Histidinkináza - Histidine kinase
| protein histidinkináza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
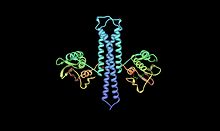 Krystalografické struktura ATP: protein-L-histidin N-fosfotransferáza na základě PDB: 2c2aSouřadnice. | |||||||||
| Identifikátory | |||||||||
| EC číslo | 2.7.13.3 | ||||||||
| Číslo CAS | 99283-67-7 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Histidinkinázy (HK) jsou multifunkční a typicky v královstvích jiných než zvířat transmembránový, proteiny z transferáza třída enzymy které hrají roli v signální transdukce přes buněčnou membránu.[1] Drtivá většina HK jsou homodimery, které vykazují autokináza, fosfotransfer a aktivita fosfatázy. HK mohou fungovat jako buněčné receptory pro signalizaci molekul analogickým způsobem receptory tyrosinkinázy (RTK). Multifunkční receptorové molekuly, jako jsou HK a RTK, mají obvykle části na vnější straně buňky (extracelulární doména), které se váží na molekuly podobné hormonům nebo růstovým faktorům, částem, které překlenují buněčnou membránu (transmembránová doména ) a části v buňce (intracelulární doména), které obsahují enzymatickou aktivitu. Navíc kináza Aktivita má intracelulární domény obvykle oblasti, které se vážou na sekundární efektorovou molekulu nebo komplex molekul, které dále šíří signální transdukci v buňce. Na rozdíl od jiných tříd proteinové kinázy, HK jsou obvykle součástí a dvousložkové mechanismy přenosu signálu ve kterém HK přenáší fosfátovou skupinu ATP na histidinový zbytek v kináze a poté na aspartátový zbytek v přijímací doméně a regulátor odezvy protein (nebo někdy na samotné kináze). V nedávné době byla v lidských buňkách rozpoznána rozšířená existence fosforylace proteinů histidinu odlišná od dvoukomponentních histidinkináz.[2][3] Na rozdíl od fosforylace Ser, Thr a Tyr je analýza fosforylovaného histidinu pomocí standardních biochemických a hmotnostních spektrometrických přístupů mnohem náročnější,[4][5] a pro jejich konzervaci jsou vyžadovány speciální postupy a separační techniky spolu s klasickou fosforylací Ser, Thr a Tyr na proteinech izolovaných z lidských buněk.[6]
Ve smyslu enzymologie, histidinkináza (ES 2.7.13.3, EnvZ, histidinkin kináza, protein histidinkináza, protein kináza (histidin), HK1, HP165, Sln1p) je enzym že katalyzuje the chemická reakce
- ATP + protein L-histidin ADP + protein N-fosfo-L-histidin.

Tedy dva substráty tohoto enzymu jsou ATP a protein L-histidin, zatímco jeho dva produkty jsou ADP a protein N-fosfo-L-histidin.
Tento typ enzymu se podílí na signálních transdukčních drahách před mnoha buněčnými procesy, včetně různých metabolických, virulenčních a homeostatických drah.
Mechanismus

Mechanismus reakcí katalyzovaných histidinkinázou nebyl zcela objasněn, ale současné důkazy naznačují, že katalytická doména jednoho dimerní jednotka se může otáčet takovým způsobem, že ATP vazebná kapsa této jednotky může přijít do kontaktu s konkrétním histidinovým zbytkem na opačné jednotce a nukleofilní adice vede k fosforylovanému histidinu.[7]
Struktura a funkce
HK se skládá z několika domén počínaje krátkým N-terminál cytoplazmatická část připojená k extracelulární snímací doméně přes transmembránu α šroubovice. Druhá transmembránová α spirála spojuje extracelulární doménu s C-terminál cytoplazmatická katalytická doména. Je známo, že HK slouží rolím v mnoha různých drahách signální transdukce, takže není divu, že extracelulární detekční doména není v rodině HK velmi dobře zachována. Naproti tomu cytoplazmatická doména má tendenci mít vysokou sekvenci homologie a obsahuje několik známých motivy. Mezi tyto motivy patří pole H, N, G1, F a G2.[8] Autofosforylační H-box je obsažen v N-terminální dimerizační a histidinové fosfotransferové (DHp) doméně. V HK853-CD krystalizovalo z Thermotoga maritima, tato doména je spirálovitá-sponka do vlasů a je tvořen zbytky 232-317. Místo histidinové fosforylace je umístěno na His-260. Krabice N, G1, F a G2 jsou obsaženy v C-koncové katalytické doméně a doméně vázající ATP (CA). Tato doména je tvořena zbytky 323-489 a tvoří strukturu známou jako sendvičový záhyb α / β. Tento konkrétní záhyb má jednu vrstvu složenou z 5 pramenů β list a druhá vrstva je tvořena třemi α šroubovicemi.
Dimerická jednotka je držena pohromadě čtyřšroubovicovým svazkem, vytvořeným při interakci C-koncových segmentů spirál α1 na každé podjednotce v antiparalelní způsobem s oběma helixy α2. Stabilitě dimeru napomáhá několik interakcí na rozhraní mezi DHps každého monomeru. Patří mezi ně hydrofobní interakce mezi konzervovanými hydrofobní zbytky stejně jako dva Vodíkové vazby (Thr-252...Glu-316 'a Arg-263...Asn-307 “) a jedna solný most (Lys-270...Glu-303 “). Další interakce jsou zprostředkovány vodíkovými vazbami na vodu v dutině uvnitř svinuté cívky a lemovány hydrofobními zbytky.


The nukleotid /ATP vazebná kapsa je obsažena v doméně CA a strukturní podobnost této kapsy je vysoká mezi většinou HK. Dutina CheA, také krystalizovaná z T. maritima, je nejprve tvořena β vrstvou P4 v zadní části a boky dutiny jsou tvořeny 4 motivy zmíněnými výše, krabicemi N, G1, F a G2.[9] Většina zbytků pocházejících z p listu je hydrofobní, s výjimkou Asp449. Tento zbytek je neměnný a tvoří vodíkovou vazbu s molekulou vody k adenin aminová skupina. Tři další molekuly vody tvoří přímé vodíkové vazby s adeninovou bází. A Mg2+ iont tvoří můstek mezi všemi třemi fosfáty a neměnným Asnovým zbytkem. Nakonec další dvě molekuly vody dokončují oktaedrickou koordinaci s Mg2+ a jsou spojeny s Arg-408 a His-405. Když je γ fosfát ATP destabilizován, Mg2+ již není pozorován kvůli jeho neschopnosti oktaedricky koordinovat. Marina a kol. tvrdí, že podobná koordinace Mg2+ se vyskytuje v HK853, ale že je nepozorován kvůli použití ATP analogový AMPPNP v krystalové struktuře.[7] Během krystalizace byl analog hydrolyzován na produkt podobný ADP.
Poslední strana vázací kapsy ATP se pohodlně jmenuje „víko ATP“. Stabilita této struktury je zprostředkována přítomností y fosfátu a tím i Mg2+ ion ve vazebném místě. Ukázalo se také, že přítomnost nukleotidové báze hraje významnou roli při stabilizaci víka v uzavřeném stavu konformace. Víčko ATP je připojeno přes hydrofobní zbytky ke zbytku proteinu. Γ fosfát ATP je poněkud exponovaný, což umožňuje defosforylace Po navázání ATP v této kapse se předpokládá, že dojde ke konformační změně, která umožní rotaci CA domény přijít do kontaktu s DHp jiného monomeru, a tím umožní konzervovanému His-260 odpočívat v blízkosti y fosfátu. Nε His-260 pak napadá y fosfát ATP v a nukleofilní adice a narazí ADP jako jeho odstupující skupina.
Úloha plísňových infekcí
A dvousložkový systém zahrnující histidinkinázu a proměnnou regulátor odezvy protein může být rozhodující pro virulenci některých kmenů hub, jako je Candida albicans, který je často zodpovědný za způsobení kandidóza v imunokompromitovaný osob.[10] C. albicans s delecí CHK1, dvousložkového genu histidinkinázy, vykazuje defekty morfogeneze a drastické snížení schopnosti buňky odolat eliminaci člověkem neutrofily. Protože lidem chybí tento dvousložkový systém, může to být dobrý cíl antimikrobiální agenti za účelem léčby kandidóza.
Reference
- ^ Wolanin PW, Thomason PA, Stock JB (2002). „Histidinové proteinové kinázy: klíčové převodníky signálu mimo zvířecí říši“. Genome Biology. 3 (10): reviews3013.1–3013.8. doi:10.1186 / gb-2002-3-10-reviews3013. PMC 244915. PMID 12372152.
- ^ Fuhs SR, Hunter T (2017). „pHisforylace: vznik histidinové fosforylace jako reverzibilní regulační modifikace“. Curr Opin Cell Biol. 45: 8–16. doi:10.1016 / j.ceb.2016.12.010. PMC 5482761. PMID 28129587.
- ^ Fuhs SR, Meisenhelder J, Aslanian A, Ma L, Zagorska A, Stankova M, Binnie A, Al-Obeidi F, Mauger J, Lemke G, Yates JR 3rd, Hunter T (2015). „Monoklonální 1- a 3-fosfohistidinové protilátky: nové nástroje ke studiu histidinové fosforylace“. Buňka. 162 (1): 198–210. doi:10.1016 / j.cell.2015.05.046. PMC 4491144. PMID 26140597.
- ^ Gonzalez-Sanchez MB, Lanucara F, Hardman GE, Eyers CE (2014). „Přenos intermolekulárního fosfátu v plynné fázi v dimeru fosfohistidinu a fosfopeptidu“. Hmotnostní spektrum Int J. 367: 28–34. Bibcode:2014 IJMSp.367 ... 28G. doi:10.1016 / j.ijms.2014.04.015. PMC 4375673. PMID 25844054.
- ^ Gonzalez-Sanchez MB, Lanucara F, Helm M, Eyers CE (2013). "Pokus o přepsání historie: výzvy s analýzou histidin-fosforylovaných peptidů". Biochem Soc Trans. 41 (4): 1089–1095. doi:10.1042 / bst20130072. PMID 23863184.
- ^ Hardman G, Perkins S, Ruan Z, Kannan N, Brownridge P, Byrne DP, Eyers PA, Jones AR, Eyers CE (13. října 2017). „Rozsáhlá nekanonická fosforylace v lidských buňkách odhalena pomocí fosfoproteomiky zprostředkované silnou aniontovou výměnou“. bioRxiv 10.1101/202820.
- ^ A b Marina A, Waldburger CD, Hendrickson WA (prosinec 2005). „Struktura celé cytoplazmatické části proteinu histidinkinázového senzoru“. EMBO J.. 24 (24): 4247–59. doi:10.1038 / sj.emboj.7600886. PMC 1356327. PMID 16319927.
- ^ Parkinson JS, Kofoid EC (1992). "Komunikační moduly v bakteriálních signálních proteinech". Annu. Genet. 26: 71–112. doi:10.1146 / annurev.ge.26.120192.000443. PMID 1482126.
- ^ Bilwes AM, Quezada CM, Croal LR, Crane BR, Simon MI (duben 2001). "Nukleotidová vazba histidinkinázou CheA". Nat. Struct. Biol. 8 (4): 353–60. doi:10.1038/86243. PMID 11276258. S2CID 25434861.
- ^ Torosantucci A, Chiani P, De Bernardis F, Cassone A, Calera JA, Calderone R (únor 2002). „Odstranění dvousložkového genu histidinkinázy (CHK1) z Candida albicans přispívá ke zvýšené inhibici růstu a usmrcování lidskými neutrofily in vitro“. Infikovat. Immun. 70 (2): 985–7. doi:10.1128 / IAI.70.2.985-987.2002. PMC 127696. PMID 11796636.
Další čtení
- Kowluru A (2002). „Identifikace a charakterizace nové proteinové histidinkinázy v beta buňce ostrůvků: důkazy o její regulaci mastoparanem, aktivátorem G-proteinů a sekrecí inzulínu“. Biochem. Pharmacol. 63 (12): 2091–100. doi:10.1016 / S0006-2952 (02) 01025-0. PMID 12110368.
- Yoshimi A, Tsuda M, Tanaka C (2004). „Klonování a charakterizace genu histidinkinázy Dic1 z Cochliobolus heterostrophus, který propůjčuje rezistenci na dikarboximid a osmotickou adaptaci“. Mol. Genet. Genomika. 271 (2): 228–36. doi:10.1007 / s00438-003-0974-4. PMID 14752661. S2CID 26038953.
- Beier D, Frank R (2000). "Molekulární charakterizace dvousložkových systémů Helicobacter pylori". J. Bacteriol. 182 (8): 2068–76. doi:10.1128 / JB.182.8.2068-2076.2000. PMC 111253. PMID 10735847.
- Pflock M, Dietz P, Schar J, Beier D (2004). „Genetické důkazy o tom, že histidinkináza HP165 je kyselým senzorem Helicobacter pylori“. FEMS Microbiol. Lett. 234 (1): 51–61. doi:10.1111 / j.1574-6968.2004.tb09512.x. PMID 15109719.
- Roberts DL, Bennett DW, Forst SA (1994). "Identifikace místa fosforylace na osmosenzoru, EnvZ, z Escherichia coli". J. Biol. Chem. 269 (12): 8728–33. PMID 8132603.
- Alexandrine M. Bilwes; Lisa A. Alex; Brian R. Crane; Melvin I. Simon (1999). "Struktura CheA, signální transdukční histidinkinázy". Buňka. 96 (1): 131–41. doi:10.1016 / S0092-8674 (00) 80966-6. PMID 9989504. S2CID 16842653.
- Ryan L. Brunsing; Chandra La Clair; Sharon Tang; Christina Chiang; Lynn E. Hancock; Marta Perego; James A Hoch (2005). "Charakterizace sporulačních histidinkináz z Bacillus anthracis". J. Bacteriol. 187 (20): 6972–81. doi:10.1128 / JB.187.20.6972-6981.2005. PMC 1251614. PMID 16199567.
- Amr Eldakak; F. Marion Hulett (2007). „Cys303 v histidin kináze PhoR je klíčový pro reakci fosfotransferu ve dvousložkovém systému PhoPR v Bacillus subtilis“. J. Bacteriol. 189 (2): 410–21. doi:10.1128 / JB.01205-06. PMC 1797398. PMID 17085571.
- Hirschman A, Boukhvalova M, VanBruggen R, Wolfe AJ, Stewart RC (listopad 2001). „Mutace aktivního místa v CheA, signální transdukční protein kináze chemotaxního systému v Escherichia coli“. Biochemie. 40 (46): 13876–87. doi:10.1021 / bi0113622. PMID 11705377.