Proteinogenní aminokyselina - Proteinogenic amino acid

Proteinogenní aminokyseliny jsou aminokyseliny které jsou biosynteticky začleněny do bílkoviny v době překlad. Slovo „proteinogenní“ znamená „vytváření bílkovin“. Skrz známé život, ve standardu je 22 geneticky kódovaných (proteinogenních) aminokyselin, 20 genetický kód a další 2, které lze začlenit pomocí speciálních překladových mechanismů.[1]
V porovnání, neproteinogenní aminokyseliny jsou aminokyseliny, které buď nejsou zabudovány do proteinů (jako GABA, L-DOPA nebo trijodtyronin ), nesprávně zabudované místo geneticky kódované aminokyseliny nebo nevyráběné přímo a izolovaně standardním buněčným mechanizmem (jako hydroxyprolin ). Ta často vyplývá z posttranslační modifikace bílkovin. Některé neproteinogenní aminokyseliny jsou začleněny do nonribozomální peptidy které jsou syntetizovány non-ribozomálními peptidovými syntetázami.
Oba eukaryoty a prokaryoty může začlenit selenocystein do svých proteinů prostřednictvím nukleotidové sekvence známé jako a Prvek SECIS, který směruje buňku k překladu blízkého UGA kodon tak jako selenocystein (UGA je obvykle a stop kodon ). V některých methanogenní prokaryoty, může být také přeložen kodon UAG (obvykle stop kodon) pyrrolysin.[2]
V eukaryotech je pouze 21 proteinogenních aminokyselin, 20 standardního genetického kódu plus selenocystein. Lidé mohou syntetizovat 12 z nich navzájem nebo z jiných molekul intermediárního metabolismu. Dalších devět musí být spotřebováno (obvykle jako jejich proteinové deriváty), a tak se jim říká esenciální aminokyseliny. Esenciální aminokyseliny jsou histidin, isoleucin, leucin, lysin, methionin, fenylalanin, threonin, tryptofan, a valin (tj. H, I, L, K, M, F, T, W, V).[3]
Bylo zjištěno, že proteinogenní aminokyseliny souvisí se sadou aminokyseliny které lze rozpoznat podle ribozym autoaminoacylační systémy.[4] Neproteinogenní aminokyseliny by tedy byly vyloučeny případným evolučním úspěchem životních forem založených na nukleotidech. Byly nabídnuty další důvody, které vysvětlují, proč určité specifické neproteinogenní aminokyseliny nejsou obecně zabudovány do proteinů; například, ornitin a homoserin cyklizovat proti peptidové kostře a fragmentovat protein relativně krátkou poločasy rozpadu zatímco jiné jsou toxické, protože mohou být omylem zabudovány do proteinů, jako je analog argininu kanavanin.
Struktury
Následující text ilustruje struktury a zkratky 21 aminokyselin, které jsou přímo kódovány pro syntézu proteinů genetickým kódem eukaryot. Níže uvedené struktury jsou standardní chemické struktury, nikoli typické zwitterion formy, které existují ve vodných roztocích.
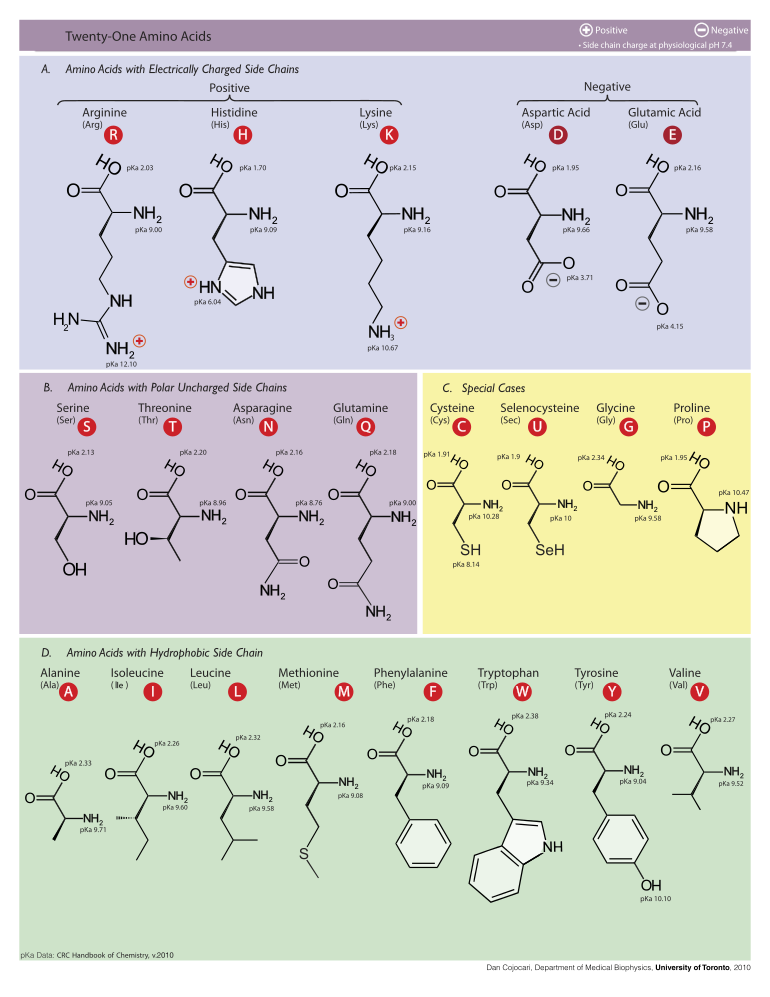

L-Alanin
(Ala / A)
L-Arginin
(Arg / R)
L-Asparagin
(Asn / N)
L-Aspartová kyselina
(Asp / D)
L-Cystein
(Cys / C)
L-Kyselina glutamová
(Glu / E)
L-Glutamin
(Gln / Q)
Glycin
(Gly / G)
L-Histidin
(His / H)
L-Isoleucine
(Ile / I)
L-Leucin
(Leu / L)
L-Lysin
(Lys / K)
L-Methionin
(Met / M)
L-Fenylalanin
(Phe / F)
L-Proline
(Pro / P)
L-Serine
(Ser / S)
L-Treonin
(Thr / T)
L- Tryptofan
(Trp / W)
L-Tyrosin
(Tyr / Y)
L-Valine
(Val / V)

















IUPAC /IUBMB nyní také doporučuje standardní zkratky pro následující dvě aminokyseliny:

L-Selenocystein
(Sec / U)
L-Pyrrolysin
(Pyl / O)

Chemické vlastnosti
Následuje tabulka se seznamem jednopísmenných symbolů, třípísmenných symbolů a chemických vlastností postranních řetězců standardních aminokyselin. Uvedené hmotnosti jsou založeny na vážených průměrech elementů izotopy na jejich přirozené množství. Formování a peptidová vazba vede k eliminaci molekuly voda. Proto je hmotnost proteinu rovna hmotnosti aminokyselin, z nichž je protein složen, minus 18,01524 Da na peptidovou vazbu.
Obecné chemické vlastnosti
| Aminokyselina | Krátký | Zkratka. | Prům. Hmotnost (Da ) | pI | pK1 (α-COOH) | pK2 (α-+NH3) |
|---|---|---|---|---|---|---|
| Alanin | A | Ala | 89.09404 | 6.01 | 2.35 | 9.87 |
| Cystein | C | Cys | 121.15404 | 5.05 | 1.92 | 10.70 |
| Kyselina asparagová | D | Asp | 133.10384 | 2.85 | 1.99 | 9.90 |
| Kyselina glutamová | E | Glu | 147.13074 | 3.15 | 2.10 | 9.47 |
| Fenylalanin | F | Phe | 165.19184 | 5.49 | 2.20 | 9.31 |
| Glycin | G | Gly | 75.06714 | 6.06 | 2.35 | 9.78 |
| Histidin | H | Jeho | 155.15634 | 7.60 | 1.80 | 9.33 |
| Isoleucin | Já | Ile | 131.17464 | 6.05 | 2.32 | 9.76 |
| Lysin | K. | Lys | 146.18934 | 9.60 | 2.16 | 9.06 |
| Leucin | L | Leu | 131.17464 | 6.01 | 2.33 | 9.74 |
| Methionin | M | Se setkal | 149.20784 | 5.74 | 2.13 | 9.28 |
| Asparagin | N | Asn | 132.11904 | 5.41 | 2.14 | 8.72 |
| Pyrrolysin | Ó | Pyl | 255.31 | ? | ? | ? |
| Prolin | P | Pro | 115.13194 | 6.30 | 1.95 | 10.64 |
| Glutamin | Q | Gln | 146.14594 | 5.65 | 2.17 | 9.13 |
| Arginin | R | Arg | 174.20274 | 10.76 | 1.82 | 8.99 |
| Serine | S | Ser | 105.09344 | 5.68 | 2.19 | 9.21 |
| Threonin | T | Thr | 119.12034 | 5.60 | 2.09 | 9.10 |
| Selenocystein | U | Sec | 168.053 | 5.47 | 1.91 | 10 |
| Valine | PROTI | Val | 117.14784 | 6.00 | 2.39 | 9.74 |
| Tryptofan | Ž | Trp | 204.22844 | 5.89 | 2.46 | 9.41 |
| Tyrosin | Y | Tyr | 181.19124 | 5.64 | 2.20 | 9.21 |
Vlastnosti postranního řetězce
| Aminokyselina | Krátký | Zkratka. | Boční řetěz | Hydro- fobický | pKa§ | Polární | pH | Malý | Drobný | Aromatický nebo Alifatický | van der Waals objem (A3) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alanin | A | Ala | -CH3 | - | - | Alifatický | 67 | ||||
| Cystein | C | Cys | -CH2SH | 8.55 | kyselé | - | 86 | ||||
| Kyselina asparagová | D | Asp | -CH2COOH | 3.67 | kyselé | - | 91 | ||||
| Kyselina glutamová | E | Glu | -CH2CH2COOH | 4.25 | kyselé | - | 109 | ||||
| Fenylalanin | F | Phe | -CH2C6H5 | - | - | Aromatický | 135 | ||||
| Glycin | G | Gly | -H | - | - | - | 48 | ||||
| Histidin | H | Jeho | -CH2-C3H3N2 | 6.54 | slabý základní | Aromatický | 118 | ||||
| Isoleucin | Já | Ile | -CH (CH3) CH2CH3 | - | - | Alifatický | 124 | ||||
| Lysin | K. | Lys | - (CH2)4NH2 | 10.40 | základní | - | 135 | ||||
| Leucin | L | Leu | -CH2CH (CH3)2 | - | - | Alifatický | 124 | ||||
| Methionin | M | Se setkal | -CH2CH2S CH3 | - | - | Alifatický | 124 | ||||
| Asparagin | N | Asn | -CH2CONH2 | - | - | - | 96 | ||||
| Pyrrolysin | Ó | Pyl | - (CH2)4NHCOC4H5N CH3 | N.D. | slabý základní | - | ? | ||||
| Prolin | P | Pro | -CH2CH2CH2- | - | - | - | 90 | ||||
| Glutamin | Q | Gln | -CH2CH2CONH2 | - | - | - | 114 | ||||
| Arginin | R | Arg | - (CH2)3NH-C (NH) NH2 | 12.3 | silně základní | - | 148 | ||||
| Serine | S | Ser | -CH2ACH | - | - | - | 73 | ||||
| Threonin | T | Thr | -CH (OH) CH3 | - | - | - | 93 | ||||
| Selenocystein | U | Sec | -CH2SeH | 5.43 | kyselé | - | ? | ||||
| Valine | PROTI | Val | -CH (CH3)2 | - | - | Alifatický | 105 | ||||
| Tryptofan | Ž | Trp | -CH2C8H6N | - | - | Aromatický | 163 | ||||
| Tyrosin | Y | Tyr | -CH2-C6H4ACH | 9.84 | slabě kyselý | Aromatický | 141 |
§: Hodnoty pro Asp, Cys, Glu, His, Lys a Tyr byly stanoveny pomocí aminokyselinového zbytku umístěného centrálně v alanin pentapeptidu.[5] Hodnota pro Arg je z Pace et al. (2009).[6] Hodnota pro Sec je od Byun & Kang (2011).[7]
N.D .: Hodnota pKa pyrrolysinu nebyla hlášena.
Poznámka: Hodnota pKa aminokyselinového zbytku v malém peptidu se obvykle mírně liší, pokud je uvnitř proteinu. Výpočty proteinu pKa se někdy v této situaci používají k výpočtu změny hodnoty pKa aminokyselinového zbytku.
Genová exprese a biochemie
| Aminokyselina | Krátký | Zkratka. | Codon (s) | Výskyt | Nezbytný‡ u lidí | |||
|---|---|---|---|---|---|---|---|---|
| v archaeanských proteinech (%)& | v proteinech bakterií (%)& | v proteinech eukaryotu (%)& | Výskyt v lidských bílkovinách (%)& | |||||
| Alanin | A | Ala | GCU, GCC, GCA, GCG | 8.2 | 10.06 | 7.63 | 7.01 | Ne |
| Cystein | C | Cys | UGU, UGC | 0.98 | 0.94 | 1.76 | 2.3 | Podmíněně |
| Kyselina asparagová | D | Asp | GAU, GAC | 6.21 | 5.59 | 5.4 | 4.73 | Ne |
| Kyselina glutamová | E | Glu | GAA, GAG | 7.69 | 6.15 | 6.42 | 7.09 | Podmíněně |
| Fenylalanin | F | Phe | UUU, UUC | 3.86 | 3.89 | 3.87 | 3.65 | Ano |
| Glycin | G | Gly | GGU, GGC, GGA, GGG | 7.58 | 7.76 | 6.33 | 6.58 | Podmíněně |
| Histidin | H | Jeho | CAU, CAC | 1.77 | 2.06 | 2.44 | 2.63 | Ano |
| Isoleucin | Já | Ile | AUU, AUC, AUA | 7.03 | 5.89 | 5.1 | 4.33 | Ano |
| Lysin | K. | Lys | AAA, AAG | 5.27 | 4.68 | 5.64 | 5.72 | Ano |
| Leucin | L | Leu | UUA, UUG, CUU, CUC, CUA, CUG | 9.31 | 10.09 | 9.29 | 9.97 | Ano |
| Methionin | M | Se setkal | SRPEN | 2.35 | 2.38 | 2.25 | 2.13 | Ano |
| Asparagin | N | Asn | AAU, AAC | 3.68 | 3.58 | 4.28 | 3.58 | Ne |
| Pyrrolysin | Ó | Pyl | UAG* | 0 | 0 | 0 | 0 | Ne |
| Prolin | P | Pro | CCU, CCC, CCA, CCG | 4.26 | 4.61 | 5.41 | 6.31 | Ne |
| Glutamin | Q | Gln | CAA, CAG | 2.38 | 3.58 | 4.21 | 4.77 | Ne |
| Arginin | R | Arg | CGU, CGC, CGA, CGG, AGA, AGG | 5.51 | 5.88 | 5.71 | 5.64 | Podmíněně |
| Serine | S | Ser | UCU, UCC, UCA, UCG, AGU, AGC | 6.17 | 5.85 | 8.34 | 8.33 | Ne |
| Threonin | T | Thr | ACU, ACC, ACA, ACG | 5.44 | 5.52 | 5.56 | 5.36 | Ano |
| Selenocystein | U | Sec | UGA** | 0 | 0 | 0 | >0 | Ne |
| Valine | PROTI | Val | GUU, GUC, GUA, GUG | 7.8 | 7.27 | 6.2 | 5.96 | Ano |
| Tryptofan | Ž | Trp | UGG | 1.03 | 1.27 | 1.24 | 1.22 | Ano |
| Tyrosin | Y | Tyr | UAU, UAC | 3.35 | 2.94 | 2.87 | 2.66 | Podmíněně |
| Stop kodonu† | - | Období | UAA, UAG, UGA†† | ? | ? | ? | N / A | N / A |
* UAG je obvykle jantarový stop kodon, ale v organismech obsahujících biologický aparát kódovaný klastrem genů pylTSBCD bude začleněna aminokyselina pyrrolysin.[8]
** UGA je obvykle opálový (nebo umber) stop kodon, ale kóduje selenocystein, pokud a Prvek SECIS je přítomen.
† The stop kodon není aminokyselina, ale je zahrnuta pro úplnost.
†† UAG a UGA ne vždy fungují jako stop kodony (viz výše).
‡ Esenciální aminokyselinu nelze u lidí syntetizovat, a proto musí být dodávána ve stravě. Podmíněně esenciální aminokyseliny nejsou obvykle ve stravě vyžadovány, ale musí být dodány exogenně konkrétním populacím, které jej nesyntetizují v dostatečném množství.
& Výskyt aminokyselin je založen na 135 Archeaách, 3775 bakteriích, 614 Eukaryota proteomech a na lidském proteomu (21 006 proteinů).[9]
Hmotnostní spektrometrie
v hmotnostní spektrometrie Z peptidů a proteinů je užitečná znalost hmot zbytků. Hmotnost peptidu nebo proteinu je součtem zbytkových hmot plus hmotnosti voda (Monoizotopová hmota = 18,01056 Da; průměrná hmotnost = 18,0153 Da). Hmotnosti zbytků se počítají z chemických vzorců uvedených v tabulce a atomových hmotností.[10] v hmotnostní spektrometrie ionty mohou také zahrnovat jeden nebo více protony (Monoizotopová hmota = 1,00728 Da; průměrná hmotnost = 1,0074 Da).
| Aminokyselina | Krátký | Zkratka. | Vzorec | Pondělí Hmotnost§ (Da) | Prům. Hmotnost (Da) |
|---|---|---|---|---|---|
| Alanin | A | Ala | C3H5NE | 71.03711 | 71.0779 |
| Cystein | C | Cys | C3H5NOS | 103.00919 | 103.1429 |
| Kyselina asparagová | D | Asp | C4H5NE3 | 115.02694 | 115.0874 |
| Kyselina glutamová | E | Glu | C5H7NE3 | 129.04259 | 129.1140 |
| Fenylalanin | F | Phe | C9H9NE | 147.06841 | 147.1739 |
| Glycin | G | Gly | C2H3NE | 57.02146 | 57.0513 |
| Histidin | H | Jeho | C6H7N3Ó | 137.05891 | 137.1393 |
| Isoleucin | Já | Ile | C6H11NE | 113.08406 | 113.1576 |
| Lysin | K. | Lys | C6H12N2Ó | 128.09496 | 128.1723 |
| Leucin | L | Leu | C6H11NE | 113.08406 | 113.1576 |
| Methionin | M | Se setkal | C5H9NOS | 131.04049 | 131.1961 |
| Asparagin | N | Asn | C4H6N2Ó2 | 114.04293 | 114.1026 |
| Pyrrolysin | Ó | Pyl | C12H19N3Ó2 | 237.14773 | 237.2982 |
| Prolin | P | Pro | C5H7NE | 97.05276 | 97.1152 |
| Glutamin | Q | Gln | C5H8N2Ó2 | 128.05858 | 128.1292 |
| Arginin | R | Arg | C6H12N4Ó | 156.10111 | 156.1857 |
| Serine | S | Ser | C3H5NE2 | 87.03203 | 87.0773 |
| Threonin | T | Thr | C4H7NE2 | 101.04768 | 101.1039 |
| Selenocystein | U | Sec | C3H5Nos | 150.95364 | 150.0489 |
| Valine | PROTI | Val | C5H9NE | 99.06841 | 99.1311 |
| Tryptofan | Ž | Trp | C11H10N2Ó | 186.07931 | 186.2099 |
| Tyrosin | Y | Tyr | C9H9NE2 | 163.06333 | 163.1733 |
Stechiometrie a metabolické náklady v buňce
Níže uvedená tabulka uvádí množství aminokyselin v E-coli buňky a metabolické náklady (ATP) pro syntézu aminokyselin. Záporná čísla znamenají, že metabolické procesy jsou energeticky příznivé a nestojí čistou ATP buňky.[11] Množství aminokyselin zahrnuje aminokyseliny ve volné formě a v polymerační formě (proteiny).
| Aminokyselina | Krátký | Zkratka. | Hojnost (počet molekul (× 108) za E-coli buňka) | Náklady ATP v syntéze | |
|---|---|---|---|---|---|
| Aerobní podmínky | Anaerobní podmínky | ||||
| Alanin | A | Ala | 2.9 | -1 | 1 |
| Cystein | C | Cys | 0.52 | 11 | 15 |
| Kyselina asparagová | D | Asp | 1.4 | 0 | 2 |
| Kyselina glutamová | E | Glu | 1.5 | -7 | -1 |
| Fenylalanin | F | Phe | 1.1 | -6 | 2 |
| Glycin | G | Gly | 3.5 | -2 | 2 |
| Histidin | H | Jeho | 0.54 | 1 | 7 |
| Isoleucin | Já | Ile | 1.7 | 7 | 11 |
| Lysin | K. | Lys | 2.0 | 5 | 9 |
| Leucin | L | Leu | 2.6 | -9 | 1 |
| Methionin | M | Se setkal | 0.88 | 21 | 23 |
| Asparagin | N | Asn | 1.4 | 3 | 5 |
| Pyrrolysin | Ó | Pyl | - | - | - |
| Prolin | P | Pro | 1.3 | -2 | 4 |
| Glutamin | Q | Gln | 1.5 | -6 | 0 |
| Arginin | R | Arg | 1.7 | 5 | 13 |
| Serine | S | Ser | 1.2 | -2 | 2 |
| Threonin | T | Thr | 1.5 | 6 | 8 |
| Selenocystein | U | Sec | - | - | - |
| Valine | PROTI | Val | 2.4 | -2 | 2 |
| Tryptofan | Ž | Trp | 0.33 | -7 | 7 |
| Tyrosin | Y | Tyr | 0.79 | -8 | 2 |
Poznámky
| Aminokyselina | Zkratka. | Poznámky | |
|---|---|---|---|
| Alanin | A | Ala | Je velmi bohatý a velmi univerzální, je tužší než glycin, ale dostatečně malý, aby představoval jen malé sterické limity pro konformaci proteinu. Chová se poměrně neutrálně a může být umístěn jak v hydrofilních oblastech na proteinu venku, tak v hydrofobních oblastech uvnitř. |
| Asparagin nebo kyselina asparagová | B | Asx | Zástupný symbol, když kterákoli aminokyselina může zaujímat pozici |
| Cystein | C | Cys | Atom síry se snadno váže těžký kov ionty. Za oxidačních podmínek se mohou dva cysteiny spojit dohromady v a disulfidová vazba za vzniku aminokyseliny cystin. Pokud jsou cystiny součástí proteinu, inzulín například terciární struktura je stabilizovaný, což zvyšuje odolnost proteinu denaturace; proto jsou disulfidové vazby běžné v proteinech, které musí fungovat v drsném prostředí včetně trávicích enzymů (např. pepsin a chymotrypsin ) a strukturní proteiny (např. keratin ). Disulfidy se také nacházejí v peptidech, které jsou příliš malé na to, aby si samy udržovaly stabilní tvar (např. inzulín ). |
| Kyselina asparagová | D | Asp | Asp se chová podobně jako kyselina glutamová a nese hydrofilní kyselou skupinu se silným negativním nábojem. Obvykle je umístěn na vnějším povrchu proteinu, což z něj činí rozpustný ve vodě. Váže se na pozitivně nabité molekuly a ionty a často se používá v enzymech k fixaci kovového iontu. Pokud jsou umístěny uvnitř proteinu, jsou aspartát a glutamát obvykle spárovány s argininem a lysinem. |
| Kyselina glutamová | E | Glu | Glu se chová podobně jako kyselina asparagová a má delší, mírně pružnější postranní řetězec. |
| Fenylalanin | F | Phe | Fenylalanin, tyrosin a tryptofan, které jsou pro člověka nezbytné, obsahují velké a tuhé látky aromatický skupina na postranním řetězci. Jedná se o největší aminokyseliny. Stejně jako isoleucin, leucin a valin jsou hydrofobní a mají tendenci se orientovat směrem dovnitř složené proteinové molekuly. Fenylalanin lze převést na tyrosin. |
| Glycin | G | Gly | Vzhledem ke dvěma atomům vodíku na uhlíku α glycin není opticky aktivní. Je to nejmenší aminokyselina, snadno se otáčí a dodává pružnost proteinovému řetězci. Je schopen zapadnout do nejužších prostor, např. Trojité šroubovice kolagen. Protože příliš velká flexibilita obvykle není požadována, je jako strukturální složka méně častá než alanin. |
| Histidin | H | Jeho | Jeho je pro lidi zásadní. I v mírně kyselých podmínkách protonace dusíku dochází ke změně vlastností histidinu a polypeptidu jako celku. Používá jej mnoho proteinů jako regulační mechanismus, který mění konformaci a chování polypeptidu v kyselých oblastech, jako je pozdní endosom nebo lysozom, vynucení změny konformace v enzymech. K tomu je však zapotřebí jen několik histidinů, takže je to poměrně vzácné. |
| Isoleucin | Já | Ile | Ile je pro lidi nezbytný. Isoleucin, leucin a valin mají velké alifatické hydrofobní postranní řetězce. Jejich molekuly jsou tuhé a jejich vzájemné hydrofobní interakce jsou důležité pro správné skládání proteinů, protože tyto řetězce mají tendenci být umístěny uvnitř molekuly proteinu. |
| Leucin nebo isoleucin | J | Xle | Zástupný symbol, když může kterákoli aminokyselina zaujímat pozici |
| Lysin | K. | Lys | Lys je pro člověka nezbytný a chová se podobně jako arginin. Obsahuje dlouhý, pružný boční řetěz s kladně nabitým koncem. Flexibilita řetězce činí lysin a arginin vhodným pro vazbu na molekuly s mnoha negativními náboji na jejich površích. Např., DNA vazebné proteiny mají aktivní oblasti bohaté na arginin a lysin. Díky silnému náboji jsou tyto dvě aminokyseliny náchylné k umístění na vnějších hydrofilních površích proteinů; když se nacházejí uvnitř, jsou obvykle spárovány s odpovídající záporně nabitou aminokyselinou, např. aspartát nebo glutamát. |
| Leucin | L | Leu | Leu je nezbytný pro člověka a chová se podobně jako isoleucin a valin. |
| Methionin | M | Se setkal | Met je pro člověka zásadní. Vždy první aminokyselina, která je začleněna do proteinu, je po translaci někdy odstraněna. Stejně jako cystein obsahuje síru, ale s a methyl skupina místo vodíku. Tuto methylovou skupinu lze aktivovat a používá se v mnoha reakcích, kdy se k jiné molekule přidává nový atom uhlíku. |
| Asparagin | N | Asn | Podobně jako kyselina asparagová obsahuje Asn amide skupina, kde má Asp a karboxyl. |
| Pyrrolysin | Ó | Pyl | Podobný lysin, ale má pyrrolin prsten připojen. |
| Prolin | P | Pro | Pro obsahuje neobvyklý kruh k aminové skupině na N-konci, který nutí sekvenci amidu CO-NH do pevné konformace. Může narušit struktury skládání bílkovin α šroubovice nebo β list, nutí požadovaný zlom v proteinovém řetězci. Obyčejný v kolagen, často prochází a posttranslační modifikace na hydroxyprolin. |
| Glutamin | Q | Gln | Podobně jako kyselina glutamová obsahuje Gln an amide skupina, kde má Glu a karboxyl. Používá se v bílkovinách a jako úložiště pro amoniak, je to nejhojnější aminokyselina v těle. |
| Arginin | R | Arg | Funkčně podobné lysinu. |
| Serine | S | Ser | Serin a threonin mají krátkou skupinu zakončenou hydroxylovou skupinou. Jeho vodík se snadno odstraňuje, takže serin a threonin často působí jako donory vodíku v enzymech. Oba jsou velmi hydrofilní, takže vnější oblasti rozpustných proteinů mají tendenci být s nimi bohaté. |
| Threonin | T | Thr | Thr je pro člověka zásadní a chová se podobně jako serin. |
| Selenocystein | U | Sec | The selen analog cysteinu, ve kterém selen nahrazuje síra atom. |
| Valine | PROTI | Val | Val je nezbytný pro člověka a chová se podobně jako isoleucin a leucin. |
| Tryptofan | Ž | Trp | Trp, nezbytný pro člověka, se chová podobně jako fenylalanin a tyrosin. Je předchůdcem serotonin a je přirozeně fluorescenční. |
| Neznámý | X | Xaa | Zástupný symbol, když je aminokyselina neznámá nebo nedůležitá. |
| Tyrosin | Y | Tyr | Tyr se chová podobně jako fenylalanin (předchůdce tyrosinu) a tryptofan a je předchůdcem melanin, epinefrin, a hormony štítné žlázy. Přirozeně fluorescenční, jeho fluorescence je obvykle uhasena přenosem energie na tryptofany. |
| Kyselina glutamová nebo glutamin | Z | Glx | Zástupný symbol, když kterákoli aminokyselina může zaujímat pozici |
Katabolismus

Aminokyseliny lze klasifikovat podle vlastností jejich hlavních produktů:[12]
- Glukogenní, s produkty, které mají schopnost se tvořit glukóza podle glukoneogeneze
- Ketogenní, s produkty, které nejsou schopné tvořit glukózu: Tyto produkty lze stále používat ketogeneze nebo syntéza lipidů.
- Aminokyseliny se katabolizovaly na glukogenní i ketogenní produkty
Viz také
Reference
- ^ Ambrogelly A, Palioura S, Söll D (leden 2007). "Přirozené rozšíření genetického kódu". Přírodní chemická biologie. 3 (1): 29–35. doi:10.1038 / nchembio847. PMID 17173027.
- ^ Lobanov AV, Turanov AA, Hatfield DL, Gladyshev VN (srpen 2010). „Duální funkce kodonů v genetickém kódu“. Kritické recenze v biochemii a molekulární biologii. 45 (4): 257–65. doi:10.3109/10409231003786094. PMC 3311535. PMID 20446809.
- ^ Young VR (srpen 1994). „Požadavky na aminokyseliny pro dospělé: důvod pro zásadní revizi současných doporučení“ (PDF). The Journal of Nutrition. 124 (8 doplňků): 1517S – 1523S. doi:10.1093 / jn / 124.suppl_8.1517S. PMID 8064412.
- ^ Erives A (srpen 2011). „Model proto-antikodonových RNA enzymů vyžadujících L-aminokyselinovou homochiralitu“. Journal of Molecular Evolution. 73 (1–2): 10–22. Bibcode:2011JMolE..73 ... 10E. doi:10.1007 / s00239-011-9453-4. PMC 3223571. PMID 21779963.
- ^ Thurlkill RL, Grimsley GR, Scholtz JM, Pace CN (květen 2006). „hodnoty pK ionizovatelných skupin proteinů“. Věda o bílkovinách. 15 (5): 1214–8. doi:10.1110 / ps.051840806. PMC 2242523. PMID 16597822.
- ^ Pace CN, Grimsley GR, Scholtz JM (květen 2009). „Protein ionizovatelné skupiny: hodnoty pK a jejich příspěvek ke stabilitě a rozpustnosti proteinu“. The Journal of Biological Chemistry. 284 (20): 13285–9. doi:10,1074 / jbc.R800080200. PMC 2679426. PMID 19164280.
- ^ Byun BJ, Kang YK (květen 2011). "Konformační preference a hodnota pK (a) zbytku selenocysteinu". Biopolymery. 95 (5): 345–53. doi:10,1002 / bip.21581. PMID 21213257.
- ^ Rother M, Krzycki JA (srpen 2010). "Selenocystein, pyrrolysin a jedinečný energetický metabolismus methanogenní archea". Archaea. 2010: 1–14. doi:10.1155/2010/453642. PMC 2933860. PMID 20847933.
- ^ Kozlowski LP (leden 2017). "Proteome-pI: proteome isoelectric point database". Výzkum nukleových kyselin. 45 (D1): D1112 – D1116. doi:10.1093 / nar / gkw978. PMC 5210655. PMID 27789699.
- ^ „Atomové váhy a izotopové směsi pro všechny prvky“. NIST. Citováno 2016-12-12.
- ^ Phillips R, Kondev J, Theriot J, Garcia HG, Orme N (2013). Fyzikální biologie buňky (Druhé vydání.). Věnec věnec. p. 178. ISBN 978-0-8153-4450-6.
- ^ Ferrier DR (2005). „Kapitola 20: Degradace a syntéza aminokyselin“. V Champe PC, Harvey RA, Ferrier DR (eds.). Lippincott's Illustrated Reviews: Biochemistry (Lippincott's Illustrated Reviews). Hagerstwon, MD: Lippincott Williams & Wilkins. ISBN 978-0-7817-2265-0.
Obecné odkazy
- Nelson, David L .; Cox, Michael M. (2000). Lehningerovy principy biochemie (3. vyd.). Worth Publishers. ISBN 978-1-57259-153-0.
- Kyte J, Doolittle RF (květen 1982). "Jednoduchá metoda pro zobrazení hydropatického charakteru proteinu". Journal of Molecular Biology. 157 (1): 105–32. CiteSeerX 10.1.1.458.454. doi:10.1016/0022-2836(82)90515-0. PMID 7108955.
- Meierhenrich, Uwe J. (2008). Aminokyseliny a asymetrie života (1. vyd.). Springer. ISBN 978-3-540-76885-2.
- Biochemistry, Harpers (2015). Harpers Illustrated Biochemistry (30. vydání). Lange. ISBN 978-0-07-182534-4.
externí odkazy
| Obecná témata |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Podle vlastností |
| ||||||||||
| |||||||||||