Peripatrická speciace - Peripatric speciation
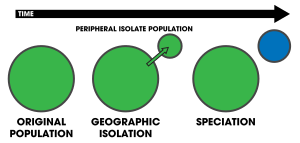


Peripatrická speciace je režim speciace ve kterém nový druh je tvořen z izolované periferní populace.[1]:105 Protože peripatrická speciace se podobá alopatrická speciace, v tom jsou populace izolované a je jim zabráněno výměna genů, může být často obtížné je rozlišit.[2] Nicméně primární charakteristika peripatrické speciace naznačuje, že jedna z populací je mnohem menší než druhá. Podmínky peripatrický a peripatry jsou často používány v biogeografie, s odkazem na organismy jejichž rozsahy těsně sousedí, ale nepřekrývají se, přičemž se oddělují tam, kde se tyto organismy nevyskytují - například na oceánský ostrov ve srovnání s pevninou. Tyto organismy jsou obvykle úzce spjaty (např. sesterské druhy ); jejich distribuce je výsledkem peripatrické speciace.
Koncept peripatrické speciace byl poprvé načrtnut evolučním biologem Ernst Mayr v roce 1954.[3] Od té doby byly vyvinuty další alternativní modely, jako např odstředivá speciace, což předpokládá, že populace druhu zažívá geografická období rozšíření rozsahu následovala zmenšující se období, zanechávající za sebou malé izolované populace na periferii hlavní populace. Další modely zahrnovaly účinky sexuální výběr na omezené velikosti populace. Byly vyvinuty další související modely periferně izolovaných populací založené na chromozomálních přeskupeních, jako např začínající speciace a kvantová speciace.
Existenci peripatrické speciace podporují pozorovací důkazy a laboratorní experimenty.[1]:106 Vědci sledující vzorce biogeografické distribuce druhů a její fylogenetické vztahy jsou schopny rekonstruovat historický proces, kterým jsou rozcházely se. Dále jsou oceánské ostrovy často předmětem peripatrického výzkumu speciace kvůli jejich izolovaným stanovištím - s Havajské ostrovy široce zastoupen ve velké části vědecké literatury.
Dějiny
Peripatrickou speciaci původně navrhl Ernst Mayr v roce 1954,[3] a plně teoreticky modelován v roce 1982.[4] Souvisí to s zakladatelský efekt, kde mohou malé žijící populace procházet úzkými místy pro výběr.[5] Efekt zakladatele je založen na modelech, které naznačují, že peripatrická speciace může nastat interakcí výběru a genetický drift,[1]:106 což může hrát významnou roli.[6] Mayr tuto myšlenku poprvé pochopil svými pozorováními ledňáček populace na Nové Guineji a okolních ostrovech.[1]:389 Tanysiptera galatea byl do značné míry jednotný v morfologii na pevnině, ale populace na okolních ostrovech se významně lišily - odkazovaly na tento vzor jako na „peripatric“.[1]:389 Stejný vzorec byl v té době pozorován mnoha Mayrovými současníky, jako například E. B. Forda studie Maniola jurtina.[7]:522 Přibližně ve stejnou dobu botanik Verne Grant vyvinul model kvantové speciace velmi podobný Mayrovu modelu v kontextu rostlin.[8]
V takzvaných Mayrových genetických revolucích předpokládal, že primární roli, která vyústila v tento vzorec, hraje genetický drift.[1]:389 Vidět, že druhová soudržnost je udržována konzervativními silami, jako je epistáza a pomalé tempo šíření příznivých alel ve velké populaci (silně založené na J. B. S. Haldane Výpočty), usoudil, že speciace může probíhat pouze tehdy, když a zúžení populace došlo.[1]:389 Například malá izolovaná zakladatelská populace by se mohla usadit na ostrově. Obsahující menší genetické variace od hlavní populace, může dojít k posunům ve frekvencích alel z různých selekčních tlaků.[1]:390 To k dalším změnám v síti propojených lokusů, k nastartování kaskády genetických změn nebo k „genetické revoluci“ - rozsáhlé reorganizaci celého genomu periferní populace.[1]:391 Mayr si uvědomil, že šance na úspěch jsou neuvěřitelně nízké a že vyhynutí je pravděpodobné; ačkoli poznamenal, že v té době existovaly některé příklady úspěšných populací zakladatelů.[7]:522
Krátce poté, co Mayr, William Louis Brown, Jr. navrhl v roce 1957 alternativní model peripatrické speciace zvaný odstředivá speciace. V letech 1976 a 1980 vyvinul Kaneshiroův model peripatrické speciace Kenneth Y. Kaneshiro která se zaměřila na sexuální výběr jako hnací sílu speciace během úzkých míst populace.[9][10][11]
Modely
Peripatric
Peripatrické speciační modely jsou totožné s modely vikářství (alopatrická speciace).[1]:105 Speciace vyžaduje geografické oddělení i čas a může být výsledkem předvídatelného vedlejšího produktu.[12] Peripatrii lze odlišit od alopatrické speciace třemi klíčovými rysy:[1]:105
- Velikost izolované populace
- Silný výběr způsobené rozptýlením a kolonizací nových prostředí,
- Účinky genetický drift na malé populace.
Velikost populace je důležitá, protože jednotlivci kolonizující nové stanoviště pravděpodobně obsahují jen malý vzorek genetické variace původní populace. To podporuje divergenci v důsledku silných selektivních tlaků, což vede k rychlému fixace z alela v potomstvu populace. To vede k možnosti genetické nekompatibility rozvíjet se. Tyto nekompatibility způsobují reprodukční izolace, což vede k - někdy rychlým - speciačním událostem.[1]:105 Kromě toho se uplatňují dvě důležité předpovědi, a to, že geologické nebo klimatické změny způsobují, že populace se lokálně fragmentují (nebo regionálně při zvažování alopatrické speciace) a že reprodukční vlastnosti izolované populace se vyvíjejí natolik, aby se zabránilo křížení potenciálu sekundární kontakt.[13]
Výsledkem peripatrického modelu jsou páry progenitor-derivátových druhů, které se nazývají, přičemž se derivátové druhy (periferně izolovaná populace) - geograficky a geneticky izolované od progenitorových druhů - rozcházejí.[14] Specifický fylogenetické podpis je výsledkem tohoto způsobu speciace: stane se geograficky rozšířeným předkem paraphyletic (čímž se stává paraspecies ), pokud jde o derivátové druhy (periferní izolát).[1]:470 Pojem paraspecies je tedy logickým důsledkem koncept evolučního druhu, kterým jeden druh vede k dceřinému druhu.[15] Předpokládá se, že charakterové vlastnosti periferně izolovaných druhů se stávají apomorfní, zatímco centrální populace zůstává pleisomorfní.[16]
Moderní kladistické metody vyvinuly definice, které mimochodem odstranily odvozené druhy definováním kladů způsobem, který předpokládá, že když dojde ke speciační události, původní druh již neexistuje, zatímco vznikají dva nové druhy; toto není případ peripatrické speciace.[8] Mayr před tím varoval, protože to způsobí, že druh ztratí status klasifikace.[17] Loren H. Rieseberg a Luc Brouillet uznali stejné dilema v klasifikaci rostlin.[18]
Kvantová a začínající speciace
Botanik Verne Grant navrhl termín kvantová speciace, který spojil myšlenky o J. T. Gulick (jeho pozorování variace druhů v poloizolaci), Sewall Wright (jeho modely genetického driftu), Mayr (oba jeho modely peripatrické a genetické revoluce) a George Gaylord Simpson (jeho vývoj myšlenky kvantová evoluce ).[19]:114 Kvantová speciace je rychlý proces s velkými genotypovými nebo fenotypovými účinky, přičemž nový rostlinný druh s křížovým hnojením se uvolňuje z větší populace jako částečně izolovaná periferní populace.[20][19]:114 Křížení a genetický drift probíhá kvůli snížené velikosti populace, což vede ke změnám v genomu, které by s největší pravděpodobností vyhynuly (kvůli nízké adaptivní hodnotě).[19]:115 Ve vzácných případech mohou nastat chromozomální znaky s adaptivní hodnotou, což vede k původu nového derivátového druhu.[8][21] Důkazy o výskytu tohoto typu speciace byly nalezeny u několika párů rostlinných druhů: Layia discoidea a L. glandulosa, Clarkia lingulata a C. biloba, a Stephanomeria malheurensis a S. exigua ssp. coronaria.[8]
Úzce související model peripatrické speciace se nazývá nadějná speciace - do značné míry se používá v kontextu speciace rostlin.[22] Začínající proces, kde nový druh pochází z okraje předků, je považován za běžný v rostlinách[22]—Zejména v párech druhů progenitor-derivát.[23]
Odstředivá speciace
William Louis Brown, Jr. navrhl v roce 1957 alternativní model peripatrické speciace zvaný odstředivá speciace. Tento model kontrastuje s peripatrickou speciací na základě původu genetické novinky, která vede k reprodukční izolaci.[24] Populace druhu zažívá období rozšiřování geografického rozsahu následované obdobími kontrakce. Během fáze kontrakce se fragmenty populace izolují jako malé uprchlický populace na periferii centrální populace. Vzhledem k velké velikosti a potenciálně větší genetické variabilitě v centrální populaci mutace vstávat snadněji. Tyto mutace jsou ponechány v izolovaných periferních populacích, čímž podporují reprodukční izolaci. V důsledku toho Brown navrhl, že během další fáze expanze centrální populace přemůže periferní populace a znemožní speciaci. Pokud však tento druh najde specializované ekologické místo, mohou dva koexistovat.[25][26] Fylogenetickým podpisem tohoto modelu je, že se stane centrální populací odvozený, zatímco periferní izoláty zůstávají pleisomorfní[16]—Severní strana obecného modelu. Na rozdíl od odstředivé speciace byla peripatrická speciace někdy označována jako dostředivá speciace (kontrast viz obrázky 1 a 2).[27] Odstředivá speciace byla ve vědecké literatuře do značné míry ignorována, často dominovala tradiční model peripatrické speciace.[28][24][16] Navzdory tomu Brown uvedl řadu důkazů na podporu svého modelu, které dosud nebyly vyvráceny.[25]
Peromyscus polionotus a P. melanotis (periferně izolované druhy z centrální populace P. maniculatus) vznikly prostřednictvím odstředivého speciačního modelu.[29] Odstředivá speciace mohla proběhnout v stromové klokani, Jihoamerické žáby (Ceratophrys ), rejsci (Crocidura ) a primáti (Presbytis melalophos ).[28] John C. Briggs sdružuje odstředivou speciaci s centra původu, tvrdí, že odstředivý model je lépe podporován údaji, s odvoláním na druhové vzory z navrhovaného „centra původu“ v rámci Indo-západní Pacifik[28]
Kaneshiro model

Když sexuální druhy zažívá úzké místo populace - to znamená, když je genetická variace snížena kvůli malé velikosti populace -páření diskriminace mezi ženami může být změněn poklesem námluvy chování mužů.[11] Tím může dojít k oslabení tlaků na sexuální výběr u izolované periferní populace a jako vedlejší produkt změněného systému rozpoznávání páření, sekundární sexuální vlastnosti může se objevit.[9] Nakonec se růst velikosti populace spojil s románem preference ženského partnera povede k reprodukční izolaci od hlavní populace - čímž se dokončí proces peripatrické speciace.[10] Podpora tohoto modelu pochází z experimentů a pozorování druhů, které vykazují asymetrické vzory páření, jako je havajský Drosophila druh[30][31] nebo havajský kriket Laupala.[32] Tento model však nebyl zcela podporován experimenty, a proto nemusí představovat věrohodný proces peripatrické speciace, který probíhá v přírodě.[11]
Důkaz
Pozorování a laboratorní experimenty podporují výskyt peripatrické speciace. Ostrovy a souostroví jsou často předmětem speciačních studií v tom smyslu, že představují izolované populace organismů. Ostrovní druhy poskytují přímý důkaz o speciaci vyskytující se peripatricky tak, že „přítomnost endemický druhy na oceánských ostrovech, jejichž nejbližší příbuzní obývají blízké kontinent "musí pocházet z kolonizační události.[1]:106–107 Srovnávací fylogeografie oceánské souostroví ukazuje konzistentní vzorce postupné kolonizace a speciace podél ostrovních řetězců, zejména na Azory ostrovy, Kanárské ostrovy, Společenské ostrovy, Markézské ostrovy, Galapágy, Australské ostrovy a Havajské ostrovy - všechny vyjadřují geologické vzory prostorové izolace a v některých případech lineární uspořádání.[33] Peripatrická speciace se vyskytuje také na kontinentech, protože k izolaci malých populací může docházet prostřednictvím různých geografických a disperze Události. Byly provedeny laboratorní studie, kde populace Drosophila jsou například od sebe odděleny a vyvíjejí se v reprodukční izolaci.
Havajské souostroví



Drosophila druhy na Havajské souostroví pomohli vědcům pochopit speciační procesy velmi podrobně. Je dobře známo, že Drosophila prošel adaptivní záření na stovky endemické druhy na havajském ostrovním řetězci;[1]:107[34] pocházející od jediného společného předka (podporováno molekulární analýzou).[35] Studie důsledně zjišťují, že ke kolonizaci každého ostrova došlo od starších k mladším ostrovům a v Drosophila, peripatricky specifikující nejméně padesát procent času.[1]:108 Ve spojení s Drosophila, Havajské lobeliady (Cyanea ) také prošly adaptivním zářením, s více než dvaceti sedmi procenty existující druhy vznikající po nové kolonizaci ostrovů - příkladem peripatrické speciace - opět se vyskytující ve starém až mladém ostrovním směru.[36][37][38]
Jiné endemické druhy na Havaji také poskytují důkazy o peripatrické speciaci, jako jsou endemické nelétavé cvrčci (Laupala ). Odhaduje se, že „17 druhů z 36 dobře prostudovaných případů [Laupala] speciace byly peripatrické ".[1]:108[39] Druhy rostlin v rodech jako např Dubautia, Wilkesia, a Argyroxiphium také vyzařovaly podél souostroví.[40] Jiná zvířata kromě hmyzu vykazují stejný vzorec, například hlemýžď havajský jantarový (Succinea caduca ),[41] a „Elepaio lapače.[42]
Tetragnatha pavouci se také peripatricky zaměřili na havajské ostrovy,[43][44] Bylo zdokumentováno mnoho členovců, kteří existovali ve vzorcích odpovídajících geologickému vývoji ostrovního řetězce, takže fylogenetické rekonstrukce nacházejí mladší druhy obývající geologicky mladší ostrovy a starší druhy obývající starší ostrovy[45] (nebo v některých případech předkové pocházejí z doby, kdy byly vystaveny ostrovy v současné době pod hladinou moře). Pavouci, jako jsou ti z rodu Orsonwelles vzory exponátů kompatibilní s geologií starých i mladých.[46] Jiné endemické rody jako např Argyrodes bylo prokázáno, že se specializovali podél ostrovního řetězce.[47] Pagiopalus, Pedinopistha a část rodiny Thomisidae adaptivně vyzařovaly podél ostrovního řetězce,[48] stejně jako rodina vlčích pavouků, Lycosidae.[49]
Řada dalších havajských endemických druhů a rodů členovců si nechala prostudovat své speciační a fylogeografické vzorce: Drosophila grimshawi druhový komplex,[50] motýlice (Megalagrion xanthomelas a Megalagrion pacificum),[51] Doryonychus raptor, Littorophiloscia hawaiiensis, Anax strenuus, Nesogonia blackburni, Theridion Grallator,[52] Vanessa tameamea, Hyalopeplus pellucidus, Coleotichus blackburniae, Labula, Hawaiioscia, Banza (v rodině Tettigoniidae ), Caconemobius, Eupethicea, Ptycta, Megalagrion, Prognathogryllus, Nesosydne, Cephalops, Trupanea a kmen Platynini —Všechny naznačují opakované záření mezi ostrovy.[53]
Ostatní ostrovy
Fylogenetické studie druhu krabů (Misumenops rapaensis ) v rodu Thomisidae nachází se na Australské ostrovy založili „postupnou kolonizaci linie po australském souostroví směrem k mladším ostrovům“. M. rapaensis tradičně byl považován za jediný druh; vzhledem k tomu, že tato konkrétní studie zjistila výrazné genetické rozdíly odpovídající postupnému věku ostrovů.[54] The fík druhy rostlin Scrophularia lowei Předpokládá se, že vznikly peripatrickou speciační událostí s rozšířenějšími druhy na pevnině, Scrophularia arguta rozptýlení do Macaronesian ostrovy.[55][56] Další členové stejného rodu také vznikli při jednotlivých kolonizačních událostech mezi ostrovy.[57][58]
Druhy druhů na kontinentech


Výskyt peripatrie na kontinentech je obtížnější zjistit kvůli možnosti pravděpodobného vysvětlení vikariata.[1]:110 Studie týkající se kalifornských druhů rostlin Clarkia biloba a C. lingulata silně naznačují peripatrický původ.[59] Kromě toho bylo provedeno velké množství výzkumu na několika druzích suchozemských hlemýžďů chirality což naznačuje peripatrii (přičemž někteří autoři berou na vědomí další možné interpretace).[1]:111
The kaštan-sledoval antbird (Sciaphylax hemimelaena) se nachází v Národní park Noel Kempff Mercado (Serrania de Huanchaca) v Bolívii. V této oblasti existuje fragment lesa, který byl podle odhadů izolován po dobu 1000–3000 let. Populace S. hemimelaena protivníci, kteří se nacházejí v izolované záplatě, vyjadřují významnou divergenci písní; považována za „raný krok“ v procesu peripatrické speciace. Dále, periferní izolace "může částečně vysvětlit dramatickou diverzifikaci subosciny v Amazonia ".[13]
Montane ostnatý throated žába rákosu druhový komplex (rod: Hyperolius ) vznikly výskytem peripatrických speciačních událostí. Lucinda P. Lawson tvrdí, že zeměpisné rozpětí tohoto druhu na východě Afromontan Biodiversity Hotspot podporuje peripatrický model, který podporuje speciaci; což naznačuje, že tento způsob speciace může hrát významnou roli ve „vysoce fragmentovaných ekosystémech“.[2]
Ve studii o fylogenezi a biogeografii rodu suchozemských hlemýžďů Monacha druh M. ciscaucasica Předpokládá se, že se peripatricky specializoval na populaci M. roseni. Navíc, M. claussi Skládá se z malé populace na okraji mnohem většího rozsahu M. subcarthusiana což naznačuje, že také vznikla peripatrickou speciací.[60]


Červený smrk (Picea rubens ) vznikl z izolované populace smrku černého (Picea mariana ). Během Pleistocén Populace černého smrku se geograficky izolovala, pravděpodobně kvůli zalednění. Zeměpisný rozsah černého smrku je mnohem větší než červeného smrku. Červený smrk má výrazně nižší genetickou rozmanitost jak v DNA, tak i v ní mitochondriální DNA než černý smrk.[61][62] Kromě toho genetická variace červeného smrku nemá žádný jedinečný mitochondriální haplotypy, pouze podmnožiny těch v černém smrku; což naznačuje, že červený smrk se peripatricky specializoval na populaci černého smrku.[63][64][65] Předpokládá se, že celý rod Picea v severní Americe se zpestřil procesem peripatrické speciace, protože četné páry blízce příbuzných druhů rodu mají menší rozsahy jižní populace; a osoby s překrývajícími se rozsahy často vykazují slabou reprodukční izolaci.[66][62]
Pomocí fylogeografického přístupu spárovaného s ekologické nika modely (tj. predikce a identifikace druhů rozšiřujících se a stahujících se na základě aktuálních sahá do vhodných stanovišť ekologické výklenky, korelovaný s fosilními a molekulárními daty), vědci zjistili, že prerijní pes druh Cynomys mexicanus specializován peripatricky od Cynomys ludovicianus přibližně před 230 000 lety. Severoamerické ledovcové cykly podporovaly expanzi a kontrakci prérijních psů, což vedlo k izolaci reliktní populace v refugium nachází se v současnosti Coahuila, Mexiko.[67] Tato distribuce a paleobiogeografický vzor koreluje s jinými druhy vyjadřujícími podobné vzory biografického rozsahu[67] například s Sorex cinereus komplex.[68]
Laboratorní experimenty
| Druh | Replikuje | Rok |
|---|---|---|
| Drosophila adiastola | 1 | 1979[69] |
| Drosophila silvestris | 1 | 1980[70] |
| Drosophila pseudoobscura | 8 | 1985[71] |
| Drosophila simulans | 8 | 1985[72] |
| Musca domestica | 6 | 1991[73] |
| Drosophila pseudoobscura | 42 | 1993[74] |
| Drosophila melanogaster | 50 | 1998[75] |
| Drosophila melanogaster | 19; 19 | 1999[76] |
| Drosophila grimshawi | 1 | N / A[11] |
Peripatrická speciace byla zkoumána jak v laboratorních studiích, tak v přírodě. Jerry Coyne a H. Allen Orr v Speciace naznačují, že většina laboratorních studií alopatrické speciace je také příkladem peripatrické speciace kvůli její malé velikosti populace a nevyhnutelné divergentní selekci, kterou procházejí.[1]:106 Velká část laboratorního výzkumu týkajícího se peripatrie je neoddělitelně spjata zakladatelský efekt výzkum. Coyne a Orr docházejí k závěru, že role výběru ve speciaci je dobře zavedená genetický drift Role není podporována experimentálními a terénními daty - což naznačuje, že nedochází ke speciaci na efekt zakladatele.[1]:410 V této věci však bylo provedeno velké množství výzkumu a jedna studie byla provedena úzké místo populace Drosophila pseudoobscura našel důkaz izolace po jediném úzkém místě.[77][78]
Tabulka je neúplnou tabulkou laboratorních experimentů zaměřených výslovně na peripatrickou speciaci. Většina studií také prováděla experimenty také se speciací vikariátu. Sloupec „replikáty“ označuje počet řádků použitých v experimentu - to znamená, kolik nezávislých populací bylo použito (nikoli velikost populace nebo počet provedených generací).[11]
Reference
![]() Tento článek byl odeslán na adresu WikiJournal of Science pro externí akademické vzájemné hodnocení v roce 2018 (zprávy recenzenta ). Aktualizovaný obsah byl znovu integrován na stránku Wikipedie pod a CC-BY-SA-3.0 licence (2018 ). Verze záznamu v revidovaném znění je: „Peripatrická speciace“ (PDF), WikiJournal of Science, 1 (2): 008, 14. srpna 2018, doi:10.15347 / WJS / 2018.008, ISSN 2470-6345, Wikidata Q56553060
Tento článek byl odeslán na adresu WikiJournal of Science pro externí akademické vzájemné hodnocení v roce 2018 (zprávy recenzenta ). Aktualizovaný obsah byl znovu integrován na stránku Wikipedie pod a CC-BY-SA-3.0 licence (2018 ). Verze záznamu v revidovaném znění je: „Peripatrická speciace“ (PDF), WikiJournal of Science, 1 (2): 008, 14. srpna 2018, doi:10.15347 / WJS / 2018.008, ISSN 2470-6345, Wikidata Q56553060
- ^ A b C d E F G h i j k l m n Ó str q r s t u Jerry A. Coyne; H. Allen Orr (2004), Speciace, Sinauer Associates, s. 1–545, ISBN 978-0-87893-091-3
- ^ A b Lucinda P. Lawson, John M. Bates, Michele Menegon a Simon P. Loader (2015), „Divergence na okrajích: peripatrická izolace v montánním ostnatém throated reed žabím komplexu“, BMC Evoluční biologie, 15 (128): 128, doi:10.1186 / s12862-015-0384-3, PMC 4487588, PMID 26126573CS1 maint: více jmen: seznam autorů (odkaz)
- ^ A b Ernst Mayr. (1954). Změna genetického prostředí a evoluce. V J. Huxley, A. C. Hardy a E. B. Ford. (eds) Evoluce jako proces, Unwin Brothers, Londýn. Str. 157–180.
- ^ Ernst Mayr. (1982). Procesy speciace u zvířat. In A. R. I. Liss. (eds) Mechanismy speciace, Alan R. Liss Inc., New York. Str. 1–19.
- ^ W. B. Provine (1. července 2004). „Ernst Mayr: Genetika a speciace“. Genetika. 167 (3): 1041–6. PMC 1470966. PMID 15280221.
- ^ Alan R. Templeton (1. dubna 1980). „Teorie speciace prostřednictvím principu zakladatele“. Genetika. 94 (4): 1011–38. PMC 1214177. PMID 6777243.
- ^ A b Ernst Mayr (1963), Druhy zvířat a evoluce, Harvard University Press, s. 1–797
- ^ A b C d L. D. Gottlieb (2003), „Přehodnocení klasických příkladů nedávné speciace v rostlinách“, Nový fytolog, 161: 71–82, doi:10.1046 / j.1469-8137.2003.00922.x
- ^ A b Kenneth Y. Kaneshiro (1976), „Etologická izolace a fylogeneze v podskupině havajských rostlin Plantibia Drosophila", Vývoj, 30 (4): 740–745, doi:10.1111 / j.1558-5646.1976.tb00954.x, PMID 28563322, S2CID 205773169
- ^ A b Kenneth Y. Kaneshiro (1980), „Sexuální výběr, speciace a směr vývoje“, Vývoj, 34 (3): 437–444, doi:10.1111 / j.1558-5646.1980.tb04833.x, PMID 28568697, S2CID 28701838
- ^ A b C d E F Anders Ödeen a Ann-Britt Florin (2002), „Sexuální výběr a peripatrická speciace: znovu se objevil model Kaneshiro“, Journal of Evolutionary Biology, 15 (2): 301–306, doi:10.1046 / j.1420-9101.2002.00378.x, S2CID 82095639
- ^ Michael Turelli, Nicholas H. Barton a Jerry A. Coyne (2001), „Teorie a speciace“, Trendy v ekologii a evoluci, 16 (7): 330–343, doi:10.1016 / s0169-5347 (01) 02177-2, PMID 11403865CS1 maint: více jmen: seznam autorů (odkaz)
- ^ A b Nathalie Seddon a Joseph A. Tobias (2007), „Divergence písní na okraji Amazonie: empirický test peripatrického modelu speciace“, Biologický žurnál společnosti Linnean, 90: 173–188, doi:10.1111 / j.1095-8312.2007.00753.x
- ^ Daniel J. Crawford (2010), „Progenitor-derivátové páry druhů a speciace rostlin“, Taxon, 59 (5): 1413–1423, doi:10,1002 / daň 595008
- ^ James S. Albert & Roberto E. Reis (2011). Historická biogeografie neotropických sladkovodních ryb. ISBN 978-0-520-26868-5.
- ^ A b C Jennifer K. Frey (1993), „Režimy tvorby a speciace periferních izolátů“, Systematická biologie, 42 (3): 373–381, doi:10.1093 / sysbio / 42.3.373, S2CID 32546573
- ^ Ernst Mayr (1992), „Koncept místní flóry a biologických druhů“, American Journal of Botany, 79 (2): 222–238, doi:10.2307/2445111, JSTOR 2445111
- ^ Loren H. Rieseberg a Luc Brouillet (1994), „Existuje mnoho druhů rostlin paraphyletic?“, Taxon, 43 (1): 21–32, doi:10.2307/1223457, JSTOR 1223457
- ^ A b C Verne Grant (1971), Speciace rostlin, New York: Columbia University Press, str. 432, ISBN 978-0231083263
- ^ Douglas J. Futuyma (1989), „Speciační trendy a role druhů v makroevoluci“, Americký přírodovědec, 134 (2): 318–321, doi:10.1086/284983
- ^ Loren H. Rieseberg (2001), „Chromozomální přesmyky a speciace“, Trendy v ekologii a evoluci, 16 (7): 351–358, doi:10.1016 / S0169-5347 (01) 02187-5, PMID 11403867
- ^ A b Brian L. Anacker a Sharon Y. Strauss (2013), „Geografie a ekologie speciace rostlin: překrytí rozsahu a divergence niky u sesterských druhů“, Sborník královské společnosti B, 281 (1778): 20132980, doi:10.1098 / rspb.2013.2980, PMC 3906944, PMID 24452025
- ^ Daniel J. Crawford (2010), „Progenitor-derivátové páry druhů a speciace rostlin“, Taxon, 59 (5): 1413–1423, doi:10,1002 / daň 595008
- ^ A b Sergey Gavrilets, Hai Li a Michael D. Vose (2000), „Patterns of Parapatric Speciation“, Vývoj, 54 (4): 1126–1134, CiteSeerX 10.1.1.42.6514, doi:10.1554 / 0014-3820 (2000) 054 [1126: pops] 2.0.co; 2, PMID 11005282CS1 maint: více jmen: seznam autorů (odkaz)
- ^ A b Daniel J. Howard (2003). "Speciace: Allopatrická". Encyclopedia of Life Sciences. eLS. doi:10.1038 / npg.els.0001748. ISBN 978-0470016176.
- ^ W. L. Brown Jr. (1957), „Odstředivá speciace“, Čtvrtletní přehled biologie, 32 (3): 247–277, doi:10.1086/401875
- ^ Brian W. Bowen (2010), „Rozhovor s Johnem C. Briggsem, držitelem ceny Alfreda Russella Wallacee z roku 2005“, Hranice biogeografie, 2 (3): 78–80, ISSN 1948-6596
- ^ A b C John C. Briggs (2000), „Odstředivá speciace a střediska původu“, Časopis biogeografie, 27 (5): 1183–1188, doi:10.1046 / j.1365-2699.2000.00459.x
- ^ Ira F. Greenbaum, Robert J. Baker a Paul R. Ramsey (1978), „Chromozomální evoluce a způsob speciace u tří druhů Peromyscus“, Vývoj, 32 (3): 646–654, doi:10.1111 / j.1558-5646.1978.tb04609.x, PMID 28567964, S2CID 27865356
- ^ Kenneth Y. Kaneshiro (1983), „Sexuální výběr a směr evoluce v biosystematice havajských Drosophilidae“, Každoroční přezkum entomologie, 28: 161–178, doi:10.1146 / annurev.en.28.010183.001113
- ^ Luther Val Giddings a Alan R. Templeton (1983), „Behaviorální fylogeneze a směr vývoje“, Věda, 220 (4595): 372–378, Bibcode:1983Sci ... 220..372G, doi:10.1126 / science.220.4595.372, PMID 17831399, S2CID 45100702
- ^ Kerry L. Shaw & Ezequiel Lugo (2001), „Páření asymetrie a směr vývoje v havajském kriketovém rodu Laupala", Molekulární ekologie, 10 (3): 751–759, doi:10.1046 / j.1365-294x.2001.01219.x, PMID 11298985, S2CID 38590572
- ^ Kerry L. Shaw & Rosemary G. Gillespie (2016), „Comparative phylogeography of oceanic archipelagos: Hotspots for inferences of evolution process“, PNAS, 113 (29): 7986–7993, doi:10.1073 / pnas.1601078113, PMC 4961166, PMID 27432948
- ^ Hannes Schuler, Glen R. Hood, Scott P. Egan a Jeffrey L. Feder (2016), Meyers, Robert A (ed.), „Režimy a mechanismy speciace“, Recenze v buněčné biologii a molekulární medicíně, 2 (3): 60–93, doi:10.1002/3527600906, ISBN 9783527600908CS1 maint: více jmen: seznam autorů (odkaz)
- ^ DeSalle R. (1995). Molekulární přístupy k biogeografické analýze havajských Drosophilidae. Str. 72-89 v W.L. Wagner a V.A. Funk (eds.) Hawaiian Biogeography: Evolution on a Hot-Spot Archipeligo. Smithsonian Institution Press, Washington DC.
- ^ T. J. Givnish (1998). „Adaptivní evoluce rostlin na ostrovech: klasické vzory, molekulární data, nové poznatky“. Evoluce na ostrovech. 281: 304.
- ^ T. J. Givnish, K. J. Sytsma, W. J. Hahn a J. F. Smith. (1995). Molekulární evoluce, adaptivní záření a geografická speciace v Cyanea (Campanulaceae, Lobeliodeae). Str. 259-301 v W.L. Wagner a V.A. Funk (eds.) Hawaiian Biogeography: Evolution on a Hot-Spot Archipeligo. Smithsonian Institution Press, Washington DC.
- ^ Thomas J. Givnish, Kendra C. Millam, Austin R. Mast, Thomas B. Paterson, Terra J. Theim, Andrew L. Hipp, Jillian M. Henss, James F. Smith, Kenneth R. Wood a Kenneth J. Sytsma (2009), „Původ, adaptivní záření a diverzifikace havajských lobeliad (Asterales: Campanulaceae)“, Proc. R. Soc. B, 276 (1656): 407–416, doi:10.1098 / rspb.2008.1204, PMC 2664350, PMID 18854299CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Kerry L. Shaw (2002), „Konflikt mezi jadernými a mitochondriálními fylogeniemi DNA nedávného záření druhů: Co mtDNA odhaluje a zakrývá způsoby speciace v havajských cvrčcích“, PNAS, 99 (25): 16122–16127, Bibcode:2002PNAS ... 9916122S, doi:10.1073 / pnas.242585899, PMC 138575, PMID 12451181
- ^ Martha S. Witter (1990), „Evolution in the Madiinae: Evidence from Enzyme Electrophoresis“, Annals of the Missouri Botanical Garden, 77 (1): 110–117, doi:10.2307/2399630, JSTOR 2399630
- ^ Brenden S. Holland a Robert H. Cowie (2007), „Geografická mozaika pasivního rozptýlení: struktura populace u endemického havajského jantarového šneka Succinea caduca (Mighels, 1845)“, Molekulární ekologie, 16 (12): 2422–2435, doi:10.1111 / j.1365-294X.2007.03246.x, PMID 17561903, S2CID 32193624
- ^ Eric A. VanderWerf, Lindsay C. Young, Norine W. Yeung a David B. Carlon (2010), „Speciace odrazových můstků v havajských muškách: molekulární divergence podporuje nové ostrovní endemity v elepaiu“, Genetika ochrany, 11 (4): 1283–1298, doi:10.1007 / s10592-009-9958-1, S2CID 35883704CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Rosemary G. Gillespie a H. B. Croom. (1995). Porovnání speciačních mechanismů ve skupinách vytvářejících web a netvořících web v rámci řady pavouků. Ve W.L. Wagner & V.A. Funk (eds.) Hawaiian Biogeography: Evolution on a Hot-Spot Archipeligo, Smithsonian Institution Press, Washington DC. Str. 121-146.
- ^ Rosemary G. Gillespie (2005), „Geografický kontext speciace v havajském záření Tetragnatha pavouci (Aranae, Tetragnathidae ", The Journal of Arachnology, 33 (2): 313–322, doi:10.1636/05-15.1, S2CID 11856750
- ^ Rosemary G. Gillespie (2004), „Shromáždění komunity prostřednictvím adaptivního záření u havajských pavouků“, Věda, 303 (5656): 356–359, Bibcode:2004Sci ... 303..356G, doi:10.1126 / science.1091875, PMID 14726588, S2CID 7748888
- ^ Gustavo Hormiga, Miquel Arnedo a Rosemary G. Gillespie (2003), „Speciace na dopravním pásu: Postupná kolonizace Havajských ostrovů pavouky Orsonwelles (Araneae, Linyphiidae)“, Systematická biologie, 52 (1): 70–88, doi:10.1080/10635150390132786, PMID 12554442CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Rosemary G. Gillespie, Malia A. J. Rivera a Jessica E. Garb. (1998). Slunce, surfování a pavouci: taxonomie a fylogeografie havajských Araneae. Sborník 17. evropského arachnologického kolokvia.
- ^ Jessica E. Garb (1999), „Adaptivní záření havajských Thomisidae: biogreografické a genetické důkazy“, The Journal of Arachnology, 27: 71–78
- ^ W. J. Gertsch (1973), "Cavernikózní fauna havajských lávových trubic. 3. Araneae (pavouci)", Tichomořský hmyz, 15: 163–180
- ^ Fabio Piano, Elysse M. Craddock a Michael P. Kambysellis (1997), „Fylogeneze ostrovních populací havajského komplexu Drosophila grimshawi: důkazy z kombinovaných údajů“, Molekulární fylogenetika a evoluce, 7 (2): 173–184, doi:10,1006 / mpev.1996.0387, PMID 9126558CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Steve Jordan, Chris Simon, David Foote a Ronald A. Englund (2005), „Fylogeografické vzory havajských motýlích megalagrionů (Odonata: Coenagrionidae) korelují s hranicemi pleistocénních ostrovů“, Molekulární ekologie, 14 (11): 3457–3470, doi:10.1111 / j.1365-294X.2005.02669.x, PMID 16156815, S2CID 42614215CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Peter J. P. Croucher, Geoff S. Oxford, Athena Lam, Neesha Mody a Rosemary G. Gillespie (2012), „Historie kolonizace a populační genetika barevně polymorfního havajského pavouka šťastného obličeje Theridion Grallator (Araneae, Theridiidae) ", Vývoj, 66 (9): 2815–2833, doi:10.1111 / j.1558-5646.2012.01653.x, PMID 22946805, S2CID 28684202CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Rodák a K. G. Gillespie (1998), „Speciace a fylogeografie havajských suchozemských členovců“, Molekulární ekologie, 7 (4): 519–531, doi:10.1046 / j.1365-294x.1998.00309.x, PMID 9628003, S2CID 29359389
- ^ Jessica E. Garb & Rosemary G. Gillespie (2006), "Ostrov skákající přes centrální Pacifik: mitochondriální DNA detekuje sekvenční kolonizaci australských ostrovů krabovými pavouky (Araneae: Thomisidae)", Časopis biogeografie, 33 (2): 201–220, doi:10.1111 / j.1365-2699.2005.01398.x, S2CID 43087290
- ^ Francisco J. Valtueña, Tomás Rodríguez-Riaño, Josefa López, Carlos Mayo a Ana Ortega-Olivencia (2017), „Peripatrická speciace v endemické makaronéské rostlině po nedávných odchylkách od rozšířeného příbuzného“, PLOS ONE, 12 (6): e0178459, Bibcode:2017PLoSO..1278459V, doi:10.1371 / journal.pone.0178459, PMC 5456078, PMID 28575081CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Francisco J. Valtueña, Josefa López, Juan Álvarez, Tomás Rodríguez-Riaño a Ana Ortega-Olivencia (2016), "Scrophularia arguta, rozšířená jednoletá rostlina na Kanárských ostrovech: jediná nedávná kolonizační událost nebo složitější fylogeografický vzor? ", Ekologie a evoluce, 6 (13): 4258–4273, doi:10,1002 / ece3,2109, PMC 4930978, PMID 27386073CS1 maint: více jmen: seznam autorů (odkaz)
- ^ María L. Navarro-Péreza, Pablo Vargas, Mario Fernández-Mazuecos, Josefa López, Francisco J. Valtueña a Ana Ortega-Olivencia (2015), „Více oken kolonizace do Makaronésie od nespecializovaného šíření Scrophularia od pozdního miocénu “, Perspektivy v ekologii rostlin, evoluci a systematice, 17 (4): 263–273, doi:10.1016 / j.ppees.2015.05.002CS1 maint: více jmen: seznam autorů (odkaz)
- ^ AgnesScheunert a Günther Heubl (2014), „Diverzifikace Scrophularia (Scrophulariaceae) v západním Středomoří a Makaronésii - fylogenetické vztahy, vývoj síťky a biogeografické vzorce ", Molekulární fylogenetika a evoluce, 70: 296–313, doi:10.1016 / j.ympev.2013.09.023, PMID 24096055
- ^ H. Lewis & M. R. Roberts (1956), „Původ Clarkia lingulata", Vývoj, 10 (2): 126–138, doi:10.2307/2405888, JSTOR 2405888
- ^ Marco T. Neiber a Bernhard Hausdorf (2016), „Molekulární fylogeneze a biogeografie rodu Monacha (Gastropoda, Hygromiidae)“, Zoologica Scripta, 46 (3): 1–14, doi:10.1111 / zsc.12218, S2CID 88655961
- ^ Gary J. Hawley a Donald H. DeHayes (1994), „Genetická rozmanitost a populační struktura smrku červeného (Picea rubens)“, Canadian Journal of Botany, 72 (12): 1778–1786, doi:10.1139 / b94-219
- ^ A b Juan P. Jaramillo-Correa a Jean Bousquet (2003), „Nové důkazy mitochondriální DNA o vztahu mezi původcem a derivátem mezi černým a červeným smrkem (Pinaceae)“, American Journal of Botany, 90 (12): 1801–1806, doi:10,3732 / ajb.90.12.1801, PMID 21653356
- ^ JP Jaramillo-Correa, J. Bousquet, J. Beaulieu, N. Isabel, M. Perron a M. Bouillé (2003), „Cross-species amplification of mitochondrial DNA sequence-tagged-site-markers in conifers: the nature of polymorphism a variace uvnitř a mezi druhy v Picea ", Teoretická a aplikovaná genetika, 106 (8): 1353–1367, doi:10.1007 / s00122-002-1174-z, PMID 12750779, S2CID 21097661CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Isabelle Gamache, Juan P. Jaramillo-Correa, Sergey Payette a Jean Bousquet (2003), „Odlišné vzorce rozmanitosti mitochondrií a jaderné DNA v subarktickém černém smrku: otisk zakladatelského efektu spojeného s postglaciální kolonizací“, Molekulární ekologie, 12 (4): 891–901, doi:10.1046 / j.1365-294x.2003.01800.x, PMID 12753210, S2CID 20234158CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Martin Perron, Daniel J. Perry, Christophe Andalo a Jean Bousquet (2000), „Důkazy ze sekvenčních značených stránek nedávného páru progenitor-derivát v jehličnanech“, PNAS, 97 (21): 11331–11336, Bibcode:2000PNAS ... 9711331P, doi:10.1073 / pnas.200417097, PMC 17200, PMID 11016967CS1 maint: více jmen: seznam autorů (odkaz)
- ^ J. W. Wright (1955), „Druhová křížitelnost smrku ve vztahu k rozšíření a taxonomii“, Forest Science, 1 (4): 319–349
- ^ A b Gabriela Castellanos-Morales, Niza Gámez, Reyna A. Castillo-Gámez a Luis E. Eguiarte (2016), „Peripatrická speciace endemického druhu způsobeného pleistocénními klimatickými změnami: Případ mexického prérijního psa (Cynomys mexicanus)", Molekulární fylogenetika a evoluce, 94 (Pt A): 171–181, doi:10.1016 / j.ympev.2015.08.027, PMID 26343460CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Andrew G. Hope, Kelly A. Speer, John R. Demboski, Sandra L. Talbot a Joseph A. Cook (2012), „Klima pro speciaci: Rychlá prostorová diverzifikace v rámci Sorex cinereus komplex rejsků ", Molekulární fylogenetika a evoluce, 64 (3): 671–684, doi:10.1016 / j.ympev.2012.05.021, PMID 22652055CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Lorna H. Arita a Kenneth Y. Kaneshiro (1979), „Etologická izolace mezi dvěma zásobami Drosophila Adiastola Hardy ", Proc. Havaj. Entomol. Soc., 13: 31–34
- ^ J. N. Ahearn (1980), „Evoluce behaviorální reprodukční izolace v laboratorních zásobách Drosophila silvestris", Experientia, 36 (1): 63–64, doi:10.1007 / BF02003975, S2CID 43809774
- ^ Diane M. B. Dodd a Jeffrey R. Powell (1985), „Speciace zakladatele Flush: Aktualizace experimentálních výsledků s Drosophila", Vývoj, 39 (6): 1388–1392, doi:10.1111 / j.1558-5646.1985.tb05704.x, PMID 28564258, S2CID 34137489
- ^ John Ringo, David Wood, Robert Rockwell a Harold Dowse (1985), „Experiment testující dvě hypotézy speciace“, Americký přírodovědec, 126 (5): 642–661, doi:10.1086/284445CS1 maint: více jmen: seznam autorů (odkaz)
- ^ L. M. Meffert a E. H. Bryant (1991), „sklon k páření a námluvy v sériově zúžených liniích mouchy domácí“, Vývoj, 45 (2): 293–306, doi:10.1111 / j.1558-5646.1991.tb04404.x, PMID 28567864, S2CID 13379387
- ^ A. Galiana, A. Moya a F. J. Ayala (1993), „Speciace zakladatelské flush v Drosophila pseudoobscura: rozsáhlý experiment ", Vývoj, 47 (2): 432–444, doi:10.1111 / j.1558-5646.1993.tb02104.x, PMID 28568735, S2CID 42232235CS1 maint: více jmen: seznam autorů (odkaz)
- ^ H. D. Rundle, A. Ø. Mooers, & M. C. Whitlock (1998), „Single found-flush events and the evolution of reproductive izolace“, Vývoj, 52 (6): 1850–1855, doi:10.1111 / j.1558-5646.1998.tb02263.x, PMID 28565304, S2CID 24502821CS1 maint: více jmen: seznam autorů (odkaz)
- ^ A. Ø. Mooers, H. D. Rundle a M. C. Whitlock (1999), „Účinky výběru a překážek na úspěch páření u periferních izolátů“, Americký přírodovědec, 153 (4): 437–444, doi:10.1086/303186, PMID 29586617CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Jeffrey R. Powell (1978), "The Founder-Flush Speciation Theory: An Experimental Approach", Vývoj, 32 (3): 465–474, doi:10.1111/j.1558-5646.1978.tb04589.x, PMID 28567948, S2CID 30943286
- ^ Diane M. B. Dodd & Jeffrey R. Powell (1985), "Founder-Flush Speciation: An Update of Experimental Results with Drosophila", Vývoj, 39 (6): 1388–1392, doi:10.1111/j.1558-5646.1985.tb05704.x, PMID 28564258, S2CID 34137489
| Základní pojmy | ||
|---|---|---|
| Geografické režimy | ||
| Izolační faktory | ||
| Hybridizace | ||
| Speciace v taxonech | ||
| Vývoj |  | |
|---|---|---|
| Populace genetika | ||
| Rozvoj | ||
| Z taxony | ||
| Z orgány | ||
| Z procesy | ||
| Tempo a režimy | ||
| Speciace | ||
| Dějiny | ||
| Filozofie | ||
| Příbuzný | ||
| ||