Enzym aktivující ubikvitin - Ubiquitin-activating enzyme
| Enzymy aktivující ubikvitin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Krystalová struktura komplexu E1 / ubikvitin aktivujícího enzym kvasinek ubikvitin.[1] | |||||||||
| Identifikátory | |||||||||
| EC číslo | 6.2.1.45 | ||||||||
| Číslo CAS | 74812-49-0 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Enzymy aktivující ubikvitin, také známý jako E1 enzymy, katalyzují první krok v ubikvitinace reakce, která (mimo jiné) může zacílit protein na degradaci prostřednictvím a proteazom. Tento kovalentní vazba z ubikvitin nebo ubikvitin podobné proteiny cílené proteiny je hlavním mechanismem pro regulaci funkce proteinu v eukaryotické organismy.[2] Mnoho procesů, jako je buněčné dělení, imunitní reakce a embryonální vývoj jsou rovněž regulovány posttranslační modifikace ubikvitinem a ubikvitinovými proteiny.[2]
Ubikvitinace (ubikvitylace)
Enzym aktivující ubikvitin (E1) zahajuje proces ubikvitinace (obrázek 1). Enzym E1 spolu s ATP se váže na ubikvitinový protein. Enzym E1 poté předá ubikvitinový protein druhému proteinu, který se nazývá ubiquitinový nosič nebo konjugační protein (E2). E2 proteinové komplexy s a ubikvitinová proteinová ligáza (E3). Tato ubikvitinová proteinová ligáza rozpoznává, který protein je třeba označit, a katalyzuje přenos ubikvitinu na tento protein. Tato cesta se opakuje, dokud cílový protein nemá k sobě připojený celý řetězec ubikvitinu.[3]
Struktura a mechanismus
Na začátku ubikvitinační kaskády se enzym E1 (obrázek 2) váže na ATP-Mg2+ a ubikvitin a katalyzuje C-koncovou acylovou adenylaci ubikvitinu.[4] V dalším kroku katalytický cystein (Obrázek 3) na enzymu E1 napadá komplex ubikvitin-AMP prostřednictvím acylové substituce a současně vytváří thioesterová vazba a odcházející skupina AMP.[2] Nakonec komplex E1-ubikvitin přenáší ubikvitin na enzym E2 transthioesterifikační reakcí, při které E2 katalytický cystein útočí na zadní stranu komplexu E1-ubikvitin.[5] Proces transthioesterifikace je však velmi komplikovaný, protože jak enzymy E1, tak E2 tvoří meziproduktový komplex, kde oba enzymy procházejí řadou konformační změny aby se navzájem svázaly.[5]
V průběhu tohoto mechanismu je enzym E1 vázán na dvě molekuly ubikvitinu. I když je tento sekundární ubikvitin podobně adenylovaný, netvoří stejný thioesterový komplex, který byl popsán výše. Funkce sekundárního ubikvitinu zůstává do značné míry neznámá, předpokládá se však, že může usnadnit konformační změny pozorované v enzymu E1 během transthioesterifikačního procesu.[2]

Obrázek 1. ukazuje sekvenci toho, jak se enzym aktivující ubikvitin váže na ATP a ubikvitinový substrát. Ukazuje také, jak lze najednou vázat dva ubikvitinové substráty.[6]

Obrázek 2. Protein E1 váže molekulu ubikvitinu v každém ze dvou identických aktivních míst (zvýrazněno). Důležité zbytky, cystein a arginin, jsou označeny červeně.[2]

Obrázek 3. Detailní pohled na nevázaný aktivní web. Předpokládá se, že Arg (603) dobije katalytický Cys (600), jakmile byl ubikvitin přenesen do enzymu E2.[2]
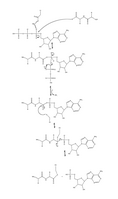
Obrázek 4. Plný mechanismus pro adenylaci ubikvitinu a následnou vazbu ubikvitinu na E1.
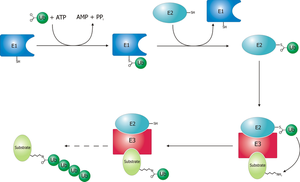
Schematické znázornění ubikvitylačního systému.





Isozymes
Následující geny kódují enzymy aktivující ubikvitin:
Sdružení nemocí
Ubiquitin-proteazomový systém je velmi důležitý degradace bílkovin uvnitř buněk. Dysfunkce tohoto systému mohou narušit buněčnou homeostázu a vést k řadě poruch. V normálně fungujících buňkách mění kovalentní vazba ubikvitinu nebo proteinu podobného ubikvitinu na cílový protein povrch cílového proteinu. Tyto ubikvitinované proteiny podléhají degradaci proteolytickými a neproteolytickými cestami.[7] Pokud tento systém nefunguje správně, může dojít k mnoha dědičným a získaným chorobám, jako je rakovina, cukrovka, mrtvice, Alzheimerova choroba, Amyotrofní laterální skleróza, roztroušená skleróza, astma, zánětlivé onemocnění střev, autoimunitní tyroiditida, zánětlivá artritida, a lupus.[7]
Missense in UBE1 a X-spojená infantilní spinální svalová atrofie (XL-SMA)
Mezi různé poruchy spojené s ubikvitin-proteazomovou cestou patří X-vázaný infantilní spinální svalová atrofie (XL-SMA).[8] Fatální dětská porucha je spojena se ztrátou buňky předního rohu a dětská smrt. Mezi klinické příznaky patří hypotonie, areflexie a mnohočetné vrozené kontraktury. Při analýze mutací ve velkém měřítku poskytl screening šesti rodin XL-SMA výsledky naznačující dva nové missense mutace ve dvou rodinách a nová synonymní substituce C → T v dalších třech rodinách. Všechny tyto detekované mutace byly lokalizovány v exonu 15 UBE1 gen (gen kódující enzym aktivující ubikvitin) a bylo pozorováno, že se v rodinách segregují s onemocněním. Stručně řečeno, UBE1 missense může vést k narušené složité budově s gigaxonin, protein zapojený do axonální struktury a udržování neuronů. To může vést ke zhoršené degradaci protein 1B spojený s mikrotubuly (MAP1B), což vede k hromadění proteinu MAP1B, což může zvýšit smrt neuronových buněk.[8] Mutace tedy UBE1 existuje podezření, že jsou příčinou genetických vad u jedinců XL-SMA.
Reference
- ^ PDB: 3 CMM; Lee I, Schindelin H (červenec 2008). „Strukturální pohledy na aktivaci ubikvitinu katalyzovanou E1 a přenos na konjugující enzymy“. Buňka. 134 (2): 268–78. doi:10.1016 / j.cell.2008.05.046. PMID 18662542.
- ^ A b C d E F Schulman BA, Harper JW (květen 2009). „Aktivace proteinu podobného ubikvitinu enzymy E1: vrchol pro signální dráhy po proudu“. Nature Reviews Molecular Cell Biology. 10 (5): 319–31. doi:10.1038 / nrm2673. PMC 2712597. PMID 19352404.
- ^ Lecker SH, Goldberg AL, Mitch WE (červenec 2006). „Degradace proteinu cestou ubikvitin-proteazomu v normálním a chorobném stavu“ (PDF). Časopis Americké nefrologické společnosti. 17 (7): 1807–19. doi:10.1681 / ASN.2006010083. PMID 16738015.
- ^ Tokgöz Z, Bohnsack RN, Haas AL (květen 2006). "Pleiotropní účinky vazby ATP.Mg2 + v katalytickém cyklu enzymu aktivujícího ubikvitin". The Journal of Biological Chemistry. 281 (21): 14729–37. doi:10,1074 / jbc.M513562200. PMID 16595681.
- ^ A b Lee I, Schindelin H (červenec 2008). „Strukturální pohledy na aktivaci ubikvitinu katalyzovanou E1 a přenos na konjugující enzymy“. Buňka. 134 (2): 268–78. doi:10.1016 / j.cell.2008.05.046. PMID 18662542.
- ^ Tento obrázek je převzat ze schématu 1 „Mechanismu enzymu aktivujícího ubikvitin: analýza kinetiky a rovnováhy“ od Arthura Haase. Haas AL, Rose IA (září 1982). "Mechanismus enzymu aktivujícího ubikvitin. Kinetická a rovnovážná analýza". The Journal of Biological Chemistry. 257 (17): 10329–37. PMID 6286650.
- ^ A b Wang J, Maldonado MA (srpen 2006). „Systém ubikvitin-proteazom a jeho role při zánětlivých a autoimunitních onemocněních“ (PDF). Buněčná a molekulární imunologie. 3 (4): 255–61. PMID 16978533.
- ^ A b Ramser J, Ahearn ME, Lenski C a kol. (Leden 2008). „Vzácná missense a synonymní varianty v UBE1 jsou spojeny s infantilní spinální svalovou atrofií spojenou s X“. American Journal of Human Genetics. 82 (1): 188–93. doi:10.1016 / j.ajhg.2007.09.009. PMC 2253959. PMID 18179898.
externí odkazy
- Ubiquitin-Activating + Enzymes v americké národní lékařské knihovně Lékařské předměty (Pletivo)