoje způsobují specifikaci tkáně v příslušných oblastech. Ačkoli byl vyvinut studiem A. thaliana květiny, tento model je obecně použitelný pro ostatní kvetoucí rostliny.
Vývoj listů
Studie z A. thaliana poskytly značné poznatky, pokud jde o genetiku morfogeneze listů, zejména v dvouděložného typu rostliny.[57][58] Velká část porozumění pochází z analýzy mutantů ve vývoji listů, z nichž některé byly identifikovány v šedesátých letech minulého století, ale byly analyzovány genetickými a molekulárními technikami až v polovině 90. let. A. thaliana listy jsou velmi vhodné pro studium vývoje listů, protože jsou relativně jednoduché a stabilní.
Použitím A. thaliana, genetika za vývojem tvaru listů se stala jasnější a byla rozdělena do tří fází: Zahájení list primordium, založení dorziventrality, a vývoj marginální meristem. Listové primordie jsou iniciovány potlačením genů a proteinů třídy I. KNOX rodina (např VYSTŘELTE APIKÁLNÍ NEPROSTŘEDNĚ). Tyto proteiny třídy I KNOX přímo potlačují gibberellin biosyntéza v listovém primordiu. Bylo zjištěno, že na potlačení těchto I. třídy se podílí mnoho genetických faktorů KNOX geny v listových primordiích (např ASYMETRICKÉ LISTY1, BLADE-ON-PETIOLE1, SAWTOOTH1, atd.). S tímto potlačením se tedy zvyšuje hladina giberelinu a listový primordium iniciuje růst.
Zřízení listové dorziventrality je důležité, protože hřbetní (adaxiální) povrch listu se liší od ventrálního (abaxiálního) povrchu.[59]
Mikroskopie
A. thaliana je vhodný pro světelná mikroskopie analýza. Mladá sazenice celkově a zejména jejich kořeny jsou relativně průsvitné. To spolu s jejich malou velikostí usnadňuje zobrazování živých buněk pomocí obou fluorescence a konfokální laserová skenovací mikroskopie.[60] Nasazením sazenic za mokra do vody nebo do kultivačního média mohou být rostliny zobrazeny neinvazivně, což odstraňuje potřebu fixace a krájení a umožňující časová prodleva Měření.[61] Prostřednictvím mohou být zavedeny fluorescenční proteinové konstrukty proměna. The vývojový fázi každé buňky lze odvodit z jejího umístění v rostlině nebo pomocí fluorescenční protein markery, umožňující podrobné vývojová analýza.
Fyziologie
Snímání světla, emise světla a cirkadiánní biologie
Fotoreceptory fytochromy A, B, C, D a E zprostředkovávají fototropní reakci založenou na červeném světle. Pochopení funkce těchto receptorů pomohlo rostlinným biologům pochopit signální kaskády, které regulují fotoperiodismus, klíčení, deetiolace, a vyhýbání se stínu v rostlinách.
The UVR8 detekuje bílkoviny UV-B světlo a zprostředkovává reakci na tuto vlnovou délku poškozující DNA.
A. thaliana byl široce používán při studiu genetického základu fototropismus, chloroplast zarovnání a stomatální clona a další procesy ovlivněné modrým světlem.[62] Tyto vlastnosti reagují na modré světlo, které je vnímáno fototropin světelné receptory. Arabidopsis byla také důležitá pro pochopení funkcí dalšího receptoru modrého světla, kryptochrom, což je zvláště důležité pro unášení světla k ovládání rostlin cirkadiánní rytmy.[63] Když je nástup temnoty neobvykle brzo, A. thaliana snižuje jeho metabolismus škrobu o množství, které účinně vyžaduje divize.[64]
Světelné reakce byly dokonce nalezeny v kořenech, které byly dříve považovány za do značné míry necitlivé na světlo. Zatímco gravitropní odpověď uživatele A. thaliana kořenových orgánů je jejich převládající tropická odpověď, vzorky ošetřené mutageny a vybrané pro absenci gravitropního působení vykazovaly negativní fototropní reakci na modré nebo bílé světlo a pozitivní reakci na červené světlo, což naznačuje, že kořeny také vykazují pozitivní fototropismus.[65]
V roce 2000 Dr. Janet Braam z Rice University geneticky upraveno A. thaliana při dotyku zářit ve tmě. Efekt byl viditelný na ultravysokých kamerách.[66]
Několik úsilí, včetně Projekt Glowing Plant, se snažili použít A. thaliana zvýšit intenzitu luminiscence rostlin na komerčně životaschopné úrovně.
Na Měsíci
2. ledna 2019 v Číně Chang'e-4 přinesl přistávací modul A. thaliana na měsíc.[67] Malý mikrokosmos „cín“ v přistávacím modulu obsahoval A. thaliana, semena brambory, a bource morušového vejce. Protože rostliny by podporovaly bource morušového kyslíkem a bource morušového by zase dodávaly rostlinám potřebný oxid uhličitý a živiny prostřednictvím jejich odpadu,[68] vědci vyhodnotí, zda rostliny úspěšně fungují fotosyntéza a rostou a kvetou v měsíčním prostředí.[67]
Interakce rostlin a patogenů
Je důležité si uvědomit, jak rostliny dosahují rezistence k ochraně světové produkce potravin i zemědělství. Abychom lépe porozuměli interakcím mezi rostlinami a rostlinami, bylo vyvinuto mnoho modelových systémů bakteriální, plísňový, oomycete, virový, a hlístice patogeny. Arabidopsis thaliana byl mocným nástrojem pro studium subdisciplíny rostlinná patologie, tj. interakce mezi rostlinami a choroboplodnými chorobami patogeny.
| Typ patogenu | Příklad v Arabidopsis thaliana |
|---|---|
| Bakterie | Pseudomonas syringae, Xanthomonas campestris |
| Houby | Colletotrichum destructivum, Botrytis cinerea, Golovinomyces orontii |
| Oomycete | Hyaloperonospora arabidopsidis |
| Virový | Virus mozaiky květáku (CaMV), virus rajčatové mozaiky (TMV) |
| Hlístice | Meloidogyne incognita, Heterodera schachtii |

Schéma imunity spouštěné PAMP, konkrétně rozpoznávání bičíku FLS2 (vlevo nahoře), imunita spouštěná efektorem znázorněná rozpoznáním avrRpt2 pomocí RPS2 přes RIN4 (vpravo nahoře), mikroskopický pohled na ukládání kalózy v A. thaliana list (vlevo dole), příklad bez hypersenzitivní odpovědi (HR), horní a HR v A. thaliana listy (vpravo dole)

na kořenech Arabidopsis thaliana
a) Přehled an A. thaliana kořen (primární kořen) s mnoha kořenovými chloupky. b) Bakterie tvořící biofilm. c) Plísňové nebo oomycetové hyfy obklopující povrch kořene. d) Primární kořen hustě pokrytý výtrusy a protisty. e, f) Protisti, pravděpodobně patřící do třídy Bacillariophyceae. g) Bakterie a bakteriální vlákna. h, i) Různí bakteriální jedinci vykazující velké rozmanitosti tvarů a morfologických znaků.[69]
Použití A. thaliana vedlo k mnoha průlomům v rozvoji znalostí o tom, jak se rostliny projevují odolnost vůči chorobám rostlin. Důvodem, proč je většina rostlin rezistentních na většinu patogenů, je rezistence bez hostitele. To znamená, že ne všechny patogeny infikují všechny rostliny. Příklad kde A. thaliana byl použit ke stanovení genů odpovědných za nonhost rezistenci Blumeria graminis, původce padlí trav. A. thaliana mutanti byli vyvinuti pomocí mutagen ethylmethansulfonát a testovány k identifikaci mutantů se zvýšenou infekcí do B. graminis.[70][71][72] Mutanti s vyšší mírou infekce se kvůli schopnosti označují jako mutanti PEN B. graminis proniknout A. thaliana zahájit proces nemoci. Geny PEN byly později mapovány, aby identifikovaly geny odpovědné za odolnost vůči nonhost B. graminis.
Obecně platí, že když je rostlina vystavena patogenu, nebo nepatogenní mikrob, existuje počáteční reakce, známá jako imunita vyvolaná PAMP (PTI), protože rostlina detekuje konzervované motivy známé jako s patogeny spojené molekulární vzorce (PAMP).[73] Tyto PAMP jsou detekovány specializovanými receptory v hostiteli známém jako receptory rozpoznávání vzoru (PRR) na povrchu rostlinné buňky.
Nejlépe charakterizovaný PRR v A. thaliana je FLS2 (Flagellin-Sensing2), který rozpoznává bakteriální bičík,[74][75] specializovaná organela používaná mikroorganismy za účelem motility, stejně jako ligand flg22, který obsahuje 22 aminokyselin rozpoznávaných FLS2. Objev FLS2 byl usnadněn identifikací A. thaliana ekotyp, Ws-0, který nebyl schopen detekovat flg22, což vedlo k identifikaci genu kódujícího FLS2. FLS2 vykazuje nápadnou podobnost s rýží XA21, první PRR izolovanou v roce 1995
Druhý PRR, EF-Tu receptor (EFR), identifikovaný v A. thaliana, rozpoznává bakteriální EF-Tu protein, prokaryotický elongační faktor používaný v proteosyntéza, stejně jako laboratorně používaný ligand elf18.[76] Použitím Agrobacteriumzprostředkovaná transformace, technika, která využívá přirozeného procesu, kterým Agrobacterium přenáší geny do hostitelských rostlin, byl transformován gen EFR Nicotiana benthamiana rostlina tabáku, která nerozpoznává EF-Tu, čímž umožňuje rozpoznání bakteriálního EF-Tu[77] čímž se potvrzuje EFR jako receptor EF-Tu.
FLS2 i EFR používají podobné signální transdukce cesty k zahájení PTI. A. thaliana pomohl rozebrat tyto cesty, aby lépe porozuměl regulaci imunitních odpovědí, z nichž nejpozoruhodnější je mitogenem aktivovaná protein kináza (MAP kináza) kaskáda. Následné reakce PTI zahrnují callose depozice, oxidační výbuch a transkripce genů souvisejících s obranou.[78]
PTI je schopna bojovat proti patogenům nespecifickým způsobem. Silnější a specifičtější odezvou u rostlin je imunita vyvolaná efektorem (ETI). ETI závisí na rozpoznávání patogenních efektorů, proteinů vylučovaných patogenem, které mění funkce v hostiteli, rostlinou geny rezistence (R-geny), často popisován jako vztah gen-pro-gen. K tomuto rozpoznání může dojít přímo nebo nepřímo prostřednictvím strážného proteinu v hypotéze známé jako strážní hypotéza. Klonoval se první R gen A. thaliana byl RPS2 (odolnost vůči Pseudomonas syringae 2), který je odpovědný za rozpoznání efektoru avrRpt2.[79] Bakteriální efektor avrRpt2 je dodáván do A. thaliana přes Sekreční systém typu III z P. syringae pv rajče kmen DC3000. K rozpoznání avrRpt2 pomocí RPS2 dochází prostřednictvím strážného proteinu RIN4, který je štěpen. Rozpoznání efektoru patogenu vede k dramatické imunitní odpovědi známé jako hypersenzitivní reakce, ve kterém infikované rostlinné buňky podstupují buněčnou smrt, aby se zabránilo šíření patogenu.[80]
Systémová získaná rezistence (SAR) je dalším příkladem rezistence, který je u rostlin lépe pochopen díky výzkumu provedenému v A. thaliana. Benzothiadiazol (BTH), a kyselina salicylová (SA) analog, se historicky používá jako antifungální sloučenina v plodinách. Ukázalo se, že BTH stejně jako SA indukuje SAR v rostlinách. Zahájení dráhy SAR bylo poprvé prokázáno v roce A. thaliana ve kterém jsou zvýšené hladiny SA rozpoznávány žádným expresorem PR genů 1 (NPR1)[81] v důsledku redoxní změny v cytosolu, což má za následek snížení NPR1. NPR1, který obvykle existuje v multiplexním (oligomerním) stavu, se po redukci stává monomerním (jedna jednotka).[82] Když se NPR1 stane monomerním, je to přemístí do jádra, kde interaguje s mnoha TGA transkripční faktory, a je schopen indukovat geny související s patogeny, jako je PR1.[83] Dalším příkladem SAR by mohl být výzkum prováděný s transgenními rostlinami tabáku, které exprimují bakteriální salicyláthydroxylázu, gen nahG, vyžadují pro svou expresi akumulaci SA[84]
Evoluční aspekt rezistence rostlinných patogenů
Rostliny jsou ovlivněny několika patogeny po celou dobu jejich životnosti. V reakci na přítomnost patogenů si rostliny vyvinuly receptory na buněčném povrchu, aby detekovaly patogeny a reagovaly na ně.[85] Arabidopsis Thaliana je modelový organismus používaný ke stanovení specifických obranných mechanismů rezistence na rostlinné patogeny.[86] Tyto rostliny mají na svých buněčných površích speciální receptory, které umožňují detekci patogenů a iniciují mechanismy inhibující růst patogenů.[86] Obsahují dva receptory, FLS2 (bakteriální bičíkový receptor) a EF-Tu (bakteriální EF-Tu protein), které využívají signální transdukční dráhy k zahájení dráhy reakce na nemoc.[86] Cesta vede k rozpoznání patogenu, který způsobí, že infikované buňky podstoupí buněčnou smrt a zastaví šíření patogenu.[86] Rostliny s receptory FLS2 a EF-Tu prokázaly zvýšenou kondici v populaci.[84] To vedlo k přesvědčení, že rezistence vůči rostlinným patogenům je evoluční mechanismus, který se buduje po generace a reaguje na dynamická prostředí, jako je zvýšená predace a extrémní teploty.[84]
A. thaliana byl také použit ke studiu systémová získaná rezistence (SAR).[87]Tato cesta využívá benzothiadiazol, chemický induktor, k indukci transkripčních faktorů, mRNA, genů SAR. Tato akumulace transkripčních faktorů vede k inhibici genů souvisejících s patogeny.[87]
Interakce rostlin a patogenů jsou důležité pro pochopení toho, jak se rostliny vyvinuly v boji proti různým typům patogenů, které na ně mohou mít vliv.[84] Rozdíly v odolnosti rostlin napříč populacemi jsou způsobeny změnami faktorů prostředí. Rostliny, u nichž se vyvinula rezistence, ať už jde o obecnou variantu nebo variantu SAR, dokázaly žít déle a zadržovat nekrózu své tkáně (předčasné odumírání buněk), což vede k lepší adaptaci a vhodnosti pro populace, které se rychle nacházejí měnící se prostředí.[84]
Jiný výzkum
Probíhající výzkum Arabidopsis thaliana se provádí na Mezinárodní vesmírná stanice podle Evropská kosmická agentura. Cílem je studovat růst a reprodukci rostlin od semene k semenu v mikrogravitace.[88][89]
Zařízení „na čipu“, ve kterém A. thaliana tkáně lze kultivovat za podmínek in vitro.[90] Použití těchto zařízení může pomoci našemu porozumění vedení pylové trubice a mechanismu sexuální reprodukce v A. thaliana.
Samoopylení
A. thaliana je převážně samoopelivá rostlina s mírou křížení odhadovanou na méně než 0,3%.[91] Analýza genomového vzorce vazebné nerovnováhy naznačila, že samoopylení se vyvinulo zhruba před milionem let nebo více.[92] Je nepravděpodobné, že by meiózy, které vedou k samoopylení, měly významnou genetickou variabilitu. Tyto meiózy však mohou poskytnout adaptivní výhodu rekombinační opravy poškození DNA během tvorby zárodečných buněk v každé generaci.[Citace je zapotřebí ] Taková výhoda mohla stačit k tomu, aby umožňovala dlouhodobé přetrvávání meióz, i když následuje samooplodnění. Fyzikální mechanismus pro samoopylení v A. thaliana probíhá autogamií před syntézou, takže k oplodnění dochází převážně před otevřením květu.
Databáze a další zdroje
- TAIR a NASC: vybrané zdroje různých genetických a molekulárně biologických informací, odkazy na genovou expresi databáze, atd.
- Centrum biologických zdrojů Arabidopsis (Zásoby osiva a DNA)
- Akciové centrum v Nottinghamu Arabidopsis (Zásoby osiva a DNA)
Viz také
Reference
- ^ Warwick SI, Francis A, Al-Shehbaz IA (2016). "Kontrolní seznam a databáze druhů Brassicaceae". Species 2000 & ITIS Katalog života (26 ed.). ISSN 2405-8858.
- ^ "Arabidopsis thaliana". Informační síť zdrojů Germplasm (ÚSMĚV). Služba zemědělského výzkumu (ARS), Ministerstvo zemědělství USA (USDA). Citováno 11. prosince 2017.
- ^ Hoffmann MH (2002). „Biogeografie Arabidopsis thaliana (L.) Heynh. (Brassicaceae)“. Časopis biogeografie. 29: 125–134. doi:10.1046 / j.1365-2699.2002.00647.x.
- ^ Mitchell-Olds T (prosinec 2001). „Arabidopsis thaliana a její divocí příbuzní: modelový systém pro ekologii a evoluci“. Trendy v ekologii a evoluci. 16 (12): 693–700. doi:10.1016 / s0169-5347 (01) 02291-1.
- ^ Sharbel TF, Haubold B, Mitchell-Olds T (2000). „Genetická izolace podle vzdálenosti u Arabidopsis thaliana: biogeografie a postglaciální kolonizace Evropy“. Molekulární ekologie. 9 (12): 2109–2118. doi:10.1046 / j.1365-294x. 2000.01122.x. PMID 11123622. S2CID 1788832.
- ^ A b Krämer U (březen 2015). „Výsadba molekulárních funkcí v ekologickém kontextu s Arabidopsis thaliana“. eLife. 4: –06100. doi:10,7554 / eLife.06100. PMC 4373673. PMID 25807084.
- ^ Durvasula A, Fulgione A, Gutaker RM, Alacakaptan SI, Flood PJ, Neto C, Tsuchimatsu T, Burbano HA, Picó FX, Alonso-Blanco C, Hancock AM (květen 2017). "Arabidopsis thaliana". Sborník Národní akademie věd Spojených států amerických. 114 (20): 5213–5218. doi:10.1073 / pnas.1616736114. PMC 5441814. PMID 28473417.
- ^ A b "Shromáždění genomu". Informační zdroj Arabidopsis. Citováno 29. března 2016.
- ^ Flóra NW Evropy: Arabidopsis thaliana Archivováno 8. Prosince 2007 v Wayback Machine
- ^ Blamey, M. & Gray-Wilson, C. (1989). Flóra Británie a severní Evropy. ISBN 0-340-40170-2
- ^ Flóra Pákistánu: Arabidopsis thaliana
- ^ Flóra Číny: Arabidopsis thaliana
- ^ López-Bucio J, Campos-Cuevas JC, Hernández-Calderón E, Velásquez-Becerra C, Farías-Rodríguez R, Macías-Rodríguez LI, Valencia-Cantero E (únor 2007). „Bacillus megaterium rhizobacteria podporuje růst a mění architekturu kořenového systému prostřednictvím signálního mechanismu nezávislého na auxinu a ethylenu u Arabidopsis thaliana“. Molekulární interakce rostlin a mikrobů. 20 (2): 207–17. doi:10.1094 / MPMI-20-2-0207. PMID 17313171.
- ^ Meinke DW, Cherry JM, Dean C, Rounsley SD, Koornneef M (říjen 1998). „Arabidopsis thaliana: modelová rostlina pro analýzu genomu“. Věda. 282 (5389): 662, 679–82. Bibcode:1998Sci ... 282..662M. CiteSeerX 10.1.1.462.4735. doi:10.1126 / science.282.5389.662. PMID 9784120.
- ^ A b Konsorcium 1001 genomů (červenec 2016). "1 135 genomů odhaluje globální vzor polymorfismu v roce 2006 Arabidopsis thaliana". Buňka. 166 (2): 481–491. doi:10.1016 / j.cell.2016.05.063. PMC 4949382. PMID 27293186.
- ^ "Arabidopsis thaliana (L.) Heynh". www.gbif.org. Citováno 8. prosince 2018.
- ^ Hedberg, Olov (1957). „Cévní rostliny afroalpinu: taxonomická revize“. Acta Universitatis Upsaliensis: Symbolae Botanicae Upsalienses. 15 (1): 1–144.
- ^ Fulgione A, Hancock AM (září 2018). „Archaické linie rozšiřují náš pohled na historii Arabidopsis thaliana“. Nový fytolog. 219 (4): 1194–1198. doi:10.1111 / nph.15244. PMID 29862511.
- ^ A b "Arabidopsis thaliana - přehled". Encyklopedie života.
- ^ Exposito-Alonso M, Becker C, Schuenemann VJ, Reiter E, Setzer C, Slovak R, Brachi B, Hagmann J, Grimm DG, Chen J, Busch W, Bergelson J, Ness RW, Krause J, Burbano HA, Weigel D ( Února 2018). „Míra a potenciální význam nových mutací v linii kolonizujících rostlin“. Genetika PLOS. 14 (2): e1007155. doi:10.1371 / journal.pgen.1007155. PMC 5825158. PMID 29432421.
- ^ "Arabidopsis thaliana (řeřicha thale)". Zahrady Kew.
- ^ "Státní a federální seznam škodlivých plevelů | USDA ROSTLINY". rostliny.sc.egov.usda.gov. Citováno 8. prosince 2018.
- ^ „IRMNG“. Encyklopedie života. Archivovány od originál dne 1. dubna 2018.
- ^ [1] TAIR: O Arabidopsis
- ^ Rensink WA, Buell CR (červen 2004). „Arabidopsis na rýži. Uplatňování poznatků z plevelů pro lepší pochopení druhu plodiny“. Fyziologie rostlin. 135 (2): 622–9. doi:10.1104 / pp.104.040170. PMC 514098. PMID 15208410.
- ^ Coelho SM, Peters AF, Charrier B, Roze D, Destombe C, Valero M, Cock JM (prosinec 2007). „Složité životní cykly mnohobuněčných eukaryot: nové přístupy založené na použití modelových organismů“. Gen. 406 (1–2): 152–70. doi:10.1016 / j.gene.2007.07.025. PMID 17870254.
- ^ Platt A, Horton M, Huang YS, Li Y, Anastasio AE, Mulyati NW, Agren J, Bossdorf O, Byers D, Donohue K, Dunning M, Holub EB, Hudson A, Le Corre V, Loudet O, Roux F, Warthmann N, Weigel D, Rivero L, Scholl R, Nordborg M, Bergelson J, Borevitz JO (únor 2010). Novembre J (ed.). „Rozsah struktury populace v Arabidopsis thaliana“. Genetika PLOS. 6 (2): e1000843. doi:10.1371 / journal.pgen.1000843. PMC 2820523. PMID 20169178.
- ^ Yanofsky MF, Ma H, Bowman JL, Drews GN, Feldmann KA, Meyerowitz EM (červenec 1990). „Protein kódovaný agamem homeotického genu Arabidopsis se podobá transkripčním faktorům“. Příroda. 346 (6279): 35–9. Bibcode:1990 Natur.346 ... 35Y. doi:10.1038 / 346035a0. PMID 1973265. S2CID 4323431.
- ^ A b Meyerowitz EM (leden 2001). "Prehistorie a historie výzkumu Arabidopsis". Fyziologie rostlin. 125 (1): 15–9. doi:10.1104 / pp.125.1.15. PMC 1539315. PMID 11154286.
- ^ Lloyd AM, Barnason AR, Rogers SG, Byrne MC, Fraley RT, Horsch RB (říjen 1986). "Transformace Arabidopsis thaliana pomocí Agrobacterium tumefaciens". Věda. 234 (4775): 464–6. Bibcode:1986Sci ... 234..464L. doi:10.1126 / science.234.4775.464. PMID 17792019. S2CID 22125701.
- ^ Chang C, Meyerowitz EM (březen 1986). "Molekulární klonování a sekvence DNA genu pro alkohol dehydrogenázu Arabidopsis thaliana". Sborník Národní akademie věd Spojených států amerických. 83 (5): 1408–12. Bibcode:1986PNAS ... 83.1408C. doi:10.1073 / pnas.83.5.1408. PMC 323085. PMID 2937058.
- ^ A b "Arabidopsis thaliana chloroplast, kompletní genom - přístupové číslo NCBI NC_000932.1 ". Národní centrum pro biotechnologické informace. Citováno 4. listopadu 2018.
- ^ A b Sato S, Nakamura Y, Kaneko T, Asamizu E, Tabata S (1999). "Kompletní struktura chloroplastového genomu Arabidopsis thaliana". Výzkum DNA. 6 (5): 283–290. doi:10.1093 / dnares / 6.5.283. ISSN 1340-2838. PMID 10574454.
- ^ Bennett MD, Leitch IJ, Price HJ, Johnston JS (duben 2003). „Srovnání s Caenorhabditis (přibližně 100 Mb) a Drosophila (přibližně 175 Mb) pomocí průtokové cytometrie ukazují, že velikost genomu u Arabidopsis je přibližně 157 Mb, a tedy přibližně o 25% větší než odhad iniciativy genomů Arabidopsis přibližně 125 Mb.“. Annals of Botany. 91 (5): 547–57. doi:10.1093 / aob / mcg057. PMC 4242247. PMID 12646499.
- ^ (Leutwileret al., 1984). V našem průzkumu Arabidopsis ...
- ^ Fleischmann A, Michael TP, Rivadavia F, Sousa A, Wang W, Temsch EM, Greilhuber J, Müller KF, Heubl G (prosinec 2014). „Vývoj velikosti genomu a počtu chromozomů v rodu masožravých rostlin Genlisea (Lentibulariaceae), s novým odhadem minimální velikosti genomu v krytosemenných rostlinách“. Annals of Botany. 114 (8): 1651–63. doi:10.1093 / aob / mcu189. PMC 4649684. PMID 25274549.
- ^ Arabidopsis Genome Initiative (prosinec 2000). „Analýza genomové sekvence kvetoucí rostliny Arabidopsis thaliana“. Příroda. 408 (6814): 796–815. Bibcode:2000Natur.408..796T. doi:10.1038/35048692. PMID 11130711.
- ^ „TAIR - Anotace genomu“.
- ^ "Integr8 - A.thaliana Genome Statistics".
- ^ Bundy JG, Davey MP, Viant MR (2009). „Metabolomika životního prostředí: kritická revize a budoucí perspektivy. (Pozvaná revize)“. Metabolomika. 5 (3–21): 3–21. doi:10.1007 / s11306-008-0152-0. S2CID 22179989.
- ^ Lake JA, Field KJ, Davey MP, Beerling DJ, Lomax BH (2009). „Metabolomické a fyziologické reakce odhalují vícefázovou aklimatizaci Arabidopsis thaliana na chronické UV záření“. Rostlina, buňka a životní prostředí. 32 (10): 1377–1389. doi:10.1111 / j.1365-3040.2009.02005.x. PMID 19558413.
- ^ "Arabidopsis thaliana ekotyp mitochondrie Col-0, kompletní genom - přístupové číslo NCBI BK010421 ". Národní centrum pro biotechnologické informace. Citováno 10. dubna 2019.
- ^ Klein M, Eckert-Ossenkopp U, Schmiedeberg I, Brandt P, Unseld M, Brennicke A, Schuster W (1994). "Fyzické mapování mitochondriálního genomu Arabidopsis thaliana pomocí kosmidových a YAC klonů". Plant Journal. 6 (3): 447–455. doi:10.1046 / j.1365-313X.1994.06030447.x. PMID 7920724.
- ^ Gualberto JM, Mileshina D, Wallet C, Niazi AK, Weber-Lotfi F, Dietrich A (2014). "The plant mitochondrial genome: dynamics and maintenance". Biochimie. 100: 107–120. doi:10.1016/j.biochi.2013.09.016. PMID 24075874.
- ^ Clough SJ, Bent AF (December 1998). "Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana". The Plant Journal. 16 (6): 735–43. doi:10.1046/j.1365-313x.1998.00343.x. PMID 10069079.
- ^ Zhang X, Henriques R, Lin SS, Niu QW, Chua NH (2006). "Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method". Nature Protocols. 1 (2): 641–6. doi:10.1038/nprot.2006.97. PMID 17406292. S2CID 6906570.
- ^ A b NASC-Nottingham Arabidopsis Stock Center - http://arabidopsis.info
- ^ Magliano TM, Botto JF, Godoy AV, Symonds VV, Lloyd AM, Casal JJ (June 2005). "New Arabidopsis recombinant inbred lines (Landsberg erecta x Nossen) reveal natural variation in phytochrome-mediated responses". Fyziologie rostlin. 138 (2): 1126–35. doi:10.1104/pp.104.059071. PMC 1150426. PMID 15908601.
- ^ The Arabidopsis Biological Resource Center (ABRC), http://abrc.osu.edu
- ^ NASC-Nottingham Arabidopsis Stock Center-Background Lines-Description- http://arabidopsis.info/CollectionInfo?id=94
- ^ Hahn F, Mantegazza O, Greiner A, Hegemann P, Eisenhut M, Weber AP (2017). "Arabidopsis thaliana". Hranice ve vědě o rostlinách. 8: 39. doi:10.3389/fpls.2017.00039. PMC 5258748. PMID 28174584.
- ^ Hahn F, Eisenhut M, Mantegazza O, Weber AP (5 April 2018). "Arabidopsis With Cas9-Based Gene Targeting". Hranice ve vědě o rostlinách. 9: 424. doi:10.3389/fpls.2018.00424. PMC 5895730. PMID 29675030.
- ^ Lolle SJ, Victor JL, Young JM, Pruitt RE (March 2005). "Genome-wide non-mendelian inheritance of extra-genomic information in Arabidopsis". Příroda. 434 (7032): 505–9. Bibcode:2005Natur.434..505L. doi:10.1038/nature03380. PMID 15785770. S2CID 1352368.Washington Post summary.
- ^ Peng P, Chan SW, Shah GA, Jacobsen SE (September 2006). "Plant genetics: increased outcrossing in hothead mutants". Příroda. 443 (7110): E8, discussion E8–9. Bibcode:2006Natur.443E...8P. doi:10.1038/nature05251. PMID 17006468. S2CID 4420979.
- ^ Pennisi E (September 2006). "Genetics. Pollen contamination may explain controversial inheritance". Věda. 313 (5795): 1864. doi:10.1126/science.313.5795.1864. PMID 17008492. S2CID 82215542.
- ^ Coen ES, Meyerowitz EM (September 1991). "The war of the whorls: genetic interactions controlling flower development". Příroda. 353 (6339): 31–7. Bibcode:1991Natur.353...31C. doi:10.1038/353031a0. PMID 1715520. S2CID 4276098.
- ^ Tsukaya H (7 June 2013). "Leaf development". The Arabidopsis Book. 11: e0163. doi:10.1199/tab.0163. PMC 3711357. PMID 23864837.
- ^ Turner S, Sieburth LE (22 March 2003). "Vascular patterning". The Arabidopsis Book. 2: e0073. doi:10.1199/tab.0073. PMC 3243335. PMID 22303224.
- ^ Efroni I, Eshed Y, Lifschitz E (April 2010). "Morphogenesis of simple and compound leaves: a critical review". Rostlinná buňka. 22 (4): 1019–32. doi:10.1105/tpc.109.073601. PMC 2879760. PMID 20435903.
- ^ Moreno N, Bougourd S, Haseloff J and Fiejo JA. 2006. Chapter 44: Imaging Plant Cells. In: Pawley JB (Editor). Handbook of Biological Confocal Microscopy - 3rd edition. SpringerScience+Business Media, New York. p769-787
- ^ Shaw SL (February 2006). "Imaging the live plant cell". The Plant Journal. 45 (4): 573–98. doi:10.1111/j.1365-313X.2006.02653.x. PMID 16441350.
- ^ Sullivan JA, Deng XW (August 2003). "From seed to seed: the role of photoreceptors in Arabidopsis development". Vývojová biologie. 260 (2): 289–97. doi:10.1016/S0012-1606(03)00212-4. PMID 12921732.
- ^ Más P (2005). "Circadian clock signaling in Arabidopsis thaliana: from gene expression to physiology and development". International Journal of Developmental Biology. 49 (5–6): 491–500. doi:10.1387/ijdb.041968pm. PMID 16096959.
- ^ Scialdone A, Mugford ST, Feike D, Skeffington A, Borrill P, Graf A, Smith AM, Howard M (June 2013). "Arabidopsis plants perform arithmetic division to prevent starvation at night". eLife. 2: e00669. arXiv:1306.5148. doi:10.7554/eLife.00669. PMC 3691572. PMID 23805380.
- ^ Ruppel NJ, Hangarter RP, Kiss JZ (February 2001). "Red-light-induced positive phototropism in Arabidopsis roots". Planta. 212 (3): 424–30. doi:10.1007/s004250000410. PMID 11289607. S2CID 28410755.
- ^ "Plants that Glow in the Dark", Bioresearch Online, 18 May 2000
- ^ A b Letzter, Rafi (4 January 2019). "There Are Plants and Animals on the Moon Now (Because of China)". ProfoundSpace.org. Citováno 15. ledna 2019.
- ^ Connor, Neil (13 April 2018). "China plans to grow flowers and silkworms on the dark side of the moon". The Telegraph. ISSN 0307-1235. Citováno 15. ledna 2019.
- ^ Hassani, M.A., Durán, P. and Hacquard, S. (2018) "Microbial interactions within the plant holobiont". Mikrobiom, 6(1): 58. doi:10.1186/s40168-018-0445-0.
 Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0. - ^ Collins NC, Thordal-Christensen H, Lipka V, Bau S, Kombrink E, Qiu JL, Hückelhoven R, Stein M, Freialdenhoven A, Somerville SC, Schulze-Lefert P (October 2003). "SNARE-protein-mediated disease resistance at the plant cell wall". Příroda. 425 (6961): 973–7. Bibcode:2003Natur.425..973C. doi:10.1038/nature02076. PMID 14586469. S2CID 4408024.
- ^ Lipka V, Dittgen J, Bednarek P, Bhat R, Wiermer M, Stein M, Landtag J, Brandt W, Rosahl S, Scheel D, Llorente F, Molina A, Parker J, Somerville S, Schulze-Lefert P (November 2005). "Pre- and postinvasion defenses both contribute to nonhost resistance in Arabidopsis". Věda. 310 (5751): 1180–3. Bibcode:2005Sci...310.1180L. doi:10.1126/science.1119409. hdl:11858/00-001M-0000-0012-3A32-0. PMID 16293760. S2CID 35317665.
- ^ Stein M, Dittgen J, Sánchez-Rodríguez C, Hou BH, Molina A, Schulze-Lefert P, Lipka V, Somerville S (March 2006). "Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to nonhost resistance to inappropriate pathogens that enter by direct penetration". Rostlinná buňka. 18 (3): 731–46. doi:10.1105/tpc.105.038372. PMC 1383646. PMID 16473969.
- ^ Knepper C, Day B (March 2010). "From perception to activation: the molecular-genetic and biochemical landscape of disease resistance signaling in plants". The Arabidopsis Book. 8: e012. doi:10.1199/tab.0124. PMC 3244959. PMID 22303251.
- ^ Gómez-Gómez L, Felix G, Boller T (May 1999). "A single locus determines sensitivity to bacterial flagellin in Arabidopsis thaliana". The Plant Journal. 18 (3): 277–84. doi:10.1046/j.1365-313X.1999.00451.x. PMID 10377993.
- ^ Gómez-Gómez L, Boller T (June 2000). "FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis". Molekulární buňka. 5 (6): 1003–11. doi:10.1016/S1097-2765(00)80265-8. PMID 10911994.
- ^ Zipfel C, Kunze G, Chinchilla D, Caniard A, Jones JD, Boller T, Felix G (May 2006). "Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation". Buňka. 125 (4): 749–60. doi:10.1016/j.cell.2006.03.037. PMID 16713565. S2CID 6856390.
- ^ Lacombe S, Rougon-Cardoso A, Sherwood E, Peeters N, Dahlbeck D, van Esse HP, Smoker M, Rallapalli G, Thomma BP, Staskawicz B, Jones JD, Zipfel C (April 2010). "Interfamily transfer of a plant pattern-recognition receptor confers broad-spectrum bacterial resistance". Přírodní biotechnologie. 28 (4): 365–9. doi:10.1038/nbt.1613. PMID 20231819. S2CID 7260214.,
- ^ Zhang J, Zhou JM (September 2010). "Plant immunity triggered by microbial molecular signatures". Molecular Plant. 3 (5): 783–93. doi:10.1093/mp/ssq035. PMID 20713980.
- ^ Kunkel BN, Bent AF, Dahlbeck D, Innes RW, Staskawicz BJ (August 1993). "RPS2, an Arabidopsis disease resistance locus specifying recognition of Pseudomonas syringae strains expressing the avirulence gene avrRpt2". Rostlinná buňka. 5 (8): 865–75. doi:10.1105/tpc.5.8.865. PMC 160322. PMID 8400869.
- ^ Axtell MJ, Staskawicz BJ (Únor 2003). "Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4". Buňka. 112 (3): 369–77. doi:10.1016/S0092-8674(03)00036-9. PMID 12581526. S2CID 1497625.
- ^ Cao H, Bowling SA, Gordon AS, Dong X (November 1994). "Characterization of an Arabidopsis Mutant That Is Nonresponsive to Inducers of Systemic Acquired Resistance". Rostlinná buňka. 6 (11): 1583–1592. doi:10.1105/tpc.6.11.1583. PMC 160545. PMID 12244227.
- ^ Mou Z, Fan W, Dong X (June 2003). "Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes". Buňka. 113 (7): 935–44. doi:10.1016/S0092-8674(03)00429-X. PMID 12837250. S2CID 1562690.
- ^ Johnson C, Boden E, Arias J (August 2003). "Salicylic acid and NPR1 induce the recruitment of trans-activating TGA factors to a defense gene promoter in Arabidopsis". Rostlinná buňka. 15 (8): 1846–58. doi:10.1105/tpc.012211. PMC 167174. PMID 12897257.
- ^ A b C d E Delaney TP, Uknes S, Vernooij B, Friedrich L, Weymann K, Negrotto D, Gaffney T, Gut-Rella M, Kessmann H, Ward E, Ryals J (November 1994). "A central role of salicylic Acid in plant disease resistance". Věda. 266 (5188): 1247–50. Bibcode:1994Sci...266.1247D. doi:10.1126/science.266.5188.1247. PMID 17810266. S2CID 15507678.
- ^ Bent AF, Kunkel BN, Dahlbeck D, Brown KL, Schmidt R, Giraudat J, Leung J, Staskawicz BJ (Září 1994). "RPS2 of Arabidopsis thaliana: a leucine-rich repeat class of plant disease resistance genes". Věda. 265 (5180): 1856–60. Bibcode:1994Sci...265.1856B. doi:10.1126/science.8091210. PMID 8091210.
- ^ A b C d Zipfel C, Robatzek S, Navarro L, Oakeley EJ, Jones JD, Felix G, Boller T (April 2004). "Bacterial disease resistance in Arabidopsis through flagellin perception". Příroda. 428 (6984): 764–7. Bibcode:2004Natur.428..764Z. doi:10.1038/nature02485. PMID 15085136. S2CID 4332562.
- ^ A b Lawton K, Friedrich L, Hunt M (1996). "Benzothiadizaole induces disease resistance by a citation of the systemic acquired resistance signal transduction pathway". The Plant Journal. 10 (1): 71–82. doi:10.1046/j.1365-313x.1996.10010071.x. PMID 8758979.
- ^ Link BM, Busse JS, Stankovic B (2014). "Seed-to-Seed-to-Seed Growth and Development of Arabidopsis in Microgravity". Astrobiologie. 14 (10): 866–875. Bibcode:2014AsBio..14..866L. doi:10.1089/ast.2014.1184. PMC 4201294. PMID 25317938.
- ^ Ferl RJ, Paul AL (April 2010). "Lunar plant biology--a review of the Apollo era". Astrobiologie. 10 (3): 261–74. Bibcode:2010AsBio..10..261F. doi:10.1089/ast.2009.0417. PMID 20446867.
- ^ Yetisen AK, Jiang L, Cooper JR, Qin Y, Palanivelu R, Zohar Y (May 2011). "A microsystem-based assay for studying pollen tube guidance in plant reproduction". J. Micromech. Microeng. 25 (5): 054018. Bibcode:2011JMiMi..21e4018Y. doi:10.1088/0960-1317/21/5/054018.CS1 maint: používá parametr autoři (odkaz)
- ^ Abbott RJ, Gomes MF (1989). "Population genetic structure and outcrossing rate of Arabidopsis thaliana (L.) Heynh". Dědičnost. 62 (3): 411–418. doi:10.1038/hdy.1989.56.
- ^ Tang C, Toomajian C, Sherman-Broyles S, Plagnol V, Guo YL, Hu TT, Clark RM, Nasrallah JB, Weigel D, Nordborg M (August 2007). "The evolution of selfing in Arabidopsis thaliana". Věda. 317 (5841): 1070–2. Bibcode:2007Sci...317.1070T. doi:10.1126/science.1143153. PMID 17656687. S2CID 45853624.
externí odkazy
- Arabidopsis transcriptional regulatory map
- The Arabidopsis Information Resource (TAIR)
- Salk Institute Genomic Analysis Laboratory
- What Makes Plants Grow? The Arabidopsis genome knows Featured article in Genome News Network
- The Arabidopsis book - A comprehensive review published yearly related to research in Arabidopsis
- A. thaliana protein abundance
- The Arabidopsis Information Portal (Araport)
| Kontrolní úřad |
|---|