Ribonukleotid reduktáza - Ribonucleotide reductase
| ribonukleosid-difosfátreduktáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||||
| EC číslo | 1.17.4.1 | ||||||||
| Číslo CAS | 9047-64-7sy | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Ribonukleotid reduktáza (RNR), také známý jako ribonukleotid difosfátreduktáza (rNDP), je enzym který katalyzuje vznik deoxyribonukleotidy z ribonukleotidy.[1] Katalyzuje tuto tvorbu odstraněním 2'-hydroxylové skupiny ribózového kruhu nukleosiddifosfátů. Tato redukce produkuje deoxyribonukleotidy.[2] Deoxyribonukleotidy se zase používají při syntéze DNA. Reakce katalyzovaná RNR je přísně zachována u všech živých organismů.[3] Kromě toho hraje RNR zásadní roli v regulaci celkové rychlosti syntézy DNA tak, aby během dne byla DNA k buněčné hmotě udržována v konstantním poměru buněčné dělení a Oprava DNA.[4] Poněkud neobvyklým rysem enzymu RNR je, že katalyzuje reakci, která probíhá prostřednictvím a volné radikály mechanismus účinku.[5][6] Substráty pro RNR jsou ADP, HDP, CDP a UDP. dTDP (deoxythymidíndifosfát) je syntetizován jiným enzymem (thymidylát kináza ) z dTMP (deoxytymidinmonofosfát).
Struktura
Ribonukleotid reduktázy jsou rozděleny do tří tříd. Enzymy RNR třídy I jsou konstruovány z velké podjednotky alfa a malých podjednotek beta, které se sdružují za vzniku aktivní látky heterodimerní tetramer. Snížením NDP na 2'-dNDPs enzym katalyzuje de novo syntéza deoxyribonukleotidů (dNTP), které jsou prekurzory syntézy DNA a jsou nezbytné pro proliferace buněk.[7] RNR třídy II produkují 5'-deoxyadenosylový radikál homolytickým štěpením vazby C-Co v adenosylkobalaminu. Kromě toho RNR třídy III obsahují stabilní glycylový radikál.[8]
Lidé mají RNR třídy I. Alfa podjednotka je kódována genem RRM1, zatímco existují dvě izoformy beta podjednotky kódované geny RRM2 a RRM2B:
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Každá třída I alfa monomer skládá se ze tří domén:[9]
- jedna převážně spirálovitá doména zahrnující 220 N-terminál zbytky,
- druhou velkou desetvláknovou strukturu α / β obsahující 480 zbytků,
- a třetí malou pětřetězcovou strukturu a / p obsahující 70 zbytků.
v Pfam, druhá doména byla interpretována jako dvě samostatné domény:
- kratší all-alfa N-terminální doména,
- a delší hlavní C-koncová doména.
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
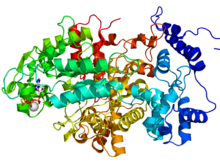
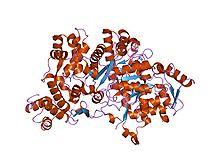


Beta podjednotka třídy I obvykle obsahuje dvojkovový střed a stáj tyrosyl radikální. U lidí se beta podjednotka spoléhá na kofaktor s dvojitým obsahem železa. v E-coli, tyrosyl radikál je umístěn v poloze 122 (Y122) poskytující stabilní radikál pro podjednotky RNR2 třídy I.[13] v A. aegypti, tento tyrosyl radikál je umístěn v poloze 184 (Y184).[14] Tyrosyl radikál je hluboce pohřben uvnitř proteinu v hydrofobním prostředí, které se nachází v blízkosti centra železa, které se používá ke stabilizaci tyrosyl radikálu. Ve struktuře dvou μ-oxo vázaných žehliček dominují ligandy, které slouží jako vazebná místa pro železo: čtyři karboxyláty [aspartát (D146), glutamát (E177, E240 a E274)] a dva histidiny (H180 a H277).[14] Asociace nastává mezi C-koncem RNR2 a C-konec RNR1.[9] Enzymatická aktivita závisí na asociaci podjednotek RNR1 a RNR2. Aktivní místo se skládá z aktivních dithiolových skupin z RNR1 a také z diferenciálního centra a tyrosylového radikálu z podjednotky RNR2.
Další zbytky RNR2, jako je aspartát (D273), tryptofan (W48) a tyrosin (Y356) dále stabilizují tyrosylový radikál v aktivním místě, což umožňuje přenos elektronů.[9] Tyto zbytky pomáhají při přenosu radikálového elektronu z tyrosinu (Y122) RNR2 na cystein (C439) RNR1. Přenos elektronů začíná na RNR2 tyrosinu (Y122) a pokračuje v RNR2 na tryptofan (W48), který je oddělen od RNR1 tyrosinu (Y731) o 2,5 nanometry. K přenosu elektronů z RNR2 na RNR1 dochází prostřednictvím tyrosinu (Y356 až Y731) a pokračuje dále prostřednictvím tyrosinu (Y730) na cystein (C439) v aktivním místě.[15] Místně cílené mutace primární struktury RNR naznačují, že všechny zbytky uvedené výše se účastní přenosu volných radikálů na dlouhé vzdálenosti do aktivního místa.[9]
v A. aegypti komáři, RNR1 zachovává většinu rozhodujících aminokyselinových zbytků, včetně aspartátu (D64) a valinu (V292 nebo V284), které jsou nezbytné alosterická regulace; prolin (P210 a P610), leucin (L453 a L473) a methionin (M603) zbytky, které jsou umístěny v hydrofobním aktivním místě; cysteinové (C225, C436 a C451) zbytky, které se podílejí na odstraňování atomu vodíku a přenosu radikálového elektronu na aktivním místě; cystein (C225 a C436), asparagin (N434) a zbytky glutamátu (E441), které se vážou na ribonukleotidový substrát; tyrosinové (Y723 a Y743) zbytky, které určují přenos radikálů; a cysteinové (C838 a C841) zbytky, které se používají při regeneraci dithiolových skupin v aktivním místě.[14]
Funkce
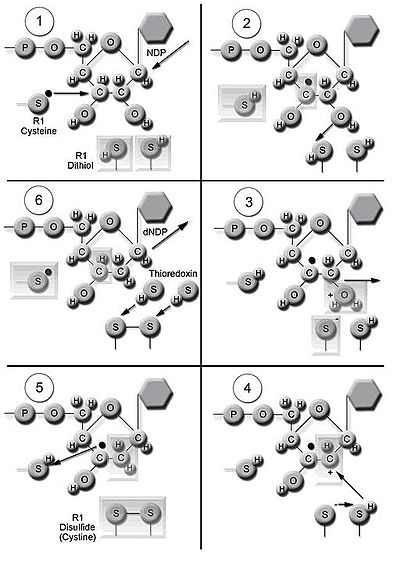
Enzym ribonukleotidreduktáza (RNR) katalyzuje de novo syntézu dNDP.[16] Katalýza ribonukleosidových 5’-difosfátů (NDP) zahrnuje redukci na 2’-uhlíku ribóza-5-fosfát za vzniku 2’-deoxyderivátu redukovaného 2’-deoxyribonukleosidového 5’-difosfátu (dNDP). Tato redukce je zahájena generací volných radikálů. Po jediné redukci vyžaduje RNR elektrony darované z dithiolových skupin proteinu thioredoxin. Regenerace thioredoxinu nastává, když je nikotinamid adenin dinukleotid fosfát (NADPH ) poskytuje dva atomy vodíku, které se používají ke snížení disulfid skupiny thioredoxinu.
Tři třídy RNR mají podobné mechanismy pro snížení NDP, ale liší se v doméně, která generuje volný radikál, specifický kov v metaloprotein struktura a dárci elektronů. Všechny třídy používají chemii volných radikálů.[9] Reduktázy třídy I používají železné centrum s přeměnou ze železa na železo k vytvoření volného radikálu tyrosyl. Redukce NDP substrátů nastává za aerobních podmínek. Reduktázy třídy I se dělí na IA a IB kvůli rozdílům v regulaci. Reduktázy třídy IA jsou distribuovány v eukaryoty, eubakterie, bakteriofágy, a viry. Reduktázy třídy IB se nacházejí v eubakteriích. Reduktázy třídy IB mohou také použít radikál generovaný stabilizací binukleária mangan centrum. Reduktázy třídy II generují volný radikál 5’-deoxyadenosyl radikál z kobalamin (koenzym B12) a mají jednodušší strukturu než reduktázy třídy I a třídy III. Ke snížení NDP nebo ribonukleotidových 5’-trifosfátů (NTP) dochází buď aerobní nebo anaerobní podmínky. Reduktázy třídy II jsou distribuovány v archaebakterie, eubakterie a bakteriofágy. Reduktázy třídy III používají glycinový radikál vytvořený pomocí S-adenosyl methionin a centrum železné síry. Snížení NTP je omezeno na anaerobní podmínky. Reduktázy třídy III jsou distribuovány v archebakteriích, eubakteriích a bakteriofágech.[9][14] Organismy se neomezují pouze na jednu třídu enzymů. Například, E-coli mít RNR třídy I i třídy III.
Katalytický redukční mechanismus
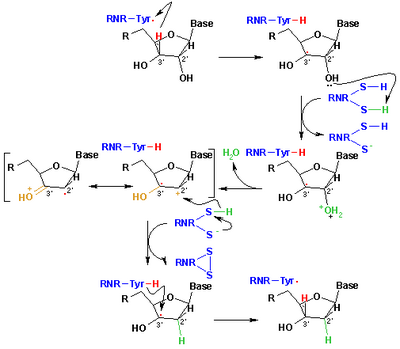
Mechanismus, který je v současné době přijímán pro redukci ribonukleotidů na deoxyribonukleotidy, je znázorněn v následujícím schématu. První krok zahrnuje abstrakci 3’- H substrátu 1 radikálem Cys439. Následně reakce zahrnuje eliminaci jedné molekuly vody z uhlíku C-2 'ribonukleotidu, katalyzovaného Cys225 a Glu441. Ve třetím kroku dochází k přenosu atomu vodíku z Cys225 na uhlík C-2 '2’-ketylového radikálu 3 po předchozím přenosu protonů z Cys462 na Cys225. Na konci tohoto kroku se získá radikál aniontový disulfidový můstek a ketonový meziprodukt 4 s uzavřeným obalem. Tento meziprodukt byl identifikován během převodu několika 2’-substituovaných analogů substrátu, stejně jako u přírodního substrátu[17] interakce s enzymovými mutanty. Dalším krokem je oxidace aniontového disulfidového můstku se současnou redukcí substrátu, generování 5. Spinální hustota se posune z atomů síry na atom C-3 'substrátu se současným přenosem protonů z Glu441 na uhlík C -3 '. Poslední krok je opakem prvního kroku a zahrnuje přenos vodíku z Cys439 na C-3 ', regeneraci počátečního radikálu a výsledný produkt 6.
Teoretické modely některých kroků těchto mechanismů využívajících plný model proteinu R1 lze nalézt ve studiích provedených Cerqueirou et al..[18][19]
Nařízení
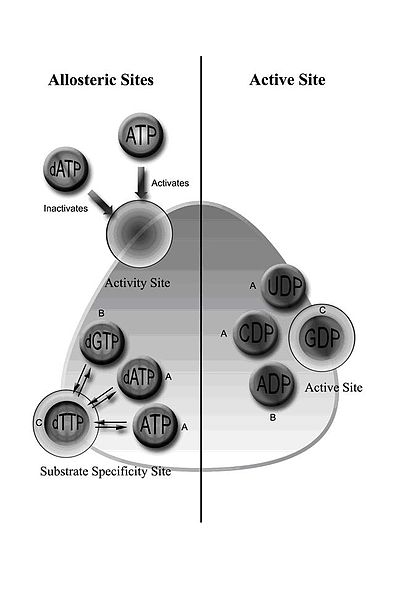
Třída I RNR zahrnuje RNR1 a RNR2 podjednotky, které se mohou sdružovat za vzniku heterodimerního tetrameru.[5] RNR1 obsahuje obě alosterická místa zprostředkující regulaci specificity a aktivity substrátu.[11] V závislosti na alosterické konfiguraci se jeden ze čtyř ribonukleotidů váže na aktivní místo.
Regulace RNR je navržena tak, aby udržovala vyvážené množství dNTP. Vazba efektorových molekul buď zvyšuje nebo snižuje aktivitu RNR. Když se ATP váže na místo alosterické aktivity, aktivuje RNR. Naproti tomu, když se dATP váže na tuto stránku, deaktivuje RNR.[9] Kromě kontroly aktivity alosterický mechanismus také reguluje specificitu substrátu a zajišťuje, že enzym produkuje stejné množství každého dNTP pro syntézu DNA.[9] Ve všech třídách navázání ATP nebo dATP na alosterické místo indukuje redukci cytidinu 5’-difosfátu (CDP) a uridinu 5’-difosfátu (UDP); 2’-deoxyguanosin 5’-trifosfát (dGTP) indukuje redukci adenosin 5’-difosfátu (ADP); a 2’-deoxythymidin 5’-trifosfát (dTTP) indukuje snížení guanosin 5’-difosfátu (GDP) (obrázek 1).
Reduktázy třídy IB nejsou inhibovány dATP, protože jim chybí přibližně 50 N-koncových aminokyselin požadovaných pro místo alosterické aktivity.[20] Dále je důležité, aby aktivita ribonukleotidreduktázy byla pod transkripční a posttranskripční kontrolou, protože syntéza DNA bez poškození závisí na vyváženém množství deoxyribonukleotidů.[21] Eukaryotické buňky s reduktázami třídy IA mají mechanismus negativní kontroly k vypnutí syntézy dNTP při jejich akumulaci. Tento mechanismus chrání buňku před toxickými a mutagenními účinky, které mohou vzniknout z nadprodukce dNTP, protože změny ve vyvážených zásobách dNTP vedou k poškození DNA a buněčné smrti.[22][23] I když nadprodukce dNTP nebo jejich nevyvážený přísun může vést k nesprávné inkorporaci nukleotidů do DNA, může dodávka dodávek dNTP umožnit opravu DNA. p53R2 je malá podjednotka ribonukleotidreduktázy, která může tuto opravu vyvolat. Změny v tomto homologu R2 indukovaném p53 mohou způsobit vyčerpání mitochondriální DNA a následně p53R2 slouží jako hlavní faktor v zásobování dNTP.[24]
RNR může používat morpheein model alosterická regulace.[25]
Inhibitory RNR1 a RNR2
Obecně lze inhibitory třídy I RNR rozdělit do tří hlavních skupin: inhibitory translace, které blokují syntézu enzymu; inhibitory dimerizace, které zabraňují asociaci dvou podjednotek RNR (R1 a R2); a katalytické inhibitory, které inaktivují podjednotku R1 a / nebo podjednotku R2.[18]
RNR třídy I může být inhibována peptidy podobně jako C-konec RNR2. Tyto peptidy mohou soutěžit s RNR2 o vazbu na RNR1 a ve výsledku RNR1 netvoří enzymaticky aktivní komplex s RNR2.[26][27] Ačkoli je C-konec proteinů RNR2 u různých druhů odlišný, může RNR2 interagovat s RNR1 u různých druhů.[28] Když byl myší RNR2 C-konec nahrazen E-coli C-terminální (7 nebo 33) aminokyselinové zbytky RNR2, chimérická podjednotka RNR2 se stále váže na myší podjednotky RNR1. Chybí jim však enzymatická aktivita pravděpodobně kvůli eliminaci zbytků podílejících se na přenosu elektronu volných radikálů z RNR2 na podjednotku RNR1.[27]
Malé peptidy mohou specificky inhibovat podjednotky RNR2 ve vazbě s RNR1, když mají významnou podobnost s normálním C-koncem RNR2.[29] Tato inhibiční vazba RNR2 na RNR1 byla úspěšně testována na RNR viru herpes simplex (HSV). Když byl v kompetičních testech použit 7 aminokyselinový oligomer (GAVVNDL) zkrácený z C-konce podjednotky RNR2, zabránilo to normálnímu RNR2 ve vytvoření enzymaticky aktivního komplexu s RNR1.[30] Jiné malé peptidové inhibitory podobné C-konci RNR2 byly také úspěšně použity k inhibici enzymatické aktivity HSV RNR a tím i replikace HSV.[31] U myší modely stromální keratitida a neovaskularizace rohovky (HSV oční onemocnění ), malý RNR2 C-terminální analog BILD 1263 uvádí, že inhibuje RNR a je účinný při prevenci těchto onemocnění.[32] V některých případech, i když léčba malými C-koncovými analogy nemusí zastavit šíření nemoci, mohou stále pomáhat při hojení. V acyklovir - rezistentní HSV (PAAr5), malý peptidový inhibitor BILD 1633 byl hlášen 5 až 10krát účinněji než BILD 1263 proti kožní infekci PAAr5.[33] Přístup kombinované terapie (BILD 1633 a acyklovir) je účinnější při hojení lokálních lézí u myší. Tato data naznačují, že malé peptidové inhibitory, které soutěží s RNR2 o vazbu na RNR1, jsou užitečné při prevenci šíření HSV.
Gallium inhibuje RNR2 nahrazením Fe3+ na aktivním webu. Gallium maltolát je orálně biologicky dostupná forma gália, která využívá tuto inhibiční aktivitu k léčbě rakoviny, infekcí a dalších nemocí.[34]
Drogy hydroxymočovina[35] a Motexafin gadolinium interferovat s působením tohoto enzymu.[36]
Reference
- ^ Elledge SJ, Zhou Z, Allen JB (březen 1992). „Ribonukleotid reduktáza: regulace, regulace, regulace“. Trendy v biochemických vědách. 17 (3): 119–23. doi:10.1016/0968-0004(92)90249-9. PMID 1412696.
- ^ Sneeden JL, Loeb LA (září 2004). „Mutace v podjednotce R2 ribonukleotidreduktázy, které propůjčují rezistenci vůči hydroxymočovině“. The Journal of Biological Chemistry. 279 (39): 40723–8. doi:10,1074 / jbc.M402699200. PMID 15262976.
- ^ Torrents E, Aloy P, Gibert I, Rodríguez-Trelles F (srpen 2002). „Ribonukleotid reduktázy: divergentní vývoj starověkého enzymu“. Journal of Molecular Evolution. 55 (2): 138–52. doi:10.1007 / s00239-002-2311-7. PMID 12107591. S2CID 24603578.
- ^ Herrick J, Sclavi B (leden 2007). „Ribonukleotid reduktáza a regulace replikace DNA: starý příběh a starověké dědictví“. Molekulární mikrobiologie. 63 (1): 22–34. doi:10.1111 / j.1365-2958.2006.05493.x. PMID 17229208. S2CID 9473163.
- ^ A b Eklund H, Eriksson M, Uhlin U, Nordlund P, Logan D (srpen 1997). „Ribonukleotid reduktáza - strukturální studie radikálního enzymu“. Biologická chemie. 378 (8): 821–5. doi:10.1515 / bchm.1997.378.8.815. PMID 9377477.
- ^ Stubbe J, Riggs-Gelasco P (listopad 1998). "Využití volných radikálů: tvorba a funkce tyrosylového radikálu v ribonukleotidreduktáze". Trendy v biochemických vědách. 23 (11): 438–43. doi:10.1016 / S0968-0004 (98) 01296-1. PMID 9852763.
- ^ Fairman JW, Wijerathna SR, Ahmad MF, Xu H, Nakano R, Jha S, Prendergast J, Welin RM, Flodin S, Roos A, Nordlund P, Li Z, Walz T, Dealwis CG (březen 2011). „Strukturální základ pro alosterickou regulaci lidské ribonukleotidreduktázy nukleotidem indukovanou oligomerací“. Přírodní strukturní a molekulární biologie. 18 (3): 316–22. doi:10.1038 / nsmb.2007. PMC 3101628. PMID 21336276.
- ^ Larsson KM, Jordan A, Eliasson R, Reichard P, Logan DT, Nordlund P (listopad 2004). "Strukturální mechanismus regulace specificity alosterického substrátu v ribonukleotidreduktáze". Přírodní strukturní a molekulární biologie. 11 (11): 1142–9. doi:10.1038 / nsmb838. PMID 15475969. S2CID 1025702.
- ^ A b C d E F G h Jordan A, Reichard P (1998). "Ribonukleotid reduktázy". Roční přehled biochemie. 67 (1): 71–98. doi:10.1146 / annurev.biochem.67.1.71. PMID 9759483.
- ^ A b PDB: 1PEU; Uppsten M, Färnegårdh M, Jordan A, Eliasson R, Eklund H, Uhlin U (červen 2003). „Struktura velké podjednotky ribonukleotidreduktázy třídy Ib ze Salmonella typhimurium a jejích komplexů s alosterickými efektory“. Journal of Molecular Biology. 330 (1): 87–97. doi:10.1016 / S0022-2836 (03) 00538-2. PMID 12818204.
- ^ A b Uhlin U, Eklund H (srpen 1994). "Struktura proteinu ribonukleotid reduktázy R1". Příroda. 370 (6490): 533–9. doi:10.1038 / 370533a0. PMID 8052308. S2CID 8940689.
- ^ Nordlund P, Eklund H (červenec 1993). "Struktura a funkce proteinu ribonukleotid reduktázy Escherichia coli R2". Journal of Molecular Biology. 232 (1): 123–64. doi:10.1006 / jmbi.1993.1374. PMID 8331655.
- ^ Högbom M, Andersson ME, Nordlund P (březen 2001). „Krystalové struktury oxidovaných dinukleárních center manganu v Mn-substituované ribonukleotidové reduktáze třídy I z Escherichia coli: karboxylátové posuny s důsledky pro aktivaci O2 a tvorbu radikálů“. Journal of Biological Anorganic Chemistry. 6 (3): 315–23. doi:10,1007 / s007750000205. PMID 11315567. S2CID 20748553.
- ^ A b C d Pham DQ, Blachuta BJ, Nichol H, Winzerling JJ (září 2002). „Ribonukleotid reduktázové podjednotky od komára žluté zimnice, Aedes aegypti: klonování a exprese“. Hmyzí biochemie a molekulární biologie. 32 (9): 1037–44. doi:10.1016 / S0965-1748 (02) 00041-3. PMID 12213240.
- ^ Chang MC, Yee CS, Stubbe J, Nocera DG (květen 2004). „Zapnutí ribonukleotid reduktázy generováním radikálů aminokyselin iniciovaných světlem“. Sborník Národní akademie věd Spojených států amerických. 101 (18): 6882–7. doi:10.1073 / pnas.0401718101. PMC 406436. PMID 15123822.
- ^ Cox M, Nelson DR (2008). Lehningerovy principy biochemie. San Francisco: W. H. Freeman. ISBN 978-0-7167-7108-1.
- ^ Cerqueira NM, Fernandes PA, Eriksson LA, Ramos MJ (prosinec 2004). "Aktivace ribonukleotidů enzymem ribonukleotidreduktázou: porozumění úloze enzymu". Journal of Computational Chemistry. 25 (16): 2031–7. doi:10.1002 / jcc.20127. PMID 15481089. S2CID 19665974.
- ^ A b Cerqueira NM, Pereira S, Fernandes PA, Ramos MJ (2005). „Přehled inhibitorů ribonukleotid reduktázy: přitažlivý cíl v protinádorové terapii“. Současná léčivá chemie. 12 (11): 1283–94. doi:10.2174/0929867054020981. PMID 15974997.
- ^ Cerqueira NM, Fernandes PA, Eriksson LA, Ramos MJ (březen 2006). „Dehydratace ribonukleotidů katalyzovaných ribonukleotidreduktázou: úloha enzymu“. Biofyzikální deník. 90 (6): 2109–19. doi:10.1529 / biophysj.104.054627. PMC 1386789. PMID 16361339.
- ^ Eliasson R, Pontis E, Jordan A, Reichard P (říjen 1996). „Alosterická regulace třetí ribonukleotidreduktázy (enzymu NrdEF) z enterobacteriaceae“. The Journal of Biological Chemistry. 271 (43): 26582–7. doi:10.1074 / jbc.271.43.26582. PMID 8900130.
- ^ Thelander L (červen 2007). "Ribonukleotidreduktáza a syntéza mitochondriální DNA". Genetika přírody. 39 (6): 703–4. doi:10.1038 / ng0607-703. PMID 17534360. S2CID 22565931.
- ^ Kunz BA (1988). "Mutageneze a nerovnováha zásoby deoxyribonukleotidů". Mutační výzkum. 200 (1–2): 133–47. doi:10.1016/0027-5107(88)90076-0. PMID 3292903.
- ^ Meuth M (duben 1989). „Molekulární základ mutací vyvolaných nerovnováhou deoxyribonukleosid trifosfátu v nerovnováze v buňkách savců“. Experimentální výzkum buněk. 181 (2): 305–16. doi:10.1016/0014-4827(89)90090-6. PMID 2647496.
- ^ Bourdon A, Minai L, Serre V, Jais JP, Sarzi E, Aubert S, Chrétien D, de Lonlay P, Paquis-Flucklinger V, Arakawa H, Nakamura Y, Munnich A, Rötig A (červen 2007). „Mutace RRM2B, kódující p53-kontrolovanou ribonukleotidreduktázu (p53R2), způsobuje závažné vyčerpání mitochondriální DNA“. Genetika přírody. 39 (6): 776–80. doi:10.1038 / ng2040. PMID 17486094. S2CID 22103978.
- ^ Selwood T, Jaffe EK (březen 2012). „Dynamické disociační homo-oligomery a řízení funkce proteinu“. Archivy biochemie a biofyziky. 519 (2): 131–43. doi:10.1016 / j.abb.2011.11.020. PMC 3298769. PMID 22182754.
- ^ Climent I, Sjöberg BM, Huang CY (květen 1991). „Karboxyl-terminální peptidy jako sondy pro interakci podjednotky ribonukleotid reduktázy Escherichia coli: kinetická analýza studií inhibice“. Biochemie. 30 (21): 5164–71. doi:10.1021 / bi00235a008. PMID 2036382.
- ^ A b Hamann CS, Lentainge S, Li LS, Salem JS, Yang FD, Cooperman BS (březen 1998). „Chimérické malé podjednotkové inhibitory savčí ribonukleotidreduktázy: dvojí funkce pro C-konec R2?“. Proteinové inženýrství. 11 (3): 219–24. doi:10.1093 / protein / 11.3.219. PMID 9613846.
- ^ Cosentino G, Lavallée P, Rakhit S, Plante R, Gaudette Y, Lawetz C, Whitehead PW, Duceppe JS, Lépine-Frenette C, Dansereau N (leden 1991). „Specifická inhibice ribonukleotidreduktáz peptidy odpovídajícími C-konci jejich druhé podjednotky“. Biochemie a buněčná biologie. 69 (1): 79–83. doi:10.1139 / o91-011. PMID 2043345.
- ^ Cooperman BS (2003). "Inhibice oligopeptidů třídy I ribonukleotidreduktáz". Biopolymery. 71 (2): 117–31. doi:10,1002 / bip.10397. PMID 12767114. S2CID 25196379.
- ^ Filatov D, Ingemarson R, Gräslund A, Thelander L (srpen 1992). „Role ribonukleotidreduktázy viru herpes simplex malé podjednotky karboxylový konec v interakci podjednotky a tvorbě středové struktury železo-tyrosyl“. The Journal of Biological Chemistry. 267 (22): 15816–22. PMID 1322407.
- ^ Cohen EA, Gaudreau P, Brazeau P, Langelier Y (1986). „Specifická inhibice herpesvirové ribonukleotidreduktázy nonapeptidem odvozeným od karboxykonce podjednotky 2“. Příroda. 321 (6068): 441–3. doi:10.1038 / 321441a0. PMID 3012360. S2CID 4238076.
- ^ Brandt CR, Spencer B, Imesch P, Garneau M, Déziel R (květen 1996). „Hodnocení peptidomimetického inhibitoru ribonukleotidreduktázy s myším modelem očního onemocnění virem herpes simplex typu 1“. Antimikrobiální látky a chemoterapie. 40 (5): 1078–84. doi:10.1128 / aac.40.5.1078. PMC 163269. PMID 8723444.
- ^ Duan J, Liuzzi M, Paris W, Lambert M, Lawetz C, Moss N, Jaramillo J, Gauthier J, Déziel R, Cordingley MG (červenec 1998). „Antivirová aktivita selektivního inhibitoru ribonukleotid reduktázy proti viru herpes simplex rezistentního na acyklovir typu 1 in vivo“. Antimikrobiální látky a chemoterapie. 42 (7): 1629–35. doi:10.1128 / aac.42.7.1629. PMC 105657. PMID 9660995.
- ^ Bernstein LR (prosinec 1998). "Mechanismy terapeutické aktivity pro gallium" (PDF). Farmakologické recenze. 50 (4): 665–82. PMID 9860806.
- ^ „Informace o EC 1.17.4.1 - ribonukleosid-difosfátreduktáza“. Brenda. Citováno 25. července 2015.
- ^ Hashemy SI, Ungerstedt JS, Zahedi Avval F, Holmgren A (duben 2006). „Motexafin gadolinium, nádorově selektivní léčivo zaměřené na thioredoxin reduktázu a ribonukleotid reduktázu“. The Journal of Biological Chemistry. 281 (16): 10691–7. doi:10,1074 / jbc.M511373200. PMID 16481328.
externí odkazy
- Ribonukleotid + reduktázy v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- Databáze ribonukleotid reduktázy (RNRdb)