Proteiny vázající železo - Iron-binding proteins
Proteiny vázající železo jsou nosné proteiny a metaloproteiny které jsou důležité v železa metabolismus[1] a imunitní odpověď.[2][3] Železo je nutné pro život.
Enzymy závislé na železu katalyzují celou řadu různých biochemických reakcí a lze je rozdělit do tří širokých tříd v závislosti na struktuře jejich aktivního místa: mononukleární klastry železa, centra Rieske (Fe-S) nebo centra hemu.[4] Známá rodina enzymů závislých na železu zahrnuje oxygenázy, které usnadňují přidání hydroxylové skupiny jednoho nebo obou atomů z diatomického kyslíku. Pozoruhodné enzymy zahrnují tryptofan dioxygenázu, ferredoxin a 2-oxoglutarát dioxygenázu (hlavní enzym v biosyntéze flavonoidů).[5]
Železná biologická dostupnost
Železo je nutné pro život. Nedostatek železa vede k anémie. V životním prostředí je železo velmi hojné, ale jen málo z něj je velmi biologicky dostupné.[6] Aby bylo biologicky užitečné, anorganické železo se obvykle redukuje z železitého na železné železitými reduktázami. Železné druhy mají tendenci být rozpustnější v blízkosti neutrálního pH. „Volné železo“ je cytotoxické, protože se ho účastní Fentonova chemie k výrobě reaktivní formy kyslíku. Nadbytek železa tedy může být také škodlivý, jak dokazuje nemoc hemochromatóza.
Lidé obvykle získávají většinu své spotřeby železa z rostlinných nebo živočišných zdrojů. Toto železo může být hem (ve zvířecím mase) nebo jiné než hem (v rostlinách). Železo v lemu je mnohem více biologicky dostupné pro člověka (15-35% absorbováno), takže maso může být silným zdrojem železa v potravě.[6] Nehemové železo je obecně absorbováno pouze 2–20%. Většina železa ve stravě průměrného člověka však pochází z jiných než hemových zdrojů jednoduše proto, že se konzumuje vyšší množství rostlinných potravin.[7] Tělo pevně drží železo a jediný způsob, jak lidé vyhnat železo navíc, je ztráta krve. Proto je železo regulováno při absorpci, kde je absorbován relativně nízký podíl železa. Hepcidin je hormonální regulátor vychytávání železa ve střevě. Na střevních buněčných membránách transportér dvojmocného kovu 1 aktivně transportuje železo do buňky.[8] Poté je zachycen ve feritinu pro další použití. Prostřednictvím hepcidinu lze absorpci železa ve střevních buňkách vyladit podle fyziologických potřeb, jako je nedostatek železa nebo jeho nadbytek.
Hemové proteiny

Hemové proteiny jsou proteiny, které obsahují hemovou protetickou skupinu. Hémovou skupinu tvoří a porfyrin prsten koordinovaný s iontem železa. Čtyři atomy dusíku v porfyrinovém kruhu působí jako ligand pro železo ve středu. V mnoha případech je rovníkový porfyrin doplněn jedním nebo dvěma axiálními ligandy. Příkladem toho je hemoglobin, kde porfyrin pracuje společně s postranním řetězcem histidinu a vázaným O2 molekula, tvořící osmistěn komplex.
Syntéza porfyrinu (tj. Hemový anabolismus) je vysoce konzervativní ve všech oblastech života. Začíná to výrobou kyselina 8-aminolevulinová z glycinu a sukcinyl-CoA, následuje vytvořením tetrapyrrolového kruhu a končí přidáním atomu železa do chelatujícího kruhu. Syntéza porfyrinu je negativně regulována (tj. Nepřítomností / přítomností represoru) zvýšenými koncentracemi glukózy a hemů. Přítomnost těchto inhibitorů destabilizuje komplex mRNA a ribozomu a také brání jeho absorpci do mitochondrií pro biosyntézu mitochondriálních proteinů.
Hemoglobin
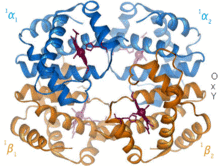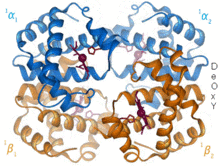
Hemoglobin je protein transportující kyslík, který se vyskytuje prakticky u všech obratlovců. Hemoglobin A je hlavní typ vyskytující se u dospělých lidí. Jedná se o tetramer skládající se ze dvou alfa a dvou beta podjednotek. Každá ze čtyř monomerních jednotek obsahuje hemovou protetickou skupinu, ve které je železitý kation vázán mezi čtyři atomy dusíku porfyrinového kruhu. Spolu s histidinem má forma apo pět ligandů obklopujících atom železa. Kyslík se váže na prázdnou šestou pozici a vytváří oktaedrický komplex v holo formě.[9] Vazba kyslíku je plně kooperativní pro každou z podjednotek, protože jelikož se první kyslík váže na jednu ze čtyř skupin hemu, protein prochází drastickou konformační změnou, která výrazně zvyšuje afinitu ke kyslíku ostatních tří podjednotek.[10]
Hemoglobin má různé afinity, v závislosti na pH, struktuře a CO2 částečný tlak. Fetální hemoglobin je varianta obsahující dvě podjednotky gama místo dvou podjednotek beta. Fetální hemoglobin je převládající formou až do několika měsíců věku kojence a má vyšší afinitu ke kyslíku, aby kompenzoval nízké kyslíkové napětí dodávané mateřské krve během těhotenství.[11] Hemoglobin má nižší afinitu ke kyslíku při nízkém pH. To umožňuje rychlou disociaci, protože okysličený hemoglobin je transportován do buněk v celém těle. Kvůli CO2 produkce a vodná tvorba kyseliny uhličité v dýchajících buňkách, okysličený hemoglobin disociuje, aby dodal buňkám potřebný kyslík.[12] Hemoglobin má vazebnou afinitu k kysličník uhelnatý to je 250krát větší než u kyslíku. To je základ otrava oxidem uhelnatým, protože hemoglobin již nemůže transportovat kyslík do buněk.
Cytochromy
Cytochromy jsou enzymy obsahující hem, které působí jako transportéry s jedním elektronem, zejména jako elektronové raketoplány při oxidační fosforylaci a fotosyntéze. Mezi typy dobře studovaných cytochromů patří cytochromy ac, cytochromoxidáza a cytochrom P450.[13] Tyto proteiny fungují jako elektronové raketoplány přepínáním oxidačního stavu atomu železa hemu na železo (Fe2+) a železitý (Fe3+). Různé cytochromy v kombinaci s jinými redox-aktivními molekulami tvoří gradient standardních redukčních potenciálů, který zvyšuje účinnost energetické vazby během událostí přenosu elektronů.
Železo-sirné proteiny
Železo-sirné proteiny jsou ty, které mají strukturu železa, která zahrnuje síru. Železo a síra mohou mít v proteinech různé formy, ale nejčastější jsou [2Fe 2S] a [4Fe 4S]. Klastry jsou často spojeny s cysteinovými zbytky v proteinovém řetězci.[14]
Nehemové proteiny
Transferrin
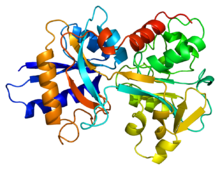
Transferrin se nachází v lidské plazmě a používá se k přenosu a importu nehemového železa.[15] Volně cestuje v extracelulárním prostoru.[16] Když jeho železo buňka potřebuje, je přivedeno do cytosol podle a receptor transferinu. Transferrin může vázat dva ionty Fe (III) spolu s aniontem (obvykle uhličitanem). K uvolnění železa je uhličitanový anion protonován. To mění interakci uhličitanu s proteinem, mění konformaci a umožňuje přenos Fe (III).
Transferin má molekulovou hmotnost přibližně 80 kDa. Je to glykoprotein, což znamená, že má na svém aminokyselinovém řetězci navázané cukry.
Laktoferin
Laktoferin je členem rodiny transferinů a je převládajícím proteinem, který se nachází v exokrinních sekrecích savců, jako jsou slzy, mléko a sliny. Skládá se z přibližně 700 zbytků a existuje hlavně jako tetramer, přičemž poměr monomer: tetramer je 1: 4 při koncentraci proteinu 10 μM.[17] Terciární struktura se skládá ze dvou laloků, nazývaných laloky N a C, z nichž každý obsahuje jednu kapsu vázající železo. Každá kapsa přispívá čtyřmi aminokyselinami (dva tyrosiny, jeden histidin a jeden aspartát) a spolu se dvěma uhličitanovými nebo hydrogenuhličitanovými anionty tvoří šestičlennou souřadnici kolem kationu železa. Je to tato specifická kombinace, která zvyšuje afinitu laktoferrinu k železa 300krát vyšší než transferin.[18]

Laktoferin má významné antimikrobiální vlastnosti. Nachází se v nejvyšší koncentraci 150 ng / ml v lidském mlezivu (typ mléka produkovaného v konečných stádiích těhotenství), který poskytuje tolik potřebnou imunitní podporu nově narozeným dětem.[19] Všeobecně se věřilo, že laktoferin je pouze bakteriostatickým činidlem kvůli jeho vysoké afinitě k železu a jeho schopnosti izolovat volné atomy železa od patogenních mikrobů. Nyní je však známo, že hlavní antimikrobiální hnací síla spočívá v baktericidních vlastnostech kapsy vázané na železo a specifického peptidu laktoferricinu umístěného na N-laloku. Laktoferin je schopen se vázat na LPS (lipopolysacharid ) vrstva bakterií a ve své holo formě atom železa oxiduje lipopolysacharidy, aby lyžovaly vnější membránu a současně produkovaly toxický peroxid vodíku.[20] Navíc se štěpením laktoferinu trypsinem vytvoří peptid laktoferricin, který se váže na H+-ATPáza, narušuje translokaci protonů a nakonec zabíjí buňku.[21]
Feritin

Ferritin je rezervoár železa pro jednotlivé buňky. Nachází se ve všech typech buněk a je lokalizován v cytosolu. Ferritin je velký protein složený z 24 podjednotek obklopujících jádro plné atomů železa. Je schopen pojmout 0-4500 atomů železa,[22] který lze použít jako rezervoár pro buněčné potřeby. Železo se skladuje, když je nadbytek, a znovu se vyhledává, když je železo znovu potřeba.[15] Podjednotky jsou směsí H (těžká nebo srdeční) a L (lehká nebo játrová). Podjednotky tvoří shluk široký 70-80 Angstromů, který je poté naplněn železitým ferithydritem.[23]
Ferritin je vysoce konzervovaný protein ve všech oblastech života. Je to tak konzervativní, že podjednotky koní a lidí se mohou spojit do funkčního proteinu.[15] Každá podjednotka se skládá z pěti alfa šroubovic.
Ferritin se používá k diagnostice nízké hladiny železa u lidí.[22] Může být použit k indikaci hladiny biologicky dostupného železa, což je užitečné při diagnostice anémie. Obvyklé rozmezí pro muže je 18-270 ng / ml a rozmezí pro ženy je 18-160 ng / ml.[24]
Viz také
externí odkazy
- Vazba na železo + proteiny v americké národní lékařské knihovně Lékařské předměty (Pletivo)
Reference
- ^ Brock JH (1989). "Proteiny vázající železo". Acta Paediatrica Scandinavica. Doplněk. 361: 31–43. doi:10.1111 / apa.1989.78.s361.31. PMID 2485582. S2CID 44752615.
- ^ De Sousa M, Breedvelt F, Dynesius-Trentham R, Trentham D, Lum J (1988). „Železo, proteiny vázající železo a buňky imunitního systému“. Annals of the New York Academy of Sciences. 526 (1): 310–22. Bibcode:1988NYASA.526..310S. doi:10.1111 / j.1749-6632.1988.tb55515.x. PMID 3291685. S2CID 12756539.
- ^ Kaplan J, Ward DM (srpen 2013). „Základní podstata používání a regulace železa“. Aktuální biologie. 23 (15): R642-6. doi:10.1016 / j.cub.2013.05.033. PMC 3928970. PMID 23928078.
- ^ Solomon EI, Brunold TC, Davis MI, Kemsley JN, Lee SK, Lehnert N, Neese F, Skulan AJ, Yang YS, Zhou J (leden 2000). "Geometrická a elektronová struktura / funkce korelace v nehemových železných enzymech". Chemické recenze. 100 (1): 235–350. doi:10.1021 / cr9900275. PMID 11749238.
- ^ Cheng AX, Han XJ, Wu YF, Lou HX (leden 2014). „Funkce a katalýza 2-oxoglutarát-dependentních oxygenáz podílejících se na biosyntéze rostlinných flavonoidů“. International Journal of Molecular Sciences. 15 (1): 1080–95. doi:10,3390 / ijms15011080. PMC 3907857. PMID 24434621.
- ^ A b Abbaspour N, Hurrell R, Kelishadi R (únor 2014). „Recenze železa a jeho významu pro lidské zdraví“. Journal of Research in Medical Sciences. 19 (2): 164–74. PMC 3999603. PMID 24778671.
- ^ Monsen ER, Hallberg L, Layrisse M, Hegsted DM, Cook JD, Mertz W, Finch CA (leden 1978). "Odhad dostupného dietního železa". American Journal of Clinical Nutrition. 31 (1): 134–41. doi:10.1093 / ajcn / 31.1.134. PMID 619599.
- ^ "Absorpce železa". kurzy. washington.edu. Citováno 2018-11-17.
- ^ Menis, Oscar (1968). „Technická poznámka 454“. Sekce analytické koordinační chemie: Shrnutí činností od července 1967 do června 1968. Gaithersburg, MD: Oddělení analytické koordinační chemie Oddělení analytické chemie Ústav pro výzkum materiálů. doi:10,6028 / nbs.tn.454.
- ^ Mihailescu MR, Russu IM (březen 2001). „Podpis přechodu T ---> R v lidském hemoglobinu“. Sborník Národní akademie věd Spojených států amerických. 98 (7): 3773–7. Bibcode:2001PNAS ... 98,3773M. doi:10.1073 / pnas.071493598. PMC 31128. PMID 11259676.
- ^ Berg JM, Tymoczko JL, Stryer L (2002). Biochemie (5. vydání). New York: W.H. Freemane. ISBN 0716730510. OCLC 48055706.
- ^ Hasselbalch KA (prosinec 1964). „Výpočet koncentrace vodíkových iontů v krvi z vazby volného a vázaného kyslíku na kyslík jako funkci Ph“. Průzkum anesteziologie. 8 (6): 607–32. doi:10.1097/00132586-196412000-00059.
- ^ L., Lehninger, Albert (2000). Lehningerovy principy biochemie. Nelson, David L. (David Lee), 1942-, Cox, Michael M. (3. vyd.). New York: Worth Publishers. ISBN 1572591536. OCLC 42619569.
- ^ Johnson, Deborah C .; Dean, Dennis R .; Smith, Archer D .; Johnson, Michael K. (18. února 2005). "Struktura, funkce a tvorba biologických klastrů železa a síry". Roční přehled biochemie. 74 (1): 247–281. doi:10,1146 / annurev.biochem.74.082803.133518. ISSN 0066-4154. PMID 15952888.
- ^ A b C Aisen P, Enns C, Wessling-Resnick M (říjen 2001). "Chemie a biologie eukaryotického metabolismu železa". International Journal of Biochemistry & Cell Biology. 33 (10): 940–59. doi:10.1016 / s1357-2725 (01) 00063-2. PMID 11470229.
- ^ "TF - prekurzor serotransferinu - Homo sapiens (člověk) - TF gen a protein". www.uniprot.org. Citováno 2018-11-11.
- ^ Baker EN, Baker HM (listopad 2005). "Molekulární struktura, vazebné vlastnosti a dynamika laktoferinu". Buněčné a molekulární biologické vědy. 62 (22): 2531–9. doi:10.1007 / s00018-005-5368-9. PMID 16261257.
- ^ Mazurier J, Spik G (květen 1980). "Srovnávací studie vazebných vlastností železa pro lidské transferiny. I. Úplná a postupná saturace a desaturace laktotransferrinu železem". Biochimica et Biophysica Acta. 629 (2): 399–408. doi:10.1016/0304-4165(80)90112-9. PMID 6770907.
- ^ Sánchez L, Calvo M, Brock JH (květen 1992). "Biologická role laktoferinu". Archiv nemocí v dětství. 67 (5): 657–61. doi:10.1136 / př. 67.5.657. PMC 1793702. PMID 1599309.
- ^ Farnaud S, Evans RW (listopad 2003). "Laktoferin - multifunkční protein s antimikrobiálními vlastnostmi". Molekulární imunologie. 40 (7): 395–405. doi:10.1016 / S0161-5890 (03) 00152-4. PMID 14568385.
- ^ Kuwata H, Yip TT, Yip CL, Tomita M, Hutchens TW (duben 1998). „Baktericidní doména laktoferinu: detekce, kvantifikace a charakterizace laktoferricinu v séru pomocí afinitní hmotnostní spektrometrie SELDI“. Sdělení o biochemickém a biofyzikálním výzkumu. 245 (3): 764–73. doi:10.1006 / bbrc.1998.8466. PMID 9588189.
- ^ A b Andrews NC (červenec 2008). „Kování pole: zlatý věk biologie železa“. Krev. 112 (2): 219–30. doi:10.1182 / krev-2007-12-077388. PMC 2442739. PMID 18606887.
- ^ Crichton RR, Charloteaux-Wauters M (květen 1987). "Přeprava a skladování železa". European Journal of Biochemistry. 164 (3): 485–506. doi:10.1111 / j.1432-1033.1987.tb11155.x. PMID 3032619.
- ^ „Co je to feritinový krevní test? Co znamenají výsledky?“. WebMD. Citováno 2018-11-11.