Turn (biochemie) - Turn (biochemistry)
A otáčet se je prvek sekundární struktura v proteinech, kde polypeptidový řetězec obrací svůj celkový směr.
Definice
Podle jedné definice[1] zatáčka je strukturální motiv, kde je Cα atomy dvou zbytků oddělených několika (obvykle 1 až 5) peptidové vazby jsou blízko (méně než 7A [0.70 nm ]). Blízkost terminálu Cα atomy často korelují s vytvořením inter hlavního řetězce vodíková vazba mezi odpovídajícími zbytky. Taková vodíková vazba je základem pro původní, snad lépe známou definici odbočky. V mnoha případech, ale ne ve všech, jsou vodíkové vazby a Cα- definice vzdáleností jsou ekvivalentní.
Druhy zatáček
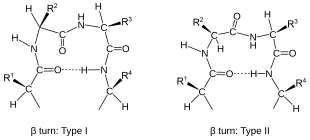
Tahy jsou klasifikovány[2] podle oddělení mezi dvěma koncovými zbytky:
- V α-turn koncové zbytky jsou odděleny čtyři peptidové vazby (i → i ± 4).
- V β-turn (nejběžnější forma) podle tři vazby (i → i ± 3).
- V γ-turntím, že dva vazby (i → i ± 2).
- V δ-turntím, že jeden vazba (i → i ± 1), což je stericky nepravděpodobné.
- V π-turntím, že Pět vazby (i → i ± 5).
| Typ | φi + 1 | ψi + 1 | φi + 2 | ψi + 2 |
|---|---|---|---|---|
| Já | −60° | −30° | −90° | 0° |
| II | −60° | 120° | 80° | 0° |
| VIII | −60° | −30° | −120° | 120° |
| Já | 60° | 30° | 90° | 0° |
| II ' | 60° | −120° | −80° | 0° |
| VIa1 | −60° | 120° | −90° | 0°* |
| VIa2 | −120° | 120° | −60° | 0°* |
| VIb | −135° | 135° | −75° | 160°* |
| IV | tahy vyloučeny ze všech výše uvedených kategorií | |||
Tahy jsou klasifikovány podle jejich páteře vzepětí (vidět Ramachandran spiknutí ). Tahu lze převést na jeho inverzní obrat (ve kterém mají atomy hlavního řetězce opak chirality ) změnou znaménka na jeho vzepětí. (Inverzní obrat není pravdivý enantiomer protože C.α atom chirality γ-turn má tedy dvě formy, klasickou formu s (φ, ψ) roztečné úhly zhruba (75 °, -65 °) a inverzní tvar s roztečnými úhly (-75 °, 65 °). Nejméně osm forem beta turn nastat, lišící se v tom, zda a cis je zahrnut izomer peptidové vazby a na dihedrálních úhlech dvou centrálních zbytků. Klasické a inverzní β-otáčky se vyznačují prvočíslem, např., typ I a typ I ' beta zatáčky. Pokud i → i + 3 vodíková vazba je považována za kritérium pro zatáčky, čtyři kategorie Venkatachalamu[4] (I, II, II ', I') stačí[5] popsat vše možné beta zatáčky. Všechny čtyři se často vyskytují v proteinech, ale I je nejčastější, následovaný II, I 'a II' v tomto pořadí.
Smyčky
An ω smyčka je univerzální termín pro delší, prodlouženou nebo nepravidelnou smyčku bez pevné vnitřní vodíkové vazby.
Více zatáček
V mnoha případech je jeden nebo více zbytků zahrnuto ve dvou částečně se překrývajících zatáčkách. Například v sekvenci 5 zbytků tvoří zbytky 1 až 4 a zbytky 2 až 5 obrat; v takovém případě se mluví o (i, i + 1) dvojité otočení. V proteinech se běžně vyskytuje několik otáček (až sedmkrát).[6] Beta ohýbejte pásky jsou jiný typ vícenásobného otočení.
Vlásenky
A sponka do vlasů je speciální případ obratu, ve kterém se směr proteinové páteře obrací a hraniční prvky sekundární struktury interagují. Například a vlásenka beta spojuje dva vodíkově vázané, antiparalelní β-vlákna (poněkud matoucí název, protože β-vlásenka může obsahovat mnoho typů závitů - α, β, γ atd.).
Beta vlásenky lze klasifikovat podle počtu zbytků, které tvoří tah - to znamená, že jsou ne část lemujících β-řetězců.[7] Pokud je toto číslo X nebo Y (podle dvou různých definic β listů), β vlásenka je definována jako X: Y.
Beta se otočí na konci smyčky vlásenky beta mít odlišné rozdělení typů od ostatních; typ I ′ je nejběžnější, následovaný typy II ′, I a II.
Flexibilní linkery
Obraty se někdy nacházejí uvnitř flexibilní linkery nebo smyčky připojení proteinové domény. Linkerové sekvence mají různou délku a jsou obvykle bohaté na polární náboje aminokyseliny. Flexibilní linkery umožňují připojování domén k volnému kroucení a otáčení za účelem náboru jejich vazebných partnerů dynamika proteinové domény. Umožňují také svým vazebným partnerům vyvolat větší rozsah konformační změny dálkovým dosahem allostery[8][9][10]
Role ve skládání bílkovin
Byly navrženy dvě hypotézy týkající se role obratů skládání bílkovin. V jednom pohledu hrají otočky zásadní roli při skládání spojením a povolením nebo povolením interakcí mezi běžnými prvky sekundární struktury. Tento názor podporují studie mutageneze, které naznačují kritickou roli konkrétních zbytků v obloucích některých proteinů. Také nepůvodní izomery X−Pro peptidové vazby zase může zcela blokovat konformační skládání některých proteinů. V opačném pohledu hrají zatáčky pasivní roli při skládání. Tento názor podporuje špatná ochrana aminokyselin pozorovaná ve většině obratů. Také nepůvodní izomery mnoha X-Pro peptidové vazby v zatáčkách mají malý nebo žádný vliv na skládání.
Metody predikce beta otočení
V průběhu let bylo vyvinuto mnoho metod predikce beta tahů. Nedávno, Skupina Dr. Raghavy rozvinutý BetaTPred3 metoda, která předpovídá spíše kompletní beta tah než jednotlivé zbytky spadající do beta kola. Metoda také dosahuje dobré přesnosti a je první metodou, která předpovídá všech 9 typů beta tahů. Kromě predikce lze tuto metodu také použít k vyhledání minimálního počtu mutací potřebných k zahájení nebo rozbití beta zatáčky v proteinu na požadovaném místě.
Viz také
Poznámky
- ^ viz Rose a kol. 1985 v referencích
- ^ Toniolo 1980
- ^ Venkatachalam 1968; Richardson 1981; Hutchinson a Thornton 1994
- ^ Venkatachalam, CM (1968). "Sterochemická kritéria pro polypeptidy a proteiny. V. Konformace systému tří spojených peptidových jednotek" (PDF). Biopolymery. 6 (10): 1425–1436. doi:10,1002 / bip.1968.360061006. hdl:2027.42/37819. PMID 5685102.
- ^ Richardson, JS (1981). Anatomie a taxonomie proteinové struktury. Adv Protein Chem. Pokroky v chemii proteinů. 34. 167–339. doi:10.1016 / s0065-3233 (08) 60520-3. ISBN 9780120342341. PMID 7020376.
- ^ Hutchinson 1994, s. 2213
- ^ Sibanda 1989
- ^ Dunker AK, Lawson JD, Brown CJ, Williams RM, Romero P, Oh JS, Oldfield CJ, Campen AM, Ratliff CM, Hipps KW, Ausio J, Nissen MS, Reeves R, Kang C, Kissinger CR, Bailey RW, Griswold MD , Chiu W, Garner EC, Obradovic Z (2001). "Jiskrově neuspořádaný protein". Journal of Molecular Graphics & Modeling. 19 (1): 26–59. CiteSeerX 10.1.1.113.556. doi:10.1016 / s1093-3263 (00) 00138-8. PMID 11381529.
- ^ Bu Z, Callaway DJ (2011). "Proteiny se pohybují! Dynamika proteinů a dálková dálnice v buněčné signalizaci". Struktura bílkovin a nemoci. Pokroky v chemii proteinů a strukturní biologii. 83. 163–221. doi:10.1016 / B978-0-12-381262-9.00005-7. ISBN 9780123812629. PMID 21570668.
- ^ Compiani M, Capriotti E (prosinec 2013). "Výpočetní a teoretické metody skládání proteinů" (PDF). Biochemie. 52 (48): 8601–24. doi:10.1021 / bi4001529. PMID 24187909. Archivovány od originál (PDF) dne 04.09.2015.
externí odkazy
- BetaTPred3 - platforma Insilico pro předpovídání a zahájení betaturnů v proteinu na požadovaném místě Odkaz na článek
- NetTurnP - Predikce Beta-turn oblastí v proteinových sekvencích
- BetaTPred - Predikce beta změn v proteinech pomocí statistických algoritmů
Reference
Tyto reference jsou seřazeny podle data.
- Venkatachalam CM. (1968). "Stereochemická kritéria pro polypeptidy a proteiny. V. Konformace systému tří spojených peptidových jednotek". Biopolymery. 6 (10): 1425–36. doi:10,1002 / bip.1968.360061006. hdl:2027.42/37819. PMID 5685102.
- Némethy, George; Printz, Morton P. (1972). „The -Turn, možná složená konformace polypeptidového řetězce. Srovnání s β-Turnem “. Makromolekuly. 5 (6): 755–758. doi:10.1021 / ma60030a017.
- Lewis PN, Momany FA, Scheraga HA (1973). "Řetězové obraty v proteinech". Biochim Biophys Acta. 303 (2): 211–29. doi:10.1016/0005-2795(73)90350-4. PMID 4351002.
- Toniolo C .; Benedetti, Ettore (1980). "Intramolekulárně vodíkově vázané peptidové konformace". CRC Crit Rev Biochem. 9 (1): 1–44. doi:10.3109/10409238009105471. PMID 6254725.
- Richardson JS. (1981). Anatomie a taxonomie proteinové struktury. Adv Protein Chem. Pokroky v chemii proteinů. 34. 167–339. doi:10.1016 / S0065-3233 (08) 60520-3. ISBN 9780120342341. PMID 7020376.
- Rose GD, Gierasch LM, Smith JA (1985). Obrací peptidy a bílkoviny. Adv Protein Chem. Pokroky v chemii proteinů. 37. s. 1–109. doi:10.1016 / S0065-3233 (08) 60063-7. ISBN 9780120342372. PMID 2865874.
- Milner-White EJ, Poet R (1987). "Smyčky, boule, zatáčky a vlásenky v bílkovinách". Trends Biochem Sci. 12: 189–192. doi:10.1016/0968-0004(87)90091-0.
- Wilmot CM, Thornton JM (1988). "Analýza a predikce různých typů beta-turn v proteinech". J Mol Biol. 203 (1): 221–32. doi:10.1016/0022-2836(88)90103-9. PMID 3184187.
- Sibanda, B.L .; Blundell, T.L .; Thornton, J. M. (1989). "Konformace β-vlásenek v proteinových strukturách: Systematická klasifikace s aplikacemi pro modelování pomocí homologie, přizpůsobení elektronové hustoty a proteinového inženýrství". Journal of Molecular Biology. 206 (4): 759–777. doi:10.1016/0022-2836(89)90583-4. PMID 2500530.
- Milner-White, E (1990). „Situace gama-obratů v proteinechJejich vztah k alfa-helixům, beta-listům a vazebným místům pro ligand“. J. Mol. Biol. 216 (2): 385–397. doi:10.1016 / S0022-2836 (05) 80329-8. PMID 2254936.
- Hutchinson, E.G .; Thornton, J.M. (1994). „Revidovaná sada potenciálů pro tvorbu β-obratu v proteinech“. Věda o bílkovinách. 3 (12): 2207–2216. doi:10.1002 / pro.5560031206. PMC 2142776. PMID 7756980.
- Pavone V, Gaeta G, Lombardi A, Nastri F, Maglio O, Isernia C, Saviano M (1996). "Objevování sekundárních struktur proteinů: klasifikace a popis izolovaných alfa-obratů". Biopolymery. 38 (6): 705–21. doi:10.1002 / (SICI) 1097-0282 (199606) 38: 6 <705 :: AID-BIP3> 3.0.CO; 2-V. PMID 8652792.
- Rajashankar KR, Ramakumar S (1996). „Pi-obraty v proteinech a peptidech: Klasifikace, konformace, výskyt, hydratace a sekvence“. Protein Sci. 5 (5): 932–46. doi:10.1002 / pro.5560050515. PMC 2143406. PMID 8732765. Archivovány od originál dne 2009-05-24.
