Protocell - Protocell

A protocell (nebo protobiont) je samoorganizovaná, endogenně uspořádaná sférická sbírka lipidy navrhl jako odrazový můstek směrem k původ života.[1][2] Ústřední otázka v vývoj je to, jak jednoduché protokoly nejprve vznikly a jak se mohou lišit v reprodukčním výstupu, což umožňuje akumulaci nových biologických objevů v čase, tj. biologická evoluce. Přestože v laboratorním prostředí dosud nebyl dosažen funkční protokol, cíl porozumět procesu se zdá být na dosah.[3][4][5][6]
Přehled
Rozdělení bylo[když? ] důležité v počátcích života.[Citace je zapotřebí ] Membrány tvoří uzavřené oddíly, které jsou oddělené od vnějšího prostředí, a poskytují tak buňce funkčně specializované vodné prostory. Protože lipidová dvojvrstva membrán je pro většinu nepropustná hydrofilní molekuly (rozpuštěné ve vodě), mají buňky membránové transportní systémy, které zajišťují dovoz výživných molekul i vývoz odpadu.[7][potřebujete nabídku k ověření ]Je velmi náročné konstruovat protokoly z molekulárních sestav. Důležitým krokem v této výzvě je dosažení dynamiky vezikul, které jsou relevantní pro buněčné funkce, jako je membránový transport a autoreprodukce, za použití amfifilních molekul. Na primitivní Zemi vytvořily složky života četné chemické reakce organických sloučenin.[Citace je zapotřebí ] Z těchto látek mohou být amfifilní molekuly prvním hráčem ve vývoji od molekulárního shromáždění k buněčnému životu.[8][9] Krokem od vezikul k protokolům může být vývoj samoreprodukujících se vezikul spolu s metabolickým systémem.[10]
Selektivita pro rozčlenění

Samostatně sestavené vezikuly jsou základními složkami primitivních buněk.[1] The druhý zákon termodynamiky vyžaduje, aby se vesmír pohyboval ve směru, ve kterém porucha (nebo entropie ) se zvyšuje, ale život se vyznačuje velkou mírou organizace. Proto je potřeba oddělit hranici životní procesy z neživé hmoty.[11] The buněčná membrána je jedinou buněčnou strukturou, která se nachází ve všech buňkách všech organismů na Zemi.[12]
Výzkumní pracovníci Irene A. Chen a Jack W. Szostak (Nobelova cena za fyziologii nebo medicínu 2009) mimo jiné prokázala, že jednoduché fyzikálně-chemické vlastnosti elementárních protokolů mohou vést k jednodušším koncepčním analogům základního buněčného chování, včetně primitivních forem darwinovské konkurence a ukládání energie. Takové kooperativní interakce mezi membránou a zapouzdřeným obsahem by mohly výrazně zjednodušit přechod z replikujících se molekul do skutečných buněk.[4] Konkurence o membránové molekuly by upřednostňovala stabilizované membrány, což naznačuje selektivní výhodu pro vývoj zesítěných mastných kyselin a dokonce fosfolipidy dneška.[4] Tento mikroenkapsulace povoleno pro metabolismus uvnitř membrány, výměna malých molekul a zabránění průchodu velkých látek přes ni.[13] Mezi hlavní výhody zapouzdření patří zvýšená rozpustnost nákladu a vytváření energie ve formě chemického gradientu. Energie se tak často říká, že ji akumuluje buňky ve strukturách molekul látek jako např sacharidy (včetně cukrů), lipidy, a bílkoviny, které uvolňují energii, když jsou chemicky kombinovány s kyslík v době buněčné dýchání.[14][15]
Energetický gradient
Studie laboratoře Jet Propulsion Laboratory NASA z března 2014 prokázala jedinečný způsob studia původu života: palivové články.[16] Palivové články jsou podobné biologickým buňkám v tom, že elektrony jsou také přenášeny do az molekul. V obou případech to vede k elektřině a energii. Studie uvádí, že jedním důležitým faktorem bylo, že Země poskytuje elektrickou energii na mořském dně. „Tato energie mohla nastartovat život a mohla udržet život i po jeho vzniku. Nyní máme způsob testování různých materiálů a prostředí, které by mohly pomoci životu vzniknout nejen na Zemi, ale možná i na Mars, Evropa a na dalších místech v Sluneční Soustava."[16]
Vezikuly, micely a kapičky bez membrány
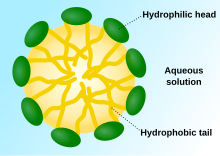
Když fosfolipidy jsou umístěny ve vodě, molekuly se spontánně uspořádají tak, že ocasy jsou chráněny před vodou, což vede k tvorbě membránových struktur, jako jsou dvojvrstvy, vezikuly a micely.[2] V moderních buňkách vezikuly jsou zapojeni do metabolismus doprava, kontrola vztlaku,[17] a enzym úložný prostor. Mohou také působit jako komory pro přirozenou chemickou reakci. Typický váček nebo micela v vodný roztok tvoří agregát s hydrofilní „hlavové“ oblasti v kontaktu s okolím solventní, zabavující hydrofobní oblasti jednoho ocasu ve středu micely. Tato fáze je způsobena chování balení jednoho ocasu lipidy v dvouvrstvá. Přestože se jedná o protocelulární proces samo-sestavování, který spontánně tvoří lipid jednovrstvá vezikuly a micely v přírodě připomínají druhy prvotních vezikul nebo protocelul, které mohly existovat na začátku evoluce, nejsou tak sofistikované jako dvouvrstvá membrány dnešních živých organismů.[18]
Spíše než aby byly tvořeny fosfolipidy, mohly se časné membrány tvořit z monovrstev nebo dvouvrstev mastné kyseliny, které se snadněji vytvořily v prebiotickém prostředí.[19] Mastné kyseliny byly syntetizovány v laboratořích za různých prebiotických podmínek a byly nalezeny na meteoritech, což naznačuje jejich přirozenou syntézu v přírodě.[4]
Kyselina olejová vezikuly představují dobré modely membránových protocelul, které mohly existovat v prebiotických dobách.[20]
Elektrostatické interakce vyvolané krátkými, pozitivně nabitými hydrofobními peptidy obsahujícími 7 nebo více aminokyselin mohou připojit RNA k membráně vezikul, základní buněčné membráně.[21][22]
Geotermální rybníky a jíl
Vědci naznačili, že život začal v hydrotermální průduchy v hlubokém moři, ale studie z roku 2012 naznačuje, že vnitrozemské bazény kondenzovaných a chlazených geotermálních par mají ideální vlastnosti pro vznik života.[23] Závěr je založen hlavně na chemii moderních buněk, kde je cytoplazma bohatá na ionty draslíku, zinku, manganu a fosfátů, které nejsou v mořském prostředí příliš rozšířené. Vědci tvrdí, že takové podmínky se vyskytují pouze tam, kde horká hydrotermální tekutina přináší ionty na povrch - místa jako gejzíry bahenné hrnce, fumaroly a další geotermální funkce. V těchto dýmavých a bublajících povodích se voda naložená ionty zinku a manganu mohla shromažďovat, ochlazovat a kondenzovat v mělkých bazénech.[23]
Ukázala to další studie z 90. let montmorillonit jíl může pomoci vytvořit RNA řetězce až 50 nukleotidů, které se spontánně spojily do jedné molekuly RNA.[5] Později, v roce 2002, bylo zjištěno, že přidáním montmorillonitu do roztoku mastné kyseliny micely (lipidové kuličky), jíl zrychlil rychlost váček stokrát.[5]
Výzkum ukázal, že některé minerály mohou katalyzovat postupnou tvorbu uhlovodíkových zbytků mastné kyseliny z plynů vodíku a oxidu uhelnatého - plyny, ze kterých se mohly uvolnit hydrotermální průduchy nebo gejzíry. Mastné kyseliny různých délek se nakonec uvolní do okolní vody,[19] ale tvorba vezikul vyžaduje vyšší koncentraci mastných kyselin, proto se předpokládá, že tvorba protocellu začala u pozemních hydrotermálních průduchů, jako je gejzíry bahenné hrnce, fumaroly a další geotermální vlastnosti, při kterých se voda odpařuje a koncentruje rozpuštěnou látku.[5][24][25]
Montmorillonitové bubliny
Další skupina naznačuje, že primitivní buňky se mohly tvořit uvnitř anorganických jílovitých mikrokompartmentů, které mohou poskytnout ideální obal pro syntézu a rozčlenění komplexních organických molekul.[26] Hliněné obrněné bubliny přirozeně se tvoří, když částice montmorillonit jíl se hromadí na vnějším povrchu vzduchových bublin pod vodou. Tím se vytvoří polopropustný vezikul z materiálů, které jsou snadno dostupné v prostředí. Autoři poznamenávají, že je známo, že montmorillonit slouží jako chemický katalyzátor, který podporuje lipidy ve vytváření membrán a jednotlivé nukleotidy, aby se spojily do řetězců RNA. Primitivní reprodukci si lze představit, když prasknou hliněné bubliny a uvolní produkt vázaný na lipidovou membránu do okolního média.[26]
Membránové kapičky
Dalším způsobem, jak vytvořit primitivní kompartmenty, které mohou vést k tvorbě protocell, jsou polyesterové membránové struktury, které mají schopnost hostit biochemické látky (proteiny a RNA) a / nebo lešení kolem lipidů.[27][28] I když jsou tyto kapičky netěsné vůči genetickým materiálům, tato netěsnost mohla usnadnit hypotézu progenotů.[29]
Membránový transport

Pro buněčné organismy je transport specifických molekul přes rozdělovací membránové bariéry nezbytný pro výměnu obsahu s jejich prostředím a s jinými jedinci. Například umožňuje výměna obsahu mezi jednotlivci horizontální přenos genů, důležitý faktor ve vývoji buněčného života.[30] Zatímco moderní buňky se mohou spolehnout na komplikované proteinové stroje, které katalyzují tyto rozhodující procesy, protokoly to musely dosáhnout pomocí jednodušších mechanismů.
Protocelly složené z mastné kyseliny[31] by byli schopni snadno vyměňovat malé molekuly a ionty s jejich prostředím.[1] Membrány sestávající z mastných kyselin mají relativně vysokou permeabilitu pro molekuly, jako jsou nukleosidmonofosfát (NMP), nukleosid difosfát (NDP) a nukleosid trifosfát (NTP), a může vydržet milimolární koncentrace Mg2+.[32] Osmotický tlak může také hrát významnou roli, pokud jde o tento pasivní transport membrány.[1]
Účinky na životní prostředí byly navrženy ke spuštění podmínek, za kterých je transport větších molekul, jako je DNA a RNA, přes membrány protokolů je možné. Například bylo navrženo, že elektroporace vyplývající z Blesk takové přepravy by mohly umožnit stávky.[33] Elektroporace je rychlé zvýšení propustnosti dvouvrstvy vyvolané aplikací velkého umělého elektrického pole přes membránu. Během elektroporace se molekuly lipidů v poloze membrány posouvají a otevírají póry (díry), které působí jako vodivá cesta, kterou hydrofobní molekuly jako nukleové kyseliny může projít lipidovou dvojvrstvou.[34] Podobný přenos obsahu mezi protokoly as okolním řešením může být způsoben zmrazením a následným rozmrazením. Může k tomu dojít například v prostředí, ve kterém denní a noční cykly způsobují opakované mrznutí. Laboratorní experimenty ukázaly, že takové podmínky umožňují výměnu genetických informací mezi populacemi protocelul.[35] To lze vysvětlit skutečností, že membrány jsou vysoce propustné při teplotách mírně pod teplotou fázového přechodu. Je-li tohoto bodu dosaženo během cyklu zmrazení-rozmrazení, mohou dokonce i velké a vysoce nabité molekuly dočasně projít membránou protocell.
Některé molekuly nebo částice jsou příliš velké nebo příliš hydrofilní, aby prošly lipidovou dvojvrstvou i za těchto podmínek, ale lze je přes membránu pohybovat fúze nebo nadějně vezikuly,[36] události, které byly také pozorovány u cyklů zmrazení a rozmrazení.[37] To nakonec mohlo vést k mechanismům, které usnadňují pohyb molekul dovnitř protocely (endocytóza ) nebo uvolnit jeho obsah do extracelulárního prostoru (exocytóza ).[36]
Umělé modely
Langmuir-Blodgettova depozice
Počínaje technikou běžně používanou k ukládání molekul na pevný povrch, Langmuir-Blodgettovou depozicí, jsou vědci schopni sestavit fosfolipidové membrány libovolné složitosti vrstvu po vrstvě.[38][39] Tyto umělé fosfolipidové membrány podporují funkční inzerci jak purifikovaných, tak i in situ vyjádřený membránové proteiny.[39] Technika by mohla pomoci astrobiologové pochopit, jak vznikly první živé buňky.[38]
Jecewanské protokoly

Jeewanu protocell jsou syntetické chemické částice, které mají buňka -jako struktura a zdá se, že mají nějaké funkční životní vlastnosti.[40] Nejprve syntetizován v roce 1963 z jednoduchých minerálů a základních organických látek, zatímco byl vystaven sluneční světlo, stále se uvádí, že má určité metabolické schopnosti, přítomnost Semipermeabilní membrána, aminokyseliny, fosfolipidy, sacharidy a molekuly podobné RNA.[40][41] Je však třeba vyjasnit povahu a vlastnosti Jeewanu.[40][41][42]
V podobném experimentu syntézy zmrazená směs vody, methanolu, čpavek a kysličník uhelnatý byl vystaven ultrafialovému (UV) záření. Tato kombinace poskytla velké množství organického materiálu, který se po ponoření do vody samoorganizoval a vytvořil globule nebo vezikuly.[43] Vyšetřující vědec považoval tyto kuličky za podobné buněčným membránám, které obklopují a koncentrují chemii života a oddělují jejich vnitřek od vnějšího světa. Globuly byly mezi 10 až 40 mikrometry (0,00039 až 0,00157 palce) nebo přibližně o velikosti červených krvinek. Pozoruhodné je, že kuličky fluorescenční, nebo svítil, když je vystaven UV záření. Absorpce UV a jeho přeměna na viditelné světlo tímto způsobem byla považována za jeden z možných způsobů poskytnutí energie primitivní buňce. Pokud by takové kuličky hrály roli v počátcích života, mohla by být fluorescence předchůdcem primitiv fotosyntéza. Taková fluorescence také poskytuje tu výhodu, že působí jako opalovací krém a rozptyluje jakékoli poškození, které by jinak bylo způsobeno UV zářením. Taková ochranná funkce by byla životně důležitá pro život na počátku Země, protože ozónová vrstva, který blokuje nejničivější sluneční paprsky slunce, se vytvořil až po fotosyntetickém životě začal produkovat kyslík.[44]
Biologické struktury
Byla popsána syntéza tří druhů „jeewanu“; dva z nich byly organické a druhý byl anorganický. Byly také vyrobeny další podobné anorganické struktury. Vyšetřující vědec (V. O. Kalinenko) je označil jako „biologické struktury“ a „umělé buňky“. Tvoří se v destilované vodě (stejně jako na agarovém gelu) pod vlivem elektrického pole a chybí jim bílkoviny, aminokyseliny, purinové nebo pyrimidinové báze a určité enzymatické aktivity. Podle vědců NASA „v současnosti známé vědecké principy biologie a biochemie nemohou odpovídat za živé anorganické jednotky“ a „postulovaná existence těchto živých jednotek nebyla prokázána“.[42]
Etika a kontroverze
Výzkum protocellu vyvolal polemiku a protichůdné názory, včetně kritiků vágní definice „umělého života“.[45] Vytvoření základní jednotky života je nejnaléhavějším etickým zájmem, ačkoli nejrozšířenější starostí o protokoly je jejich potenciální hrozba pro lidské zdraví a životní prostředí prostřednictvím nekontrolované replikace.[46]
Viz také
- Abiogeneze - Přirozený proces, při kterém život vzniká z neživé hmoty
- Umělá buňka
- Vznik - Fenomén ve složitých systémech, kde interakce vyvolávají účinky, které nelze přímo předvídat ze subsystémů
- Entropie a život
- Poslední univerzální předek
- Protocellový cirkus, film
- Pseudo-panspermie
- Světová hypotéza RNA
- Syntetická biologie - Interdisciplinární obor biologie a inženýrství
Reference
- ^ A b C d Chen, Irene A .; Walde, Peter (červenec 2010). „Od samo-sestavených vezikul po protokoly“. Cold Spring Harb Perspect Biol. 2 (7): a002170. doi:10.1101 / cshperspect.a002170. PMC 2890201. PMID 20519344.
- ^ A b Garwood, Russell J. (2012). „Vzory v paleontologii: první 3 miliardy let evoluce“. Paleontologie online. 2 (11): 1–14. Citováno 25. června 2015.
- ^ National Science Foundation (2013). „Zkoumání původu života - protokoly“. Citováno 2014-03-18.
- ^ A b C d Chen, Irene A. (8. prosince 2006). „Vznik buněk během vzniku života“. Věda. 314 (5805): 1558–59. doi:10.1126 / science.1137541. PMID 17158315.
- ^ A b C d Zimmer, Carl (26. června 2004). „Co přišlo před DNA?“. Objevte časopis: 1–5.
- ^ Rasmussen, Steen (2. července 2014). „Vědci vytvářejí možného předchůdce života“. Letters Journal Exploring the Frontiers of Physics. 107 (2). Astrobiologický web. Citováno 2014-10-24.
- ^ Alberts, Bruce; Johnson, Alexander; Lewis, Juliane; Morgan, David; Raff, Martin; Roberts, Keith; Walter, Peter (2014). Molekulární biologie buňky (6. vyd.). New York: Garland Science. ISBN 978-1317563754. Citováno 2018-06-15.
- ^ Deamer, D.W .; Dworkin, J.P. (2005). "Chemie a fyzika primitivních membrán". Horní. Curr. Chem. Témata ze současné chemie. 259: 1–27. doi:10.1007 / b136806. ISBN 3-540-27759-5.
- ^ Walde, P (2006). "Složení povrchově aktivních látek a jejich různé možné role pro původ (y) života". Orig. Life Evol. Biosf. 36 (2): 109–50. Bibcode:2006OLEB ... 36..109 W.. doi:10.1007 / s11084-005-9004-3. hdl:20.500.11850/24036. PMID 16642266. S2CID 8928298.
- ^ Sakuma, Yuka; Imai, Masayuki (2015). „Od vezikul k protokolům: role amfifilních molekul“. Život. 5 (1): 651–75. doi:10,3390 / život5010651. PMC 4390873. PMID 25738256.
- ^ Shapiro, Robert (12. února 2007). „Jednodušší původ pro život“. Scientific American. 296 (6): 46–53. Bibcode:2007SciAm.296f..46S. doi:10.1038 / scientificamerican0607-46. PMID 17663224.
- ^ Vodopich, Darrell S .; Moore., Randy (2002). „Důležitost membrán“. Biologická laboratorní příručka, 6 / a. McGraw-Hill. Citováno 2014-03-17.
- ^ Chang, Thomas Ming Swi (2007). Umělé buňky: biotechnologie, nanomedicína, regenerativní medicína, krevní náhrady, bioenkapsulace, terapie buňkami / kmenovými buňkami. Hackensack, NJ: World Scientific. ISBN 978-981-270-576-1.
- ^ Knowles, JR (1980). "Enzymem katalyzované reakce přenosu fosforylu". Annu. Biochem. 49: 877–919. doi:10.1146 / annurev.bi.49.070180.004305. PMID 6250450.
- ^ Campbell, Neil A .; Williamson, Brad; Heyden, Robin J. (2006). Biology: Exploring Life. Boston, MA: Pearson Prentice Hall. ISBN 978-0-13-250882-7.
- ^ A b Clavin, Whitney (13. března 2014). „Jak vznikl život? Na palivové články mohou mít odpovědi“. NASA.
- ^ Walsby, AE (1994). "Plynové vezikuly". Mikrobiologické recenze. 58 (1): 94–144. doi:10.1128 / MMBR.58.1.94-144.1994. PMC 372955. PMID 8177173.
- ^ Szostak, Jack W. (3. září 2004). „Bitva o bubliny mohla vyvolat evoluci“. Howard Hughes Medical Institute.
- ^ A b National Science Foundation (2013). „Membránové lipidy minulosti a současnosti“. Exploring Life's Origins Project - a timeline of Life's Evolution. Citováno 2014-03-17.
- ^ Douliez, Jean-Paul; Zhendre, Vanessa; Grélard, Axelle; Dufourc, Erick J. (24. listopadu 2014). "Aminosilan / vezikuly kyseliny olejové jako modelové membrány protocelul". Langmuir. 30 (49): 14717–24. doi:10.1021 / la503908z. PMID 25420203.
- ^ "Peptidové lepidlo mohlo držet první komponenty protocell pohromadě".
- ^ Kamat, Neha P .; Tobé, Sylvia; Hill, Ian T .; Szostak, Jack W. (2015). „Elektrostatická lokalizace RNA na protocellové membrány kationtovými hydrofobními peptidy“. Angewandte Chemie International Edition. 54 (40): 11735–39. doi:10,1002 / anie.201505742. PMC 4600236. PMID 26223820.
- ^ A b Switek, Brian (13. února 2012). „Debata o původu života“. Příroda -! Novinky.
- ^ Szostak, Jack W. (4. června 2008). „Vědci vytvářejí modelový protokol schopný kopírovat DNA“. Zprávy HHMI. Howard Hughes Medical Institute.
- ^ Cohen, Philip (23. října 2003). "Hliněné dohazování mohlo vyvolat život". Nový vědec.
Odkaz na deník: Věda (svazek 302, s. 618)
- ^ A b Kámen, Howard A. (7. února 2011). "Hliněné obrněné bubliny mohly tvořit první protokoly". Harvardská škola inženýrství a aplikovaných věd.
- ^ Jia, Tony Z .; Chandru, Kuhan; Hongo, Yayoi; Afrin, Rehana; Usui, Tomohiro; Myojo, Kunihiro; Cleaves, H. James (22. července 2019). „Membránové polyesterové mikrokapky jako prvotní oddíly na počátku života“. Sborník Národní akademie věd. 116 (32): 15830–35. doi:10.1073 / pnas.1902336116. PMC 6690027. PMID 31332006.
- ^ Tokijský technologický institut (23. července 2019). „Vědci ELSI objevují novou chemii, která může pomoci vysvětlit počátky buněčného života - Chemici zjišťují, že nejjednodušší organické molekuly se mohou samy sestavit, aby poskytly struktury podobné buňkám za časných podmínek Země. EurekAlert!. Citováno 23. července 2019.
- ^ Woese, Carl R .; Fox, George E. (březen 1977). "Koncept buněčné evoluce". Journal of Molecular Evolution. 10 (1): 1–6. Bibcode:1977JMolE..10 .... 1W. doi:10.1007 / BF01796132. PMID 903983. S2CID 24613906.
- ^ Gyles, C .; Boerlin, P. (06.12.2013). "Horizontálně přenesené genetické prvky a jejich role v patogenezi bakteriální nemoci". Veterinární patologie. 51 (2): 328–40. doi:10.1177/0300985813511131. ISSN 0300-9858. PMID 24318976. S2CID 206510894.
- ^ Müller, A. W. (červen 2006). „Znovu vytvořit svět RNA“. Cell Mol Life Sci. 63 (11): 1278–93. doi:10.1007 / s00018-006-6047-1. PMID 16649141. S2CID 36021694.
- ^ Ma, Wentao; Yu, Chunwu; Zhang, Wentao; Hu., Jiming (listopad 2007). „Ribozymy nukleotidové syntetázy se mohly ve světě RNA objevit jako první“. RNA. 13 (11): 2012–19. doi:10,1261 / rna.658507. PMC 2040096. PMID 17878321.
- ^ Demanèche, S; Bertolla, F; Buret, F; et al. (Srpen 2001). „Laboratorní důkazy pro přenos genů v půdě bleskem“. Appl. Environ. Microbiol. 67 (8): 3440–44. doi:10.1128 / AEM.67.8.3440-3444.2001. PMC 93040. PMID 11472916.
- ^ Neumann, E; Schaefer-Ridder, M; Wang, Y; Hofschneider, PH (1982). „Přenos genů do buněk myšího lyomu elektroporací ve vysokých elektrických polích“. EMBO J.. 1 (7): 841–45. doi:10.1002 / j.1460-2075.1982.tb01257.x. PMC 553119. PMID 6329708.
- ^ Litschel, Thomas; Ganzinger, Kristina A .; Movinkel, Torgeir; Heymann, Michael; Robinson, Tom; Hannes Mutschler; Schwille, Petra (2018). „Cykly zmrazení a rozmrazení indukují výměnu obsahu mezi lipidovými váčky velikosti buněk“. New Journal of Physics. 20 (5): 055008. Bibcode:2018NJPh ... 20e5008L. doi:10.1088 / 1367-2630 / aabb96. ISSN 1367-2630.
- ^ A b Norris, V .; Raine, D.J. (Říjen 1998). „Původ štěpení-fúze po celý život“. Orig Life Evol Biosph. 28 (4): 523–37. doi:10.1023 / A: 1006568226145. PMID 9742727. S2CID 24682163.
- ^ Tsuji, Gakushi; Fujii, Satoshi; Sunami, Takeshi; Yomo, Tetsuya (2016-01-19). „Udržitelná proliferace liposomů kompatibilní s vnitřní replikací RNA“. Sborník Národní akademie věd. 113 (3): 590–95. Bibcode:2016PNAS..113..590T. doi:10.1073 / pnas.1516893113. ISSN 0027-8424. PMC 4725462. PMID 26711996.
- ^ A b „Vědci vytvářejí umělé buněčné membrány“. Astrobiologický časopis. 4. října 2014. Citováno 2014-05-07.
- ^ A b Matosevic, Sandro; Paegel, Brian M. (29. září 2013). "Sestava buněčné membrány vrstva po vrstvě". Přírodní chemie. 5 (11): 958–63. Bibcode:2013NatCh ... 5..958M. doi:10.1038 / nchem.1765. PMC 4003896. PMID 24153375.
- ^ A b C Grote, M (září 2011). „Jeewanu neboli„ částice života “'" (PDF). Journal of Biosciences. 36 (4): 563–70. doi:10.1007 / s12038-011-9087-0. PMID 21857103. S2CID 19551399. Archivovány od originál (PDF) dne 2014-03-23.
- ^ A b Gupta, V. K .; Rai, R. K. (2013). „Histochemická lokalizace materiálu podobného RNA ve fotochemicky vytvořených soběstačných, abiogenních supramolekulárních sestavách Jeewanu'". Int. Res. J. Of Science & Engineering. 1 (1): 1–4. ISSN 2322-0015.
- ^ A b Caren, Linda D .; Ponnamperuma, Cyril (1967). „Přehled některých experimentů se syntézou„ Jeewanu “'" (PDF). Technické memorandum NASA X-1439.
- ^ Dworkin, Jason P .; Deamer, David W .; Sandford, Scott A .; Allamandola, Louis J. (30. ledna 2001). „Samoskladající se amfifilní molekuly: Syntéza v simulovaných mezihvězdných / předkometárních ledech“. Sborník Národní akademie věd Spojených států amerických. 98 (3): 815–19. Bibcode:2001PNAS ... 98..815D. doi:10.1073 / pnas.98.3.815. PMC 14665. PMID 11158552.
- ^ Mullen, L (5. září 2005). „Budování života z hvězdných věcí“. Astrobiologický časopis.
- ^ Bedau, M .; Church, G .; Rasmussen, S .; Caplan, A .; Benner, S .; Fussenegger, M .; Collins, J .; Deamer, D. (27. května 2010). "Život po syntetické buňce". Příroda. 465 (7297): 422–24. Bibcode:2010Natur.465..422.. doi:10.1038 / 465422a. PMID 20495545. S2CID 27471255.
- ^ Bedau, Mark A .; Parke, Emily C. (2009). Etika protokolů, morální a sociální důsledky vytváření života v laboratoři (Online ed.). Cambridge, MA: MIT Press. ISBN 978-0-262-51269-5.
externí odkazy
- „Protokoly: Přemostění neživých a živých věcí.“ Editace: Steen Rasmussen, Mark A. Bedau, Liaochai Chen, David Deamer, David Krakauer, Norman, H. Packard a Peter F. Stadler. MIT Press, Cambridge, Massachusetts. 2008.
- "Living Chemistry & A Natural History of Protocells. „Synth-ethic: Exhibition Art and Synthetic Biology (2013) v Natural History Museum, Vídeň, Rakousko.
- Kenyon, DH; Nissenbaum, A (duben 1976). "Melanoidin a aldokyanoinové mikrosféry: důsledky pro chemickou evoluci a ranou prekambrickou mikropaleontologii". J. Mol. Evol. 7 (3): 245–51. Bibcode:1976JMolE ... 7..245K. doi:10.1007 / bf01731491. PMID 778393. S2CID 2995886.
| Koncepty | ||
|---|---|---|
| Hypotézy | ||
| Výzkum | ||
