Superoxid dismutáza - Superoxide dismutase
| Superoxid dismutáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Struktura lidského tetrameru superoxid dismutázy 2 Mn.[1] | |||||||||
| Identifikátory | |||||||||
| EC číslo | 1.15.1.1 | ||||||||
| Číslo CAS | 9054-89-1 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Superoxid dismutáza (DRN, ES 1.15.1.1 ) je enzym který střídavě katalyzuje vyvrácení (nebo rozdělení) na superoxid (Ó2−) radikální do obyčejných molekulárních kyslík (Ó2) a peroxid vodíku (H2Ó2). Superoxid se vyrábí jako vedlejší produkt metabolismu kyslíku a pokud není regulován, způsobuje mnoho druhů poškození buněk.[2] Peroxid vodíku je také škodlivý a je degradován jinými enzymy, jako je kataláza. SOD je tedy důležitý antioxidant obrana téměř ve všech živých buňkách vystavených kyslíku. Jedna výjimka je Lactobacillus plantarum a související laktobacily, které používají jiný mechanismus k zabránění poškození reaktivním O2−.
Chemická reakce
SOD katalyzují nepřiměřenost superoxidu:
- 2 HO2 → O.2 + H2Ó2
Tímto způsobem, O2− se přeměňuje na dva méně škodlivé druhy.
Cesta, kterou katalyzoval SOD vyvrácení z superoxid lze psát pro Cu, Zn SOD, s následujícími reakcemi:
- Cu2+-SOD + O.2− → Cu+-SOD + O.2 (redukce mědi; oxidace superoxidu)
- Cu+-SOD + O.2− + 2 hodiny+ → Cu2+-SOD + H2Ó2 (oxidace mědi; redukce superoxidu)
Obecný formulář, který lze použít pro všechny různé formy SOD koordinované kovem, lze napsat následovně:
- M(n + 1) +-SOD + O.2− → M.n +-SOD + O.2
- Mn +-SOD + O.2− + 2 hodiny+ → M.(n + 1) +-SOD + H2Ó2.
kde M = Cu (n = 1); Mn (n = 2); Fe (n = 2); Ni (n = 2).
V sérii takových reakcí oxidační stav a náboj kovu kation osciluje mezi n a n + 1: +1 a +2 pro Cu, nebo +2 a +3 pro ostatní kovy.
Typy
Všeobecné
Irwin Fridovich a Joe McCord na Duke University objevil enzymatickou aktivitu superoxiddismutázy v roce 1968.[3] SOD byly dříve známé jako skupina metaloproteiny s neznámou funkcí; například CuZnSOD byl známý jako erythrocuprein (nebo hemocuprein nebo cytocuprein) nebo jako veterinární protizánětlivé léčivo "Orgotein".[4] Podobně Brewer (1967) identifikoval protein, který se později stal známým jako superoxiddismutáza jako indofenoloxidáza, pomocí proteinové analýzy škrobových gelů pomocí techniky fenazin-tetrazolium.[5]
Existují tři hlavní rodiny superoxiddismutázy, v závislosti na složení proteinu a kovu kofaktor: typ Cu / Zn (který váže obojí měď a zinek ), Typy Fe a Mn (které se vážou buď žehlička nebo mangan ) a typ Ni (který se váže nikl ).
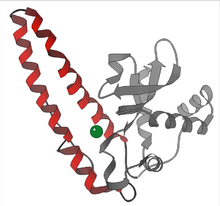
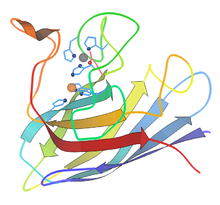
 Pás karet diagram hovězí podjednotky Cu-Zn SOD[6] |  Aktivní stránka lidského manganu SOD, mangan zobrazený fialově[7] |  Dimery Mn-SOD vs Fe-SOD |
- Měď a zinek - nejčastěji používané eukaryoty, včetně lidí. The cytosoly prakticky všech eukaryotický buňky obsahují enzym SOD s měď a zinek (Cu-Zn-SOD). Například komerčně dostupný Cu-Zn-SOD se obvykle čistí z kravských červených krvinek. Hovězí enzym Cu-Zn je homodimer o molekulové hmotnosti 32 500. Jednalo se o první SOD, jehož struktura atomových detailů byla vyřešena v roce 1975.[8] Je to 8vláknové “Řecký klíč „Beta-hlaveň, přičemž aktivní místo je udržováno mezi válcem a dvěma povrchovými smyčkami. Obě podjednotky jsou pevně spojeny zády k sobě, většinou hydrofobními a některými elektrostatickými interakcemi. Ligandů mědi a zinku je šest histidin a jeden aspartát postranní řetězce; jeden histidin je vázán mezi dvěma kovy.[9]
- Železo nebo mangan - používané společností prokaryoty a protistů a v mitochondrie a chloroplasty
 Aktivní místo pro superoxid dismutázu železa
Aktivní místo pro superoxid dismutázu železa- Železo - Mnoho bakterií obsahuje formu enzymu s žehlička (Fe-SOD); některé bakterie obsahují Fe-SOD, jiné Mn-SOD a některé (např E-coli ) obsahují obě. Fe-SOD lze také nalézt v chloroplasty rostlin. 3D struktury homologních Mn a Fe superoxiddismutáz mají stejné uspořádání alfa-šroubovic a jejich aktivní místa obsahují stejný typ a uspořádání postranních řetězců aminokyselin. Obvykle jsou to dimery, ale občas tetramery.
- Mangan - téměř vše mitochondrie, a mnoho bakterie, obsahují formulář s mangan (Mn-SOD): Například Mn-SOD nalezený v lidských mitochondriích. Ligandy manganových iontů jsou 3 histidin postranní řetězy, an aspartát postranní řetězec a molekula vody nebo hydroxy ligand, v závislosti na oxidačním stavu Mn (respektive II a III).[10]
- Nikl - prokaryotický. To má hexamerickou strukturu (6 kopií) postavenou z pravostranných svazků se 4 šroubovicemi, z nichž každý obsahuje N-koncové háky, které chelatují Ni iont. Ni-háček obsahuje motiv His-Cys-X-X-Pro-Cys-Gly-X-Tyr; poskytuje většinu interakcí kritických pro vazbu kovu a katalýzu, a je proto pravděpodobnou diagnostikou NiSOD.[11][12]

|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



U vyšších rostlin byly SOD izozymy lokalizovány v různých buněčných kompartmentech. Mn-SOD je přítomen v mitochondriích a peroxisomy. Fe-SOD byl nalezen hlavně v chloroplastech, ale byl také detekován v peroxisomech a CuZn-SOD byl lokalizován v cytosol, chloroplasty, peroxisomy a apoplast.[14][15]
Člověk
Ve všech ostatních jsou u lidí přítomny tři formy superoxiddismutázy savci a většina strunatci. SOD1 se nachází v cytoplazma, SOD2 v mitochondrie, a SOD3 je extracelulární. První je a dimer (skládá se ze dvou jednotek), zatímco ostatní jsou tetramery (čtyři podjednotky). SOD1 a SOD3 obsahují měď a zinek, zatímco SOD2, mitochondriální enzym, obsahuje mangan ve svém reaktivním centru. The geny jsou umístěny na chromozomech 21, 6 a 4 (21q22.1, 6q25.3 a 4p15.3-p15.1).
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Rostliny
Ve vyšších rostlinách působí enzymy superoxiddismutázy (SOD) jako antioxidanty a chrání buněčné složky před oxidací reaktivní formy kyslíku (ROS).[18] ROS se může tvořit v důsledku sucha, poranění, herbicidů a pesticidů, ozonu, metabolické aktivity rostlin, nedostatku živin, fotoinhibice, teploty nad a pod zemí, toxických kovů a UV nebo gama záření.[19][20] Konkrétně molekulární O2 se redukuje na O2− (ROS zvaný superoxid), když absorbuje vzrušený elektron uvolněný ze sloučenin elektronového transportního řetězce. Je známo, že superoxid denaturuje enzymy, oxiduje lipidy a fragmentuje DNA.[19] SOD katalyzují produkci O2 a H2Ó2 ze superoxidu (O2−), což má za následek méně škodlivé reaktanty.
Při aklimatizaci na zvýšené hladiny oxidačního stresu se koncentrace SOD typicky zvyšují se stupněm stresových podmínek. Rozdělení různých forem SOD v celé rostlině umožňuje velmi účinně působit proti stresu. V rostlinách existují tři známé a studované třídy kovových koenzymů SOD. Nejprve Fe SOD sestávají ze dvou druhů, jednoho homodimeru (obsahujícího 1-2 g Fe) a jednoho tetrameru (obsahujícího 2-4 g Fe). Jsou považovány za nejstarší metaloenzymy SOD a nacházejí se v prokaryotech i eukaryotech. Fe SOD jsou nejhojněji lokalizovány uvnitř rostlinných chloroplastů, kde jsou původní. Zadruhé, Mn SOD se skládají z druhů homodimeru a homotetrameru, z nichž každý obsahuje jeden atom Mn (III) na podjednotku. Vyskytují se převážně v mitochondriích a peroxisomech. Za třetí, Cu-Zn SOD mají elektrické vlastnosti velmi odlišné od vlastností ostatních dvou tříd. Ty jsou soustředěny v chloroplast, cytosol, a v některých případech mimobuněčný prostor. Pamatujte, že Cu-Zn SOD poskytují menší ochranu než Fe SOD, pokud jsou lokalizovány v chloroplastu.[18][19][20]
Bakterie
Lidské bílé krvinky používají enzymy, jako je NADPH oxidáza generovat superoxid a další reaktivní formy kyslíku k zabíjení bakterií. Během infekce mohou být některé bakterie (např. Burkholderia pseudomallei ) proto produkují superoxid dismutázu, aby se ochránily před zabitím.[21]
Biochemie
SOD konkuruje škodlivým reakcím superoxidu, čímž chrání buňku před toxicitou superoxidu. Reakce superoxidu s neradikály je zakázáno otáčení. V biologických systémech to znamená, že jeho hlavní reakce jsou samy se sebou (dismutace) nebo s jiným biologickým radikálem, jako je oxid dusnatý (NO) nebo s kovem přechodové řady. Superoxidový aniontový radikál (O2−) spontánně dismutuje na O2 a peroxid vodíku (H2Ó2) poměrně rychle (~ 105 M−1s−1 při pH 7).[Citace je zapotřebí ] SOD je nezbytný, protože superoxid reaguje s citlivými a kritickými buněčnými cíli. Například reaguje s NO radikálem a je toxický peroxynitrit.
Protože nekatalyzovaná dismutační reakce na superoxid vyžaduje vzájemnou reakci dvou molekul superoxidu, je rychlost dismutace druhého řádu vzhledem k počáteční koncentraci superoxidu. Takže poločas superoxidu, i když je velmi vysoký při vysokých koncentracích (např. 0,05 sekundy při 0,1 mM), je ve skutečnosti poměrně dlouhý při nízkých koncentracích (např. 14 hodin při 0,1 nM). Naproti tomu reakce superoxidu s SOD je prvního řádu s ohledem na koncentraci superoxidu. Superoxiddismutáza má navíc největší kkočka/K.M (přibližná katalytická účinnost) jakéhokoli známého enzymu (~ 7 x 109 M−1s−1),[22] tato reakce je omezena pouze frekvencí srážky mezi sebou a superoxidem. To znamená, že reakční rychlost je „omezena difúzí“.
Vysoká účinnost superoxiddismutázy se jeví jako nezbytná: i při subnanomolárních koncentracích dosažených vysokými koncentracemi SOD v buňkách superoxid inaktivuje enzym cyklu citronové kyseliny akonitáza, může otrávit energetický metabolismus a uvolňuje potenciálně toxické železo. Akonitáza je jednou z několika (de) hydratáz obsahujících železo a síru v metabolických drahách, u nichž se prokázalo, že jsou inaktivovány superoxidem.[23]
Stabilita a skládací mechanismus
SOD1 je extrémně stabilní protein. V holoformě (vázané na měď i zinek) je teplota tání> 90 ° C. V apo formě (bez vazby na měď nebo zinek) je teplota tání ~ 60 ° C.[24] Podle diferenční skenovací kalorimetrie (DSC), holo SOD1 se odvíjí dvoustavovým mechanismem: od dimeru po dva rozložené monomery.[24] Chemicky denaturace experimenty, holo SOD1 se odvíjí pomocí třístupňového mechanismu s pozorováním složeného monomerního meziproduktu.[25]
Fyziologie
Superoxid je jedním z hlavních reaktivní formy kyslíku v cele. V důsledku toho má SOD klíčovou antioxidační roli. Fyziologický význam SOD je ilustrován závažnými patologiemi evidentními u myší geneticky upravených pro nedostatek těchto enzymů. Myši postrádající SOD2 umírají několik dní po narození, uprostřed masy oxidační stres.[26] U myší postrádajících SOD1 se vyvíjí široká škála patologických stavů, včetně hepatocelulárního karcinomu,[27] zrychlení úbytku svalové hmoty související s věkem,[28] dřívější výskyt katarakty a zkrácení životnosti. Myši postrádající SOD3 nevykazují žádné zjevné vady a vykazují normální životnost, i když jsou citlivější na hyperoxické poškození.[29] Vyřazené myši kteréhokoli enzymu SOD jsou citlivější na letální účinky sloučenin generujících superoxid, jako je např parakvat a diquat (herbicidy ).
Drosophila bez SOD1 mají dramaticky zkrácenou životnost, zatímco mouchy bez SOD2 umírají před narozením. Vyčerpání SOD1 a SOD2 v nervovém systému a svalech Drosophila je spojena se sníženou životností.[30] Akumulace neuronů a svalů ROS Zdá se, že přispívá k poškozením souvisejícím s věkem. Když je indukována nadměrná exprese mitochondriálního SOD2, životnost dospělého Drosophila je prodloužena.[31]
Mezi černé zahradní mravenci (Lasius niger), životnost královny je řádově větší než u pracovníků, i když mezi nimi není žádný systematický rozdíl v sekvenci nukleotidů.[32] The SOD3 Bylo zjištěno, že gen je nejvíce diferencovaně nadměrně exprimován v mozcích mravenců královny vs. Toto zjištění zvyšuje možnost důležité role antioxidační funkce v modulaci délky života.[32]
SOD knockdowny v červu C. elegans nezpůsobují závažné fyziologické poruchy. Životnost C. elegans lze prodloužit superoxidem /kataláza mimetici tomu nasvědčují oxidační stres je hlavním určujícím činitelem míry stárnutí.[33]
Knockout nebo nulové mutace v SOD1 jsou vysoce škodlivé pro aerobní růst začínajících kvasinek Saccharomyces cerevisiae a vést k dramatickému snížení postdiauxické délky života. V divokém typu S. cerevisiae, Poškození DNA sazby se s věkem zvýšily 3krát, ale více než 5krát u mutantů odstraněných buď pro SOD1 nebo SOD2 geny.[34] Reaktivní formy kyslíku hladiny se s věkem u těchto mutantních kmenů zvyšují a vykazují podobný vzorec jako vzorec Poškození DNA s věkem roste. Zdá se tedy, že superoxiddismutáza hraje podstatnou roli při zachování integrity genomu během stárnutí v S. cerevisiae.SOD2 knockout nebo nulové mutace způsobují kromě snížené post-diauxické délky života také inhibici růstu na dýchacích zdrojích uhlíku.
Ve štěpném droždí Schizosaccharomyces pombe nedostatek mitochondriální superoxiddismutázy SOD2 urychluje chronologické stárnutí.[35]
Bylo vygenerováno několik prokaryotických nulových mutantů, včetně E-coli. Ztráta periplazmatického CuZnSOD způsobuje ztrátu virulence a může být atraktivním cílem pro nová antibiotika.
Role v nemoci
Mutace v prvním enzymu SOD (SOD1 ) může způsobit familiární Amyotrofní laterální skleróza (ALS, forma onemocnění motorických neuronů ).[36][37][38][39] Nejběžnější mutací v USA je A4V, zatímco nejintenzivněji studovaný je G93A. Další dvě izoformy SOD nebyly spojeny s mnoha lidskými chorobami, avšak u myší inaktivace SOD2 způsobuje perinatální letalitu[26] a deaktivace příčin SOD1 hepatocelulární karcinom.[27] Mutace v SOD1 může způsobit familiární ALS (několik důkazů také ukazuje, že divoký typ SOD1 je za podmínek buněčného stresu zapojen do významné části sporadických případů ALS, což představuje 90% pacientů s ALS.),[40] mechanismem, který v současné době není znám, ale nikoli kvůli ztrátě enzymatické aktivity nebo snížení konformační stability proteinu SOD1. Nadměrná exprese SOD1 byla spojena s nervovými poruchami pozorovanými u Downův syndrom.[41] U pacientů s talasemií se SOD zvýší jako forma kompenzačního mechanismu. V chronickém stadiu se však SOD nejeví jako dostatečný a má tendenci se snižovat v důsledku destrukce proteinů masivní reakcí oxidantu s antioxidantem.[42]
U myší přispívá extracelulární superoxiddismutáza (SOD3, ecSOD) k rozvoji hypertenze.[43][44] Snížená aktivita SOD3 byla spojena s plicními chorobami, jako je syndrom akutní respirační tísně (ARDS) nebo chronická obstrukční plicní nemoc (COPD).[45][46][47]
Superoxid dismutáza také není ve vývoji v buňkách nervového hřebenu exprimována plod. Vysoké hladiny volných radikálů proto mohou způsobit jejich poškození a vyvolat dysrafické anomálie (defekty neurální trubice).[Citace je zapotřebí ]
Farmakologická aktivita
SOD má silnou protizánětlivou aktivitu. Například SOD je vysoce účinná experimentální léčba chronického zánětu v kolitida.[Citace je zapotřebí ] Léčba SOD klesá reaktivní formy kyslíku generace a oxidační stres a tím inhibuje aktivaci endotelu. Proto mohou být tyto antioxidanty důležitými novými terapiemi pro léčbu zánětlivé onemocnění střev.[48]
Podobně má SOD několik farmakologických aktivit. Např. Zlepšuje cis-platina -indukovaný nefrotoxicita u hlodavců.[49] Jako „Orgotein" nebo „ontosein", farmakologicky aktivní čištěná hovězí játra SOD, je také účinný při léčbě zánětlivých onemocnění močových cest u člověka.[50] Po určitou dobu měl SOD z hovězích jater pro takové použití dokonce regulační schválení v několika evropských zemích. Toto bylo zkráceno obavami o prionová nemoc.[Citace je zapotřebí ]
An SOD-mimetikum činidlo, TEMPOL, je v současné době v klinických studiích pro radioprotekci a pro prevenci radiační indukce dermatitida.[51] TEMPOL a podobné SOD-mimetické nitroxidy vykazují mnohočetné účinky u onemocnění zahrnujících oxidační stres.[52]
Kosmetické použití
SOD může snížit poškození kůže volnými radikály - například ke snížení fibrózy po ozařování rakoviny prsu. Studie tohoto druhu musí být považovány za předběžné, protože ve studii nebyly adekvátní kontroly, včetně nedostatečné randomizace, dvojitého zaslepení nebo placeba.[53] Je známo, že superoxiddismutáza reverzuje fibróza, možná prostřednictvím de-diferenciace z myofibroblasty zpět k fibroblasty.[54][je třeba další vysvětlení ]
Komerční zdroje
SOD je komerčně získáván z námořnictva fytoplankton, hovězí játra, křen, ananasový meloun a určité bakterie. Z terapeutických důvodů se SOD obvykle injektuje lokálně. Neexistují žádné důkazy o tom, že požití nechráněného SOD nebo potravin bohatých na SOD může mít nějaké fyziologické účinky, protože všechny požité SOD jsou zlomený do aminokyseliny před vstřebává. Avšak požití SOD vázaného na pšeničné proteiny by mohlo zlepšit jeho terapeutickou aktivitu, alespoň teoreticky.[55]
Viz také
- Kataláza
- Glutathionperoxidáza
- Jiaogulan
- NADPH oxidáza enzym, který vyrábí superoxid
- Peroxidáza
Reference
- ^ A b PDB: 1VAR; Borgstahl GE, Parge HE, Hickey MJ, Johnson MJ, Boissinot M, Hallewell RA, Lepock JR, Cabelli DE, Tainer JA (duben 1996). "Lidská mitochondriální polymorfní varianta Ile58Thr superoxid dismutázy manganu snižuje aktivitu destabilizací tetramerního rozhraní". Biochemie. 35 (14): 4287–97. doi:10.1021 / bi951892w. PMID 8605177. S2CID 7450190.
- ^ Hayyan M, Hashim MA, Al Nashef IM (2016). „Superoxidový ion: tvorba a chemické důsledky“. Chem. Rev. 116 (5): 3029–3085. doi:10.1021 / acs.chemrev.5b00407. PMID 26875845.
- ^ McCord JM, Fridovich I (listopad 1969). "Superoxid dismutáza. Enzymatická funkce pro erytrocuprein (hein)". The Journal of Biological Chemistry. 244 (22): 6049–55. PMID 5389100.
- ^ McCord JM, Fridovich I (1988). „Superoxid dismutáza: prvních dvacet let (1968–1988)“. Radikální biologie a medicína zdarma. 5 (5–6): 363–9. doi:10.1016/0891-5849(88)90109-8. PMID 2855736.
- ^ Brewer GJ (září 1967). „Achromatické oblasti škrobových gelů obarvených tetrazoliem: zděděná elektroforetická variace“. American Journal of Human Genetics. 19 (5): 674–80. PMC 1706241. PMID 4292999.
- ^ PDB: 2SOD;Tainer JA, Getzoff ED, Beem KM, Richardson JS, Richardson DC (září 1982). "Stanovení a analýza struktury 2 A mědi, disperzi oxidu zinečnatého zinečnatého". J. Mol. Biol. 160 (2): 181–217. doi:10.1016/0022-2836(82)90174-7. PMID 7175933.
- ^ Quint P, Reutzel R, Mikulski R, McKenna R, Silverman DN (únor 2006). "Krystalová struktura nitrované lidské manganové superoxid dismutázy: mechanismus inaktivace". Radikální biologie a medicína zdarma. 40 (3): 453–8. doi:10.1016 / j.freeradbiomed.2005.08.045. PMID 16443160.
- ^ Richardson J, Thomas KA, Rubin BH, Richardson DC (duben 1975). „Krystalová struktura hovězího Cu, superoxiddismutáza Zn při rozlišení 3 A: trasování řetězce a kovové ligandy“. Sborník Národní akademie věd Spojených států amerických. 72 (4): 1349–53. doi:10.1073 / pnas.72.4.1349. PMC 432531. PMID 1055410..
- ^ Tainer JA, Getzoff ED, Richardson JS, Richardson DC (1983). "Struktura a mechanismus mědi, superoxiddismutázy zinečnaté". Příroda. 306 (5940): 284–7. Bibcode:1983 Natur.306..284T. doi:10.1038 / 306284a0. PMID 6316150. S2CID 4266810.
- ^ A b C PDB: 1N0J; Borgstahl GE, Parge HE, Hickey MJ, Beyer WF, Hallewell RA, Tainer JA (říjen 1992). „Struktura lidské mitochondriální superoxiddismutázy manganu odhaluje nové tetramerní rozhraní dvou svazků se 4 šroubovicemi“. Buňka. 71 (1): 107–18. doi:10.1016 / 0092-8674 (92) 90270-M. PMID 1394426. S2CID 41611695.
- ^ Barondeau DP, Kassmann CJ, Bruns CK, Tainer JA, Getzoff ED (červen 2004). "Struktura a mechanismus superoxiddismutázy niklu". Biochemie. 43 (25): 8038–47. doi:10.1021 / bi0496081. PMID 15209499. S2CID 10700340.
- ^ A b PDB: 1Q0M; Wuerges J, Lee JW, Yim YI, Yim HS, Kang SO, Djinovic Carugo K (červen 2004). „Krystalová struktura superoxiddismutázy obsahující nikl odhaluje jiný typ aktivního místa“. Sborník Národní akademie věd Spojených států amerických. 101 (23): 8569–74. Bibcode:2004PNAS..101 8669 W.. doi:10.1073 / pnas.0308514101. PMC 423235. PMID 15173586.
- ^ PDB: 1 SDY; Djinović K, Gatti G, Coda A, Antolini L, Pelosi G, Desideri A, Falconi M, Marmocchi F, Rolilio G, Bolognesi M (prosinec 1991). "Strukturální řešení a zdokonalení molekulární dynamiky kvasinek Cu, Zn, enzym superoxid dismutáza". Acta Crystallogr. B. 47 (6): 918–27. doi:10.1107 / S0108768191004949. PMID 1772629.
- ^ Corpas FJ, Barroso JB, del Río LA (duben 2001). „Peroxisomy jako zdroj reaktivních forem kyslíku a signálních molekul oxidu dusnatého v rostlinných buňkách“. Trendy ve vědě o rostlinách. 6 (4): 145–50. doi:10.1016 / S1360-1385 (01) 01898-2. PMID 11286918.
- ^ Corpas FJ, Fernández-Ocaña A, Carreras A, Valderrama R, Luque F, Esteban FJ, Rodríguez-Serrano M, Chaki M, Pedrajas JR, Sandalio LM, del Río LA, Barroso JB (červenec 2006). „Exprese různých forem superoxiddismutázy je závislá na buněčném typu v listech oliv (Olea europaea L.)“. Fyziologie rostlin a buněk. 47 (7): 984–94. doi:10.1093 / pcp / pcj071. PMID 16766574.
- ^ PDB: 3CQQ; Cao X, Antonyuk SV, Seetharaman SV, Whitson LJ, Taylor AB, Holloway SP, Strange RW, Doucette PA, Valentine JS, Tiwari A, Hayward LJ, Padua S, Cohlberg JA, Hasnain SS, Hart PJ (červen 2008). "Struktury varianty G85R SOD1 u familiární amyotrofické laterální sklerózy". J. Biol. Chem. 283 (23): 16169–77. doi:10,1074 / jbc.M801522200. PMC 2414278. PMID 18378676.
- ^ PDB: 2JLP; Antonyuk SV, Strange RW, Marklund SL, Hasnain SS (květen 2009). "Struktura lidské extracelulární měď-zinek-superoxid dismutázy při rozlišení 1,7 A: pohledy na vazbu heparinu a kolagenu". J. Mol. Biol. 388 (2): 310–26. doi:10.1016 / j.jmb.2009.03.026. PMID 19289127.
- ^ A b Alscher RG, Erturk N, Heath LS (květen 2002). „Role superoxiddismutáz (SOD) při kontrole oxidačního stresu v rostlinách“. Journal of Experimental Botany. 53 (372): 1331–41. doi:10.1093 / jexbot / 53.372.1331. PMID 11997379.
- ^ A b C Smirnoff, Nicholas (1993). „Tansley Review No. 52 Role aktivního kyslíku v reakci rostlin na nedostatek vody a vysychání“. Nový fytolog. 125: 27–58. doi:10.1111 / j.1469-8137.1993.tb03863.x.
- ^ A b Raychaudhuri SS, Deng XW (2008). „Role superoxiddismutázy v boji proti oxidačnímu stresu u vyšších rostlin“. The Botanical Review. 66 (1): 89–98. doi:10.1007 / BF02857783. S2CID 7663001.
- ^ Vanaporn M, Wand M, Michell SL, Sarkar-Tyson M, Irsko P, Goldman S, Kewcharoenwong C, Rinchai D, Lertmemongkolchai G, Titball RW (srpen 2011). „Superoxid dismutáza C je nutná pro intracelulární přežití a virulenci Burkholderia pseudomallei“. Mikrobiologie. 157 (Pt 8): 2392–400. doi:10.1099 / mic.0.050823-0. PMID 21659326.
- ^ Heinrich PC, Löffler G, Petrifies PE (2006). Biochemie und Pathobiochemie (Springer-Lehrbuch) (Německé vydání). Berlín: Springer. p. 123. ISBN 978-3-540-32680-9.
- ^ Gardner PR, Raineri I, Epstein LB, White CW (červen 1995). „Superoxidový radikál a železo modulují aktivitu akonitázy v buňkách savců“. The Journal of Biological Chemistry. 270 (22): 13399–405. doi:10.1074 / jbc.270.22.13399. PMID 7768942.
- ^ A b Stathopulos PB, Rumfeldt JA, Karbassi F, Siddall CA, Lepock JR, Meiering EM (březen 2006). „Kalorimetrická analýza termodynamické stability a agregace pro mutanty Gly-93 superoxid dismutázy spojené s apo a holo amyotrofickou laterální sklerózou“. The Journal of Biological Chemistry. 281 (10): 6184–93. doi:10,1074 / jbc.M509496200. PMID 16407238.
- ^ Rumfeldt JA, Stathopulos PB, Chakrabarrty A, Lepock JR, Meiering EM (leden 2006). "Mechanismus a termodynamika guanidiniumchloridu indukované denaturace ALS asociovaného mutantu Cu, Zn superoxiddismutázy". Journal of Molecular Biology. 355 (1): 106–23. doi:10.1016 / j.jmb.2005.10.042. PMID 16307756.
- ^ A b Li Y, Huang TT, Carlson EJ, Melov S, Ursell PC, Olson JL, Noble LJ, Yoshimura MP, Berger C, Chan PH, Wallace DC, Epstein CJ (prosinec 1995). „Dilatační kardiomyopatie a neonatální letalita u mutovaných myší postrádajících superoxid dismutázu manganu“. Genetika přírody. 11 (4): 376–81. doi:10.1038 / ng1295-376. PMID 7493016. S2CID 10900822.
- ^ A b Elchuri S, Oberley TD, Qi W, Eisenstein RS, Jackson Roberts L, Van Remmen H, Epstein CJ, Huang TT (leden 2005). „Nedostatek CuZnSOD vede k přetrvávajícímu a rozšířenému oxidačnímu poškození a hepatokarcinogenezi později v životě“. Onkogen. 24 (3): 367–80. doi:10.1038 / sj.onc.1208207. PMID 15531919.
- ^ Muller FL, Song W, Liu Y, Chaudhuri A, Pieke-Dahl S, Strong R, Huang TT, Epstein CJ, Roberts LJ, Csete M, Faulkner JA, Van Remmen H (červen 2006). „Absence CuZn superoxiddismutázy vede ke zvýšenému oxidačnímu stresu a zrychlení atrofie kosterního svalstva závislé na věku“. Radikální biologie a medicína zdarma. 40 (11): 1993–2004. doi:10.1016 / j.freeradbiomed.2006.01.036. PMID 16716900.
- ^ Sentman ML, Granström M, Jakobson H, Reaume A, Basu S, Marklund SL (březen 2006). „Fenotypy myší bez extracelulární superoxiddismutázy a superoxiddismutázy obsahující měď a zinek“. The Journal of Biological Chemistry. 281 (11): 6904–9. doi:10,1074 / jbc.M510764200. PMID 16377630.
- ^ Oka S, Hirai J, Yasukawa T, Nakahara Y, Inoue YH (srpen 2015). „Korelace akumulace reaktivních druhů kyslíku vyčerpáním superoxiddismutáz s věkem závislým poškozením v nervovém systému a svalech dospělých Drosophila“. Biogerontologie. 16 (4): 485–501. doi:10.1007 / s10522-015-9570-3. PMID 25801590. S2CID 18050827.
- ^ Sun J, Folk D, Bradley TJ, Tower J (červen 2002). „Indukovaná nadměrná exprese mitochondriální Mn-superoxid dismutázy prodlužuje životnost dospělého Drosophila melanogaster“. Genetika. 161 (2): 661–72. PMC 1462135. PMID 12072463.
- ^ A b Lucas ER, Keller L (červenec 2018). "Zvýšené vyjádření genů stárnutí a imunity u královen mravence černé zahradní". Experimentální gerontologie. 108: 92–98. doi:10.1016 / j.exger.2018.03.020. PMID 29625209. S2CID 5045743.
- ^ Melov S, Ravenscroft J, Malik S, Gill MS, Walker DW, Clayton PE, Wallace DC, Malfroy B, Doctrow SR, Lithgow GJ (září 2000). "Prodloužení délky života s mimetiky superoxiddismutázy / katalázy". Věda. 289 (5484): 1567–9. Bibcode:2000 sci ... 289,1567 mil. doi:10.1126 / science.289.5484.1567. PMID 10968795. S2CID 21519801.
- ^ Muid KA, Karakaya HÇ, Koc A (únor 2014). „Absence aktivity superoxiddismutázy způsobuje fragmentaci jaderné DNA během procesu stárnutí“ (PDF). Biochem. Biophys. Res. Commun. 444 (2): 260–3. doi:10.1016 / j.bbrc.2014.01.056. hdl:11147/5542. PMID 24462872.
- ^ Ogata T, Senoo T, Kawano S, Ikeda S (leden 2016). „Nedostatek mitochondriální superoxiddismutázy urychluje chronologické stárnutí štěpných kvasinek Schizosaccharomyces pombe“. Cell Biol. Int. 40 (1): 100–6. doi:10,1002 / cbin.10556. PMID 26507459. S2CID 205563521.
- ^ Milani P, Gagliardi S, Cova E, Cereda C (2011). „Transkripční a posttranskripční regulace SOD1 a její možné důsledky v ALS“. Neurology Research International. 2011: 1–9. doi:10.1155/2011/458427. PMC 3096450. PMID 21603028.
- ^ Deng HX, Hentati A, Tainer JA, Iqbal Z, Cayabyab A, Hung WY, Getzoff ED, Hu P, Herzfeldt B, Roos RP (srpen 1993). "Amyotrofická laterální skleróza a strukturální vady Cu, Zn superoxid dismutázy". Věda. 261 (5124): 1047–51. Bibcode:1993Sci ... 261.1047D. doi:10.1126 / science.8351519. PMID 8351519.
- ^ Conwit RA (prosinec 2006). „Prevence familiární ALS: klinické hodnocení může být proveditelné, ale je nutné provést hodnocení účinnosti?“. Časopis neurologických věd. 251 (1–2): 1–2. doi:10.1016 / j.jns.2006.07.009. PMID 17070848. S2CID 33105812.
- ^ Al-Chalabi A, Leigh PN (srpen 2000). "Nedávný pokrok v amyotrofické laterální skleróze". Aktuální názor na neurologii. 13 (4): 397–405. doi:10.1097/00019052-200008000-00006. PMID 10970056. S2CID 21577500.
- ^ Gagliardi S, Cova E, Davin A, Guareschi S, Abel K, Alvisi E, Laforenza U, Ghidoni R, Cashman JR, Ceroni M, Cereda C (srpen 2010). "Exprese mRNA SOD1 při sporadické amyotrofické laterální skleróze". Neurobiologie nemocí. 39 (2): 198–203. doi:10.1016 / j.nbd.2010.04.008. PMID 20399857. S2CID 207065284.
- ^ Groner Y, Elroy-Stein O, Avraham KB, Schickler M, Knobler H, Minc-Golomb D, Bar-Peled O, Yarom R, Rotshenker S (1994). "Poškození buněk nadměrným CuZnSOD a Downovým syndromem". Biomedicína a farmakoterapie. 48 (5–6): 231–40. doi:10.1016/0753-3322(94)90138-4. PMID 7999984.
- ^ Rujito L, Mulatsih S, Sofro AS (květen 2015). „Stav superoxiddismutázy u talasemie závislé na transfuzi“. North American Journal of Medical Sciences. 7 (5): 194–8. doi:10.4103/1947-2714.157480. PMC 4462814. PMID 26110130.
- ^ Gongora MC, Qin Z, Laude K, Kim HW, McCann L, Folz JR, Dikalov S, Fukai T, Harrison DG (září 2006). „Role extracelulární superoxid dismutázy při hypertenzi“. Hypertenze. 48 (3): 473–81. doi:10.1161 / 01.HYP.0000235682.47673.ab. PMID 16864745.
- ^ Lob HE, Marvar PJ, Guzik TJ, Sharma S, McCann LA, Weyand C, Gordon FJ, Harrison DG (únor 2010). „Indukce hypertenze a periferního zánětu snížením extracelulární superoxid dismutázy v centrálním nervovém systému“. Hypertenze. 55 (2): 277–83, 6p po 283. doi:10.1161 / HYPERTENSIONAHA.109.142646. PMC 2813894. PMID 20008675.
- ^ Young RP, Hopkins R, Black PN, Eddy C, Wu L, Gamble GD, Mills GD, Garrett JE, Eaton TE, Rees MI (květen 2006). „Funkční varianty antioxidačních genů u kuřáků s CHOPN au těch s normální funkcí plic“. Hrudník. 61 (5): 394–9. doi:10.1136 / tis.2005.048512. PMC 2111196. PMID 16467073.
- ^ Ganguly K, Depner M, Fattman C, Bein K, Oury TD, Wesselkamper SC, Borchers MT, Schreiber M, Gao F, von Mutius E, Kabesch M, Leikauf GD, Schulz H (květen 2009). „Superoxid dismutáza 3, extracelulární varianty (SOD3) a funkce plic“. Fyziologická genomika. 37 (3): 260–7. doi:10.1152 / physiolgenomics.90363.2008. PMC 2685504. PMID 19318538.
- ^ Gongora MC, Lob HE, Landmesser U, Guzik TJ, Martin WD, Ozumi K, Wall SM, Wilson DS, Murthy N, Gravanis M, Fukai T, Harrison DG (říjen 2008). „Ztráta extracelulární superoxiddismutázy vede k akutnímu poškození plic v přítomnosti okolního vzduchu: potenciální mechanismus syndromu respirační tísně dospělých“. American Journal of Pathology. 173 (4): 915–26. doi:10.2353 / ajpath.2008.080119. PMC 2543061. PMID 18787098.
- ^ Seguí J, Gironella M, Sans M, Granell S, Gil F, Gimeno M, Coronel P, Piqué JM, Panés J (září 2004). „Superoxiddismutáza zlepšuje kolitidu vyvolanou TNBS snížením oxidačního stresu, exprese adhezních molekul a náboru leukocytů do zaníceného střeva“. Journal of Leukocyte Biology. 76 (3): 537–44. doi:10.1189 / jlb.0304196. PMID 15197232. S2CID 15028921.
- ^ McGinness JE, Proctor PH, Demopoulos HB, Hokanson JA, Kirkpatrick DS (1978). „Zlepšení nefrotoxicity cis-platiny orgoteinem (superoxid dismutázou)“. Fyziologická chemie a fyzika. 10 (3): 267–77. PMID 733940.
- ^ Marberger H, Huber W, Bartsch G, Schulte T, Swoboda P (1974). „Orgotein: nové protizánětlivé hodnocení metaloproteinových léků z hlediska klinické účinnosti a bezpečnosti při zánětlivých stavech močových cest“. Mezinárodní urologie a nefrologie. 6 (2): 61–74. doi:10.1007 / bf02081999. PMID 4615073. S2CID 23880216.
- ^ Číslo klinického hodnocení NCT01324141 pro „Aktuální MTS-01 pro dermatitidu během záření a chemoterapii pro rakovinu konečníku“ na ClinicalTrials.gov
- ^ Wilcox CS (květen 2010). "Účinky tempolu a redox cyklujících nitroxidů v modelech oxidačního stresu". Farmakologie a terapeutika. 126 (2): 119–45. doi:10.1016 / j.pharmthera.2010.01.003. PMC 2854323. PMID 20153367.
- ^ Campana F, Zervoudis S, Perdereau B, Gez E, Fourquet A, Badiu C, Tsakiris G, Koulaloglou S (2004). „Topická superoxiddismutáza snižuje po ozáření fibrózu rakoviny prsu“. Journal of Cellular and Molecular Medicine. 8 (1): 109–16. CiteSeerX 10.1.1.336.8033. doi:10.1111 / j.1582-4934.2004.tb00265.x. PMC 6740277. PMID 15090266.
- ^ Vozenin-Brotons MC, Sivan V, Gault N, Renard C, Geffrotin C, Delanian S, Lefaix JL, Martin M (leden 2001). „Antifibrotický účinek Cu / Zn SOD je zprostředkován represí TGF-beta1 a fenotypovou reverzí myofibroblastů.“ Radikální biologie a medicína zdarma. 30 (1): 30–42. doi:10.1016 / S0891-5849 (00) 00431-7. PMID 11134893.
- ^ Romao S (březen 2015). „Terapeutická hodnota orálního doplňku s kombinací meloun-superoxid dismutázy a pšeničného gliadinu“. Výživa. 31 (3): 430–6. doi:10.1016 / j.nut.2014.10.006. PMID 25701330.
externí odkazy
- Online Mendelian Inheritance in Man (OMIM): 105400 (ALS)
- Online databáze ALS
- Krátký, ale věcný přehled SOD a jeho literatury.
- Teorie stárnutí založené na poškození Zahrnuje diskusi o rolích SOD1 a SOD2 ve stárnutí.
- Komunikace lékařů Za odpovědnou med.
- Obrázek dráhy SOD a oxidačního stresu
- Historické informace o výzkumu SOD "Vývoj Radikální biologie a medicína zdarma: 20letá historie „a“Radikální biologie a medicína zdarma Posledních 20 let: nejcitovanější články “
- JM McCord pojednává o objevu SOD
- PDBe-KB poskytuje přehled všech strukturních informací dostupných v PDB pro lidskou superoxid dismutázu [Cu-Zn]
- PDBe-KB poskytuje přehled všech strukturních informací dostupných v PDB pro lidskou superoxid dismutázu [Mn], mitochondriální
- PDBe-KB poskytuje přehled všech strukturních informací dostupných v PDB pro lidskou extracelulární superoxid dismutázu [Cu-Zn]