Hsp90 - Hsp90 - Wikipedia
| Histidinkináza, DNA gyráza B a ATPáza podobná HSP90 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifikátory | |||||||||
| Symbol | HATPase_c | ||||||||
| Pfam | PF02518 | ||||||||
| Pfam klan | CL0025 | ||||||||
| InterPro | IPR003594 | ||||||||
| CHYTRÝ | SM00387 | ||||||||
| SCOP2 | 1ei1 / Rozsah / SUPFAM | ||||||||
| |||||||||
| Hsp90 protein | |||||||||
|---|---|---|---|---|---|---|---|---|---|
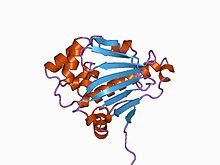 Struktura N-terminální domény kvasinkového chaperonu Hsp90.[2] | |||||||||
| Identifikátory | |||||||||
| Symbol | Hsp90 | ||||||||
| Pfam | PF00183 | ||||||||
| InterPro | IPR020576 | ||||||||
| STRÁNKA | PDOC00270 | ||||||||
| SCOP2 | 1ah6 / Rozsah / SUPFAM | ||||||||
| |||||||||




Hsp90 (protein tepelného šoku 90) je chaperonový protein který pomáhá jiným proteinům složit správně, stabilizuje proteiny proti tepelný stres a pomáhá v degradace bílkovin. Stabilizuje také řadu proteinů potřebných pro růst nádoru, a proto Inhibitory Hsp90 jsou zkoumány jako protinádorové léky.
Proteiny tepelného šoku, jako třída, patří mezi nejvíce vyjádřené buněčný bílkoviny napříč všemi druhy.[3] Jak jejich název napovídá, proteiny tepelného šoku chrání buňky, když jsou namáhány zvýšenými teplotami. Představují 1–2% celkového proteinu v nestresovaných buňkách. Když se však buňky zahřívají, zvyšuje se podíl proteinů tepelného šoku na 4–6% buněčných proteinů.[4]
Protein tepelného šoku 90 (Hsp90) je jedním z nejběžnějších proteinů souvisejících s teplem. „90“ pochází ze skutečnosti, že váží zhruba 90 kilo Daltonů. 90 kDa protein je považován za poměrně velký pro nevláknitý protein. Hsp90 se nachází v bakterie a všechny pobočky eukarya, ale zjevně chybí v archaea.[5] Zatímco cytoplazmatický Hsp90 je nezbytný pro životaschopnost za všech podmínek v eukaryoty bakteriální homolog HtpG je postradatelný v podmínkách bez tepelného stresu.[6]
Tento protein byl nejprve izolován extrakcí proteinů z buněk namáhaných zahříváním, dehydratací nebo jinými způsoby, což vše způsobilo, že proteiny buňky začaly denaturace.[7] Později však bylo zjištěno, že Hsp90 má také základní funkce v nepříznivých buňkách.
Izoformy
Hsp90 je vysoce konzervovaný a exprimovaný v různých organizmech od bakterií po savce - včetně prokaryotického analogu HtpG (vysokoteplotní protein G) se 40% sekvenční identitou a 55% podobností s lidským proteinem.[5] Droždí Hsp90 je ze 60% identický s lidským Hsp90a.
v savčí buňky, existují dva nebo více genů kódujících cytosolický Homology Hsp90,[5] s lidským Hsp90a vykazujícím 85% sekvenční identitu s Hsp90p.[8] Α- a β-formy jsou považovány za výsledek a genová duplikace událost, ke které došlo před miliony let.[5]
Pět funkčních lidských genů kódujících Hsp90 proteinové izoformy jsou uvedeny níže:[8]
| rodina | subcelulární umístění | podčeleď | gen | protein |
|---|---|---|---|---|
| HSP90A | cytosolický | HSP90AA (indukovatelné ) | HSP90AA1 | Hsp90-α1 |
| HSP90AA2 | Hsp90-α2 | |||
| HSP90AB (konstitutivně vyjádřeno ) | HSP90AB1 | Hsp90-p | ||
| HSP90B | endoplazmatický retikulum | HSP90B1 | Endoplazmin / GRP-94 | |
| PAST | mitochondriální | TRAP1 | TNF Receptor- Apřidružený Protein 1 |
Existuje 12 lidí pseudogeny (nefunkční geny), které kódují další izoformy Hsp90, které nejsou exprimovány jako proteiny.
Nedávno byla identifikována a byla pojmenována varianta cytosolického Hsp90 spojená s membránou, která postrádá vazebné místo pro ATP. Hsp90N.[9] Tento transkript HSP90α-Δ-N je chiméra s prvními 105 bp kódující sekvence odvozenou z CD47 gen na chromozomu 3q13.2 a zbývající kódující sekvence odvozená z HSP90AA1.[8] Později se však ukázalo, že Hsp90N kódující geny v lidském genomu neexistuje. Je to pravděpodobně klonovací artefakt nebo produkt chromozomálního přeskupení vyskytující se v jedné buněčné linii.[10]
Struktura
Společné rysy
Celková struktura Hsp90 je podobná struktuře jiných proteinů v tom, že obsahuje všechny běžné látky sekundární konstrukční prvky (tj., alfa šroubovice, skládané listy beta a náhodné cívky). Být a cytoplazmatický protein vyžaduje, aby protein měl strukturu kulovitého tvaru, která je z velké části nepolární na vnitřní straně a polární na vnější straně, aby byla rozpuštěna vodou. Hsp90 obsahuje devět šroubovic a osm antiparalelních skládaných listů beta, které kombinují a vytvářejí několik sendvičů alfa / beta. The 310 šroubovice tvoří přibližně 11% aminokyselinových zbytků proteinu, což je mnohem více než průměr 4% v jiných proteinech.[11]
Struktura domény
Hsp90 se skládá ze čtyř strukturálních domén:[12][13][14]
- vysoce konzervativní N-terminál doména (NTD) ~ 25 kDa
- oblast „nabitého linkeru“, která spojuje N-konec se střední doménou
- střední doména (MD) ~ 40 kDa
- A C-terminál doména (CTD) ~ 12 kDa.
Krystalové struktury jsou k dispozici pro N-koncovou doménu droždí a lidský Hsp90,[15][16][17] pro komplexy N-konce s inhibitory a nukleotidy,[15][16] a pro střední doménu kvasinek Hsp90.[18] Nedávno struktury pro celou délku Hsp90 z E-coli (2IOP, 2IOQ),[19] droždí (2CG9, 2CGE),[20] a psí endoplazmatické retikulum (2O1U, 2O1V)[21] byly objasněny.[22]
Formuláře Hsp90 homodimery kde jsou kontaktní místa lokalizována na C-konci v otevřené konformaci dimeru. N-konce také přicházejí do styku v uzavřené konformaci dimeru.[18]
N-terminální doména
Zobrazí se N-koncová doména homologie nejen mezi členy rodiny chaperonů Hsp90, ale také mezi členy ATPázy / kinázy GHKL (Gvýrok, Hsp90, Histidin K.inase, MutL ) nadčeleď.[13]
Společná vázací kapsa pro ATP a inhibitor geldanamycin se nachází v N-terminální doméně.[15][16] Aminokyseliny, které se přímo účastní interakce s ATP, jsou Leu34, Asn37, Asp79, Asn92, Lys98, Gly121 a Phe124. Kromě toho Mg2+ a několik molekul vody tvoří přemosťující elektrostatické a vodíkové vazby interakce mezi Hsp90 a ATP. Kromě toho je pro ATP vyžadován Glu33 hydrolýza.
Střední doména
Střední doména je rozdělena do tří oblastí:
- 3vrstvý sendvič α-β-α
- 3-závitová α-šroubovice a nepravidelné smyčky
- 6-závitová α-šroubovice.[13]
MD se také podílí na vazbě na klientské proteiny. Například proteiny, o nichž je známo, že interagují s Hsp90 MD, zahrnují PKB /Akt1, eNOS,[23][24] Aha1, Hch1. Dále je známo, že vazba substrátu (např. Aha1 a Hch1) na MD zvyšuje ATPáza aktivita Hsp90.[18][25]
C-koncová doména
C-koncová doména má alternativní ATP-vazebné místo, které je přístupné, když je obsazena N-koncová kapsa Bergeratu.[26][27]
Na samém C-terminálním konci proteinu je místo pro rozpoznávání motivu tetratrikopeptidové repetice (TPR), konzervovaný pentapeptid MEEVD, který je zodpovědný za interakci s kofaktory, jako je imunofiliny FKBP51 a FKBP52, stresem indukovaný fosfoprotein 1 (Sti1 / Hop), cyklofilin-40, PP5, Tom70, a mnoho dalších.[28]
Mechanismus
Protein Hsp90 obsahuje tři funkční domény, ATP - vazebná, proteinová a dimerizující doména, z nichž každá hraje klíčovou roli ve funkci proteinu.
ATP vazba
Oblast proteinu poblíž N-konec má vysoce afinitní vazebné místo pro ATP. ATP se váže na značnou štěrbinu na straně proteinu, což je 15A (1,5 nanometrů) hluboko. Tato štěrbina má vysokou afinitu k ATP, a když dostane vhodný proteinový substrát, Hsp90 štěpí ATP na ADP a Pi. Přímé inhibitory ATP vazby nebo alosterický inhibitory vazby ATP nebo aktivity ATPázy mohou blokovat funkci Hsp90.[11] Další zajímavou vlastností oblasti vázající ATP Hsp90 je to, že má „víčko“, které je otevřené během stavu vázaného na ADP a uzavřeno ve stavu vázaném na ATP.[29] V otevřené konformaci víčko nemá žádnou intraproteinovou interakci a při zavření přijde do styku s několika zbytky.[30] Byl zkoumán příspěvek tohoto víčka k aktivitě Hsp90 místně zaměřená mutageneze. Mutant Ala107Asp stabilizující uzavřenou konformaci proteinu tvorbou dalších vodíkových vazeb podstatně zvyšuje aktivitu ATPázy, přičemž ponechává konformaci AMP + PnP beze změny.[30]
The ATPáza vazebná oblast Hsp90 je v současné době intenzivně studována, protože je hlavním vazebným místem léků zaměřených na tento protein.[31] Protinádorová léčiva zaměřená na tuto část Hsp90 zahrnují antibiotika geldanamycin,[11][32] herbimycin, radicicol, deguelin,[33] derrubon,[34] makbecin,[35] a beta-laktamy.[36]
Vazba na bílkoviny
Oblast vázající protein Hsp90 je umístěna směrem k C-konec amino sekvence. Protein Hsp90 může přijmout dva hlavní konformační stavy. První je otevřený stav vázaný na ATP a druhý je uzavřený stav vázaný na ADP. Hydrolýza ATP tedy řídí to, co se běžně označuje jako konformační změna „klešťového typu“ ve vazebném místě pro bílkoviny.[37]
Hsp90, zatímco je v otevřené konformaci, některé ponechává hydrofobní vystavené zbytky, kterým jsou rozloženy a špatně složeny proteiny, které mají vystaveny neobvyklé hydrofobní oblasti, jsou získávány s vysokou afinitou.[38] Když je vázaný substrát na místě, hydrolýza ATP uvolňující energii funkcí ATPázy v blízkosti N-terminální domény nutí konformační změny, které upínají Hsp90 dolů na substrát.[30] Při reakci podobné reakci jiných molekulárních svorkových proteinů, jako je GyrB a MutL, tato stránka řídí prakticky všechny funkce skládání proteinů, ve kterých hraje roli Hsp90. Naproti tomu MutL a GyrB fungují jako topoizomerázy a použijte nabíjecí svorku s vysokým množstvím kladně nabitých postranních řetězců, která je elektrostaticky přitahována k negativnímu páteři DNA.[39]
Schopnost Hsp90 upnout na proteiny mu umožňuje provádět několik funkcí, včetně podpory skládání, prevence agregace a usnadnění transportu.
Funkce
Normální buňky
V nepříznivých buňkách hraje Hsp90 řadu důležitých rolí, mezi které patří asistence skládací, intracelulární transport, údržba a degradace proteinů a také usnadnění buněčné signalizace.
Skládání proteinů a role chaperonu
Je známo, že Hsp90 se asociuje s nepřirozenými strukturami mnoha proteinů, což vedlo k návrhu, že Hsp90 je obecně zapojen do skládání proteinů.[40] Dále bylo prokázáno, že Hsp90 potlačuje agregaci široké škály „klientských“ nebo „substrátových“ proteinů, a proto působí jako obecný ochranný chaperon.[41][42][43] Hsp90 je však o něco selektivnější než jiné chaperony.[44]
Degradace bílkovin
Eukaryotické proteiny, které již nejsou potřeba nebo jsou špatně složeny nebo jinak poškozeny, jsou obvykle označeny k ničení polyubiquitation cesta. Tyto ubikvitinované proteiny jsou 26S rozpoznávány a degradovány proteazom.[45][46] Proteasom 26S je tedy nedílnou součástí mechanismu buňky k degradaci proteinů. Kromě toho je pro udržení HSP90 zapotřebí stálý přísun funkčního Hsp90 terciární struktura proteazomu.[47] Nakonec experimenty provedené s tepelně citlivými mutanty Hsp90 a 26S proteazomem naznačují, že Hsp90 je zodpovědný za většinu, ne-li všechny, aktivity ATPázy proteazomu.[45]
Interakce se steroidními receptory
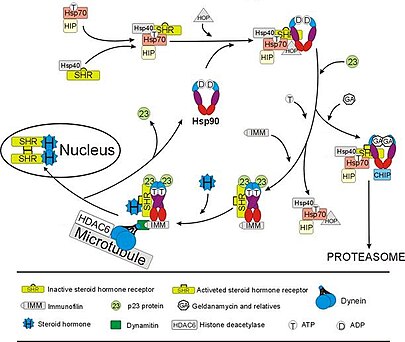
The glukokortikoidový receptor (GR) je nejdůkladněji prostudovaným příkladem a steroidní receptor jehož funkce je rozhodujícím způsobem závislá na interakcích s Hsp90.[50][51] V nepřítomnosti steroidního hormonu kortizol GR bydlí v cytosol komplexovaný s několika chaperonovými proteiny včetně Hsp90 (viz obrázek vpravo). Tyto chaperony udržují GR ve stavu schopném vázat hormon. Druhou rolí Hsp90 je vázat imunofiliny (např., FKBP52 ), které připojují komplex GR k dynein cesta obchodování s bílkovinami, která přemístí aktivovaný receptor z cytoplazmy do jádra.[52] Jakmile se nachází v jádru, GR dimerizuje a váže se na specifické sekvence DNA a tím upregulates exprese genů reagujících na GR. Hsp90 je také vyžadován pro správné fungování několika dalších steroidních receptorů, včetně těch, které jsou odpovědné za vazbu aldosteron,[53] androgen,[54] estrogen,[55] a progesteron.[56]
Rakovinné buňky
Rakovinné buňky nadměrně exprimují řadu proteinů, včetně receptorů růstových faktorů, jako je EGFR,[57] nebo proteiny signální transdukce, jako je PI3K a AKT (Může dojít ke spuštění inhibice těchto proteinů apoptóza ). Hsp90 stabilizuje receptory různých růstových faktorů[58] a některé signální molekuly včetně PI3K a AKT proteinů. Proto inhibice Hsp90 downregulates the PI3K / AKT cesta vedoucí k downregulaci anti-apoptotický protein Bcl-w vedoucí k apoptóze rakovinných a stárnoucí buňky.[59][15][60]
Další důležitou úlohou Hsp90 při rakovině je stabilizace mutantních proteinů, jako jsou v-Src, fúze onkogen Bcr / Abl a mutantní formy p53 které se objevují během transformace buněk. Ukazuje se, že Hsp90 může působit jako „ochránce“ méně stabilních proteinů produkovaných mutacemi DNA.[61]
Hsp90 je také nutný pro indukci vaskulárního endoteliálního růstového faktoru (VEGF ) a syntáza oxidu dusnatého (NOS).[24] Oba jsou důležité pro de novo angiogeneze který je nutný pro růst nádoru nad hranici difúzní vzdálenosti kyslíku v tkáních.[61] Rovněž podporuje invazní krok metastáza pomocí matricová metaloproteináza MMP2.[62] Spolu se svými spolukaperony moduluje Hsp90 apoptózu nádorových buněk zprostředkovanou účinky na AKT,[23] receptory faktoru nekrózy nádorů (TNFR) a jaderný faktor-κB (NF-κB) funkce. "[63] Hsp90 se také účastní mnoha klíčových procesů v onkogenezi, jako je soběstačnost růstových signálů, stabilizace mutantních proteinů, angiogeneze a metastázy.
Klinický význam
Hsp90 hraje v buňce zjevně protichůdné role, protože je nezbytný jak pro tvorbu a údržbu, tak pro ničení proteinů. Jeho normální funkce je zásadní pro udržení zdraví buněk, zatímco jeho dysregulace může přispívat karcinogeneze. Schopnost tohoto chaperona jak stabilizovat 26S proteazom (což umožňuje buňce degradovat nechtěné a / nebo škodlivé proteiny) a stabilizovat kinázy proti stejnému proteazomu demonstruje jeho funkční rozmanitost. Použití Inhibitory Hsp90 v léčbě rakoviny zdůrazňují význam Hsp90 jako terapeutického cíle.[64]
Cílení na Hsp90 pomocí léků ukázalo v klinických studiích slibné účinky. Například inhibitor Hsp90 geldanamycin byl použit jako protinádorové činidlo.[11] Původně se předpokládalo, že droga funguje jako a kináza inhibitor, ale následně se ukázalo, že je inhibitorem Hsp90, kde používá kompaktní konformaci k zavedení do vazebného místa ATP.[11]
HSP90 beta byl identifikován jako jeden z autoantigenních biomarkerů a cílů zapojených do autoimunitního onemocnění vaječníků u člověka, které vede k selhání vaječníků a tím k neplodnosti.[65]
Predikce a validace imunodominantního epitopu / s HSP90 beta proteinu byla prokázána pomocí sér od neplodných žen, které mají anti-HSP90 autoprotilátky. Dekapeptid EP6 (380-389) je hlavním imunogenním epitopem HSP90 následovaným EP1 (1-12) a EP8 (488-498). Znalost vázání epitopů na autoantigenu je nezbytná pro pochopení následných patologických událostí. Předpokládané 3D struktury těchto peptidů prokázaly, že existují ve smyčkové konformaci, která je nejpohyblivější částí proteinu. Také analýza sekvencí HSP90 beta u několika druhů odhalila, že peptid EP6 tvoří součást dobře konzervovaného motivu. Polyklonální protilátka vytvořená proti imunodominantnímu epitopu EP6 potvrzuje podobnou biochemickou a buněčnou imunoreaktivitu, jaká je pozorována u sér pacientů s autoprotilátkami anti-HSP90. Studie by mohla vygenerovat nové nástroje pro detekci epitopů vyvolávajících choroby a možnou terapeutickou intervenci.[66]
Vývoj
Sekvenční seřazení Hsp90 ukázalo, že protein má přibližně 40% sekvenční identitu napříč všemi homology, což naznačuje, že se jedná o vysoce konzervativní protein. V souboru jsou dva homology cytosol a endoplazmatické retikulum resp. Přítomnost těchto dvou homologů byla pravděpodobně způsobena a genová duplikace událost velmi brzy ve vývoji eukaryoty které mohly doprovázet vývoj endoplazmatického retikula nebo jádro. Tento závěr je podporován skutečností, že duplikace se nachází v Giardia lamblia, jeden z prvních rozvětvených eukaryotických druhů. Vyskytly se nejméně 2 další duplikace genů, což vysvětluje různé formy Hsp90 nalezené v houby a obratlovců. Jedna divergence produkovala příbuzné a teplem indukované formy Hsp90 v Saccharomyces cerevisiae zatímco druhá duplikace genů v cytosolické větvi produkovala podrodiny alfa a beta sekvencí, které se nacházejí u všech obratlovců. V fylogenetický strom na základě sekvencí Hsp90 bylo zjištěno, že rostliny a zvířata jsou si navzájem bližší než s houbami.[67] Podobně jako protein Hsp90, gen pro protein Hsp70 také prošel duplikací ve velmi rané fázi tvorby eukaryotických buněk a z této duplikace genu byly výsledkem homologů v cytosolu a endoplazmatickém retikulu.[68] Tyto genové duplikace jsou důležité z hlediska původu eukaryotické buňky a endoplazmatického retikula.[69][70]
Viz také
Reference
- ^ A b C PDB: 2CG9; Ali MM, Roe SM, Vaughan CK, Meyer P, Panaretou B, Piper PW, Prodromou C, Pearl LH (duben 2006). „Krystalová struktura uzavřeného chaperonového komplexu Hsp90-nukleotid-p23 / Sba1“. Příroda. 440 (7087): 1013–7. doi:10.1038 / nature04716. PMC 5703407. PMID 16625188.
- ^ Prodromou C, Roe SM, Piper PW, Pearl LH (červen 1997). „Molekulární svorka v krystalové struktuře N-koncové domény kvasinkového chaperonu Hsp90“. Nat. Struct. Biol. 4 (6): 477–82. doi:10.1038 / nsb0697-477. PMID 9187656.
- ^ Csermely P, Schnaider T, Soti C, Prohászka Z, Nardai G (srpen 1998). „Rodina molekulárních chaperonů 90 kDa: struktura, funkce a klinické aplikace. Komplexní přehled“. Pharmacol. Ther. 79 (2): 129–68. doi:10.1016 / S0163-7258 (98) 00013-8. PMID 9749880.
- ^ Crevel G, Bates H, Huikeshoven H, Cotterill S (1. června 2001). „Protein Drosophila Dpit47 je jaderný kochaperon Hsp90, který interaguje s DNA polymerázou alfa“. J. Cell Sci. 114 (Pt 11): 2015–25. PMID 11493638.
- ^ A b C d Chen B, Zhong D, Monteiro A (2006). „Srovnávací genomika a vývoj genů rodiny HSP90 ve všech královstvích organismů“. BMC Genomics. 7: 156. doi:10.1186/1471-2164-7-156. PMC 1525184. PMID 16780600.
- ^ Thomas JG, Baneyx F (říjen 1998). „Role Escherichia coli Malé proteiny tepelného šoku IbpA a IbpB při zvládání tepelného stresu: Porovnání s ClpA, ClpB a HtpG In Vivo“. J. Bacteriol. 180 (19): 5165–72. doi:10.1128 / JB.180.19.5165-5172.1998. PMC 107554. PMID 9748451.
- ^ Prodromou C, Panaretou B, Chohan S, Siligardi G, O'Brien R, Ladbury JE, Roe SM, Piper PW, Pearl LH (srpen 2000). „ATPázový cyklus Hsp90 pohání molekulární„ svorku “přechodnou dimerizací N-koncových domén“. EMBO J.. 19 (16): 4383–92. doi:10.1093 / emboj / 19.16.4383. PMC 302038. PMID 10944121.
- ^ A b C Chen B, Piel WH, Gui L, Bruford E, Monteiro A (prosinec 2005). „Rodina genů HSP90 v lidském genomu: pohledy na jejich divergenci a vývoj“. Genomika. 86 (6): 627–37. doi:10.1016 / j.ygeno.2005.08.012. PMID 16269234.
- ^ Grammatikakis N, Vultur A, Ramana CV, Siganou A, Schweinfest CW, Watson DK, Raptis L (březen 2002). „Role Hsp90N, nového člena rodiny Hsp90, v signální transdukci a neoplastické transformaci“. J. Biol. Chem. 277 (10): 8312–20. doi:10,1074 / jbc.M109200200. PMID 11751906.
- ^ Zurawska A, Urbanski J, Bieganowski P (listopad 2008). „Hsp90n - náhodný produkt náhodné chromozomální translokace místo běžného člena rodiny Hsp90 lidského proteomu“. Biochimica et Biophysica Acta (BBA) - bílkoviny a proteomika. 1784 (11): 1844–6. doi:10.1016 / j.bbapap.2008.06.013. PMID 18638579.
- ^ A b C d E Goetz MP, Toft DO, Ames MM, Erlichman C (srpen 2003). „Komplex chaperonů Hsp90 jako nový cíl pro léčbu rakoviny“. Ann. Oncol. 14 (8): 1169–76. doi:10.1093 / annonc / mdg316. PMID 12881371.
- ^ Pearl LH, Prodromou C (únor 2000). "Struktura a funkce in vivo Hsp90". Curr. Opin. Struct. Biol. 10 (1): 46–51. doi:10.1016 / S0959-440X (99) 00047-0. PMID 10679459.
- ^ A b C Prodromou C, Pearl LH (říjen 2003). "Struktura a funkční vztahy Hsp90". Curr Cancer Drug Targets. 3 (5): 301–23. doi:10.2174/1568009033481877. PMID 14529383.
- ^ Pearl LH, Prodromou C (2001). Struktura, funkce a mechanismus molekulárního chaperonu Hsp90. Adv. Protein Chem. Pokroky v chemii proteinů. 59. str. 157–86. doi:10.1016 / S0065-3233 (01) 59005-1. ISBN 978-0-12-034259-4. PMID 11868271.
- ^ A b C d Stebbins CE, Russo AA, Schneider C, Rosen N, Hartl FU, Pavletich NP (duben 1997). "Krystalová struktura komplexu Hsp90-geldanamycin: cílení na chaperon proteinu protinádorovým činidlem". Buňka. 89 (2): 239–50. doi:10.1016 / S0092-8674 (00) 80203-2. PMID 9108479.
- ^ A b C Prodromou C, Roe SM, O'Brien R, Ladbury JE, Piper PW, Pearl LH (červenec 1997). "Identifikace a strukturní charakterizace vazebného místa ATP / ADP v molekulárním chaperonu Hsp90". Buňka. 90 (1): 65–75. doi:10.1016 / S0092-8674 (00) 80314-1. PMID 9230303.
- ^ Prodromou C, Roe SM, Piper PW, Pearl LH (červen 1997). „Molekulární svorka v krystalové struktuře N-koncové domény kvasinkového chaperonu Hsp90“. Nat. Struct. Biol. 4 (6): 477–82. doi:10.1038 / nsb0697-477. PMID 9187656.
- ^ A b C Meyer P, Prodromou C, Hu B, Vaughan C, Roe SM, Panaretou B, Piper PW, Pearl LH (březen 2003). „Strukturální a funkční analýza středního segmentu hsp90: důsledky pro hydrolýzu ATP a interakce klientských proteinů a kochaperonů“ (PDF). Mol. Buňka. 11 (3): 647–58. doi:10.1016 / S1097-2765 (03) 00065-0. PMID 12667448.
- ^ Shiau AK, Harris SF, Southworth DR, Agard DA (říjen 2006). „Strukturální analýza E. coli hsp90 odhaluje dramatické konformační přesmyky závislé na nukleotidech“. Buňka. 127 (2): 329–40. doi:10.1016 / j.cell.2006.09.027. PMID 17055434.
- ^ Ali MM, Roe SM, Vaughan CK, Meyer P, Panaretou B, Piper PW, Prodromou C, Pearl LH (duben 2006). „Krystalová struktura uzavřeného chaperonového komplexu Hsp90-nukleotid-p23 / Sba1“. Příroda. 440 (7087): 1013–7. doi:10.1038 / nature04716. PMC 5703407. PMID 16625188.
- ^ Dollins DE, Warren JJ, Immormino RM, Gewirth DT (říjen 2007). „Struktury GRP94-nukleotidových komplexů odhalují mechanické rozdíly mezi chaperony hsp90“. Mol. Buňka. 28 (1): 41–56. doi:10.1016 / j.molcel.2007.08.024. PMC 2094010. PMID 17936703.
- ^ Wanderer SK, Richter K, Buchner J. (Červenec 2008). „Strojní zařízení hsp90. J. Biol. Chem. 283 (27): 18473–7. doi:10,1074 / jbc.R800007200. PMID 18442971.
- ^ A b Sato S, Fujita N, Tsuruo T (září 2000). "Modulace aktivity Akt kinázy vazbou na Hsp90". Proc. Natl. Acad. Sci. USA. 97 (20): 10832–7. doi:10.1073 / pnas.170276797. PMC 27109. PMID 10995457.
- ^ A b Fontana J, Fulton D, Chen Y, Fairchild TA, McCabe TJ, Fujita N, Tsuruo T, Sessa WC (květen 2002). „Studie mapování domén ukazují, že M doména hsp90 slouží jako molekulární lešení k regulaci Akt-dependentní fosforylace endotelové syntázy oxidu dusnatého a uvolňování NO“. Circ. Res. 90 (8): 866–73. doi:10.1161 / 01.RES.0000016837.26733 BE. PMID 11988487.
- ^ Panaretou B, Siligardi G, Meyer P, Maloney A, Sullivan JK, Singh S, Millson SH, Clarke PA, Naaby-Hansen S, Stein R, Cramer R, Mollapour M Workman P, Piper PW, Pearl LH, Prodromou C (prosinec 2002). "Aktivace aktivity ATPázy hsp90 stresově regulovaným kochaperonem aha1" (PDF). Mol. Buňka. 10 (6): 1307–18. doi:10.1016 / S1097-2765 (02) 00785-2. PMID 12504007.
- ^ Marcu MG, Chadli A, Bouhouche I, Catelli M, Neckers LM (listopad 2000). "Antagonista proteinu 90 tepelného šoku novobiocin interaguje s dříve nerozpoznanou doménou vázající ATP v karboxylovém konci chaperonu". J. Biol. Chem. 275 (47): 37181–6. doi:10,1074 / jbc.M003701200. PMID 10945979.
- ^ Söti C, Rácz A, Csermely P (březen 2002). „Nukleotidově závislý molekulární přepínač řídí navázání ATP na C-koncovou doménu Hsp90. N-koncová vazba nukleotidů odmaskuje C-koncovou vazebnou kapsu“. J. Biol. Chem. 277 (9): 7066–75. doi:10,1074 / jbc.M105568200. PMID 11751878.
- ^ Young JC, Obermann WM, Hartl FU (červenec 1998). "Specifická vazba tetratrikopeptidových opakujících se proteinů na C-koncovou 12-kDa doménu hsp90". J. Biol. Chem. 273 (29): 18007–10. doi:10.1074 / jbc.273.29.18007. PMID 9660753.
- ^ Didenko T, Duarte AM, Karagöz GE, Rüdiger SG (březen 2012). "Struktura a funkce Hsp90 studované NMR spektroskopií". Biochim. Biophys. Acta. 1823 (3): 636–47. doi:10.1016 / j.bbamcr.2011.11.009. PMID 22155720.
- ^ A b C Wegele H, Müller L, Buchner J (2004). Hsp70 a Hsp90 - přenosový tým pro skládání proteinů. Physiol. Biochem. Pharmacol. Recenze fyziologie, biochemie a farmakologie. 151. s. 1–44. doi:10.1007 / s10254-003-0021-1. ISBN 978-3-540-22096-1. PMID 14740253.
- ^ Chiosis G, Caldas Lopes E, Solit D (červen 2006). „Inhibitory proteinu 90 tepelného šoku: kronika od geldanamycinu po dnešní látky“. Curr Opin Investig Drugs. 7 (6): 534–41. PMID 16784024.
- ^ Pratt WB, Toft DO (1. února 2003). „Regulace funkce signálního proteinu a obchodování s ním pomocí chaperonového stroje na bázi hsp90 / hsp70“. Exp. Biol. Med. (Maywood). 228 (2): 111–33. CiteSeerX 10.1.1.334.341. doi:10.1177/153537020322800201. PMID 12563018.
- ^ Oh SH, Woo JK, Yazici YD, Myers JN, Kim WY, Jin Q, Hong SS, Park HJ, Suh YG, Kim KW, Hong WK, Lee HY (červen 2007). „Strukturální základ pro vyčerpání proteinu tepelného šoku 90 klientských proteinů deguelinem“. J. Natl. Cancer Inst. 99 (12): 949–61. doi:10.1093 / jnci / djm007. PMID 17565155.
- ^ Hadden MK, Galam L, Gestwicki JE, Matts RL, Blagg BS (prosinec 2007). "Derrubon, inhibitor stroje na skládání proteinů Hsp90". J. Nat. Prod. 70 (12): 2014–8. doi:10.1021 / np070190s. PMID 18020309.
- ^ Martin CJ, Gaisser S, Challis IR, Carletti I, Wilkinson B, Gregory M, Prodromou C, Roe SM, Pearl LH, Boyd SM, Zhang MQ (květen 2008). "Molekulární charakterizace makbecinu jako inhibitoru Hsp90". J. Med. Chem. 51 (9): 2853–7. doi:10.1021 / jm701558c. PMID 18357975.
- ^ O'Boyle NM, Knox AJ, Price TT, Williams DC, Zisterer DM, Lloyd DG, Meegan MJ (říjen 2011). "Identifikace olova β-laktamu a příbuzných iminových inhibitorů proteinu tepelného šoku molekulárního chaperonu 90". Bioorg. Med. Chem. 19 (20): 6055–68. doi:10.1016 / j.bmc.2011.08.048. PMID 21920765.
- ^ Grenert JP, Sullivan WP, Fadden P, Haystead TA, Clark J, Mimnaugh E, Krutzsch H, Ochel HJ, Schulte TW, Sausville E, Neckers LM, Toft DO (září 1997). „Amino-koncová doména proteinu 90 tepelného šoku (hsp90), který váže geldanamycin, je přepínací doména ATP / ADP, která reguluje konformaci hsp90“. J. Biol. Chem. 272 (38): 23843–50. doi:10.1074 / jbc.272.38.23843. PMID 9295332.
- ^ Xu Z, Horwich AL, Sigler PB (srpen 1997). „Krystalová struktura asymetrického chaperoninového komplexu GroEL-GroES- (ADP) 7“. Příroda. 388 (6644): 741–50. doi:10.1038/41944. PMID 9285585.
- ^ Kampranis SC, Bates AD, Maxwell A (červenec 1999). „Model mechanismu průchodu řetězců DNA gyrázou“. Proc. Natl. Acad. Sci. USA. 96 (15): 8414–9. doi:10.1073 / pnas.96.15.8414. PMC 17530. PMID 10411889.
- ^ Buchner J. (Duben 1999). "Hsp90 & Co. - hospodářství pro skládání". Trends Biochem. Sci. 24 (4): 136–41. doi:10.1016 / S0968-0004 (99) 01373-0. PMID 10322418.
- ^ Miyata Y, Yahara I (duben 1992). „Protein 90-kDa tepelného šoku, Hsp90, váže a chrání kasein kinázu II před vlastní agregací a zvyšuje její kinázovou aktivitu“. J. Biol. Chem. 267 (10): 7042–7. PMID 1551911.
- ^ Wiech H, Buchner J., Zimmermann R, Jakob U (červenec 1992). "Hsp90 chaperonové skládání proteinu in vitro". Příroda. 358 (6382): 169–70. doi:10.1038 / 358169a0. PMID 1614549.
- ^ Jakob U, Lilie H, Meyer I, Buchner J (březen 1995). "Přechodná interakce Hsp90 s časnými rozvíjejícími se meziprodukty citrát syntázy. Důsledky pro tepelný šok in vivo". J. Biol. Chem. 270 (13): 7288–94. doi:10.1074 / jbc.270.13.7288. PMID 7706269.
- ^ Picard D (říjen 2002). „Protein 90 tepelného šoku, chaperon pro skládání a regulaci“. Buňka. Mol. Life Sci. 59 (10): 1640–8. doi:10.1007 / PL00012491. PMID 12475174.
- ^ A b Imai J, Maruya M, Yashiroda H, Yahara I, Tanaka K (červenec 2003). „Molekulární chaperon Hsp90 hraje roli při montáži a údržbě proteazomu 26S“. EMBO J.. 22 (14): 3557–67. doi:10.1093 / emboj / cdg349. PMC 165619. PMID 12853471.
- ^ Correia MA, Sadeghi S, Mundo-Paredes E (2005). „Ubikvitinace cytochromu P450: značka pro proteolytickou porážku?“. Annu. Pharmacol. Toxicol. 45: 439–64. doi:10.1146 / annurev.pharmtox.45.120403.100127. PMID 15822184.
- ^ Kimura Y, Matsumoto S, Yahara I (březen 1994). "Teplotně citlivé mutanty hsp82 začínajících kvasinek Saccharomyces cerevisiae". Mol. Gen. Genet. 242 (5): 517–27. doi:10.1007 / BF00285275. PMID 8121410.
- ^ Davies TH, Ning YM, Sánchez ER (únor 2002). „Nový první krok v aktivaci steroidních receptorů: hormonálně indukovaná změna imunofilinů FKBP51 a FKBP52“. J. Biol. Chem. 277 (7): 4597–600. doi:10,1074 / jbc.C100531200. PMID 11751894.
- ^ Pałyga J, Kozłowski Ł (2007). "Struktura a funkce molekulárního chaperonu HSP90". Sowriemiennyj Naucznyj Wiestnik Ser. Biologija Chimija. 15 (23): 46–65.
- ^ Pratt WB, Morishima Y, Murphy M, Harrell M (2006). Spolupráce glukokortikoidových receptorů. Handb Exp Pharmacol. Příručka experimentální farmakologie. 172. 111–38. doi:10.1007/3-540-29717-0_5. ISBN 978-3-540-25875-9. PMID 16610357.
- ^ Grad I, Picard D (září 2007). "Glukokortikoidové odpovědi jsou formovány molekulárními chaperony". Mol. Buňka. Endokrinol. 275 (1–2): 2–12. doi:10.1016 / j.mce.2007.05.018. PMID 17628337.
- ^ Pratt WB, MD Galigniana, Morishima Y, Murphy PJ (2004). "Role molekulárních chaperonů při působení steroidních receptorů". Eseje Biochem. 40: 41–58. doi:10.1042 / bse0400041. PMID 15242338.
- ^ Rafestin-Oblin ME, Couette B, Radanyi C, Lombes M, Baulieu EE (Červen 1989). "Mineralokortikosteroidový receptor kuřecího střeva. Oligomerní struktura a transformace". J. Biol. Chem. 264 (16): 9304–9. PMID 2542305.
- ^ Joab I, Radanyi C, Renoir M, Buchou T, Catelli MG, Binart N, Mester J, Baulieu EE (1984). "Běžná nehormonální vazebná složka v netransformovaných kuřecích vejcovodech receptory čtyř steroidních hormonů". Příroda. 308 (5962): 850–3. doi:10.1038 / 308850a0. PMID 6201744.
- ^ Redeuilh G, Moncharmont B, Secco C, Baulieu EE (květen 1987). „Složení podjednotky molybdenanem stabilizovaného„ 8-9 S “netransformovaného estradiolového receptoru purifikovaného z lýtkové dělohy“. J. Biol. Chem. 262 (15): 6969–75. PMID 3584104.
- ^ Catelli MG, Binart N, Jung-Testas I, Renoir JM, Baulieu EE, Feramisco JR, Welch WJ (prosinec 1985). „Společnou 90kd proteinovou složkou netransformovaných steroidních receptorů„ 8S “je protein tepelného šoku.“. EMBO J.. 4 (12): 3131–5. doi:10.1002 / j.1460-2075.1985.tb04055.x. PMC 554632. PMID 2419124.
- ^ Lurje G, Lenz HJ (2009). "Signalizace EGFR a objev drog". Onkologie. 77 (6): 400–410. doi:10.1159/000279388. PMID 20130423.
- ^ Sawai A, Chandarlapaty S, Greulich H, Gonen M, Ye Q, Arteaga CL, Sellers W, Rosen N, Solit DB (leden 2008). „Inhibice Hsp90 down-reguluje mutantní expresi receptoru epidermálního růstového faktoru (EGFR) a senzibilizuje EGFR mutantní tumory na paclitaxel“. Cancer Res. 68 (2): 589–96. doi:10.1158 / 0008-5472.CAN-07-1570. PMC 4011195. PMID 18199556.
- ^ Paez-Ribes M, González-Gualda E, Doherty GJ, Muñoz-Espín D (2019). „Cílení na senescentní buňky v translační medicíně“. EMBO Molekulární medicína. 11 (12): e10234. doi:10.15252 / emmm.201810234. PMC 6895604. PMID 31746100.
- ^ Mohsin SK, Weiss HL, Gutierrez MC, Chamness GC, Schiff R, Digiovanna MP, Wang CX, Hilsenbeck SG, Osborne CK, Allred DC, Elledge R, Chang JC (duben 2005). „Neoadjuvantní trastuzumab indukuje apoptózu u primárních nádorů prsu“. J. Clin. Oncol. 23 (11): 2460–8. doi:10.1200 / JCO.2005.00.661. PMID 15710948.
- ^ A b Calderwood SK, Khaleque MA, Sawyer DB, Ciocca DR (březen 2006). "Proteiny tepelného šoku v rakovině: chaperony tumorigeneze". Trends Biochem. Sci. 31 (3): 164–72. doi:10.1016 / j.tibs.2006.01.006. PMID 16483782.
- ^ Eustace BK, Sakurai T, Stewart JK, Yimlamai D, Unger C, Zehetmeier C, Lain B, Torella C, Henning SW, Beste G, Scroggins BT, Neckers L, Ilag LL, Jay DG (červen 2004). „Funkční proteomické testy odhalují zásadní extracelulární roli hsp90 alfa při invazivitě rakovinných buněk“. Nat. Cell Biol. 6 (6): 507–14. doi:10.1038 / ncb1131. PMID 15146192.
- ^ Whitesell L, Lindquist SL (říjen 2005). "Hsp90 a doprovod rakoviny". Nat. Rev. Rakovina. 5 (10): 761–72. doi:10.1038 / nrc1716. PMID 16175177.
- ^ Kim YS, Alarcon SV, Lee S, Lee MJ, Giaccone G, Neckers L, Trepel JB (2009). „Aktualizace inhibitorů Hsp90 v klinické studii“. Curr Top Med Chem. 9 (15): 1479–92. doi:10.2174/156802609789895728. PMC 7241864. PMID 19860730.
- ^ Pires ES, Khole VV (2009). „Blok na cestě k plodnosti: autoprotilátky proti proteinu 90-beta tepelnému šoku v autoimunitě lidských vaječníků“. Plod Steril. 92 (4): 1395–1409. doi:10.1016 / j.fertnstert.2008.08.068. PMID 19022436.
- ^ Pires ES, Choudhury AK, Idicula-Thomas S, Khole VV (2011). "Anti-HSP90 autoantibodies in sera of infertile women identify a dominant, conserved epitope EP6 (380-389) of HSP90 beta protein". Reprod Biol Endocrinol. 9 (16): 13. doi:10.1186/1477-7827-9-16. PMC 3039567. PMID 21272367.
- ^ Gupta RS (November 1995). "Phylogenetic analysis of the 90 kD heat shock family of protein sequences and an examination of the relationship among animals, plants, and fungi species" (PDF). Mol. Biol. Evol. 12 (6): 1063–73. doi:10.1093/oxfordjournals.molbev.a040281. PMID 8524040.
- ^ Gupta RS, Aitken K, Falah M, Singh B (April 1994). "Cloning of Giardia lamblia heat shock protein HSP70 homologs: implications regarding origin of eukaryotic cells and of endoplasmic reticulum". Proc. Natl. Acad. Sci. USA. 91 (8): 2895–9. doi:10.1073/pnas.91.8.2895. PMC 43480. PMID 8159675.
- ^ Gupta RS, Golding GB (May 1996). "The origin of the eukaryotic cell". Trends Biochem. Sci. 21 (5): 166–71. doi:10.1016/S0968-0004(96)20013-1. PMID 8871398.
- ^ Gupta RS (December 1998). "Protein phylogenies and signature sequences: A reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes". Microbiol. Mol. Biol. Rev. 62 (4): 1435–91. doi:10.1128/MMBR.62.4.1435-1491.1998. PMC 98952. PMID 9841678.
externí odkazy
- Hsp90+Heat-Shock+Proteins v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- Didier Picard. "Hsp90 web site". VisibilityWeb. Archivovány od originál on 2004-11-23. Citováno 2008-07-07.
A web site for the community of scientists interested in the Hsp90 molecular chaperone machine.