Adenylosukcinát lyáza - Adenylosuccinate lyase




| Adenylosukcinát lyáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
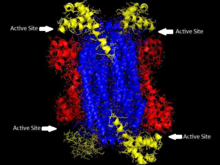 "Homotetramerická struktura ASL v Thermotoga maritima Doména 1 je červená, Doména 2 je modře, Doména 3 je žlutá. Tato struktura byla inspirována článkem Toth a Yeates[5] | |||||||||
| Identifikátory | |||||||||
| EC číslo | 4.3.2.2 | ||||||||
| Číslo CAS | 9027-81-0 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Adenylosukcinát lyáza (nebo adenylosukcináza) je enzym že u lidí je kódováno ADSL gen.[6]
Adenylosukcinát lyáza se převádí adenylosukcinát na AMP a fumarát jako součást purinový nukleotidový cyklus. ASL katalyzuje dvě reakce v purinové biosyntetické dráze, která vytváří AMP; ASL štěpí adenylosukcinát na AMP a fumarát a štěpí SAICAR do AICAR a fumarát.
Adenylosukcinát lyáza je součástí β-eliminace nadrodina enzymů a probíhá prostřednictvím E1cb mechanismus reakce. Enzym je a homotetramer se třemi doménami v každé monomer a čtyři aktivní stránky na homotetramer.
Bodové mutace v adenylosukcinátu, které způsobují snížení enzymatická aktivita způsobit klinické příznaky, které tento stav poznamená nedostatek adenylosukcinát lyázy.
Tento protein může používat morpheein model alosterická regulace.[7]
Funkce

Adenylosukcinát lyáza (ASL) je enzym, který katalyzuje dvě reakce v de novo purinový biosyntetický cesta. V obou reakcích používá E1cb eliminační reakce mechanismus k odštěpení fumarátu od substrátu. V první reakci ASL převádí 5-aminoimidazol- (N-sukcinylokarboxamid) ribotid (SAICAR) na 5-aminoimidazol-4-karboxamid ribotid (AICAR) a fumarát. AICAR pokračuje třemi dalšími reakcemi, než se z něj stane adenylosukcinát (také nazývaný sukcinyladenosin monofosfát nebo SAMP), který se ASL poté rozdělí na adenosin monofosfát (AMP) a fumarát.[8] ASL je důležitý pro buňky nejen kvůli jeho zapojení do vytváření purinů potřebných pro buněčná replikace, ale také proto, že pomáhá regulovat metabolické procesy řízením hladin AMP a fumarátu v buňce.[9]

Struktura
Podjednotky
Adenylosukcinát lyáza patří do β-eliminační nadrodiny a jako taková je její struktura homotetramer. Monomer adenylosukcinát lyázy má tři domény. v Thermotoga maritima, doména 1 obsahuje 7 α-šroubovice v zbytky 1-93, včetně His68, který je vysoce konzervovaný a dříve byl považován za katalyzátor kyselina v Aktivní stránky.[5] Novější studie předpokládají, že His171 v doméně 2, dříve považovaný za katalyzátor základna, může ve skutečnosti působit jako katalytická kyselina, alespoň v Escherichia coli.[9] Doména 2 je tvořena zbytky 94-341 a obsahuje pouze 5 α-šroubovic a monomery β-list. Doménu 3 tvoří 7 α-šroubovic. Jádro tetrameru je tvořeno čtyřmi kopiemi domény 2 a na každém konci tetrameru jsou dvě kopie každé z domén 1 a 3, které dávají tetramer D2 dihedrální symetrie. Tetramer má čtyři aktivní místa, z nichž každé se setkává se třemi doménami.[5]
Adenylosukcinát lyáza u lidí a Bacillus subtilis může být kompetitivně potlačeno podle substrát analog kyselina 2 '(3'), 5'-difosfát adenosin fosfonobutanová (APBADP). APBADP je kompetitivní inhibitor pro obě reakce katalyzované adenylosukcinát lyázou a kinetické studie s APBADP ukazují, že substráty pro obě reakce používají stejné aktivní místo.[10] V reakci katalyzované ASL štěpením adenylosukcinátu na adenosinmonofosfát (AMP) a fumarát musí AMP po ukončení reakce a před uvolněním fumarátu mírně rotovat, aby se oba produkty vešly na aktivní místo.[11]
Mutace
Adenylosukcinát lyáza mutanti může mít značně sníženou aktivitu, ať už je mutace v aktivním místě nebo mimo něj. Mutanti ASL způsobující nemoci R396C a R396H jsou u vstupu do aktivního místa a mají nižší PROTImax než ASL divokého typu, ale mutanty K246E a L311V, které jsou mimo aktivní místo, také způsobují snížení Vmax. ASL mutant R194C je mimo aktivní místo a udržuje Vmax podobně jako ASL divokého typu se ukázalo, že je nejméně konformačně stabilní z pěti mutantů in vitro a stále způsobuje nemoci.[12]
Mechanismus
Dříve se předpokládalo, že mechanismem účinku adenylosukcinát lyázy byla společná katalýza, při které byl vodík na β-uhlíku (s ohledem na odcházející dusík) abstrahován katalytickou bází současně s tím, že odcházející dusík byl protonován katalytická kyselina pro eliminaci E2.[5] Novější údaje jsou v rozporu s touto myšlenkou a potvrdily, že mechanismus není ve skutečnosti koordinovaný, ale že k abstrakci dochází jako první a existuje přechodná karbanion druh, který je rezonančně stabilizovaný. U obou reakcí katalyzovaných ASL dochází nejprve k deprotonaci uhlíku β na odcházející dusík, poté k tvorbě a rezonanční stabilizace karbaniontu a nakonec protonace odcházejícího dusíku, která způsobí rozbití vazby C-N.[9] Experimentální potvrzení deprotonace, tvorby karbaniontu a kroku omezujícího rychlost protonace způsobujícího štěpení znamená, že se jedná o mechanismus E1cb. Nejnovější údaje naznačují, že katalytickou kyselinou je His171, o které se dříve myslelo, že je katalytickou bází, a že poněkud neobvykle jde o serin v poloze 295, který působí jako katalytická báze. Štěpení adenylosukcinátu na AMP a fumarát je uspořádaný uni-bi mechanismus, což znamená, že po štěpení fumarát opouští aktivní místo před AMP.[13]

Role v nemoci
Mutovaná adenylosukcinát lyáza (ASL) způsobuje klinické onemocnění u pacientů, které se označuje jako nedostatek adenylosukcinát lyázy. Tento stav je vzácný a projevuje se různým stupněm psychomotorická retardace, autismus, ztráta svalové hmoty, a epilepsie.[14][15] Přesná příčina nemoci není známa, ale možností je málo purin nukleotid syntéza pro replikace buněk, nefunkčnost purinový nukleotidový cyklus a hromadění substrátů na toxické hladiny. Bylo identifikováno několik bodových mutací souvisejících s chorobami a ti, kteří jsou heterozygotní pro bodovou mutaci jsou zdraví, ale ti, kteří jsou homozygotní vyvinout klinické onemocnění.[16] Počet genotypů způsobujících onemocnění se neustále zvyšuje, protože je objevováno více mutací, a nyní bylo dosud identifikováno třicet různých bodových mutací a jedna delece, která způsobuje nedostatek adenylosukcinátu lyázy.[17]
Když se substráty ASL (adenylosukcinát a SAICAR) hromadí v důsledku nedostatku enzymů, jsou defosforylovaný a přeměnit se na sukcinyladenosin (S-Ado) a sukcinylaminoimidazol karboximid ribosid (SAICA ribosid).[18] Normálně tyto sloučeniny nejsou přítomny v mozkomíšním moku nebo v moči, protože ASL působí na většinu molekul substrátu, než se mohou hromadit a být fosforylovány.[15] V minulosti neexistoval dobrý test na nedostatek adenylosukcinát lyázy, který by diagnostikoval vzácné onemocnění, ale nedávno byl vyvinut test na detekci SAICA a S-Ado v moči. Test je levný a v malém vzorku výzkumníků neměl falešná pozitiva ani falešná negativa.[19]
Předpokládá se, že SAICA ribosid může být toxičtější sloučeninou, protože se vyskytuje ve vyšších hladinách u pacientů se závažnými klinickými příznaky a někteří vědci si myslí, že S-Ado může být dokonce ochranný. Je třeba provést další výzkum toho, co určuje závažnost onemocnění, ale nestabilita lidského ASL v laboratorním prostředí byla překážkou tohoto výzkumu.[17]
Terapeutické aplikace
Jak se zvyšuje odolnost proti antimalarikům, vědci hledají nové strategie, jak se zaměřit na Plasmodium parazity, které způsobují malárie, zvláště smrtící P. falciparum. Někteří vědci navrhli, aby se na ASL pohlíželo jako na potenciální drogový cíl, protože i když přerušení léčby de novo dráha biosyntézy purinu je pro hostitele toxická, Plasmodium ASL má nízkou úroveň sekvenční homologie s lidským ASL, které mohou způsobitPlasmodium ASL léky dostatečně specifické, aby nepoškodily lidské hostitele.[20]
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000239900 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000022407 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b C d E Toth EA, Yeates TO (únor 2000). "Struktura adenylosukcinát lyázy, enzymu s dvojí aktivitou v de novo purinové biosyntetické cestě". Struktura. 8 (2): 163–74. doi:10.1016 / S0969-2126 (00) 00092-7. PMID 10673438.
- ^ „Entrez Gene: Adenylosuccinate lyase“. Citováno 2012-03-01.
- ^ Selwood T, Jaffe EK (březen 2012). „Dynamické disociační homo-oligomery a řízení funkce proteinu“. Archivy biochemie a biofyziky. 519 (2): 131–43. doi:10.1016 / j.abb.2011.11.020. PMC 3298769. PMID 22182754.
- ^ Spiegel EK, Colman RF, Patterson D (2006). "Nedostatek adenylosukcinát lyázy". Molekulární genetika a metabolismus. 89 (1–2): 19–31. doi:10.1016 / j.ymgme.2006.04.018. PMID 16839792.
- ^ A b C d Tsai M, Koo J, Yip P, Colman RF, Segall ML, Howell PL (červenec 2007). „Substrát a produktové komplexy adenylosukcinátové lyázy Escherichia coli poskytují nový pohled na enzymatický mechanismus“. Journal of Molecular Biology. 370 (3): 541–54. doi:10.1016 / j.jmb.2007.04.052. PMC 4113493. PMID 17531264.
- ^ Sivendran S, Colman RF (červenec 2008). "Účinek nového neštěpitelného analogu substrátu na divoký typ a serinové mutanty v signální sekvenci adenylosukcinát lyázy z Bacillus subtilis a Homo sapiens". Věda o bílkovinách. 17 (7): 1162–74. doi:10.1110 / ps.034777.108. PMC 2442012. PMID 18469177.
- ^ Kozlov G, Nguyen L, Pearsall J, Gehring K (září 2009). „Struktura fosfátem vázané lysázy Escherichia coli adenylosukcinát identifikuje His171 jako katalytickou kyselinu“. Acta Crystallographica oddíl F. 65 (Pt 9): 857–61. doi:10.1107 / S1744309109029674. PMC 2795585. PMID 19724117.
- ^ Ariyananda Lde Z, Lee P, Antonopoulos C, Colman RF (červen 2009). „Biochemická a biofyzikální analýza pěti mutantů lidské adenylosukcinát-lyázy spojené s onemocněním“. Biochemie. 48 (23): 5291–302. doi:10,1021 / bi802321m. PMC 2745324. PMID 19405474.
- ^ Bulusu V, Srinivasan B, MP Bopanna, Balaram H (duben 2009). "Elucidace substrátové specificity, kinetického a katalytického mechanismu adenylosukcinát lyázy z Plasmodium falciparum". Biochimica et Biophysica Acta (BBA) - bílkoviny a proteomika. 1794 (4): 642–54. doi:10.1016 / j.bbapap.2008.11.021. PMID 19111634.
- ^ Maaswinkel-Mooij PD, Laan LA, Onkenhout W, Brouwer OF, Jaeken J, Poorthuis BJ (srpen 1997). "Nedostatek adenylosukcinázy s epilepsií v raném dětství". Journal of Inherited Metabolic Disease. 20 (4): 606–7. doi:10.1023 / A: 1005323512982. PMID 9266401.
- ^ A b Lee P, Colman RF (únor 2007). "Exprese, čištění a charakterizace stabilní, rekombinantní lidské adenylosukcinátové lyázy". Exprese a čištění proteinů. 51 (2): 227–34. doi:10.1016 / j.pep.2006.07.023. PMID 16973378.
- ^ Stone RL, Aimi J, Barshop BA, Jaeken J, Van den Berghe G, Zalkin H, Dixon JE (duben 1992). "Mutace v adenylosukcinát lyáze spojená s mentální retardací a autistickými rysy". Genetika přírody. 1 (1): 59–63. doi:10.1038 / ng0492-59. PMID 1302001.
- ^ A b Palenchar JB, Crocco JM, Colman RF (srpen 2003). „Charakterizace mutantních lysáz Bacillus subtilis adenylosukcinát odpovídající závažným nedostatkům lidské adenylosukcinát lyázy“. Věda o bílkovinách. 12 (8): 1694–705. doi:10.1110 / ps.0303903. PMC 2323956. PMID 12876319.
- ^ Jaeken J, Van den Berghe G (listopad 1984). "Infantilní autistický syndrom charakterizovaný přítomností sukcinylpurinů v tělních tekutinách". Lanceta. 2 (8411): 1058–61. doi:10.1016 / s0140-6736 (84) 91505-8. PMID 6150139.
- ^ Maddocks J, Reed T (leden 1989). "Test moči na nedostatek adenylosukcinázy u autistických dětí". Lanceta. 1 (8630): 158–9. doi:10.1016 / S0140-6736 (89) 91172-0. PMID 2563072.
- ^ Marshall VM, Coppel RL (září 1997). "Charakterizace genu kódujícího adenylosukcinát lyázu Plasmodium falciparum". Molekulární a biochemická parazitologie. 88 (1–2): 237–41. doi:10.1016 / S0166-6851 (97) 00054-6. PMID 9274883.
Další čtení
- Marie S, Cuppens H, Heuterspreute M, Jaspers M, Tola EZ, Gu XX, Legius E, Vincent MF, Jaeken J, Cassiman JJ, Van den Berghe G (1999). „Mutační analýza v deficitu adenylosukcinát lyázy: osm nových mutací v přehodnocené úplné kódující sekvenci ADSL“. Lidská mutace. 13 (3): 197–202. doi:10.1002 / (SICI) 1098-1004 (1999) 13: 3 <197 :: AID-HUMU3> 3.0.CO; 2-D. PMID 10090474.
- Kmoch S, Hartmannová H, Stibůrková B, Krijt J, Zikánová M, Sebesta I (červen 2000). „Lidská adenylosukcinát lyáza (ADSL), klonování a charakterizace cDNA plné délky a její izoformy, genové struktury a molekulárního základu pro nedostatek ADSL u šesti pacientů“. Lidská molekulární genetika. 9 (10): 1501–13. doi:10,1093 / hmg / 9.10.1501. PMID 10888601.
- Závod V, Marie S, Vincent MF, Van den Berghe G (září 2000). „Klinické, biochemické a molekulárně genetické korelace při nedostatku adenylosukcinát lyázy“. Lidská molekulární genetika. 9 (14): 2159–65. doi:10,1093 / hmg / 9,14 2159. PMID 10958654.
- Tabucchi A, Carlucci F, Rosi F, Guerranti R, Marinello E (červen 2001). "Stanovení, aktivita a biologická role adenylosukcinát lyázy v krevních buňkách". Biomedicína a farmakoterapie. 55 (5): 277–83. doi:10.1016 / s0753-3322 (01) 00061-0. PMID 11428554.
- Marie S, Race V, Nassogne MC, Vincent MF, Van den Berghe G (červenec 2002). „Mutace vazebného místa nukleárního respiračního faktoru 2 v 5 'nepřekládané oblasti genu ADSL u tří pacientů s nedostatkem adenylosukcinát lyázy“. American Journal of Human Genetics. 71 (1): 14–21. doi:10.1086/341036. PMC 384970. PMID 12016589.
- Castro M, Pérez-Cerdá C, Merinero B, García MJ, Bernar J, Gil Nagel A, Torres J, Bermúdez M, Garavito P, Marie S, Vincent F, Van den Berghe G, Ugarte M (srpen 2002). "Screening na nedostatek adenylosukcinát lyázy: klinické, biochemické a molekulární nálezy u čtyř pacientů". Neuropediatrie. 33 (4): 186–9. doi:10.1055 / s-2002-34493. PMID 12368987.
- Palenchar JB, Colman RF (únor 2003). „Charakterizace mutantu Bacillus subtilis adenylosukcinát lyáza ekvivalentní mutantnímu enzymu nalezenému v deficitu lidské adenylosukcinát lyázy: asparagin 276 hraje důležitou strukturální roli“. Biochemie. 42 (7): 1831–41. doi:10.1021 / bi020640 +. PMID 12590570.
- Edery P, Chabrier S, Ceballos-Picot I, Marie S, Vincent MF, Tardieu M (červenec 2003). „Intrafamiliální variabilita fenotypové exprese deficitu adenylosukcinát lyázy: zpráva o třech pacientech“. American Journal of Medical Genetics Part A. 120A (2): 185–90. doi:10.1002 / ajmg.a.2017. PMID 12833398.
- Stone RL, Aimi J, Barshop BA, Jaeken J, Van den Berghe G, Zalkin H, Dixon JE (duben 1992). "Mutace v adenylosukcinát lyáze spojená s mentální retardací a autistickými rysy". Genetika přírody. 1 (1): 59–63. doi:10.1038 / ng0492-59. PMID 1302001.
- Sivendran S, Patterson D, Spiegel E, McGown I, Cowley D, Colman RF (prosinec 2004). „Dvě nové mutantní lidské adenylosukcinátové lyázy (ASL) spojené s autismem a charakterizace ekvivalentního mutantu Bacillus subtilis ASL“. The Journal of Biological Chemistry. 279 (51): 53789–97. doi:10,1074 / jbc.M409974200. PMID 15471876.
externí odkazy
- Adenylosukcinát + lyáza v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- Člověk ADSL umístění genomu a ADSL stránka s podrobnostmi o genu v UCSC Genome Browser.
- Člověk ASL umístění genomu a ASL stránka s podrobnostmi o genu v UCSC Genome Browser.